

Abstract
The insular cortex is one of the brain regions that show consistent abnormalities in both structural and functional neuroimaging studies of schizophrenia. In healthy individuals, the insula has been implicated in a myriad of physiologic functions. The anterior cingulate cortex (ACC) and insula together constitute the salience network, an intrinsic large-scale network showing strong functional connectivity. Considering the insula as a functional unit along with the ACC provides an integrated understanding of the role of the insula in information processing. In this review, we bring together evidence from imaging studies to understand the role of the salience network in schizophrenia and propose a model of insular dysfunction in psychosis.
Introduction
Some of the most consistent and well-replicated findings in the neurobiology of psychotic disorders pertain to changes in the brain structure. In this regard, the insular cortex has emerged in recent times as a key brain region with consistent structural alterations in both schizophrenia and bipolar disorder.1 Despite recent attempts to delineate the role of the insula in various neuropsychiatric disorders, including schizophrenia,2–4 an integrative model of insular dysfunction in relation to psychosis is lacking.
The insula is a reciprocally connected brain region.5,6 A fuller understanding of insular dysfunction in the pathogenesis of psychosis can be obtained by bringing together evidence from structural and functional imaging to highlight the potential role of the disrupted interaction of this structure with other brain regions in individuals with psychosis.
In particular, the often-observed coactivation of the insula and the anterior cingulate cortex (ACC) across a variety of cognitive tasks suggests a functional network involving these 2 regions.7 Both functional and structural connectivity have been demonstrated between the insula and the ACC,7,8 with likely extension into the inferior frontal region.9,10 This intrinsic network has been variously described as a cognitive task control network and salience network.9,10 Menon and Uddin11 have recently proposed that the primary role of this network is to enable switching between the default mode and task-related states of brain connectivity. Disrupted coordination among brain circuits, in particular between the anticorrelated networks that underlie task-related activity and default modes, has been postulated as a core pathophysiologic feature of schizophrenia.12
According to an emerging Bayesian framework,13 clinical symptoms of psychosis can be explained on the basis of a mismatch between prior expectations and incoming sensory information leading to a prediction error. The insula has been repeatedly shown as an important cortical region involved in prediction error coding, although other areas, such as the ventral striatum, have also been implicated.14,15
In this review, as the primary evidence for insular dysfunction comes from structural imaging studies, we first consider in detail the nature of structural deficits across the salience network in individuals with schizophrenia. We then review the functional attributes of the insula in the context of the salience network and prediction error model along with the relation between insular deficits and clinical symptoms of psychosis. In the final section, we propose an integrated model of insular dysfunction in psychosis with suggestions for future directions necessary to confirm this model.
Structural deficits of the insula and ACC in individuals with schizophrenia
Voxel-based morphometry studies of schizophrenia
Voxel-based morphometry (VBM) is one of the most common techniques employed to study the neuroanatomy of schizophrenia. Despite the limitations inherent to the method,16 most VBM studies have implicated the insula and ACC as key regions showing consistent grey matter deficits in patients with schizophrenia.17–20 Using anatomic likelihood estimation methods on coordinates reported from 31 VBM studies (1195 patients and 1262 controls), Glahn and colleagues19 found the left and right insula to be the regions with maximum grey matter reduction among a distributed network of other regions, including the anterior cingulate and parahippocampal gyrus. A focused analysis by Fornito and colleagues21 of VBM studies of the ACC confirms the significant reduction in insula grey matter.
Grey matter density as measured by VBM is a proxy rather than absolute measure of grey matter structure owing to the VBM technique being based on the probabilistic classification of voxels subjected to affine registration. Hence, definitive conclusions regarding prominent grey matter reduction across the salience network cannot be made using VBM alone. Despite the differences in manual tracing methods and the issue of reliability while defining insular and ACC regions, region-of-interest (ROI) studies measure the absolute morphometric properties and circumvent the problems related to image registration.
Region-of-interest studies of schizophrenia
Most ROI studies exploring insular volume confirm a reduction in grey matter volume of the insula in individuals with schizophrenia.22–29 In addition, ROI studies address the issues of laterality, regional localization and the nature of the morphometric changes (thickness, area and volume) in the insula. With respect to the latter, Crespo-Facorro and colleagues22 showed significant reductions in both the volume and surface area of insular grey matter in patients with a first episode of psychosis. In a larger sample of 118 patients and 83 controls, examination of insular thickness revealed significant cortical thinning.30 Most ROI studies have reported a bilateral volume reduction,23,29 though some studies show a predominant left insular22,24 or right insular involvement.25,30
Differentiating between the anterior and posterior subdivisions of the insula, Makris and colleagues26 showed that the anterior insula had greater volume reduction than the posterior insula in individuals with schizophrenia. A moderate effect size of 0.6 was noted for left anterior insular volume reduction. Such regional differences were replicated in some,31 but not all studies.23,29 These differences are likely related to the inconsistencies in manual tracing for ROI studies in this anatomically complex region. In summary, existing evidence suggests that bilateral insular volume is reduced in individuals with schizophrenia, with both anterior and posterior subdivisions contributing to the reduction.
Goldstein and colleagues,27 who studied grey matter volume reductions using a parcellation method, found that the largest reductions in the schizophrenia group occurred in the bilateral insula and ACC. Baiano and colleagues32 systematically reviewed structural imaging studies reporting on ACC volume in individuals with schizophrenia and reported significant reduction in absolute ACC volume. They found that whereas functional subdivisions are reported within the ACC, most morphometric studies have considered the ACC as a whole (see Fornito and colleagues21 for further discussion). Taken together, ROI studies of the insula along with the numerous ROI studies reporting reductions in ACC volume suggest a significant structural deficit across the salience network in individuals with schizophrenia.
Time course of structural changes
Despite accumulating evidence regarding widespread cortical grey matter deficits in individuals with schizophrenia, the exact time course of the onset and progress of these deficits are still open to speculation. Evidence from studies of human fetal brains suggests that the process of development of cortical folding (gyrification) is first notable in the insular region and proceeds in an orderly fashion.33 The peri-insular sulci and the central (insular and cerebral) sulci are the first macroscopic structures visible on the lateral surface of the human fetal cerebral hemisphere; they can first be seen between gestational weeks 13–17. This is a crucial period for neuronal migration that is considered to be important in the etiology of schizophrenia.34,35 An abnormality in cortical development during this period is likely to affect the gyrification and subsequent formation of the insula. Unpublished data (2011) from our laboratory suggests abnormal gyrification of the in-sula is indeed a feature of schizophrenia. Further evidence for aberrant development comes from a study of deformation of the shape of the insula, which demonstrates significant reduction in the rostral end of the inferior limit of the limen in-sulae (corresponding to the anteroinferior limit of the insula) in individuals with schizophrenia.36 There is some evidence to suggest that the developmental trajectories of the insula and ACC thickness may be similar.37
To our knowledge, no systematic study has been carried out to date to estimate the onset of insular volume reduction in individuals with psychosis. Various cross-sectional comparisons add strength to the assumption that both insular and ACC deficits predate the onset of psychosis.21,38 Borgwardt and colleagues39 reported a significant deficit in left insular volume in both the high-risk group (at-risk mental state) and in the first-episode schizophrenia group. Within the at-risk group, those in whom psychosis developed after 2 years showed right insular volume reduction at initial assessment compared with those in whom psychosis did not develop. Chan and colleagues38 reviewed VBM studies of high-risk groups in addition to first-episode and chronic schizophrenia groups. High-risk individuals showed bilateral anterior cingulate and right insular deficits. Deficits in these regions were persistently noted in the first-episode and chronic schizophrenia groups too. Takahashi and colleagues40 found that in high-risk individuals who showed transition to psychosis, significant bilateral insular volume reduction was observable even before the transition compared with those in whom psychosis did not develop. In this study, greater progressive grey matter reduction of the bilateral insular cortex (−5.0% per yr) was seen in high-risk participants who showed transition to psychosis compared with controls (−0.4% per yr) or high-risk participants without transition in 4 years (−0.6% per yr).
In addition to the progressive reduction during the high-risk state, there is some evidence that insular grey matter further reduces after the first episode. In a sample different from the one reported above, Takahashi and colleagues31 compared 23 first-episode patients with 26 controls and 11 patients with chronic schizophrenia. Two years after the initial scan, the first-episode group showed the most severe loss of total insular volume (> 4%), followed by the chronic schizophrenia group (about 1.5%) and controls (about 0.3%).
Most VBM studies that compare genetically high-risk but healthy individuals with healthy controls find reduced ACC grey matter,41–43 though not all studies agree.44 Goghari and colleagues45 demonstrated bilateral reduction in ACC thickness in genetically high-risk individuals. Longitudinal studies of genetically high-risk individuals are lacking; however, studies that follow ultrahigh-risk groups based on clinical state and trait criteria demonstrate that both ACC46 and insular39 grey matter reduction predicts transition to psychosis.
Taken together, it seems that the structure of the salience network is affected at an early stage of development predating the onset of psychosis. Progressive grey matter changes in the salience network are associated with transition to psychosis; this volume loss continues at least in the first few years after the first episode.
Effect of antipsychotics
An important issue when evaluating structural deficits in individuals with schizophrenia is teasing out the confounding effect of medications. The bulk of available evidence suggests that antipsychotics have regionally specific effects on brain structure, with the basal ganglia being the most susceptible.47,48 Conflicting evidence has been presented with respect to the insula in this regard. Several authors report a lack of correlation between prescribed antipsychotic dose and insular volume.23,25,28,29,31 Pressler and colleagues49 failed to replicate the finding of reduced insular volume in a sample of 30 patients with chronic schizophrenia, but they showed that with increasing typical neuroleptic exposure the insular volume increased in their sample. In contrast, using a cross-sectional VBM analysis of a sample exposed to short-term antipsychotics, Dazzan and colleagues50 suggested that patients taking typical antipsychotics may have reduced insular grey matter when compared with drug-free patients. However, it is possible that this association between typical antipsychotics and grey matter volume is confounded by symptom burden, as there was a trend for higher positive symptom scores in the treated group (Cohen d = 0.43; 5.1 points difference in positive symptom score). In a meta-analysis, Leung and colleagues17 reviewed VBM studies of antipsychotic-naïve first-episode patients and compared the results with the pooled estimates of studies of first-episode patients treated with anti-psychotics. In both groups, bilateral insular and ACC deficits were prominent. Whereas caudate and temporal deficits were less extensive in the samples treated with antipsychotics, insular deficits (along with parahippocampal and frontal deficits) were more pronounced in the treated samples. A similar effect was not observed for the ACC. Magnetic resonance spectroscopy studies suggest that a long-term treatment with atypical antipsychotics may have a favourable effect on neuronal viability in the ACC.51,52 In line with these findings, Tomelleri and colleagues53 found a positive correlation between cumulative typical antipsychotic exposure and left anterior cingulate volume using VBM. In summary, evidence indicates an association between antipsychotic exposure and reduced insula volume, whereas there is some evidence that treatment protects the ACC. However, the evidence that the volume deficits in both the insula and ACC precede treatment in antipsychotic-naïve first-episode patients and in untreated high-risk populations suggests that the illness process itself contributes to the structural deficits in the salience network.
Specificity of the salience network deficits to schizophrenia
Most of the evidence we reviewed pertains to schizophrenia. But studies of other psychotic conditions suggest that insular deficits may not be specific to schizophrenia. Skaf and colleagues54 found reduced blood flow in the right insula (along with the inferior frontal gyrus) in patients with psychotic depression compared with nonpsychotic depression, suggesting psychosis may be related to insular dysfunction in depressed patients. Bora and colleagues,55 using a signed differential mapping procedure, showed that the anterior insular region (extending to the inferior frontal cortex) and left ACC are the regions most affected in VBM studies of bipolar disorder. A head-to-head comparison of VBM studies of bipolar disorder and schizophrenia revealed that bilateral insular and anterior cingulate lesions are common among individuals with bipolar disorder or schizophrenia.1 The 2 ROI studies that claim specificity of bilateral insular volume reduction for schizophrenia29,56 include patients who were in relatively early stages of illness. Some explanation for these conflicting results comes from Bora and colleagues55 who, using meta-regression modelling, showed that there were no frontoinsular grey matter reductions in patients with first-episode bipolar disorder; nonetheless, within this sample the left frontoinsular cortex was smaller in patients with longer durations of illness. This suggests that though not seen in relatively early phases of illness, insular grey matter deficits are seen in patients with established bipolar disorder. It is unclear whether this is related to emergence of psychotic symptoms in these patients.
Autism-spectrum disorders are often included in the extended spectrum of psychotic disorders on the basis of shared neurobiology. Interestingly, a recent meta-analysis showed that insular and cingulate deficits are shared among individuals with schizophrenia or autism-spectrum disorders, though some hemispheric differences in terms of more left lateralized deficits may occur in individuals with schizophrenia.57 This meta-analysis included mostly studies with neuroleptic-naïve participants, though the psychotic symptom burden among autistic individuals was not reported. Further, a prominent role for the anterior insula in the social processing defects of autism-spectrum disorders has been proposed on the basis of various functional magnetic resonance imaging (fMRI) studies.58 Taken together, it is plausible that insular deficits are related to expression of major psychotic disorders, such as schizophrenia and bipolar disorder, though the specificity of structural deficits to the presence of the psychotic dimension needs to be examined in heterogeneous samples.
Salience network white matter structure in individuals with schizophrenia
To our knowledge, no studies have evaluated the salience network in individuals with schizophrenia using diffusion tractography, though the connections between the ACC and insula have been traced in healthy volunteers.8,59 A meta-analysis60 reviewing voxel-based diffusion imaging studies of schizophrenia identified significant reductions in fractional anisotropy in 2 regions proximal to the insula and cingulate. Moriya and colleagues61 studied a first-episode sample and reported prominent involvement of both the insula and ACC, indicating a disruption of axonal morphology at an early stage of schizophrenia.
Neuropathology of the salience network regions in individuals with schizophrenia
The 2 nodes of the salience network are unique among other brain regions in harbouring a special class of large bipolar spindle cells called von Economo neurons (VEN). These neurons are seen only in higher primates62 and are thought to be involved in social processing in the wake of their rapid relaying ability.63,64 Patients with early-onset schizophrenia with longer durations of illness exhibited a reduced VEN density in the anterior cingulate,65 though no studies have explored this pathology in the insula. Preliminary evidence also suggests that VENs preferentially express the protein encoded by DISC1 (disrupted in schizophrenia 1).66 The only study to date exploring insular neuropathology in individuals with schizophrenia showed decreased neuronal volume in layer 2 of the insula.67 Furthermore, proteomic analysis of the insula in individuals with schizophrenia has revealed abnormal expression of multiple proteins that participate in synaptic function and neuronal morphogenesis.68
Functional attributes of the salience network
Stimulus evaluation and proximal salience
Anatomic likelihood estimation meta-analysis of fMRI studies with insula activation69 suggests that the anterior insula is concerned with a higher-level integrative process. This integration of different qualities of our coherent experience of the world sets the context for thoughts and actions. Craig70 assembles a large body of evidence from functional imaging studies demonstrating that the insula is active during the processing of many internal bodily stimuli (e.g., thirst, sexual arousal, heartbeat, visceral distension) and external stimuli (e.g., temperature, taste, pain). Similarly, Augustine5 has concluded from a review of structural connections in primates and humans that the insula is a site of multisensory integration. Furthermore, Augustine reports that the anterior insula has a strong reciprocal connectivity with prefrontal regions.5 As the prefrontal cortex contains representations of social and event models,71 this reciprocal connectivity suggests that the insula has an access to goal and plan representations. In addition, imaging studies of stimulus-independent thoughts reveal engagement of core midline structures of the default mode network (DMN) together with the anterior insula and ACC.72,73 Thus, the salience network receives information about internal and external sensations, representations of goals and plans, and stimulus-independent thoughts.
A substantial body of evidence demonstrates insular activation during a wide range of tasks that involve evaluating probability, uncertainty of an outcome,74,75 reward and risk prediction (see Bossaerts75). In particular, insular activity is often seen with prediction error coding. Prediction error refers to a discrepancy between an expectation and its occurrence. This discrepancy can lead to the updating of expectations about the external and internal milieu and, if necessary, initiates or modifies action. Murray and colleagues76 showed that reward prediction error in healthy participants was associated with activation of the ventral striatum along with the in-sula and cingulate cortex, all of which were not seen in patients with psychosis. In a different fMRI study of prediction errors, the strongest correlation between risk prediction error and blood oxygen level–dependent response was found in the insula and inferior frontal gyrus.77
Several studies suggest that the specific role of the insula in prediction error paradigms is likely to be one of updating the previously existing prediction framework.78–81 Activation of the insula during risk evaluation predicted subsequent decision-making, indicating that the insula plays a role in not only evaluating but also updating the probabilities of an outcome.78–80 Insular activity during anticipated loss predicted loss-avoidance learning several months later.81 The ACC also plays a critical role in updating the prediction models and has been shown to be involved in both social and reward-related associative learning,82 though it is unlikely to be the sole region for cognitive control.83,84 Notably, individuals who have lesions on the insula show a failure to update their prediction frameworks, despite having preserved the ability to judge the probability of events.85
In addition to the properties of the stimulus, execution of a response also depends on the context provided by the current homeostatic state of the participant. Craig’s model of “sentient self” places the insula at a central role for evaluating ongoing feeling states.70,86 Craig proposes that awareness of any object requires, first, a mental representation of oneself as a feeling (sentient) entity; second, a mental representation of that object; and third, a mental representation of the salient interrelationship between oneself and that object in the immediate moment (“now”). These 3 aspects are integrated in the anterior insula, creating a state of interoceptive awareness.70 This is consistent with evidence that the anterior insula plays a crucial role in appraisal of self and the attribution of personal relevance.87,88
These observations suggest that a primary role of the salience network is the integration of sensations, internally generated thoughts and information about goals and plans in order to update expectations about the internal and external milieu and, if necessary, to allow action to be initiated or modified. To facilitate the description of the function of the salience network, we will introduce the concept of proximal salience (Fig. 1). An event, such as an externally generated sensation, a bodily sensation or a stimulus-independent thought, attains proximal salience when it generates a momentary state of neural activity within the salience network that results in updating of expectations and, if warranted by the context, initiates or modifies action.

Proximal salience refers to a momentary state of neural activity within the salience network generated by the evaluation of external or internal stimuli. The salience network interacts with the interoceptive pathway and prefrontal system to generate the proximal salience that enables a switching between resting mode to task-processing (executive) mode or vice versa. ACC = anterior cingulate cortex; CEN = central executive network; DMN = default mode network.
Proximal salience prepares one for appropriate behavioural response. To be behaviourally effective, the brain region that integrates proximal salience must be tightly coupled to task-related brain regions. Indeed, this was clearly demonstrated by Sridharan and colleagues,89 who showed that anterior insula activation precedes activation of task-related brain networks. Using Granger causality analysis, they proposed a role for the anterior insula and anterior cingulate in switching brain states from the default mode to the task-related activity mode, although the application of Granger causality to fMRI data must be interpreted cautiously.
Several studies have shown that learning by means of developing new models of prediction involves changes in the resting state connectivity in both the DMN and task-related networks.90–93 With its unique role as a switch between these 2 networks, the salience network is crucially positioned to not only enable the behavioural response, but also to consolidate or update prediction models subsequent to stimulus evaluation. In complex social situations, an update of prediction models could involve attitudinal changes. Van Veen and colleagues94 observed that the insular activation during cognitive dissonance is inversely correlated with the degree of attitude change (rationalization) that follows, indicating that an optimal amount of insula activation is required for acquisition and/or alteration of beliefs.
Functional MRI studies of schizophrenia
Abnormalities in the functional activation of the insula and ACC have been reported across a variety of tasks ranging from working memory paradigms to complex social processing95,96 (see Wylie and Tregellas2). Studies investigating functional integration (during resting or task-related states) across multiple brain regions report a prominent reduction in connectivity involving both the insula97,98 and ACC99,100 with other brain regions.
Attenuated coactivation of the regions constituting the salience network is noted in individuals with schizophrenia compared with healthy controls during task execution101 and error processing.102 Such an effect has also been observed in patients when attending to somatosensory stimuli.103 In that study,103 the reduced engagement of the salience network when attending to the external stimuli was also associated with poor deactivation of the DMN. More direct evidence for an impaired interaction between the salience network and the DMN comes from a functional connectivity analysis of independent components in the same sample.104 Notably, failure to deactivate the DMN has been shown previously in various resting state studies of patients with schizophrenia and their relatives.105–107
In summary, disrupted functional connectivity is noted across the insula and ACC in individuals with schizophrenia. A disruption in the salience network is associated with impaired deactivation of the DMN in individuals with schizophrenia.
Correlation of insular deficits with clinical symptoms
A meta-analysis of fMRI studies of active auditory hallucinations reveals prominent involvement of the insula, bilateral Broca area and auditory cortex.108 Evidence from the analysis of whole brain structure in hallucinating patients consistently implicates insular deficits.109–115 The presence of concurrent structural and functional deficits implies an essential role for the insula in producing hallucinations.
Passivity symptoms are shown to be related to insular dysfunction. When undertaking theory of mind tasks, patients with schizophrenia and passivity symptoms show reduced activation of the right insula and anterior cingulate.116 Positron emission tomography and fMRI studies have also found abnormalities in the activation of the insula in addition to brain regions involved in action monitoring in patients with passivity symptoms.117,118 Consistent with a study by Crespo-Facorro and colleagues,22 a recent study from our group has shown significant correlation between the volume of the salience network and severity of delusions and hallucinations.119
Insular volume is related to other aspects of psychopathology as well.31 An ROI study by Makris and colleagues26 showed that the left anterior insular volume correlated with bizarre behaviour in individuals with schizophrenia. But some studies that did not distinguish subregions failed to find correlations with symptoms scores.24 Early positron emission tomography studies suggested that reduced insular blood flow was associated with both disorganization syndrome and reality distortion in individuals with schizophrenia.120 Using arterial spin labelling, Horn and colleagues121 demonstrated that, along with language areas, the anterior insula showed significant correlation of resting cerebral blood flow (rCBF) and degree of formal thought disorder. Using 2 independent cohorts of drug-free patients, Lahti and colleagues122 showed that rCBF in the Broca area, anterior insula and ACC correlated significantly with disorganization scores.
A recent VBM study of the 3 major psychopathologic dimensions of schizophrenia reported the insula to be the most prominent brain region to show deficits across the 3 clusters of positive symptoms, disorganization and negative symptoms.123 It is worth noting that most studies reporting clinical correlation of insular deficits included patients taking antipsychotic medications. Antipsychotics not only reduce the severity of psychotic symptoms (especially reality distortion and disorganization), but also affect the brain structure as discussed previously. This can introduce a variability leading to inconsistencies in the relation between clinical symptoms and brain structure. Nonetheless, a somewhat coherent pattern of relation between insular deficits and symptoms of schizophrenia has been noted. Whereas reality distortion emerges as a consistent clinical association with insular deficits, the relation with symptoms of disorganization is equally prominent and requires further study. In addition, there is some evidence for an association with negative symptoms.
Insular dysfunction model of psychosis
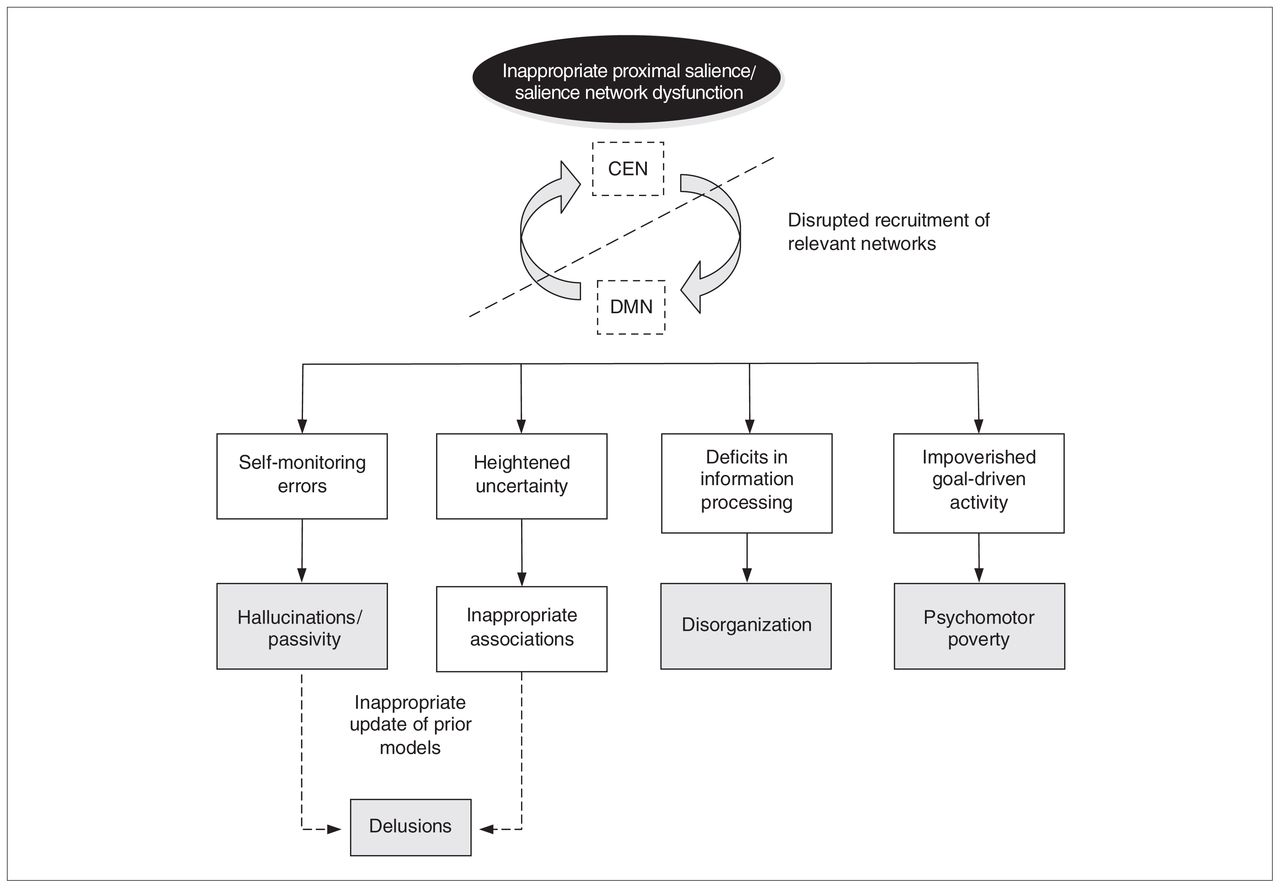
Several investigations suggest that hallucinations are related to self-generated inner speech and that passivity phenomena are related to self-generated actions.124,125 We propose that normally these self-generated internal processes do not generate proximal salience. However, insular activation during hallucinations suggests that the salience network is generating an inappropriate proximal salience during an otherwise normal activity.108 In particular, such an aberrant activation of the in-sula is noted alongside a prominent absence of cingulate activation, suggesting disruption in normal salience network activity126 and perhaps disruption to error-monitoring circuitry. The allocation of proximal salience to an event might lead to recruitment of the attentional networks required for processing the stimulus, as suggested by Seeley and colleagues.10 The faulty allocation of proximal salience to an internally generated mental event would be expected to promote recruitment of the DMN and impede the normal suppression of DMN activity during tasks requiring attention to the external world. Various groups have reported the attenuation of DMN suppression during task performance.105–107 Thus the internally generated mental activity might be further enhanced, creating a vicious cycle of inappropriate proximal salience. In a similar vein, and consistent with a recently proposed Bayesian model of positive symptoms,13 the inappropriate allocation of proximal salience to internally generated actions could explain the passivity symptoms.
Delusions are classified as primary or secondary. Primary delusions arise when significance is attached to an incidental perception without logical justification, whereas secondary delusions are secondary to other abnormal mental states, such as hallucinations or mood disturbances. A subjective state of uncertainty (delusional mood) or anxiety has been reported to precede the formation of primary delusions.127 Conrad described this state as a phase of apophenia that precedes full-blown delusions.128 It is plausible that the insular abnormality seen in susceptible individuals during prodromal states39,40,129 plays a role in this state of uncertainty. Insofar as the salience network plays a key role in the engagement of relevant distributed circuits required for processing information (and switching off less relevant circuits), salience network dysfunction and the associated failure of generation and response to proximal salience would be expected to enhance the state of uncertainty.
In the context of heightened uncertainty regarding the predicted outcome of events, seemingly irrelevant incidental stimuli (external and internal) might be allocated inappropriate proximal salience. Models of learning, such as the Rescorla–Wagner model130 (originally developed to account for reinforcement learning, but also employed to account for incidental associative learning that is not directly related to task performance or reward131), invoke the generation of a “teaching signal” when there is a discrepancy between the predicted outcome and the actual outcome of an event, resulting in the learning of a new association between the predictor and the outcome. In a similar manner, inappropriate proximal salience might lead to the incidental stimulus acquiring unwarranted causal significance, resulting in the formation of a primary delusion. Continuing deficits in the recruitment of appropriate attentional networks hamper the correct evaluation of the formed belief (updated model), whereas the associated reduction in uncertainty might serve to maintain the newly formed belief. Once an inappropriate model is formed, the process of further consolidation of this model contributes to secondary delusional elaborations, as suggested by Corlett and colleagues.132
Impaired regulation of switching between the default mode and attention to task-relevant stimuli that is hypothesized to arise from insular dysfunction11 is likely to be related to attentional impairment and distractibility. Disturbances of attention, including distractibility, are features of the disorganization syndrome in individuals with schizophrenia.133–136 Liddle and colleagues120 reported that in patients with persistent symptoms, disorganization was associated with aberrant activity in the insula, anterior cingulate and adjacent medial prefrontal cortex.
It is plausible that a defect in integration of goals and plans into the state of “interoceptive awareness” described by Craig,70 might result in the diminution of initiation of activity characteristic of the psychomotor poverty syndrome. This might arise owing to a fault within the salience network itself, though the possibility cannot be ruled out that a more diffuse impairment of frontal lobe function might contribute to the impairment of the integration of goals and plans into the state of interoceptive awareness.137 Thus, a dysfunctional salience network and associated disruption of proximal salience might account for many of the clinical features of psychosis (Fig. 2).

Insular dysfunction model of psychosis. A dysfunction of the salience network can lead to a cascade of events that result in clinical symptoms of psychosis. Inappropriate proximal salience during self-generated actions result in hallucinations and passivity experiences. Proximal salience inappropriately attached to external stimulus in the context of heightened uncertainty in unusual beliefs and delusions. A failure to engage the task-relevant network when required is associated with disorganization. Failure to generate goal-driven activity is related to psychomotor poverty. CEN = central executive network; DMN = default mode network.
It should be noted that whereas Menon and Uddin11 propose that the salience network is engaged in mediating interactions between brain networks involved in externally oriented attention and internally oriented or self-related cognition, we propose that the network has a more general switching role. We propose that the primary role of the salience network is initiating the recruitment of brain regions relevant for processing currently salient stimuli while decreasing activity in networks engaged in processing previously salient stimuli. This concept does not exclude the possibility that there are brain regions that can be engaged in attending to both internal and external stimuli. Under at least some circumstances, such as during tasks that require integrating information from the external world with information held in memory, we would anticipate simultaneous activity in nodes of the DMN and in other networks, such as those involved in perceptual or executive processing. We consider that the terms “default-mode processing” and “task-positive processing” can be misleading insofar as the DMN can be actively engaged during task performance, whereas networks other than the DMN are active during rest.89
Dopamine and the salience network
Dopamine (DA) plays a major role as the neurochemical mediator of prediction error signals,138 and DA dysfunction has been considered to have a central role in the emergence of the state of aberrant motivational salience seen in individuals with psychosis.139 The prediction error model of hallucinations and delusions13,132 is consistent with the DA hypothesis. To our knowledge, the relation between dopaminergic dysfunction and the insula has not been systematically studied in individuals with schizophrenia so far. Nonetheless, various sources of evidence suggest that a dopaminergic abnormality is likely to be associated with salience network dysfunction in individuals with schizophrenia.
Dopamine has emerged as the primary neurochemical mediator in relation to various traits and behaviours mediated by the insula (e.g., novelty-seeking,140 craving,141,142 nociception143). Moreover, various studies have demonstrated the importance of dopaminergic modulation on the ACC during executive tasks,144 suggesting that DA plays an important role in the function of the salience network. Both the insula and ACC are regions with relatively high extrastriatal DA transporters (DAT).145,146 The synaptic availability of DA may be directly related to efficient insular function. A polymorphism that is shown to be associated with higher levels of DAT, which mediate DA reuptake from the synaptic cleft into the presynaptic terminal, has been shown to be associated with greater activation of the insula and caudate along with deactivation of the cingulate during a verbal fluency task.147
A high correlation between the binding of the D2/D3 ligand [18F]-fallypride and grey matter density as measured by VBM has been observed in the anterior cingulate, insula and midbrain regions,148 raising the possibility that reduced grey matter across the salience network in individuals with schizophrenia may be directly associated with dopaminergic defects. In individuals with schizophrenia, abnormal dopaminergic transmission has been observed in the ACC.149,150 This defect may be amenable to pharmacologic manipulation, as shown by Dolan and colleagues.151
In summary, the insular dysfunction model of psychosis based on the salience network is consistent with the dopaminergic hypothesis of psychosis. Thus, the salience network provides a candidate cortical framework that is consistent with and builds on the existing dopaminergic hypothesis of schizophrenia.
Nevertheless, it is important to differentiate the concept of motivational salience traditionally associated with dopaminergic dysfunction139 from our concept of proximal salience. Proximal salience refers to a momentary state generated by evaluation of external or internal stimuli in the context of interoceptive awareness. This state precedes subsequent choice of action and/or optimization of predictive models relevant for the stimuli (learning). Kapur’s concept of motivational salience139 refers to a process that takes place once a stimulus is evaluated: it represents the assignment of a motivational value to the external object or internal representation. In other words, motivational salience refers to the process of attaching a tag of significance to the stimuli. Whereas motivational salience explains stimulus-reinforcement associations, proximal salience refers to a more fundamental step in information processing: the stimulus–response association. Whereas the concept of motivational salience places emphasis on the role of the corpus striatum,152 our concept of proximal salience places greater emphasis on the role of the insula and anterior cingulate. Nonetheless, the evidence that these brain regions interact in the attribution of salience and the notion of prediction errors and dopaminergic mediation discussed in the previous sections may serve as a common link between these 2 concepts.
Conclusion
Despite the consistency of this parsimonious model in which insula dysfunctions might lead to the major symptoms of psychosis, some key questions remain unanswered. For example, with respect to structural deficits in patients, it is unclear whether there is a critical insular grey matter volume below which psychotic symptoms emerge. Furthermore, the role of the insular dysfunction in relapses and remissions needs investigation. Longitudinal studies to clarify the time course of the structural and functional abnormalities in addition to studies employing novel imaging methods for studying cerebral connectivity might contribute to answering these questions. In addition, studies of unmedicated patients and treatment trials are required to elucidate the effect of antipsychotics on the function of the salience network.
The postulation of a specific role for the salience network in stimulus evaluation, updating of expectations and preparation for response generation suggests several specific hypotheses that might be tested using neuroimaging procedures. In particular, further investigation of the role of DA in salience network function in healthy individuals and in those with schizophrenia might not only clarify the relation between the concepts of proximal salience and motivational salience but also advance our understanding of the mechanism of psychosis.
Footnotes
Competing interests: L. Palaniyappan declares having received the Eli Lilly Young Investigator Travel Award to participate in the 9th Bipolar Conference, Pittsburgh, Pa. P.F. Liddle declares having received lecture honoraria from Bristol Meyers Squibb.
Contributors: L. Palaniyappan and P.F. Liddle both contributed to study design, wrote and reviewed the article and approved its publication.
- Received December 8, 2010.
- Revision received February 21, 2011.
- Accepted April 15, 2011.
References
In this issue

{kind=link}
{kind=link}
Article tools






Related Articles
Cited By...
- Networks extracted from nonlinear fMRI connectivity exhibit unique spatial variation and enhanced sensitivity to differences between individuals with schizophrenia and controls
- Functional alterations of two salience-related systems jointly and independently contribute to psychosis
- Spatiotemporal mapping of the neural markers of prediction error processing across multisensory and unisensory modalities
- Right anterior insula effective connectivity impairs intrinsic BOLD fluctuations in dorsal attention network in adolescents and young adults with borderline personality symptoms
- Grey matter morphometric biomarkers for classifying early schizophrenia and PD psychosis: a multicentre study
- Effective connectivity of LSD-induced ego dissolution
- Activity flow underlying abnormalities in brain activations and cognition in schizophrenia
- A meta-analytic approach to mapping co-occurrent grey matter volume increases and decreases in psychiatric disorders
- Insula Functional Connectivity in Schizophrenia
- Hubs of long-distance co-alteration in brain pathology
- Spatial patterning of tissue volume loss in schizophrenia reflects brain network architecture
- fMRI salience signals in antipsychotic naïve first episode psychosis
- Transient patterns of functional dysconnectivity in youth with psychosis spectrum symptoms
- Maternal Immune Activation During the Third Trimester Is Associated with Neonatal Functional Connectivity of the Salience Network and Fetal to Toddler Behavior
- Changes in extra-striatal functional connectivity in patients with schizophrenia in a psychotic episode
- Structural and functional brain changes in delusional disorder
- Challenges and opportunities in psychiatry today
- Challenges and opportunities in (neuro)psychiatry
- Functional connectivity and grey matter volume of the striatum in schizophrenia
- Biological vulnerability to depression: linked structural and functional brain network findings
- Functional Connectivity in Healthy Subjects Is Nonlinearly Modulated by the COMT and DRD2 Polymorphisms in a Functional System-Dependent Manner
- Tardive dyskinesia in schizophrenia