Abstract
Tuberculosis (TB) is a disease caused by Mycobacterium tuberculosis whose interaction with the host may lead to a cell-mediated protective immune response. The presence of interferon<FONT FACE=Symbol>-g</FONT> (IFN-gamma) is related to this response. With the purpose of understanding the immunological mechanisms involved in this protection, the lymphoproliferative response, IFN<FONT FACE=Symbol>-g</FONT> and other cytokines like interleukin (IL-5, IL-10), and tumor necrosis factor alpha (TNF<FONT FACE=Symbol>-a</FONT>) were evaluated before and after the use of anti-TB drugs on 30 patients with active TB disease, 24 healthy household contacts of active TB patients, with positive purified protein derivative (PPD) skin tests (induration > 10 mm), and 34 asymptomatic individuals with negative PPD skin test results (induration < 5 mm). The positive lymphoproliferative response among peripheral blood mononuclear cells of patients showed high levels of IFN<FONT FACE=Symbol>-g</FONT>, TNF<FONT FACE=Symbol>-a</FONT>, and IL-10. No significant levels of IL-5 were detected. After treatment with rifampicina, isoniazida, and pirazinamida, only the levels of IFN<FONT FACE=Symbol>-g</FONT> increased significantly (p < 0.01). These results highlight the need for further evaluation of IFN<FONT FACE=Symbol>-g</FONT> production as a healing prognostic of patients treated.
tuberculosis; interleukin-5; interleukin-10; tumor necrosis factor alpha; interferon<FONT FACE=Symbol>-g</FONT>; peripheral blood mononuclear cells
IMMUNOLOGY
Pulmonary tuberculosis: evaluation of interferon-g levels as an immunological healing marker based on the response to the Bacillus Calmette-Guerin
EP MouraI; VPCP ToledoI; MHP OliveiraIII; S Spíndola-de-MirandaII; HM AndradeIV; TMPD GuimarãesI, 1 1 Corresponding author. Fax: +55-31-3339.7644. E-mail: taniadg@farmacia.ufmg.br
IDepartamento de Análises Clínicas e Toxicológicas, Faculdade de Farmácia
IIDepartamento de Clínica Médica, Faculdade de Medicina, UFMG, Av. Olegário Maciel 2360, 30180-112 Belo Horizonte, MG, Brasil
IIICentro de Saúde Oswaldo Cruz, Prefeitura de Belo Horizonte, Belo Horizonte, MG, Brasil
IVDepartamento de Parasitologia e Microbiologia, UFPI, Teresina, PI, Brasil
ABSTRACT
Tuberculosis (TB) is a disease caused by Mycobacterium tuberculosis whose interaction with the host may lead to a cell-mediated protective immune response. The presence of interferon-g (IFN-g) is related to this response. With the purpose of understanding the immunological mechanisms involved in this protection, the lymphoproliferative response, IFN-g and other cytokines like interleukin (IL-5, IL-10), and tumor necrosis factor alpha (TNF-a) were evaluated before and after the use of anti-TB drugs on 30 patients with active TB disease, 24 healthy household contacts of active TB patients, with positive purified protein derivative (PPD) skin tests (induration > 10 mm), and 34 asymptomatic individuals with negative PPD skin test results (induration < 5 mm). The positive lymphoproliferative response among peripheral blood mononuclear cells of patients showed high levels of IFN-g, TNF-a, and IL-10. No significant levels of IL-5 were detected. After treatment with rifampicina, isoniazida, and pirazinamida, only the levels of IFN-g increased significantly (p < 0.01). These results highlight the need for further evaluation of IFN-g production as a healing prognostic of patients treated.
Key words: tuberculosis - interleukin-5 - interleukin-10 - tumor necrosis factor alpha - interferon-g - peripheral blood mononuclear cells
Tuberculosis (TB), a chronic infective-contagious disease, is caused by Mycobacterium tuberculosis and remains an important public health problem whose mechanisms related to a protective immunity in humans are not clear. The resurgence of TB has stimulated studies for the development of vaccines, new diagnostic methods and less toxic and more effective drugs for treatment (Laal et al. 1997). The cellular immunity plays an important role in TB healing (Ladel et al. 1997, Torres et al. 1998, Turner et al. 2000, Chackerian et al. 2001). Resistance to mycobacterial infections is conferred by immunological mechanisms mediated by T CD4+ lymphocytes, involving cytokines that increase the microbicide activity of macrophages (Dlugovitzky et al. 2000, Oberholzer et al. 2000, Flynn & Chan 2001). Studies in murine and human models allow differentiating two subpopulations of T CD4+ lymphocytes termed Th1 and Th2, that mediate the protection or the aggravation of the disease (Ladel et al. 1997, Kori et al. 2000). This existing dichotomy between the protective or non-protective immune responses is likely to be correlated with cytokine patterns produced by different subpopulations of lymphocytes during initial surviving stages of the pathogens inside macrophages. Interferon-g (IFN-g) acts as a powerful macrophage activator, increasing the molecule expression of the main class II histocompatibility complex and the potentialization of the cell response, including the production of cytokines, nitric oxide, and the increase of the cytolitic activity, with a main role in the Th1 type (Flesh et al. 1995). Studies carried out by Cooper et al. (1993) and Flynn et al. (1993) demonstrated that mice without the IFN-g gene were not able to fight off the infection caused by M. tuberculosis. In humans, individuals who presented genetic mutations in the receptors for IFN-g were observed to have had a high susceptibility to acquire infections caused by atypical mycobacteria (Jouanguy et al. 1996), suggesting an important role of IFN-g in the protective response against TB.
Besides the importance of better understanding the immunological mechanisms that may contribute to the healing, it becomes essential to determine markers of healing lesions, once this is currently carried out based on the clinical, radiological, and negative bacterioscopy.
In this work, we proposed the in vitro evaluation of IFN-g levels produced by peripheral blood mononuclear cells (PBMC) after Bacillus Calmette-Guerin (BCG) stimulation of patients with active pulmonary TB before and after the chemotherapeutic treatment and of those with non TB disease, reactive to purified protein derivative (PPD), aiming to relate these levels to healing prognostics of active TB patients, and within this context, we also proposed the evaluation of some immunological parameters, such as the cellular immune response, using lymphoproliferative reaction and the cytokine quantification.
MATERIALS AND METHODS
Studied group - A total of 58 people volunteered to take part in the study which was developed from June, 2000 to September, 2001. Written consent was obtained from the volunteers or from parents/guardians of those under 18 years of age. Group 1 corresponded to 30 non-treated patients, from Oswaldo Cruz Health Center in Belo Horizonte, MG, Brazil and from Santa Luzia City Laboratory in Santa Luzia, MG, Brazil. Both males and females aged between 16 and 48, negative to human immunodeficiency virus (HIV), with active pulmonary TB that fulfilled the following criteria were included: suggestive radiography images and positive sputum respiratory specimens, using Ziehl-Neelsen method. All the patients had active pulmonary disease, showing a characteristic pattern: upper lobe infiltration with cavitation. They had never received anti-TB drugs before. On the other hand, pregnant women, individuals with debilitating diseases, those that had been vaccinated with BCG recently, and those who refused to sign the written consent, were excluded from the study. Twenty milliliters of heparinized peripheral blood were collected from TB patients before the use of anti-TB drugs. Group 2, comprised TB patients treated during 6 months and who showed favorable clinical response and became smear negative using three bacilloscopies and with negative sputum culture. Group 3 included 24 healthy household contacts of active TB patients, with positive PPD skin tests (induration > 10 mm). As a control group (Group 4), 34 volunteers from Hospital das Clínicas in Belo Horizonte were asymptomatic and with negative PPD skin test results (induration < 5 mm) and normal chest X ray.
Ethics - These procedures were in accordance with the ethical standards of the Faculdade de Medicina, UFMG committee on human experimentation, protocol register ETIC 009/98, approved in 03/11/1998 and with the Helsinki Declaration of 1975, as revised in 1983.
PPD skin test - Skin reactions of more than 10 mm in volunteers after an intradermal test with 5 U of PPD (Laboratório de Extratos Alergênicos Ltda, Rio de Janeiro, RJ) were considered to be positive, and reactions of less than 5 mm were considered to be negative. Skin test response was measured with the ballpoint technique 48-72 h after PPD injection, reported as millimeters of induration and performed always by the same nurse.
Antigens - BCG: Fundação Ataulpho de Paiva, Lot 9910178; purified protein derivative PPD: Laboratório de Extratos Alergênicos Ltda, Rio de Janeiro, RJ, gently granted by Secretaria Municipal de Saúde of Belo Horizonte. BCG was used as antigen for all the tests carried out in this work.
Lymphoproliferative response - The lymphoproliferative assay was carried out with PBMC collected from patients before and after the chemotherapeutic treatment in accordance with the methodology described by Torres et al. (1998). Briefly, PBMC were separated in histopaque gradient (Sigma Chemical Co, US) in the ratio 1:3 (histopaque-blood) in polystyrene, conic bottom, sterile tubes (Falcon, Corning, US). After centrifugation at 400 g during 45 min at 18ºC, the mononuclear cells ring which was formed in the histopaque-plasma interface was withdrawn. The resuspended cells were washed three times with RPMI-1640 medium, following centrifugation at 200 g, for 10 min at 4ºC. The cell concentration was adjusted to 1.5 x 106 cells/ml in RPMI-1640 medium supplemented with 5% of human serum (AB, Rh negative), 2 Mercaptoethanol, HEPES and 10 UI of penicillin-streptomycin/ml. Two hundred microliters of the suspension of mononuclear cells were added to each well of polystyrene (96 well plates) (Costar, US). Cultures were carried out in triplicate, sensitized with 10 µl of BCG (1 mg/ml), 10 µl of PPD (174 µg/ml) and with PHA (phytohemagglutinin, Sigma Chemical Co., US), in a concentration of 0.5 µg/well incubated at 37ºC with 95% humidity containing 5% of CO2. On the 5th day, 25 µl of RPMI-1640 medium containing thymidine (specific activity of 5.3 Ci/mM, Sigma) was added to each well. After 16-18 h, cells were isolated with the help of an automatic cell harvester (Titertek Cell Harvester, Flow Laboratories, US). The radioactivity was determined in counts per minute (cpm), using beta scintillation spectrometer (model 1209, Rackbeta, LKB). The intensity of the proliferative response of the peripheral blood lymphocytes was determined by calculating the stimulation index (SI) obtained by dividing the average cpm of the three wells stimulated with antigen or mitogen by the average cpm of three non-stimulated wells. As representative values of positive lymphoproliferative response, it was considered only those which SI were < 3 (Kori et al. 2000).
Evaluation IFN-g, interleukin-5 (IL-5), IL-10, and tumor necrosis factor alpha (TNF-a) levels - Supernatants were collected 72 h after antigenic stimulus, according to a standardization carried out in preliminary studies. Concentrations of IFN-g, IL-5, IL-10, and TNF-a were determined by the immunoenzymatic method of ELISA (enzyme-linked immunosorbent assay), making use of commercial kits (Quantikine Human Immunoassay, R&D, US). The cut-off values for each of the cytokines were taken as the arithmetic mean of the absorbances of the 20 negative control samples (obtained from healthy individuals), plus two standard deviation: IFN-g = 82 pg/ml; IL-5 = 8 pg/ml; IL-10 = 29 pg/ml, and TNF-a = 42 pg/ml.
Statistical analysis - The analyzed variables were compared using non-parametric methods of Kruskall-Wallis and Mann-Whitney. A level of significance of 95% was used for all the tests. All laboratorial tests in this study were conducted in a blind way, without the previous knowledge of which group belonged to each clinical sample.
RESULTS
Lymphoproliferative response - Eleven out of 13 patients evaluated presented SI > 3.0. The arithmetic averages ± standard deviations (SD) of SI obtained before and after treatment were 17.7 ± 26.3 and 11.8 ± 15.2, respectively. Groups 3 and 4 presented average ± SD of 1.6 ± 0.9 and 1.2 ± 0.5, respectively. There was no statistically significant difference between the lymphoproliferative response before and after the chemotherapeutic treatment (p > 0.05), thus not serving as a good immunological parameter that could be correlated to the patients' healing lesions. The response to PHA was assessed and shown to be higher than 3.0 in all groups.
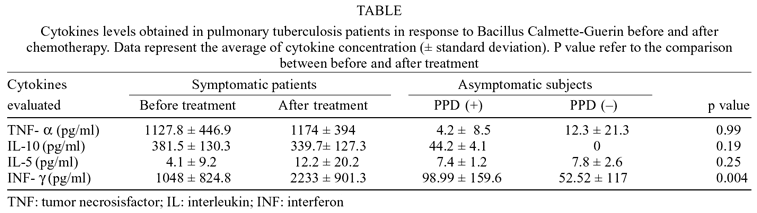
Dosage of IFN-g and other cytokines - The production of IFN-g was already high before the treatment and increased significantly (p < 0.01) after it (Figure). The levels of TNF-a and IL-10 were equally high before and after the treatment. Average levels of IL-5 obtained before the treatment were low and increased after the treatment, however, there was no significant difference between them, not distinguishing immunologically between the stage of active disease and the convalescence. Individuals reactive to PPD and the control group (non-reactive) did not present consistent levels of IFN-g, TNF-a, IL-10, and IL-5 (Table).
Interferon-g concentration in culture supernatant of mononuclear cells obtained from the peripheral blood of patients with active pulmonary tuberculosis, individuals reactive to purified protein derivative (PPD +) and control group (negative PPD), and stimulated with Bacillus Calmette-Guerin for 72 h. The horizontal bar represents the arithmetic average in each group. BT: before treatment; AT: after treatment.
DISCUSSION
In this study, the immune response to PBMC of patients with pulmonary TB facing BCG and PPD antigens was evaluated with the purpose of characterizing the immunological profile of these patients before and after the chemotherapic treatment. A better cellular response was observed when BCG was used as antigen, therefore this was the antigen chosen for all the tests carried out in this work. The difference regarding the reactivity obtained with the stimulation of the cells by these antigens could be related to the stimulation of different populations of T cells. Thus, it is known that PPD stimulates T CD4+ lymphocytes while BCG stimulates both CD4+ and CD8+ cells, in vitro (Turner & Dockrell 1996).
According to a current dogma, a vigorous Th1 response is mandatory for effective protection against Mycobacterium. This response is mediated by IFN-g secreting CD4+ T cells, which are presented with processed Mycobacterium antigens by dendritic cells and macrophages, which also secrete IL-12, thus inducing a Th1 response (Orme 1997).
In this study, the average level of IFN-g obtained in the supernatants of PBMC culture of the patients with pulmonary TB was high before the treatment and increased significantly after it. Several authors have shown that the pleural fluid of TB patients contain a high number of IFN-g producer cells (Barnes et al. 1993, Robinson et al. 1994, Raju et al. 2001), and they suggest that these cells may migrate to the lung and pleural tissue during the active disease, and therefore they may be reduced, temporarily, in the peripheral blood. This fact might explain the lower levels of IFN-g found before the use of anti-TB drugs in this study. However, in order to confirm this hypothesis, more invasive techniques may be used to collect pulmonary cells in infected individuals, if necessary. The increase in the productive capacity of IFN-g after the anti-TB treatment was also demonstrated in other studies carried out in patients with TB (Torres et al. 1998, Turner et al. 2000). Nevertheless, to date, it is still unclear if the increase in the production of IFN-g after treatment is related to cell stimulation by antigens released after the death of mycobacteria caused by chemotherapy, by reverting the antigenic status or by the reduction of the transit of T lymphocytes antigen specific to the lungs. Some authors have shown that there is a relationship between the production of some cytokines by PBMC and the severity of the disease (Wilsher et al. 1999). Dlugovitzky et al. (2000) observed that patients with the disease in a moderate stage presented significantly higher levels of IFN-g compared to the levels presented by patients with the disease in an advanced stage. The increase in the levels of IFN-g obtained after the treatment, in the present study, could be related to the use of anti-TB treatment, currently recommended by the Brazilian Health Ministry ( Ministério da Saúde 1995) and that voids the Mycobacterium after healing, which suggests an important role of the IFN-g in the immunological response against M. tuberculosis. In the present study after healing only one of the 20 patients evaluated did not increase the level of IFN-g. All other patients presented values of IFN-g above the pre-established cut-off value (82 pg/ml). Figure shows that the IFN-g levels are significantly reduced (p < 0.05) among individuals (PPD +) when compared to the group of patients with active disease. Apparently, our results are in contrast to other publications (Sanchez et al. 1994, Inokuchi et al. 2003) considering the IFN-g production in the PPD + group (household contacts). It could reflect the divergent patterns of immune reactivity against different mycobacterial antigens. Most of the studies use PPD as antigen to determine the profile of immune responsiveness that distinguishes patients with TB from healthy tuberculin-positive controls. In our study, we have employed BCG antigens in culture systems, because it seems to be more effective in eliciting a specific pattern of immune responsiveness to tuberculin-positive healthy individuals. In spite of this, IFN-g may be useful as an important immunological marker in TB healing.
TNF-a is involved both in the disease's physiopathology and in the protective response against M. tuberculosis and other mycobacterial infections (Flynn et al. 1995). Its role is complex and it is accepted that it acts in the synergism with IFN-g inducing the formation of intermediate reactive to nitrogen and oxygen, mediating the anti-TB activity of macrophages (Mohan et al. 2001, Warwick-Davies et al. 2001). But it has also been mentioned that TNF-a may be involved in the destruction of the pulmonary tissue (Flynn et al. 2001). In the present study, PBMCs of patients with active pulmonary TB were observed to produce TNF-a in high levels, however, no statistically significant difference was observed in the levels of this cytokine after the treatment (p = 0.842), reinforcing its role both in the disease physiopathology and in the protective immunity.
IL-10 cytokine has been considered as an anti-inflammatory cytokine, acting in the inactivation of macrophages through inhibiting the production of IL-12 and consequently reducing the production of IFN-g by T lymphocytes (Flynn et al. 2001). The IL-10 activity minimizes the tissular impairment occurred in the disease's site through inhibiting the production of proinflammatory cytokines (Bogdan et al. 1991, D'Andrea et al. 1993). In this study, PBMCs of patients with active pulmonary TB were observed to produce high levels of IL-10. The kinetics of the production of IL-10 did not seem to be influenced by the treatment. Constant levels of IL-10 and TNF-a obtained, suggested that the patient in order to compensate the tissular impairment caused by TNF-a might produce IL-10 cytokine. Olobo et al. (2001) showed that pleural fluid from patients contained elevated plasma levels of TNF-a, IL-10, and TGF-b, but the level of IL-10 was the highest followed by TGF-b and TNF-a. Probably, the effects induced by IFN-g and TNF-a represent bacteriostasis rather than bacterial killing. It could raise the possibility that the role of IL-10 would be to inhibit the bacterial killing by macrophages, but not to inhibit macrophage-mediated bacteriostasis. In the present study, macrophage and anti-mycobacteria activity could be principally expressed as a bacteriostatic action where the effect of IL-10 on killing might not be observed. Thus, the co-existence of IFN-g, IL-10, and TNF-a could be possible.
IL-5 cytokine is produced by Th2 lymphocytes and as IL-4, it acts in the differentiation of B lymphocytes in plasmocytes, stimulating the production of immunoglobulins. In this study, although the average concentration of IL-5 was low in all the groups analyzed, it was observed that there was a slight increase in the average level of this cytokine after the treatment, though it was not statistically significant. Other authors, in similar studies, also obtained low levels of IL-5 and IL-4 in patients with tuberculosis (Lin et al. 1996, Torres et al. 1998, Wilsher et al. 1999).
Some authors have reported a reduction in the lymphoproliferative response of PBMC of tuberculous patients facing PPD antigenic stimuli (Sanchez et al. 1994, Torres et al. 1998). In this study, no reduction was observed in this lymphoproliferative response once most patients presented positive stimulation indexes (SI > 3), showing that BCG antigens were able to induce significant levels of cellular proliferation (p < 0.05), before and after treatment. These antigens have also stimulated different clones of lymphocytes, independently of the T cell subset, because we have detected in PBMC culture supernatant, high levels of cytokines such as IFN-g and IL-10 belonging to Th1 and Th2 subsets.
Other studies have also shown a heterogeneous pattern of both Th1 and Th2 cytokine production by human T lymphocytes upon encounter with mycobacterial antigens (Haanen et al. 1991, Barnes et al. 1993, Orme et al. 1993). Further studies, investigating which kind of cells these clones are, must be done in order to clarify these results.
As mentioned above, this study has not demonstrated a clear dichotomy between the Th1 (protective immunity) and Th2 response (susceptibility to the disease), suggesting that both Th1 and Th2, are important for the immune response of the host against M. tuberculosis. Within this context, it becomes interesting to evaluate the relationship of IFN-g production as a healing prognostic for patients treated with the association of rifampicin, isoniazid and pyrazinamide, according to the recommendations of the Brazilian Health Ministry (Ministério da Saúde 1995).
ACKNOWLEDGEMENTS
To Dr Celso de Carvalho Diana from Prefeitura Municipal de Santa Luzia and Dr Helena Raquel from Centro de Saúde Oswaldo Cruz for access to the patients.
Received 10 September 2003
Accepted 27 April 2004
Financial support: Fapemig
- Barnes PF, Lu S, Abrams JS, Wang E, Yamamura M, Modlin RL 1993. Cytokine production at the site of disease in human tuberculosis. Infect Immun 61: 3482-3489.
- Bogdan C, Vodovotz Y, Nathan C 1991. Macrophage deactivation by interleukin 10. J Exp Med 174: 1549-1555.
- Chackerian AA, Perera TV, Behar SM 2001. Gamma interferon-producing CD4+ T lymphocytes in the lung correlate with resistance to infection with Mycobacterium tuberculosis. Infect Immun 69: 2666-2674.
- Cooper AM, Dalton DK, Stewart TA, Griffin JP, Russel DG, Orme IM 1993. Disseminated tuberculosis in interferon-g gene-disrupted mice. J Exp Med 178: 2243-2247.
- D'Andrea A, Aste-ameazaga M, Valiante NM, Ma X, Kubin M, Trinchieri G 1993. Interleukin 10 (IL-10) inhibits human lymphocyte interferon-g production by suppressing natural killer cell stimulatory factor/Il-12 synthesis in accessory cells. J Exp Med 178: 1041-1048.
- Dlugovitzy D, Bay ML, Rateni L, Fiorenza G, Vietti L, Farroni A, Botasso OA 2000. Influence of disease severity on nitrite and cytokine production by peripheral blood mononuclear cells (PBMC) from patients with pulmonary tuberculosis (TB). Clin Exp Immunol 122: 343-349.
- Flesch IEA, Hess JH, Huang S, Aguet M, Rothe J, Bluethmann H, Kaufmanns SH 1995. Early interleukin 12 production by macrophages in response to mycobacterial infection depends on interferon-g and tumor necrosis factor a. J Exp Med 181: 1615-1621.
- Flynn JL, Chan J 2001. Immunology of tuberculosis. Annu Rev Immunol 19: 93-129.
- Flynn JL, Chan J, Triebold KJ, Dalton DK, Stewart TA, Bloom BR 1993. An essential role for interferon-g in resistance to Mycobacterium tuberculosis infection. J Exp Med 178: 2249-2254.
- Flynn JL, Goldestein MM, Chan J, Triebold KJ, Pfelfer K, Lowenstein R, Schreiber R, Mak TW, Bloom BR 1995. Tumor necrosis factor-alpha is required in the protective immune response against. Mycobacterium tuberculosis in mice. Immunity 2: 561-572.
- Haanen JBAG, Waal Malefiji R, Res PCM, Kraakmann EM, Ottenhoff THM, Vries RRP, Spits H 1991. Selection of a human T helper type 1 like T cell subset by Mycobacteria. J Exp Med 174: 583-592.
- Inokuchi N, Sugahara, K, Soda H, Usui T, Hirakata Y, Fukushima K, Yamada Y, Kohno S, Kamihira S 2003. Relationship between whole-blood interferon-gamma production and extent of radiographic disease in patients with pulmonary tuberculosis. Diag Microbiol Infect Dis 46: 109-114.
- Jouanguy E, Altaire F, Lamhamedi S, Revy P, Emile JL, Newport MJ, Levin M, Blanche S, Seboun E, Fischer A, Casanova JL 1996. Interferon-gamma-receptor deficiency in an infant with fatal bacille Calmette-Guerin infection. N Engl J Med 335: 1956-1961.
- Kori M, Barak E, Altman Y, Eliraz A, Handzel ZT 2000. Specific in vitro proliferative immune responses and lymphokine production in Ethiopian children with and without tuberculosis. Infection 28: 42-45.
- Laal S, Samanich KM, Sonnenberg MG, Zolla-Pazner S, Phadtare JM, Belisle JT 1997. Human humoral responses to antigens of Mycobacterium tuberculosis: immunodominance of high-molecular-mass antigens. Clin Diag Lab Immunol 4: 49-56.
- Ladel CH, Szalay G, Riedel D, Kaufmann SHE 1997. Interleukin-12-secretion by Mycobacterium tuberculosis - Infected macrophages. Infect Immun 65: 1936-1938.
- Lin Y, Zhang M, Hofman FM, Gong J, Barnes PF 1996. Absence of a prominent Th2 cytokine response in human tuberculosis. Infect Immun 64: 1351-1356.
- Ministério da Saúde 1995. Normas e Manuais Técnicos 13, 44 pp.
- Mohan VP, Scanga CA, Yu K, Scott HM, Tanaka KE, Tsang E, Tsal MC, Flynn JL, Chan J 2001. Effects of tumor necrosis factor alpha on host immune response in chronic persistent tuberculosis: possible role for limiting pathology. Infect Immun 69: 1847-1855.
- Oberholzer A, Oberholzer C, Moldawer LL 2000. Cytokine signaling-regulation of the immune response in normal and critically ill states. Crit Care Med 28: 3-12.
- Olobo JO, Geletu M, Demissie A, Eguale T, Hiwot K, Aderayes G 2001. Circulating TNF-a, TGF-b, and IL-10 in tuberculosis patients and health contacts. Scand J Immunol 53: 85-91.
- Orme IM 1997. The host response to Mycobacterium tuberculosis. In SHE Kaufmann, Host Response to Intracellular Pathogens, Chapmann & Hall, New York, p. 459-471.
- Orme IM, Roberts AD, Griffin IS, Abrams IS 1993. Cytokine secretion by CD4 T lymphocytes acquired in response to Mycobacterium tuberculosis infection. J Immunol 151: 518-525.
- Raju B, Tung CF, Cheng D, Yousefzadeh N, Condos R, Rom WN, Doris BT 2001. In situ activation of helper T cells in the lung. Infect Immun 69: 4790-4798.
- Robinson D, Ying S, Taylor A, Wangoo D, Mitchell B, Kay B, Hamid Q, Shaw R 1994. Evidence for a Th1-like bronchoalveolar T-cell subset and predominance of interferon-gamma gene activation in pulmonary tuberculosis. Am J Resp Crit Care Med 149: 989-993.
- Sanchez FO, Rodrigues JL, Agudelo G, Garcai LF 1994. Immune responsiveness and lymphokine production in patients with tuberculosis and health controls. Infect Immun 62: 5673-5678.
- Torres M, Herrera T, Villareal H, Rich EA, Sada E 1998. Cytocine profiles for peripheral blood lymphocytes from patients with active pulmonary tuberculosis and health household contacts in response to the 30-kilodalton antigen of Mycobacterium tuberculosis. Infect Immun 66: 176-180.
- Turner J, Dockrell HM 1996. Stimulation of human peripheral blood mononuclear cells with live Mycobacterium bovis BCG activates cytolytic CD8+ T cells in vitro. Immunology 87: 339-342.
- Turner J, Corrah T, Sabbally S, Whittle H, Dockrell HM 2000. A longitudinal study of in vitro IFN-g production and cytotoxic T cell responses of tuberculosis patients in the Gambia. Tuber Lung Dis 80: 161-169.
- Warwick-Davies J, Watson AJ, Griffin GE, Krishna S, Shattock RJ 2001. Enhancement of Mycobacterium tuberculosis induced tumor necrosis factor alpha production from primary human monocytes by an activated T-cell membrane-mediated mechanism. Infect Immun 69: 6580-6587.
- Wilsher ML, Hagan C, Prestidge R, Wells AU, Murison G 1999. Human in vitro immune response to Mycobacterium tuberculosis. Tuber Lung Dis 79: 371-377.
Publication Dates
-
Publication in this collection
19 July 2004 -
Date of issue
May 2004
History
-
Received
10 Sept 2003 -
Accepted
27 Apr 2004