Resumo
Este artigo versa sobre aspectos socioculturais do cultivo de pequi (Caryocar brasiliense) entre os Kuikuro do alto Xingu, para os quais esta espécie possui alto valor simbólico e alimentar. O cultivo de pequi é uma prática compartilhada entre os nove povos indígenas que compõem uma sociedade pluriétnica e multilinguística na região dos formadores do Rio Xingu, da qual os Kuikuro fazem parte. A despeito da grande importância desta espécie entre os povos da região, sua diversidade morfológica nunca foi devidamente investigada em pesquisa acadêmica. Nosso objetivo é apresentar e analisar os aspectos socioculturais envolvidos no cultivo de pequi que incidem sobre a diversidade varietal da espécie, aqui tomada como a diversidade percebida e nomeada pelos Kuikuro. O estudo foi realizado na aldeia Kuikuro de Ipatse entre 2002 e 2003 (Fausto) e 2010 e 2012 (Smith), por meio de entrevistas e registro audiovisual das atividades de colheita e processamento dos frutos. Observamos que os conhecimentos e práticas de seleção e cultivo de sementes de pequi favorecem sua diversidade intraespecífica nos pequizais cultivados. As análises aqui apresentadas contribuem para demonstrar a impossibilidade de dissociar patrimônio cultural e genético no contexto dos sistemas agrícolas amazônicos.
Palavras-chave
Agrobiodiversidade; Pequi; Caryocar; Kuikuro; Patrimônio genético; Conhecimentos tradicionais associados
Abstract
This article aims to investigate and document socio-cultural aspects of the cultivation, domestication and processing of pequi (Caryocar brasiliense) among the Kuikuro Indians of the Upper Xingu, for whom this fruit has important symbolic and nutritional roles. Pequi cultivation is shared by the nine indigenous peoples who constitute the regional multiethnic system of the Upper Xingu. Despite the species’ importance among these peoples, its morphological diversity has not been the subject of intensive research yet. We intend to correlate social and cultural aspects involved in the management practices of the Kuikuro and the morphological diversity of pequi. Fieldwork was carried out between 2002 – 2003 (Fausto) and 2010 – 2012 (Smith) in the Kuikuro village of Ipatse in the Xingu Indigenous Park. Fieldwork involved interviewing families, participative observations and audiovisual recording. We observed that the knowledge and practices of seed selection and cultivation favor intraspecific diversity of the cultivated pequi groves. Our analysis, thus, supports the inextricable connection between genetic and cultural heritage in Amazonian agricultural systems.
Keywords
Agrobiodiversity; Pequi; Caryocar; Kuikuro; Genetic heritage; Traditional knowledge
INTRODUÇÃO
É comum afirmar-se que a Amazônia é uma área de ‘megabiodiversidade natural’. Nas últimas décadas, estudos têm associado tal diversidade a atividades culturais de populações tradicionais, sobretudo indígenas. A diversificação do meio amazônico estaria vinculada, assim, não exclusivamente a processos de ordem natural, mas ao impacto de práticas de cultivo e utilização do território por estas populações (Posey, 1986POSEY, D. A. Etnobiologia: teoria e prática. In: RIBEIRO, B. (Ed.). Suma etnológica brasileira: etnobiologia. Petrópolis: Vozes, 1986. v. 1, p. 15-28.; Balée, 1989BALÉE, W. The culture of Amazonian forests. In: POSEY, D. A.; BALÉE, W. (Eds.). Resource management in Amazonia: indigenous and folk strategies. New York: New York Botanical Garden, 1989. (Advances in Economic Botany, v. 7). p. 1-21., 1993BALÉE, W. Biodiversidade e os índios amazônicos. In: CARNEIRO DA CUNHA, M.; VIVEIROS DE CASTRO, E. (Orgs.). Amazônia: etnologia e história indígena. São Paulo, NHII/USP, 1993. p. 383-393.; Heckenberger et al., 2003HECKENBERGER, M. J.; KUIKURO, A.; KUIKURO, O. T.; RUSSEL, C.; SCHMIDT, M.; FAUSTO, C.; FRANCHETTO, B. Amazonia 1492: pristine forest or cultural parkland? Science, v. 301, n. 5640, p. 1710-1714, 2003.; Erickson, 2008ERICKSON, C. Amazonia: the historical ecology of a domesticated landscape. In: SILVERMAN, H.; ISBELL, W. H. (Eds.). The handbook of South American archaeology. New York: Springer, 2008. p. 157-183.; ter Steege et al., 2013TER STEEGE, H.; PITMAN, Nigel C. A.; SABATIER, Daniel; BARALOTO, Christopher; SALOMÃO, Rafael P.; GUEVARA, Juan Ernesto; PHILLIPS, Oliver L.; CASTILHO, Carolina V.; MAGNUSSON, William E.; MOLINO, Jean-François; MONTEAGUDO, Abel; VARGAS, Percy Núñez; MONTERO, Juan Carlos; FELDPAUSCH, Ted R.; CORONADO, Eurídice N. Honorio; KILLEEN, Tim J.; MOSTACEDO, Bonifacio; VASQUEZ, Rodolfo; ASSIS, Rafael L.; TERBORGH, John; WITTMANN, Florian; ANDRADE, Ana; LAURANCE, William F.; LAURANCE, Susan G. W.; MARIMON, Beatriz S.; MARIMON JR., Ben-Hur; VIEIRA, Ima Célia Guimarães; AMARAL, Iêda Leão; BRIENEN, Roel; CASTELLANOS, Hernán; LÓPEZ, Dairon Cárdenas; DUIVENVOORDEN, Joost F.; MOGOLLÓN, Hugo F.; MATOS, Francisca Dionízia de Almeida; DÁVILA, Nállarett; GARCÍA-VILLACORTA, Roosevelt; DIAZ, Pablo Roberto Stevenson; COSTA, Flávia; EMILIO, Thaise; LEVIS, Carolina; SCHIETTI, Juliana; SOUZA, Priscila; ALONSO, Alfonso; DALLMEIER, Francisco; MONTOYA, Alvaro Javier Duque; PIEDADE, Maria Teresa Fernandez; ARAUJO-MURAKAMI, Alejandro; ARROYO, Luzmila; GRIBEL, Rogerio; FINE, Paul V. A.; PERES, Carlos A.; TOLEDO, Marisol; AYMARD C., Gerardo A.; BAKER, Tim R.; CERÓN, Carlos; ENGEL, Julien; HENKEL, Terry W.; MAAS, Paul; PETRONELLI, Pascal; STROPP, Juliana; ZARTMAN, Charles Eugene; DALY, Doug; NEILL, David; SILVEIRA, Marcos; PAREDES, Marcos Ríos; CHAVE, Jerome; LIMA FILHO, Diógenes de Andrade ; JØRGENSEN, Peter Møller; FUENTES, Alfredo; SCHÖNGART, Jochen; VALVERDE, Fernando Cornejo; FIORE, Anthony Di; JIMENEZ, Eliana M.; MORA, Maria Cristina Peñuela; PHILLIPS, Juan Fernando; RIVAS, Gonzalo; VAN ANDEL, Tinde R.; VON HILDEBRAND, Patricio; HOFFMAN, Bruce; ZENT, Eglée L.; MALHI, Yadvinder; PRIETO, Adriana; RUDAS, Agustín; RUSCHELL, Ademir R.; SILVA, Natalino; VOS, Vincent; ZENT, Stanford; OLIVEIRA, Alexandre A.; SCHUTZ, Angela Cano; GONZALES, Therany; NASCIMENTO, Marcelo Trindade; RAMIREZ-ANGULO, Hirma; SIERRA, Rodrigo; TIRADO, Milton; MEDINA, María Natalia Umaña; VAN DER HEIJDEN, Geertje; VELA, César I. A.; TORRE, Emilio Vilanova; VRIESENDORP, Corine; WANG, Ophelia; YOUNG, Kenneth R.; BAIDER, Claudia; BALSLEV, Henrik; FERREIRA, Cid; MESONES, Italo; TORRES-LEZAMA, Armando; GIRALDO, Ligia Estela Urrego; ZAGT, Roderick; ALEXIADES, Miguel N.; HERNANDEZ, Lionel; HUAMANTUPA-CHUQUIMACO, Isau; MILLIKEN, William; CUENCA, Walter Palacios; PAULETTO, Daniela; SANDOVAL, Elvis Valderrama; GAMARRA, Luis Valenzuela; DEXTER, Kyle G.; FEELEY, Ken; LOPEZ-GONZALEZ, Gabriela; SILMAN, Miles R. Hyperdominance in the Amazonian tree flora. Science, v. 342, n. 6156, p. 1243092-1-1243092-9, 2013.). No que toca à agrobiodiversidade1 1 Segundo a Organização das Nações Unidas para Agricultura e Alimentação (FAO), a agrobiodiversidade é resultante de processos evolutivos, como a seleção natural e a seleção artificial por agricultores, durante milênios, e engloba todos os recursos e espaços envolvidos na produção agrícola pelas sociedades humanas (FAO, 1999). Neste trabalho, estamos restringindo o termo agrobiodiversidade à diversidade específica e intraespecífica dos recursos genéticos vegetais e aos agroecossistemas nos quais eles se reproduzem. , que é o objeto precípuo deste artigo, a diversidade de variedades, espécies e paisagens cultivadas e/ou manejadas deve ser associada também à diversidade interna às próprias culturas indígenas. Em vez de tomá-las como uma totalidade homogênea, cumpre estudá-las em sua variabilidade. Neste texto, focalizaremos a fruticultura do pequi na região do alto Xingu, Mato Grosso, Brasil.
Nas últimas décadas, o desmatamento na região amazônica, decorrente da urbanização crescente, da expansão do mercado nacional de bens agropecuários e da ampliação da malha rodoviária, tem provocado impactos sobre a diversidade biológica, não apenas “natural” como também “cultural”, acarretando perdas locais de recursos domesticados e cultivados (Thrupp, 2000THRUPP, L. A. Linking agricultural biodiversity and food security: the valuable role of agrobiodiversity for sustainable agriculture. International Affairs, v. 76, n. 2, p. 265-281, 2000.). Esse processo está intimamente associado a transformações que ocorrem, hoje, entre os povos indígenas, em função das novas oportunidades de estudo e de trabalho, da inserção na economia monetária, da crescente atração pelas cidades e do impacto da chegada de meios de comunicação nas aldeias (Gordon, 2006GORDON, C. Economia selvagem: ritual e mercadoria entre os índios Xikrin-Mebêngôkre. São Paulo: Editora UNESP/Instituto Socioambiental/Núcleo de Transformações Indígenas, 2006.; Andrello, 2006ANDRELLO, G. Cidade do índio: transformações e cotidiano em Iauaretê. São Paulo: Editora UNESP, 2006.; Fausto, 2011FAUSTO, C.; FRANCHETTO, B.; MONTAGNANI, T. Les formes de la mémoire: art verbal et musique chez les Kuikuro du Haut-Xingu (Brésil). L’Homme, n. 197, p. 41-70, 2011.). Neste contexto de mudança, a erosão da agrobiodiversidade decorre da perda de conhecimentos e de práticas tradicionais de manejo características de sistemas agrícolas indígenas, que levaram séculos ou até milênios para se desenvolver (Kerr, 1986KERR, W. Agricultura e seleções genéticas de plantas. In: RIBEIRO, B. (Ed.). Suma etnológica brasileira: etnobiologia. Petrópolis: Vozes, 1986. v. 1, p. 159-172.; Clement, 1999CLEMENT, C. R. 1492 and the loss of Amazonian crop genetic resources. I. The relation between domestication and human population decline. Economic Botany, v. 53, p. 188-202, 1999.; Clement et al., 2009cCLEMENT, C. R.; BORÉM, A.; LOPES, M. T. G. Da domesticação ao melhoramento de plantas. In: BORÉM, A.; LOPES, M. T. G.; CLEMENT, C. R. (Eds.) Domesticação e melhoramento: espécies amazônicas. Viçosa: UFV, 2009c. p. 11-38.). Embora os sistemas agrícolas indígenas sejam constantemente renovados a partir da incorporação de novos elementos, as rápidas mudanças socioambientais que estão ocorrendo na região podem comprometer sua resiliência e capacidade de reprodução.
Desde os anos 1990, a conservação da agrobiodiversidade tem recebido atenção crescente, em função de seu papel na alimentação mundial (Wood e Lenné, 1997WOOD, D.; LENNÉ, J. M. The conservation of agrobiodiversity on-farm: questioning the emerging paradigm. Biodiversity & Conservation, v. 6, n. 1, p. 109-129, 1997.)2 2 Segundo Thrupp (2000), mais de 20% dos recursos alimentares mundiais são provenientes de sistemas agrícolas tradicionais. , sendo particularmente relevantes os estudos sobre conservação em seu próprio contexto ecológico e sociocultural, a chamada conservação on-farm (Bellon et al., 1997BELLON M.R.; PHAM J. L.; JACKSON, M. T. Genetic conservation: a role for rice farmers. In: MAXTED N.; FORD-LLOYD B. V.; HAWKES J. G. Plant genetic conservation: the in situ approach. London: Chapman and Hall, 1997. p. 263-289.; Elias et al. 2001ELIAS, M.; MCKEY, Doyle; PANAUD, Olivier; ANSTETT, Marie Charlotte; ROBERT, Thierry. Traditional management of cassava morphological and genetic diversity by the Makushi Amerindians (Guyana, South America): perspectives for on-farm conservation of crop genetic resources. Euphytica, v. 120, n. 1, p. 143-157, 2001.; Clement et al., 2009aCLEMENT, C. R.; SANTOS, RonaldoP.; DESMOULIERE, Sylvain J. M.; FERREIRA, Evandro J. L.; FARIAS NETO, JoãoTomé. Ecological adaptation of wild peach palm, its in situ conservation and deforestation-mediated extinction in southern Brazilian Amazonia. PloS ONE, v. 4, n. 2, p. e4564, 2009a.). Contudo, esta literatura ainda está predominantemente centrada nas bases genéticas e agronômicas da agrobiodiversidade, sem dedicar igual atenção aos processos socioculturais responsáveis por sua origem e manutenção (Emperaire, 2006EMPERAIRE, L. Histórias de plantas, histórias de vida: uma abordagem integrada da diversidade agrícola tradicional na Amazônia. Revista da Sociedade Brasileira de Etnobiologia e Etnoecologia, v. 3, p. 189-198, 2006.; Rival, 2007RIVAL, L. Domesticating the landscape, producing crops, and reproducing society in Amazonia. In: PARKIN, D.; ULIJASZEK, S. (Eds.). Convergence and emergence: towards a new holistic anthropology? Oxford: Berghahn Books, 2007. p. 72-90.; Rival e Mckey, 2008RIVAL, L.; MCKEY, D. 2008. Domestication and diversity in manioc (Manihot esculenta Crantz ssp. esculenta, Euphorbiaceae). Current Anthropology, v. 49, n. 6, p. 1119-1128, 2008.). Estudos sobre a diversidade intraespecífica da mandioca junto às populações indígenas e locais na Amazônia têm mostrado que ações efetivas de conservação da agrobiodiversidade dependem de uma melhor compreensão sobre as relações entre diversidade per se e o sistema agrícola enquanto conjunto de práticas inseridas em um contexto sociocultural mais amplo (Emperaire, 2002EMPERAIRE, L. A agrobiodiversidade em risco: o exemplo das mandiocas na Amazônia. Ciência Hoje, v. 32, n. 187, p. 28-33, 2002., 2005EMPERAIRE, L. A biodiversidade agrícola na Amazônia brasileira: recurso e patrimônio. Revista do Patrimônio Histórico e Artístico Nacional, v. 32, p. 23-45, 2005.; Heckler e Zent, 2008HECKLER, S.; ZENT, S. Piaroa manioc varietals: hyperdiversity or social currency? Human Ecology, v. 36, n. 5, p. 679-697, 2008.).
Isso é válido não apenas para os ‘produtos das roças’, mas também para recursos florestais perenes, sobretudo árvores frutíferas, que são importantes fontes de alimentação e renda entre povos e comunidades locais na Amazônia. De acordo com Clement e colaboradores (2010), 68% dos recursos vegetais amazônicos domesticados ou em processo de domesticação são espécies lenhosas, o que demonstra a importante contribuição desse grupo de plantas à agrobiodiversidade. Segundo esses autores, há vários e importantes estudos sobre a diversidade genotípica e fenotípica de espécies arbóreas ou lenhosas na Amazônia focados em processos de domesticação de plantas e de paisagens por sociedades locais, entre os quais se incluem espécies como a castanheira-do-brasil (Bertholletia excelsa), a pupunha (Bactris gasipaes), o cacau (Theobroma cacao) e o guaraná (Paullinia cupana var. sorbilis) (Clement et al., 2010CLEMENT, C. R.; CRISTO-ARAÚJO, M.; D’EECKENBRUGGE, G. C.; PEREIRA, A. A.; PICANÇO-RODRIGUES, D. Origin and domestication of native Amazonian crops. Diversity, v. 2, p. 72-106, 2010., 2009bCLEMENT, C. R.; RIVAL, L.; COLE, D. M. Domestication of peach palm (Bactris gasipaes Kunth): the roles of human mobility and migration. In: ALEXIADES, M. N. (Ed.). Shifting spaces, changing times: mobility, migration and displacement in indigenous lowland South America, Oxford: Berghahn Books, 2009b. p. 117-140.). Contudo, há ainda poucos estudos centrados na ‘dimensão sociocultural’ do cultivo e na domesticação local de espécies arbóreas, sobretudo no contexto de sistemas agrícolas indígenas.
Neste artigo, não cabe uma discussão aprofundada sobre o que frequentemente é chamado na literatura de ‘componente’ ou ‘dimensão sociocultural’ de um sistema natural, mas é preciso notar que estas expressões são imperfeitas e, se as utilizamos aqui, o fazemos de forma provisória. Elas supõem uma dicotomia primeira entre natureza e cultura, sendo o problema subsequente o de determinar a relação entre estes termos – relação esta normalmente pensada como mera interpretação cultural, sempre variável, de fatos naturais universais. Autores contemporâneos colocam esse paradigma em questão (Latour, 1991LATOUR, B. Nous n’avons jamais été modernes: essai d’anthropologie symétrique. Paris: Editions La Découverte, 1991.; Ingold, 2000INGOLD, T. The perception of the environment: essays on livelihood, dwelling and skill. London/New York: Routledge, 2000.; Descola, 2005DESCOLA, P. Par-delà nature et culture. Paris: Gallimard, 2005.; Viveiros de Castro, 1996VIVEIROS DE CASTRO, E. Os pronomes cosmológicos e o perspectivismo ameríndio. Mana, v. 2, n. 2, p. 115-144, 1996.), atribuindo-lhe um caráter limitado a uma ontologia específica: o naturalismo ocidental moderno. Quando estudamos populações indígenas, é particularmente arriscado manter-se no interior desse paradigma, pois, como afirma Descola:
diferentemente do dualismo mais ou menos irredutível, que em nossa visão moderna do mundo, rege a distribuição dos humanos em dois campos ontologicamente distintos, as cosmologias amazônicas exibem uma escala dos seres em que as diferenças entre os homens, as plantas e os animais são de grau e não de natureza (Descola, 2000DESCOLA, P. Ecologia e cosmologia. In: DIEGUES, A. C. S. (Ed.). Etnoconservação: novos rumos para a proteção da natureza nos trópicos. São Paulo: Hucitec, 2000. p. 149-164., p. 151).
Compreender esse regime ontológico é fundamental se quisermos propor modos sui generis de proteção e salvaguarda de conhecimentos tradicionais associados à biodiversidade, que possam ser efetivamente utilizados por povos indígenas na promoção de seus direitos. Neste texto, buscamos estudar o cultivo e a domesticação do pequizeiro (Caryocar brasiliense, Caryocaraceae) entre os Kuikuro do alto Xingu, focalizando o regime de socialidade atribuído à espécie e aos espaços a ela associados. Vamos nos deter nas práticas, técnicas, preferências, ideias e representações, bem como nos conhecimentos associados às atividades de seleção, cultivo, manejo e uso da espécie, que, no contexto indígena do alto Xingu, caracteriza-se como um recurso agrícola. Na medida do possível, evitaremos a oposição entre o fato e o feito, embora, ao falarmos de ‘domesticação’, estejamos postulando um processo no qual uma população biológica natural vai sendo transformada pelo manejo humano até depender deste para a sua reprodução.
É-nos inevitável formular o problema nesses termos, pois se trata aqui de fazer os conhecimentos tradicionais dialogarem com nosso sistema de proteção e salvaguarda. Esta necessidade resulta de um processo relativamente recente de fluxo de recursos vegetais para fora do Parque Indígena do Xingu (PIX), e da percepção de que variedades de pequi selecionadas e melhoradas pelos índios estão sendo apropriadas pela população não indígena devido ao seu potencial comercial. Nosso estudo nasce dessa percepção indígena e da consequente demanda de proteção, sendo um objetivo central mostrar empiricamente que o pequi xinguano deve ser considerado mais do que um recurso genético e que as populações cultivadas dessa planta no PIX englobam em sua estrutura ecológica e biológica “conhecimentos tradicionais intrínsecos” (Emperaire, 2006EMPERAIRE, L. Histórias de plantas, histórias de vida: uma abordagem integrada da diversidade agrícola tradicional na Amazônia. Revista da Sociedade Brasileira de Etnobiologia e Etnoecologia, v. 3, p. 189-198, 2006., comunicação pessoal)3 3 Estamos chamando de ‘conhecimentos tradicionais intrínsecos’ os aspectos socioculturais que se referem não somente ao conhecimento, mas também ao trabalho indígena empregado na seleção de sementes, nas técnicas e práticas de produção de mudas, no transplante das mudas e nos tratos silviculturais envolvidos no processo de domesticação ou melhoramento genético tradicional do pequizeiro. . Cabe-nos o desafio de argumentar em favor do caráter indissociável entre patrimônio genético e cultural, a fim de estabelecer uma ponte de tradução entre a perspectiva indígena e as noções mobilizadas pelo conceito de agrobiodiversidade.
Uma última precisão antes de passarmos à análise: nosso estudo foi realizado entre os Kuikuro, povo de língua karib que, juntamente com outros nove povos, forma a constelação altoxinguana e ocupa a porção sul do Parque Indígena do Xingu (PIX), no Mato Grosso. Trata-se de um sistema multiétnico e plurilíngue, que começou a se formar no século IX d.C., com a chegada de uma primeira população colonizadora de língua arawak à qual se juntariam, a partir do século XVI, povos de língua karib e tupi (Heckenberger, 2001HECKENBERGER, M. Estrutura, história e transformação: a cultura xinguana na longue durée, 1000-2000 D.C. In: FRANCHETTO, B.; HECKENBERGER, M. (Orgs.). Os povos do alto Xingu: história e cultura. Rio de Janeiro: UFRJ, 2001. p. 21-62., 2005HECKENBERGER, M. J. The ecology of power. New York: Routledge, 2005.; Fausto et al., 2008FAUSTO, C. Donos demais: maestria e domínio na Amazônia. Mana, v. 14, n. 2, p. 329-366, 2008.). Esse sistema social único, que preserva as diferenças linguísticas – mas partilha um sistema econômico, político e ritual comum –, formou-se em uma zona igualmente singular do ponto de vista ambiental: a transição entre o cerrado e a floresta amazônica. Um dos denominadores comuns dessa constelação altoxinguana, com um valor alimentar e simbólico excepcional, é justamente a fruticultura do pequi.
Ao tratar do pequizeiro, estamos nos referindo à espécie Caryocar brasiliense Camb., uma das espécies do gênero Caryocar, que ocorre naturalmente em regiões dominadas pela fitofisionomia de cerrado no Brasil central e em parte da Amazônia legal. O pequizeiro produz um fruto – o pequi – que é bastante apreciado na cozinha regional. Embora este fruto seja geralmente explorado por meio de atividade extrativista, os povos indígenas do alto Xingu cultivam pequizeiros em antigas roças de mandioca, produzindo pomares dessa espécie em uma vasta área em torno das aldeias. Estudos morfométricos e genéticos ainda não concluídos mostram evidências de que os pequizeiros cultivados pelos povos indígenas do alto Xingu encontram-se em processo de domesticação local e podem ser considerados como componentes de uma variedade local de Caryocar brasiliense (Smith, 2013SMITH, M. Árvores de cultura: cultivo e uso do pequi (Caryocar sp., Caryocaraceae) entre os Kuikuro do alto Xingu, MT. 2013. 221 f. Tese (Doutorado em Desenvolvimento Sustentável: Política e Gestão Ambiental) – Universidade de Brasília, Brasília, 2013.).
METODOLOGIA
ÁREA DE ESTUDO
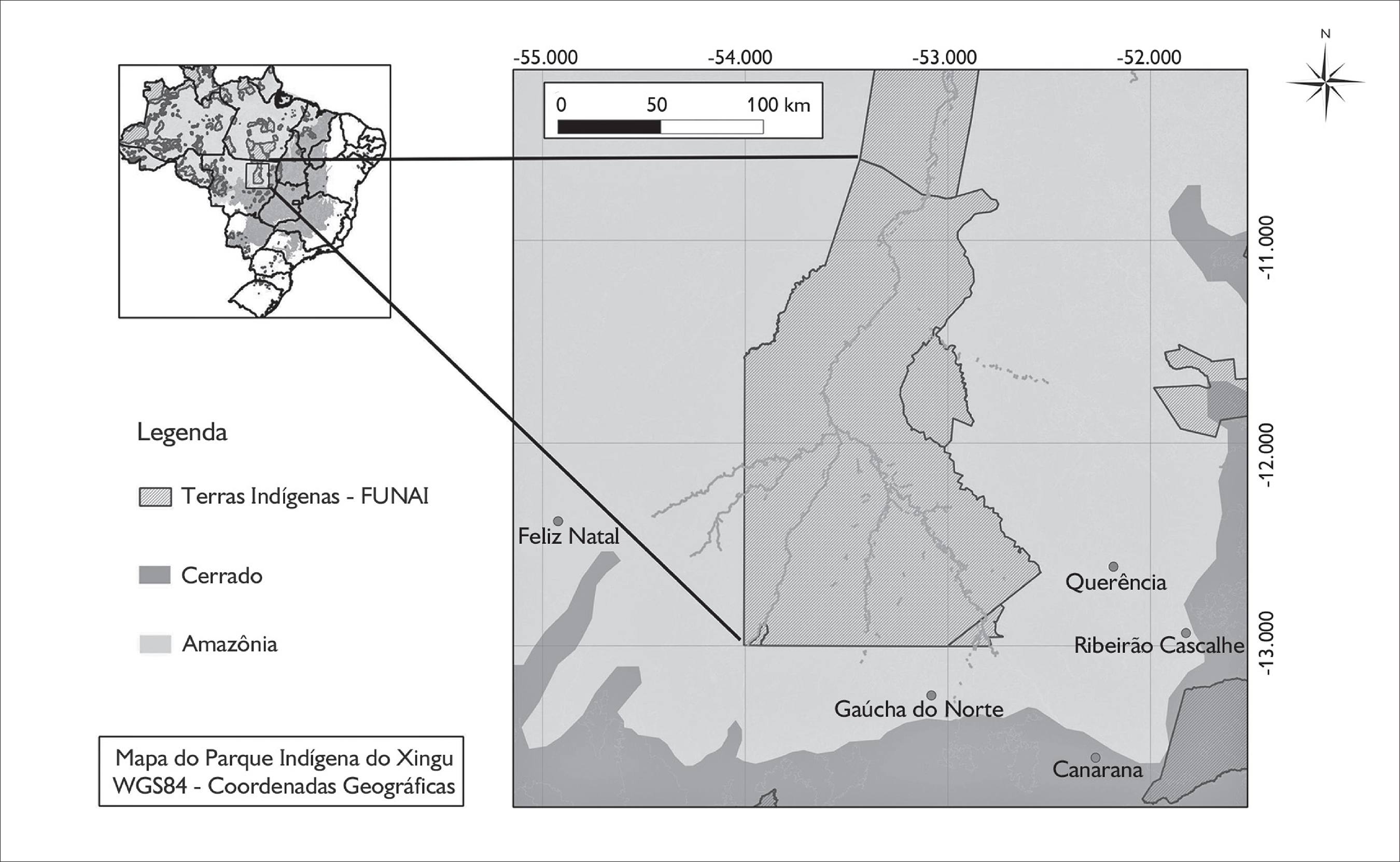
As informações aqui apresentadas foram levantadas na aldeia Kuikuro de Ipatse (12° 21’ 06.31” S, 53° 12’ 33.74” W) e em seu entorno, no Parque Indígena do Xingu (PIX). O PIX tem uma superfície de 2.642.003 ha e está localizado no nordeste do estado de Mato Grosso, com área incidente entre os municípios de Canarana, Paranatinga, São Félix do Araguaia, São José do Xingu, Gaúcha do Norte, Feliz Natal, Querência, União do Sul, Nova Ubiratã e Marcelândia (Figura 1). Falantes de uma variante dialetal do Karib Meridional (Franchetto, 2001FRANCHETTO, B. Línguas e histórias no alto Xingu. In: FRANCHETTO, B.; HECKENBERGER, M. (Orgs.). Os povos do alto Xingu: história e cultura. Rio de Janeiro: UFRJ, 2001. p. 111-156.; Meira e Franchetto, 2005MEIRA, S.; FRANCHETTO, B. The southern Cariban languages and the Cariban family. International Journal of American Linguistics, v. 71, n. 2, p. 127-192, 2005.), os Kuikuro são atualmente o povo mais numeroso do alto Xingu. Eles habitam a região compreendida entre as nascentes da lagoa de Tafununu, a leste do rio Culuene (principal formador do rio Xingu), e o rio Buriti, afluente do rio Curisevo, a oeste.
Localização da área de estudo no Parque Indígena do Xingu (PIX), Mato Grosso, região de transição entre os domínios do cerrado do Brasil central e da Amazônia.
Sua principal aldeia recebe o nome de Ipatse (‘pequena lagoa’) e localiza-se em uma “península” de terra firme, cuja vegetação original era formada predominantemente por floresta do tipo estacional perenifólia (Ivanauskas et al., 2008IVANAUSKAS, N. M.; MONTEIRO, R.; RODRIGUES, R. R. Classificação fitogeográfica das florestas do Alto Rio Xingu. Acta Amazonica, v. 38, n. 3, p. 387-402, 2008.), posicionada entre uma faixa de formações campestres e savânicas na beira do rio Culuene e um enorme buritizal, que se estende na beira do rio Buriti (Figura 2). Estas formações campestres e savânicas ocupam porção significativa das terras no alto Xingu e ocorrem em terrenos mais baixos, que sofrem inundações periódicas por água da chuva. Algumas áreas mais elevadas desses terrenos formam pequenas ilhas de vegetação (campos de murundus4 4 Os campos de murundus ocorrem geralmente no cerrado e são constituídos por áreas planas, que sofrem inundação no período de chuvas, e por morrotes (murundus), que não inundam. Os morrotes menores são cobertos por vegetação campestre e os maiores, por vegetação lenhosa do cerrado (Resende et al., 2004). ) e, quando mais extensas, variam em um gradiente entre campo sujo, cerrado típico, cerradão e matas de galeria na beira de pequenos cursos d’água.
Vista da aldeia Ipatse em uma ‘península’ de terra firme – área de floresta que os Kuikuro chamam genericamente de itsuni –, rodeada por formações savânicas que sofrem inundação sazonal denominadas óti.
Assim como os demais povos indígenas do alto Xingu, os Kuikuro praticam uma horticultura de coivara, baseada, sobretudo, no cultivo de cerca de 30 a 45 variedades diferentes de mandioca (Carneiro, 1957CARNEIRO, R. L. Subsistence and social structure: an ecological study of the Kuikuro Indians. 1957. Tese (Doutorado em Antropologia) – University of Michigan, Michigan, 1957., 1983CARNEIRO, R. L. The cultivation of manioc among the Kuikuro of the upper Xingu. In: HAMES, R. B.; VICKERS, W. T. (Eds.). Adaptive responses in native Amazonians. New York: Academic Press, 1983. p. 65-111.; Fausto, dados de campo inéditos em 2002 e 2014), às quais somam-se variedades de milho, batata-doce, abóbora, banana, entre outras espécies plantadas em terra preta (egepe5 5 Carneiro (1957, 1983) levantou 46 variedades cultivadas e nomeadas; Fausto levantou 35 variedades, em trabalho preliminar realizado em 2002 com Morgan Schmidt. Em 2014, um levantamento mais cuidadoso, realizado com Helena Cooper, permitiu corrigir as homonímias na lista original de Carneiro, confirmando o número de variedades reconhecidas a 36. Mais da metade, contudo, não está mais sendo plantada pelos Kuikuro, tornando urgente um projeto de salvaguarda. ). A proteína animal provém essencialmente da pesca, já que os altoxinguanos não comem mamíferos, à exceção de uma espécie de macaco, e consomem apenas algumas poucas espécies de aves.
Do ponto de vista fitogeográfico e morfoclimático, o PIX encontra-se em uma área de transição entre os domínios da floresta amazônica e do cerrado do Brasil central, descrita como zona de tensão ecológica (Veloso et al., 1991VELOSO, H. P.; RANGEL FILHO, A. L. R.; LIMA, J. C. A. Classificação da vegetação brasileira, adaptada a um sistema universal. Rio de Janeiro: IBGE, 1991. 112 p.apudIvanauskas et al., 2004IVANAUSKAS, N. M.; MONTEIRO, R.; RODRIGUES, R. R. Estrutura de um trecho de floresta Amazônica na bacia do alto rio Xingu. Acta Amazonica, v. 34, n. 2, p. 275-299, 2004.). Trata-se de uma região ecologicamente rica, combinando formações de cerrado lato sensu com uma área florestal (“Floresta Estacional Perenifólia”), que constitui a borda sul da floresta amazônica (Ivanauskas et al., 2004IVANAUSKAS, N. M.; MONTEIRO, R.; RODRIGUES, R. R. Estrutura de um trecho de floresta Amazônica na bacia do alto rio Xingu. Acta Amazonica, v. 34, n. 2, p. 275-299, 2004.). O PIX abrange, assim, um mosaico de vegetação com formações de cerrado, campos, floresta de várzea, florestas de terra firme, além de florestas em terras pretas arqueológicas (Villas-Bôas, 2012VILLAS-BÔAS, A. De olho na bacia do Xingu. São Paulo: Instituto Socioambiental, 2012. (Série Cartô Brasil Socioambiental, n. 5). 61 p.).
Segundo o sistema de Köppen, o clima da região é tropical chuvoso de savana (Aw), definido por médias de temperaturas superiores a 18 °C, apresentando duas estações bem definidas, uma chuvosa, que vai de novembro a abril, e outra seca, entre maio e outubro (Ivanauskas et al., 2004IVANAUSKAS, N. M.; MONTEIRO, R.; RODRIGUES, R. R. Estrutura de um trecho de floresta Amazônica na bacia do alto rio Xingu. Acta Amazonica, v. 34, n. 2, p. 275-299, 2004.). Foram caracterizados dois tipos principais de solo no alto Xingu, de acordo com a classificação da Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA): o latossolo vermelho-amarelo distrófico típico, que é predominante e ocorre nos relevos planos a suave-ondulados das áreas interfluviais; e o neossolo flúvico Tb distrófico típico nas áreas florestais sujeitas à inundação periódica (Ivanauskas et al., 2004IVANAUSKAS, N. M.; MONTEIRO, R.; RODRIGUES, R. R. Estrutura de um trecho de floresta Amazônica na bacia do alto rio Xingu. Acta Amazonica, v. 34, n. 2, p. 275-299, 2004.). Nas áreas de terra firme ocorrem, ainda, extensas manchas de solos ricos em matéria orgânica, cuja origem remete à ação antrópica passada: tratam-se das chamadas “terras pretas de índio” (Schmidt e Heckenberger, 2009SCHMIDT, M. J.; HECKENBERGER, M. J. Formação de Terra Preta na região do alto Xingu: resultados preliminares. In: TEIXEIRA, W.; KERN, D. C.; MADARI, B.; LIMA, H. N. (Eds.). As terras pretas de índio da Amazônia: sua caracterização e uso deste conhecimento na criação de novas áreas. Manaus: EDUA, 2009. p. 103-126.; Schmidt, 2010SCHMIDT, M. J. Reconstructing tropical nature: prehistoric and modern anthrosols (Terra Preta) in the Amazon rainforest, Upper Xingu river, Brazil. 2010. Tese (Doutorado em Geografia) –University of Florida, Gainesville, 2010.), que se encontram associadas às aldeias pré-históricas.
COLETA DE DADOS
As informações aqui apresentadas foram obtidas a partir do registro de narrativas, mitos, rituais, práticas de cultivo, técnicas de uso e armazenamento associadas ao pequi por meio de entrevistas semiestruturadas e registro audiovisual das atividades de colheita e processamento dos frutos. Os dados foram coletados em dois momentos distintos: primeiro, em 2002 e 2003, por Carlos Fausto, no bojo de um projeto de documentação e produção cinematográfica, em parceria com a Associação Indígena Kuikuro do alto Xingu (AIKAX) e o Vídeo nas Aldeias; segundo, em 2011 e 2012, por Maira Smith, no bojo do projeto “Documentação do conhecimento e uso de vegetais entre os Kuikuro do alto Xingu”, financiado pelo Museu do Índio6 6 As entrevistas realizadas por Maira Smith foram mediadas pelo professor indígena Sepé Ragati Kuikuro, que ajudou no planejamento das atividades, na elaboração de questões e na tradução. . Dados detalhados sobre o manejo e o uso do pequi entre os Kuikuro foram complementados pela coleta de narrativas e exegeses versando sobre a origem do pequi, seus espíritosdonos e o ritual do Hugagü, associado à sua colheita.
Em 2011, Smith e equipe realizaram um censo detalhado de todos os moradores da aldeia de Ipatse, identificando aqueles que já tinham plantado pequi no entorno da localidade, a partir do qual foram selecionadas algumas famílias para serem acompanhadas durante a safra do fruto. Delimitaram-se, assim, 12 pequizais (unidades de produção de pequi) pertencentes a dez famílias nucleares. O(a) responsável pelo plantio de cada pequizal foi entrevistado(a) em seus respectivos pomares, perguntando-se-lhes sobre: a) a data aproximada do plantio; b) a procedência das sementes utilizadas; c) os métodos e as técnicas de cultivo e de manejo; d) o dono atual do pomar; e e) os vínculos sociais (de parentesco) do dono atual com o responsável pelo plantio.
Estes dados foram cruzados com as informações levantadas no censo sobre as relações de parentesco nas famílias amostradas e sobre a história de vida de cada um dos responsáveis pelo plantio de pequi. Após as entrevistas in loco com os horticultores, realizamos o acompanhamento da etapa de coleta, do processamento e uso do pequi entre as famílias amostradas, utilizando a metodologia de observação-participante e o registro fotográfico. Pelo menos 30 árvores de pequi contidas dentro de cada parcela (pequizal individual amostrado) foram identificadas com placas de metal numeradas, as quais foram mapeadas com GPS (Figura 3). Nas parcelas, anotamos com apoio do responsável pelo plantio as variedades de cada árvore, definidas pela morfologia dos frutos. A partir da observação das características utilizadas pelos Kuikuro para classificar as variedades de pequi, elaboramos uma lista de descritores morfológicos dos frutos, com o apoio de Sepé Kuikuro.
ANÁLISE DE DADOS
Todos os dados quantitativos acerca de cada indivíduo de pequi identificado foram passados para uma planilha em Excel. As entrevistas gravadas em língua kuikuro foram traduzidas para o português por Yamaluí Mehinako, com suporte do programa ELAN 4.3.2 (Wittenburg et al., 2006WITTENBURG, P.; BRUGMAN, H.; RUSSEL, A.; KLASSMANN, A.; SLOETJES, H. Elan: a professional framework for multimodality research. In: INTERNATIONAL CONFERENCE ON LANGUAGE RESOURCES AND EVALUATION, 5., Genoa, 2006. Proceedings… Genoa: ELRA, 2006. p. 1556-1559. Disponível em: <http://www.lrec-conf.org/proceedings/lrec2006/pdf/153_pdf.pdf>. Acesso em: 10/05/2014
http://www.lrec-conf.org/proceedings/lre...
). Os dados qualitativos derivados de entrevistas e observação-participante, bem como o material fotográfico produzido foram organizados em relatórios de viagem, encaminhados aos financiadores do projeto7
7
A Associação Indígena Kuikuro do alto Xingu (AIKAX) mantém um termo de cooperação com o Museu do Índio (Fundação Nacional do Índio – FUNAI) para acomodar o acervo resultante de atividades de pesquisa e documentação cultural produzidas na aldeia.
.
Para quantificar a diversidade intraespecífica de pequis cultivados nas 12 populações (parcelas) amostradas, utilizamos os nomes kuikuro do que eles entendem como ‘variedades’, não necessariamente congruente com as noções de variedade usadas na botânica, agronomia e/ou em etnobotânica. Por se tratar de uma espécie alógama, de longo ciclo de vida e que é plantada pelos índios exclusivamente pela produção de mudas a partir de sementes, há uma variação morfológica expressiva nos frutos de pequi dentro de cada e entre as populações cultivadas, fazendo com que os nomes de cada tipo indicado não expressem a grande variação intraespecífica observada e manejada pelos Kuikuro em seus pequizais.
Como o número de indivíduos por pequizal se mostrou muito variável, limitamos o levantamento a cerca de 30 árvores em cada parcela. Duas parcelas com poucos indivíduos foram inseridas na amostra, já que as sementes ali plantadas foram trazidas de um referencial territorial importante para os povos karib do alto Xingu. Em geral, as mudas de pequi são todas plantadas em uma roça de mandioca em um único evento, fazendo com que todos os pequizeiros sejam da mesma idade dentro de cada unidade amostral (parcela)8 8 Inicialmente, as mudas de pequi são plantadas na roça consorciadas com as plantas de mandioca. Quando a roça de mandioca deixa de ser produtiva, os pequizeiros em estágio juvenil crescem, formando um pomar, os quais são manejados pelos Kuikuro de modo a desfavorecer a regeneração florestal espontânea nestas áreas. .
Com o objetivo de encontrar um indicador de diversidade intraespecífica para caracterizar cada parcela (pequizal) amostrada, fizemos uma adaptação do índice de Shannon-Weaver (Shannon e Weaver, 1949SHANNON, C. E.; WEAVER, W. The mathematical theory of communication. Urbana: Illinois University, 1949. 117 p.). Este índice é geralmente utilizado para medir a diversidade de espécies (diversidade interespecífica) em comunidades biológicas (diversidade α), mas, neste trabalho, foi usado para quantificar a diversidade intraespecífica de pequis nomeada pelos Kuikuro em seus pequizais. Para a análise, consideramos cada pequizal individual como área amostral (parcela) e nosso universo amostral foi composto por 12 parcelas com tamanhos variáveis. Ressalte-se que a identificação das variedades de pequi em cada parcela foi realizada pelo indígena responsável pelo plantio do pequizal, juntamente com o pesquisador indígena, Sepé Kuikuro, que acompanhou todas as etapas do levantamento. Na adaptação do índice de Shannon-Weaver ao contexto desse trabalho, consideramos a fórmula H’ = -∑ pi (ln pi), onde pi = abundância relativa de cada variedade nomeada, calculada pela proporção dos indivíduos de uma variedade i, identificados na parcela pelo agricultor, sobre o número total de variedades nomeadas para cada pequizal. Como houve pouca variação nos padrões de diversidade intraespecífica entre os 12 pequizais, realizamos apenas uma análise qualitativa na comparação entre eles, sem utilizar análises estatísticas multivariadas para esta finalidade.
RESULTADOS
O PEQUI NO SISTEMA AGRÍCOLA KUIKURO
Os Kuikuro nomeiam duas categorias abrangentes de unidades de paisagem: itsuni, que pode ser traduzido como ‘mata’ ou ‘floresta’, e que corresponde ao que identificamos ecologicamente como formações florestais em terra firme; e óti (‘campo’), termo geral utilizado em menção às formações campestres e savânicas que ocorrem em terreno mais baixo na região (Figura 2). Dentro de cada uma destas categorias, os Kuikuro nomeiam e diferenciam unidades de paisagem mais específicas.
Aldeias e roças são abertas em áreas de floresta de terra firme (itsuni). Há basicamente dois tipos diferentes de roça: as de mandioca, plantadas em áreas de terra ‘vermelha’, e os plantios em ‘terra preta’, denominada egepe em Kuikuro. Como dissemos, as terras pretas constituem manchas de terreno mais fértil, associadas a antigos assentamentos indígenas. A mandioca cresce em terreno menos fértil, mas o milho ou algodão são plantados nas roças de terra preta, geralmente mais distantes da aldeia. Além das roças de mandioca (tuhinhaho)9 9 O termo geral utilizado para a roça de mandioca é tuhinhaho, mas existem termos mais específicos usados para as diferentes fases da roça, desde o momento de sua abertura, passando pelo plantio, podas sucessivas, replantio até o abandono para pousio. e das roças feitas em terra preta, os Kuikuro nomeiam ainda uma terceira unidade de produção agrícola, unha, cuja tradução em português se aproxima da noção de ‘quintal’. Os quintais são muito utilizados para o plantio de frutíferas nativas e, sobretudo, exóticas, formando pomares ao redor da aldeia, de uso mais ou menos comum da comunidade. Somente o pequi possui um pomar específico na mata – o pequizal (uika) –, que pertence a uma só pessoa e cujos frutos são apropriados apenas por sua família.
O sistema agrícola kuikuro se estrutura, pois, a partir desses três espaços (tuhinhaho, egepe e unha), mas o pequizal (uika) também pode ser identificado como um agroecossistema e como um tipo de espaço de produção agrícola, vinculado temporalmente às roças de mandioca (tuhinhaho). A dinâmica de uso do itsuni faz com que este espaço se caracterize como um mosaico, composto por habitats com diferentes níveis de intervenção humana, desde áreas cultivadas temporárias (roças), pomares permanentes (pequizais), passando por capoeiras de diferentes idades e sapezais, até a floresta madura (Figura 4).
Representação esquemática da dinâmica da paisagem nos domínios do itsuni (lato sensu) – formações florestais de terra firme –, onde os Kuikuro constroem suas aldeias e praticam as atividades agrícolas. Os diferentes estágios coexistem no espaço em um mesmo período de tempo, formando um mosaico de habitats diversificados no itsuni.
No sistema agrícola kuikuro, há uma variedade significativa de plantas cultivadas, cada uma requerendo condições de solo e nutrientes específicos, estando associadas a um dos espaços produtivos anteriormente mencionados. Observamos aproximadamente 30 espécies cultivadas, destacando-se entre elas a mandioca, o pequi e o milho, não apenas pelo valor alimentício, mas por possuírem um mito de origem próprio, festas e rituais associados. Consideramos o pequi não somente uma das três espécies mais relevantes do sistema agrícola kuikuro, como também um importante marcador cultural, isto é, um dos elementos distintivos daquilo que constitui, para os próprios índios, uma ‘identidade xinguana’, que os diferencia de outros grupos indígenas da região.
A ORIGEM DO PEQUI
Os Kuikuro concebem os principais elementos biológicos de seu sistema agrícola e os espaços a eles associados em consonância com o modelo de organização das relações sociais humanas altoxinguanas. Nesse sentido, as espécies agrícolas representam muito mais do que simples recursos alimentares. A mandioca, o pequi e o milho são pivôs de um conjunto de associações que reúnem populações humanas e não humanas: associações entre variedades ou morfotipos de uma mesma espécie, entre diferentes espécies cultivadas, entre estas e certos animais, entre estes animais e suas hipóstases “espirituais” (itseke) e, finalmente, entres estas e os humanos – configurando um coletivo de atores heterogêneos (Latour, 2005LATOUR, B. Reassembling the social: an introduction to actornetwork-theory. Oxford/New York: Oxford University Press, 2005.). A socialidade atribuída às espécies agrícolas pelos Kuikuro incide diretamente nas práticas de manejo destes recursos e, consequentemente, em sua diversidade genética e morfológica.
O cultivo de mudas de pequi e os tratos silviculturais dos pequizais vinculam-se a diversos elementos do seu mito de origem, no qual a árvore de pequi brota do umbigo de um jacaré morto por um índio de nome Magiká. A seguir, resumimos uma versão padrão desta narrativa10 10 Baseada principalmente nas versões contadas por Haitsehü Kalapalo, em 2011, e Tugupé Kuikuro, em 2002, durante as filmagens de “Imbé Gikegü: cheiro de pequi”. . Conta-se que as esposas de Magiká iam todos os dias à roça para arrancar mandioca, mas antes iam à beira da lagoa, onde chamavam seu amante, o jacaré, para fazer sexo com elas. Um dia, Magiká saiu para caçar e quando estava para flechar uma cotia, esta fez-se gente e pediu para não ser morta em troca de um segredo. Contou, então, a ele que suas esposas eram amantes do jacaré. A cotia levou Magiká até a beira para ver a traição com seus próprios olhos e disse-lhe para matar o jacaré com flechas invisíveis, bem no momento em que ele estivesse gozando. Assim fez o marido traído. Ao verem o amante morto, as duas irmãs ficaram inconsoláveis e com muita raiva do marido: enterraram o jacaré e expulsaram Magiká de casa, o qual foi morar na casa dos homens, no centro da aldeia. Algum tempo depois, elas foram visitar a sepultura de seu amante e notaram que várias árvores frutíferas haviam brotado de diferentes partes de seu corpo11 11 Dos testículos, nasce a mangaba, que embora não seja plantada, encontra-se no entorno das aldeias e é importante fonte alimentar em novembro, auge da colheita do pequi. Do pênis, nasce um fruto silvestre localmente denominado akaga e dos olhos nascem ugukumi. A associação entre o pequi e esses frutos deve-se à época de frutificação. . Bem ao centro, partindo do umbigo, havia um broto avermelhado que daria origem ao primeiro pequizeiro. Elas cuidaram dessa árvore até que começou a produzir frutos e assim passaram a conhecer e usar o pequi como alimento e para a produção de óleo.
Note-se que, no mito, o pequi ocupa uma posição central em relação às outras espécies frutíferas (não cultivadas) que também surgiram a partir do jacaré: ele brota do centro (umbigo) e determina aspectos da fenologia destas outras espécies, com base em seu próprio ciclo ecológico. Algumas versões do mito narram ainda o processo de diversificação do pequizeiro altoxinguano. Este processo é pensado como uma ramificação: o tronco da variedade conhecida como suümkogu (que produz os maiores frutos) ergue-se do umbigo do jacaré e as demais variedades vão aparecendo como ramos deste tronco central.
O mito vai além da origem do pequi, pois explica sua denominação, como se estabeleceram os procedimentos associados ao cultivo e uso da espécie, a sua relação com a festa de Hugagü e também com a sexualidade12 12 Não cabe aqui desenvolver este aspecto, mas ele é central na construção do filme “Cheiro de pequi” – cheiro este que é o da vagina, a qual é modelada em cera e mostrada às mulheres durante o ritual do Hugagü. Sobre o mesmo ritual entre os povos arawak do Xingu, ver Coelho (1992) e Gregor (1985). . Todos esses episódios posteriores põem em cena os gêmeos Taũgi (sol) e Aulukumã (lua), demiurgos da cosmologia kuikuro, que ensinaram a preservar a polpa do pequi em água, evitando doenças e permitindo seu uso ao longo de muitos meses13 13 Esse é outro fato notável e singular da tecnologia xinguana associada à fruticultura do pequi. Eles constroem um cesto onde colocam a polpa, o qual é hermeticamente fechado e depositado em uma lagoa. A bebida feita a partir do pequi, conhecida como imbene, pode ser tomada durante o ritual do Quarup, na estação seca, cerca de seis meses após o final da safra. . As formas corretas de cultivar, manejar, usar e armazenar a polpa de pequi ensinadas pelos demiurgos continuam sendo adotadas hoje.
OS DONOS DO PEQUI
O mundo em que vivem os Kuikuro é recortado por domínios: tudo pode ser a casa (domus) de alguém. Por isso, todos os espaços ‘naturais’ – rios, lagos, acidentes geográficos ou mesmo aquela curva do rio, o centro daquela lagoa, aquelas árvores ali adiante – costumam ter ‘donos’ (oto), os quais são responsáveis por engendrá-los e deles cuidar. Na feliz expressão de Descola (1986)DESCOLA, P. La nature domestique: symbolisme et praxis dans l’écologie des Achuar. Paris: Maison des Sciences de l’Homme, 1986., a natureza aqui é doméstica. Estes domínios são múltiplos e não representam unidades discretas: o modelo não é o da propriedade privada exclusiva (Fausto, 2008FAUSTO, C.; FRANCHETTO, B.; HECKENBERGER, M. J. Language, ritual and historical reconstruction. In: HARRISON, K. D.; DAVID, S. R.; DWYER, A. (Eds.). Lessons from documented endangered languages. Amsterdam: John Benjamins, 2008. p. 129-157., 2012aFAUSTO, C. Too many owners: mastery and ownership in Amazonia. In: BRIGHTMAN, M.; GROTTI, V. E.; ULTURGASHEVA, O. (Orgs.). Animism in forest and tundra: personhood, animals, plants and things in contemporary Amazonia and Siberia. London: Berghahn, 2012a. p. 85-105.). Ainda assim, posto que tudo pode ser a casa de alguém e todo ente pode ter um dono, é preciso associar-se a eles – por meio de cantos, rezas e oferta alimentar –, para que permitam o uso desses espaços ou de algum recurso. É preciso movê-los para que se disponham a ceder sem causar doenças nos humanos, concedendo, assim, uma boa pesca, uma boa coleta ou uma boa safra.
No caso do pequi, esse universo é ainda mais complexo, pois ele possui vários donos, sendo o principal deles o tukuti-kuegü ou hiper-beija-flor, que é um itseke (bicho-espírito)14 14 Os Kuikuro costumam traduzir itseke por “bicho”, pois os itseke que povoam o mundo xinguano são, em sua maioria, animais em sua condição de pessoa; i.e., animais dotados de intencionalidade, reflexividade e capacidade comunicativa (ver Fausto, 2012b). Para a tradução do sufixo –kuegü como “hiper”, ver Franchetto (2003). . Além do beija-flor, contam-se ainda “a hiperarara, a hipermaritaca, o gavião [ele mesmo tolo oto, ‘dono dos pássaros], o hipermacaco e o hiperjacaré. Não é qualquer pássaro que pode ser dono do pequi, eles têm que ser poderosos e bonitos, os chefes dos pássaros” (Asahü Kuikuro, 2010 – tradução de Takumã Kuikuro). Todos esses animais estão associados ao ciclo do pequi, e em especial à polinização e dispersão de sementes15 15 Embora os morcegos sejam os principais polinizadores de C. brasiliense no período noturno, algumas espécies de beija-flor também visitam as flores desta árvore nas primeiras horas do dia para sugar néctar, agindo como polinizadores cruzados e favorecendo a diversidade genética nas populações cultivadas (Gribel e Hay, 1993). Por seu turno, a arara e a cotia possuem papel importante como dispersores naturais das sementes de pequi. , sendo que o beija-flor tem lugar de destaque, pois se atribui a ele a função de cuidar dos pequizeiros para que não sequem e produzam frutos bonitos. Como todo dono, contudo, o hiper-beija-flor é cioso de seu espaço e pode causar doença nos humanos. E é precisamente por meio da doença que emerge o ritual do Hugagü.
Quando uma pessoa adoece mais seriamente e a cura se alonga, um coletivo de pajés é contratado para ir ‘buscar a alma’ (akuãtelü) do doente, que foi raptada por algum itseke. O diagnóstico possui várias etapas e não cabe aqui descrevê-lo, mas sói desconfiar-se do hiper-beija-flor durante a safra do pequi, uma vez que as pessoas estão constantemente expostas em seu domínio. Cada pequizal tem um dono humano, mas o pequi tem donos não humanos, e é preciso haver-se com eles. Após o diagnóstico e a cura, o doente poderá se tornar dono do ritual do Hugagü, no qual vários itseke são chamados a dançar e comer. A festa não tem data certa e nem a obrigação de ser realizada todos os anos, mas, quando realizada, ocorre sempre durante a safra de pequi. Um dono poderá reencenar o ritual durante vários anos até decidir abandoná-lo, por meio de uma festa de encerramento em que se produzem miniaturas realistas dos donos do pequi, feitas em madeira (e no caso do hiper-beija-flor, em cera). O casal de beija-flores é levado por dois cantores e é o primeiro a ter seu bico molhado no caldo de pimenta e goma de mandioca, com que se alimentam os espíritos: a eles, pede-se que sejam bons e que não causem enfermidades aos humanos.
Os cuidados a serem tomados durante a safra do pequi não dizem respeito apenas aos donos. Na verdade, a safra só começa e o fruto só pode ser consumido após rezar-se a primeira bebida de pequi (imbene), que é trazida para o centro da aldeia. Esta reza, convoca uma série de personagens: o jacaré, suas amantes, o marido traído, a cotia, a cigarra (filha das amantes e onipresente nos pequizais durante a safra), a concha com que se descasca o fruto, Taũgi (por ele ter ensinado como preservar a polpa), o vento e outras figuras míticas. A reza visa afastar doenças e tempestades, pois o início da safra no começo de outubro coincide com a chegada das primeiras chuvas, acompanhadas de raios e fortes ventos, após uma longa estiagem16 16 Para uma análise desse tipo de reza de “batismo”, que ocorre também no caso do milho e da rede de pesca, ver Fausto et al. (2011). . Essa associação entre a fenologia do pequi e o regime climático, bem como a marcação de relações ecológicas por meio de narrativas e rezas apontam para a impossibilidade de dissociar conhecimento natural e cultural no alto Xingu.
PRÁTICAS DE CULTIVO
Se os conhecimentos e práticas envolvidos no cultivo e nos tratos silviculturais do pequi são indissociáveis de seus ‘aspectos socioculturais’, a própria separação entre conhecimento e prática deve ser aqui matizada, pois até a narrativa mítica tem uma orientação, por assim dizer, prático-teórica. Rezas, narrativas e ritos condensam informações técnicas e ecológicas em fórmulas memorizáveis, que são transmitidas entre gerações, conformando um modelo geral que engloba procedimentos de plantio, manejo, consumo e armazenamento do pequi. Algumas dessas técnicas incidem direta ou indiretamente sobre a diversidade intraespecífica observada em pequizais cultivados nos arredores da aldeia de Ipatse.
Como vimos, os Kuikuro mencionam que o primeiro pequizeiro, brotado do umbigo do jacaré, possuía todas as variedades de pequi dispostas em uma mesma árvore. No processo de cultivo, os Kuikuro plantam todas as sementes juntas em uma cova em formato de jacaré, aberta na roça ou na aldeia, transferindo-as posteriormente para seus lugares definitivos entremeadas às covas de mandioca (o formato visa agradar aquele que está na origem da planta e, deste modo, garantir que as sementes germinarão com sucesso)17 17 Para obterem árvores mais produtivas, os Kuikuro utilizam uma madeira chamada kuõ, a fim de cavar o buraco em forma de jacaré. Kuõ é uma árvore que dá muitos frutos e serve, por este motivo, para estimular o pequizeiro a produzir muito também (não foi possível fazer a identificação botânica da espécie). Segundo Tsaná Kuikuro, um dos principais cantores de Ipatse, seu pai, ensinou-lhe cantos que também servem para o mesmo fim. . Depois de abrir o buraco, as sementes são aí colocadas com o ‘nariz’ para cima. Em seguida, cobre-se com terra e pronuncia-se a ‘reza do lagarto’, a qual diz que o pequi lhe será útil para furar a orelha de seu filho e que, portanto, ele não deve desenterrar as sementes18 18 Hagatu, hagatu imukugu ipogohoingo higei, “lagarto, lagarto, isto servirá para a furação de seu filho”. A furação de orelha é um ritual de passagem masculino associado à chefia. .
O plantio de várias sementes juntas na produção das mudas, bem como o transplante de uma quantidade grande de mudas para a roça servem como estratégia para minimizar o efeito de predadores e garantir que pelo menos parte das sementes e plântulas sobreviva e cresça. Conforme nos sugeriu Carlos Saito (comunicação pessoal, 2013), é possível que essa prática ajude também a quebrar a dormência das sementes, por formar um ambiente propício ao choque térmico no solo. Na medida em que a dormência das sementes de pequi é geralmente referida como um empecilho para o cultivo comercial desta árvore (Pereira et al., 2004PEREIRA, A. V.; PEREIRA, E. B. C.; SILVA, D. B.; GOMES, A. C.; SOUSA SILVA, J. C. Quebra da dormência de sementes de pequi. Boletim de Pesquisa e Desenvolvimento, v. 136, p. 1-15, 2004.), caso confirme-se a hipótese de Saito, temos aqui mais uma evidência de que características diferenciadas do pequi xinguano resultam de um processo de domesticação local.
Após um ou dois meses, as sementes germinam no local do buraco-jacaré e são transferidas para as roças de mandioca, sendo plantadas a uma distância regular (em torno de cinco a seis metros) umas das outras. O plantio das mudas ocorre, geralmente, no último ano produtivo da roça, logo após o replantio da maniva. Com isso, temos condições propícias para o crescimento da plântula de pequi, a qual requer áreas abertas com muita luminosidade. Nos primeiros anos de crescimento das mudas, os donos do pequizal precisam investir no trabalho de limpeza de ervas daninhas, evitando o sombreamento das plântulas, o que é feito em conjunto com os tratos silviculturais, necessários para manter o plantio de mandioca ainda em fase produtiva. A limpeza do solo ao redor das plântulas é importante também para evitar eventuais queimadas por fogo que escape de roças abertas nas proximidades do novo pequizal, já que os Kuikuro não têm costume de fazer aceiro. São colocadas estacas ao redor das mudas de pequi recém-plantadas, para evitar que sejam pisoteadas durante o trabalho na roça.
Passada a fase inicial em que as pequenas plantas de pequi são muito vulneráveis, a fase de consolidação dos pequizais ainda requer cuidados e manejo regulares, que são feitos pelas famílias, quando vão à roça. Quando a planta já apresenta uma lignificação do tronco, os Kuikuro costumam arranhar o caule do pequi com dente de jacaré para a árvore crescer bem. Assim como os jovens xinguanos, que durante o período de reclusão pubertária devem ter a pele escarificada regularmente para ficarem grandes e fortes, os jovens pequizeiros também devem ser arranhados, a fim de superar com sucesso a transição entre a ‘puberdade’ e a vida adulta (reprodutiva)19 19 A ‘arranhadeira’ (escarificador) utilizada no Xingu é feita de dentes de uma espécie menor de peixe-cachorro, incrustrados em um pedaço de cabaça. . As árvores começam a frutificar a partir de cinco a sete anos após o plantio, e os frutos são coletados pelas mulheres na época da safra.
A DIVERSIDADE DO PEQUI ALTOXINGUANO
A diversidade intraespecífica de pequis que nos interessa aqui é aquela mapeada a partir da perspectiva kuikuro; i.e., trata-se de compreender como os Kuikuro percebem, nomeiam e manejam elementos da diversidade agrícola em uma unidade de paisagem domesticada (o pequizal), relacionando-a com aspectos da socialidade atribuída à espécie e ao agroecossistema no qual é cultivada.
Todas as variedades de pequi, plantadas ou silvestres, são denominadas imbé pelos Kuikuro. Dentro desta categoria geral, eles fazem uma primeira divisão em dois grandes grupos: o dos ‘pequis verdadeiros’ (imbé hekugu)20 20 Hekugu, assim como o sufixo eté tupi-guarani, é tanto um intensificador como um indicador “de pertencimento perfeito a uma classe ou – para usar a terminologia da teoria dos protótipos – maior representatividade como membro de uma categoria; i.e., maior grau de prototipicalidade” (Fausto, 2001, p. 262). , composto por pequizeiros cultivados, e o dos pequis silvestres, denominados kapula21 21 Kapula designa o conjunto das populações silvestres que ocorrem na unidade de paisagem denominada tahutahupe pelos Kuikuro, que corresponde ao cerrado típico. Em outro estudo em andamento, avaliamos que ambos os grupos correspondem a variantes internas dentro da espécie Caryocar brasiliense. O termo kapula refere-se especificamente às populações silvestres de C. brasiliense. , os quais ocorrem naturalmente em manchas de cerrado, entremeadas a outras espécies desta fitofisionomia. Quando utilizamos aqui o termo ‘pequi xinguano’ (ou altoxinguano), nos referimos exclusivamente ao grupo composto pelos morfotipos ou variedades de ‘pequis verdadeiros’ reconhecidos pelos Kuikuro. Além da distinção entre plantado e não plantado, entre floresta e cerrado, os Kuikuro distinguem os dois grupos em função de algumas características fenotípicas (descritores) dos frutos (Tabela 1).
Os Kuikuro não atribuem nomes varietais para agrupar subcategorias que sirvam para ordenar a variação fenotípica entre indivíduos dentro da categoria kapula, embora reconheçam a existência desta variação. Em contraste, dentro da categoria imbe hekugu, dispõem de 16 termos para designar variedades ou morfotipos de pequis cultivados (Tabela 2)22 22 Mais precisamente, são 15, pois uma delas, eungatagü ("o que foi plantado pelo besouro"), designa qualquer árvore na proximidade de um pequizal que se encontra fora da área plantada. Os Kuikuro não cortam esse pequizeiro e reconhecem os frutos como pertencendo a uma das outras categorias. .
É preciso notar, contudo, que os Kuikuro nomeiam antes características ou descritores do que propriamente variedades, ao contrário do que parece ocorrer no caso da mandioca, entre os próprios Kuikuro e entre outros grupos indígenas da Amazônia. As variedades (ou morfotipos) são nomeadas a partir da característica considerada mais proeminente do fruto e, por esse motivo, foram incluídas algumas feições que não possuíam regularidade (nem todos os frutos de uma mesma árvore apresentavam aquela característica). Algumas variações ocasionadas por fatores ambientais, como a predação de frutos antes de caírem ou indivíduos plantados por dispersores não humanos, entre outras, também foram nomeadas como ‘variedades’ ou, ainda, como ‘variantes’.
Como sói acontecer em contextos interculturais, nos quais nenhuma das partes domina perfeitamente a língua do outro, encontrar um equivalente para a noção botânica de ‘variedade’ não é jamais evidente23 23 Mesmo quando alguns interlocutores indígenas falam um bom português e um dos pesquisadores possui um domínio razoável da língua indígena, como é o nosso caso. . Neste caso, porém, parece não haver tal categoria equivalente. A noção etnobotânica de variedade, geralmente utilizada para descrever a variação intraespecífica da mandioca (Boster, 1984BOSTER, J. S. Classification, cultivation, and selection of Aguaruna cultivars of Manihot esculenta (Euphorbiaceae). Advances in Economic Botany, v. 1, p. 34-47, 1984., 1985BOSTER, J. S. Selection for perceptual distinctiveness: evidence from Aguaruna cultivars of Manihot esculenta. Economic Botany, v. 39, n. 3, p. 310-325, 1985.; Elias et al., 2000ELIAS, M.; RIVAL, L.; MCKEY, D. Perception and management of cassava (Manihot esculenta Crantz) diversity among Makushi Amerindians of Guyana (South America). Journal of Ethnobiology, v. 20, n. 2, p. 239-265, 2000.), não se mostrou de fácil adequação às formas de percepção e manejo da diversidade de pequis cultivados pelos Kuikuro. Essa diferença talvez decorra de dessemelhanças biológicas entre as espécies, pois o pequi é uma espécie perene e alógama, ou seja, que se reproduz preferencialmente de forma sexuada (fecundação cruzada) e possui alguns mecanismos de autoincompatibilidade, embora possa ocorrer autofecundação (Gribel e Hay, 1993GRIBEL, R.; HAY, J. D. Pollination ecology of Caryocar brasiliense(Caryocaraceae) in Central Brazil cerrado vegetation. Journal of Tropical Ecology, v. 9, n. 2, p. 199-211, 1993.). O mecanismo reprodutivo do pequi é diversificador e inovador, produzindo alta variação genotípica e fenotípica, e dificultando a ação dos agricultores em manter variedades estáveis. Já a mandioca possui uma combinação de dois mecanismos reprodutivos: a) um mecanismo diversificador pela produção de sementes e b) um mecanismo conservador pela reprodução vegetativa a partir do brotamento de manivas (Kerr, 1986KERR, W. Agricultura e seleções genéticas de plantas. In: RIBEIRO, B. (Ed.). Suma etnológica brasileira: etnobiologia. Petrópolis: Vozes, 1986. v. 1, p. 159-172.; Martins, 2005MARTINS, P. S. Dinâmica evolutiva em roças de caboclos amazônicos. Estudos Avançados, v. 19, n. 53, p. 209-220, 2005.). Esta combinação permite o surgimento de novos fenótipos a partir de sementes e sua incorporação na coleção de variedades cultivadas e, ao mesmo tempo, a manutenção de variedades específicas pela propagação vegetativa, o que propicia uma coleção diversificada. Observamos que as formas de classificação e ordenamento da agrobiodiversidade variam não somente entre sistemas agrícolas diversos, mas dentro do sistema agrícola de um mesmo grupo étnico, de acordo com os diferentes objetos biológicos nele englobados. Este parece ser o caso dos Kuikuro, no contraste entre a mandioca e o pequi, embora, na prática, seja uma situação muito mais complexa, como veremos na próxima seção24 24 Conforme pesquisa realizada por H. Cooper e C. Fausto, em 2014, os Kuikuro utilizam-se conscientemente dos dois mecanismos reprodutivos da mandioca. Assim, ao encontrarem um novo pé gerado por reprodução sexuada em suas roças ou em roças abandonadas, experimentam a variedade plantando as manivas. Se as novas plantas gerarem tubérculos do agrado das mulheres, repetem a ação, ampliando o número de pés até um ponto em que as manivas começam a circular em uma rede social de vizinhança e parentesco. Uma das variedades, hoje dominante nas roças kuikuro, resultou desse processo de experimentação e clonagem. .
Seja como for, as categorias kuikuro para designar diferenças internas ao pequi cultivado parecem apontar antes para um ‘campo de variações perceptíveis’ do que para variedades estáveis e consolidadas – assim, antes do que variedades, temos um conjunto de variações reconhecidas por um nome, o qual poderíamos chamar ‘variantes’, a fim de distinguir de nossa noção de variedade intraespecífica (seja ela agronômica, taxonômica ou etnobotânica). É por meio da percepção e avaliação da variação fenotípica que ocorre o processo de escolha e seleção de sementes, afetando as frequências gênicas e genotípicas das populações cultivadas de pequi, ou seja, o processo local de domesticação da planta. Isso implica, ao selecionarem sementes para o plantio, o fato de os Kuikuro valorizarem diferentes combinações de descritores, e não unicamente a característica mais proeminente. Considerando as possibilidades de combinação entre as diferentes características de frutos de pequi utilizadas como descritores, a diversidade fenotípica percebida e manejada pelos Kuikuro é maior do que a nomeada por meio de subcategorias. Seria, pois, essa variação morfológica que, em última instância, é considerada na seleção de sementes para a reprodução e não a sua subsunção a uma ou outra categoria nominal.
O pequi é utilizado para o consumo de sua polpa cozida, para a produção de óleo (da polpa crua), para a produção de uma ‘geleia’ adocicada (denominada tuma), obtida a partir da apuração do caldo, e para o aproveitamento da castanha seca25 25 A castanha seca é importante no ritual do Quarup, que ocorre cerca de seis meses após o término da safra de pequi. Ao final da festa, as meninas que deixam a reclusão são levadas pelos donos da festa até os chefes das aldeias convidadas, oferecendo-lhe castanhas de pequi. . Alguns tipos de pequi são preferencialmente utilizados para finalidades específicas: assim, o imbese é adotado na alimentação, mas, por ter coloração clara, é antes valorizado para a extração de óleo; já o kanga mukugu e o tunguĩ são considerados especialmente bons para alimentação e armazenamento de sua polpa após o cozimento. No caso do tunguĩ, a ausência de espinhos não faz tanta diferença do ponto de vista nativo, já que os Kuikuro não comem com tanta frequência o fruto inteiro in natura, como ocorre na cozinha regional26 26 Além da preferência alimentar pelo tunguĩ, os Kuikuro separam suas sementes para fazer os chocalhos de fieira, que são usados nos rituais, amarrados ao tornozelo. Isso porque é mais fácil limpá-lo, dada a ausência de espinhos. Assim, é comum ver sementes de tunguĩ separadas das demais durante a safra do pequi. . Eles sabem do valor atribuído ao tunguĩ pelos kagaiha (não indígenas) e vendem-nos esporadicamente a seus ‘amigos’ na cidade de Canarana ou Gaúcha do Norte. Diferentemente dos regionais, contudo, os Kuikuro interessam-se antes pela diversidade de pequis cultivados. Ainda que tenham preferências positivas, como é o caso do pequi carnudo (suümkogu), e negativas, como é o caso de atü, cujo gosto é amargo e ‘travoso’, continuam a plantar pequizais com uma boa diversidade fenotípica27 27 Embora não o plantem, no caso do kapula, podem utilizá-lo eventualmente para a produção de óleo, mas jamais para consumo alimentar. .
DISTRIBUIÇÃO DA DIVERSIDADE FENOTÍPICA DE PEQUIS NAS PARCELAS AMOSTRADAS
O número de árvores de pequi em cada pequizal é bastante variável, podendo chegar a 60, embora a média esteja na casa dos 30 indivíduos. Em nossa pesquisa, a grande maioria dos indivíduos foi classificada na categoria imbé hekugu pelos seus respectivos donos (Tabela 3). Esse dado, no entanto, deve ser tomado cum grano salis, pois se trata de uma categoria dada de antemão (utilizada se não acionada alguma escolha a mais), ou seja, uma categoria de fundo que se aplica a todos os pequizeiros plantados, sendo que a distinção em subcategorias aplica-se de maneira subsequente. O mesmo ocorre, por exemplo, no domínio da música ritual, em que várias peças não recebem nome e são ditas apenas egi hekugu, ‘cantos verdadeiros’ ou, mais exatamente, ‘simplesmente cantos’.
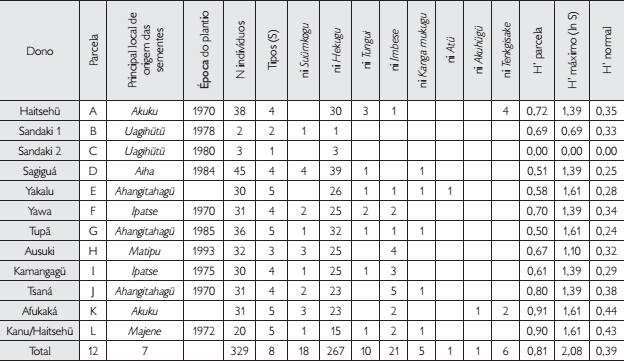
Composição de variantes de pequi indicada pelos agricultores Kuikuro nos seus respectivos pequizais amostrados (parcelas). Estimativa do índice de diversidade de Shannon (H’) modificada para cada parcela, calculada a partir da diversidade de morfotipos indicada pelo dono de cada pequizal. Legendas: N = abundância de indivíduos amostrados em cada parcela; ni = abundância de indivíduos do morfotipo i; S = número de morfotipos em cada parcela; H’ = - ∑ (ni/N)*ln(ni/N), onde ni/N = abundância relativa do morfotipo i; H’ máximo = ln S.
Embora os Kuikuro tenham listado 15 categorias de imbé hekugu em sentido amplo (Tabela 2 – excluindo-se eungatagü), durante as entrevistas nas parcelas, utilizaram apenas oito dessas categorias (Tabela 3). Observamos que, quando perguntados sobre as árvores e não sobre os frutos, os donos dos pomares tendem a categorizar seus pequizeiros a partir de uma noção de variedade semelhante àquela definida por Emperaire (2006)EMPERAIRE, L. Histórias de plantas, histórias de vida: uma abordagem integrada da diversidade agrícola tradicional na Amazônia. Revista da Sociedade Brasileira de Etnobiologia e Etnoecologia, v. 3, p. 189-198, 2006., em seus estudos sobre a mandioca, excluindo certos termos utilizados para descrever a diversidade de frutos28 28 Termos tais como tükitse hügipingo (‘com a flecha do grilo’), o qual designa um fruto machucado, ou tinaki, que descreve o fruto que, ao cair, o pedúnculo descola inteiramente do fruto. . Em outras palavras, os Kuikuro parecem operar com duas noções de diversidade ao mesmo tempo, daí decorrendo nossa dificuldade em traduzir o conceito de variedade.
A despeito dessas questões, procuramos calcular o índice de diversidade de Shannon (H’) modificado para cada pequizal plantado com base na abundância relativa de indivíduos (ni/N – Tabela 3) alocados nas diferentes variedades ou variantes de pequi indicadas pelos agricultores Kuikuro, para estimar quantitativamente a diversidade intraespecífica de pequis nas parcelas29 29 Apesar de terem identificado vários indivíduos como tsupügügi (pequenos) nas parcelas, os entrevistados mencionavam-no como hekugu tsupügügi e, por esse motivo, foram todos quantificados na categoria hekugu, tomada aqui em sentido estrito (i.e., como subcategoria do grupo de pequizeiros plantados). . O maior número de variedades (S) indicado em unidades de cultivo (pequizais) amostradas na Tabela 3 foi de cinco (parcelas E, G, K, L) e o menor número (S) foi de uma única variante ou variedade (imbé hekugu) na parcela C, a qual destoa da média dos pequizais (juntamente com a parcela B), por ter uma quantidade muito baixa de indivíduos (N). Trata-se dos dois pequizais plantados por Kamaluhé Sandaki Matipu, cujas sementes foram trazidas da antiga aldeia Uagihütü – um referencial territorial importante na história dos povos de língua karib no alto Xingu. Cumpre ressaltar que as informações sobre cada pequizal foram fornecidas pelo responsável pelo seu plantio. O índice ora mencionado foi utilizado com o intuito de associar a diversidade intraespecífica de cada pequizal à história de vida dos diferentes horticultores indígenas.
Como as unidades amostradas apresentaram uma grande variação entre as populações de dados, calculamos um valor de H’ máximo com base no número total de variantes citadas em todas as amostras (S = 8). O valor final de H’ para cada parcela (H’ normal) foi calculado como H’ parcela/H’ máximo de todas as parcelas. Com exceção das parcelas B e C, que possuem um tamanho amostral muito baixo, o índice de diversidade de Shannon modificado para analisar a diversidade morfológica de frutos de pequi (H’ normal) teve pouca variação (0,24 ≤ H’ ≤ 0,43) entre as parcelas amostradas. Embora as parcelas K (plantada por Afukaká Kuikuro) e L (plantada por Haitsehü Kuikuro) apresentem maior diversidade de variantes nomeadas pelos Kuikuro, todos os pequizais parecem ter um mesmo padrão de diversidade intraespecífica com forte dominância do tipo imbé hekugu (estrito senso). Esta dominância de um dos tipos reduz o parâmetro equitabilidade no cálculo do índice, promovendo um valor mediano (não muito alto ou muito baixo) de diversidade para as parcelas.
Como apontamos anteriormente, no entanto, a variação morfológica de pequis cultivados no entorno da aldeia de Ipatse está provavelmente subestimada quando apenas consideradas as categorias nomeadas, em particular pelo efeito do sistema de classificação e da categoria mais ampla imbé hekugu. Ademais, ela não reflete a real diversidade existente nas coleções de sementes selecionadas pelos Kuikuro em diferentes locais do alto Xingu, já que não são levadas em conta as combinações de características na classificação dos frutos, mas apenas a característica tida como principal por cada agricultor. Neste sentido, a ‘composição de variantes nomeadas’ não se mostrou um parâmetro adequado como indicador de diversidade nas áreas amostradas, embora forneça uma estimativa relevante para comparar as parcelas e identificar um padrão de diversidade de pequis na aldeia de Ipatse.
Neste trabalho, o índice foi utilizado por levar em consideração não somente a riqueza de variantes morfológicas (S) nomeadas pelos agricultores indígenas em cada pequizal, mas a equitabilidade na abundância dos indivíduos de cada variante, destacando os ‘tipos raros’ (aqueles com frequência relativa ≤ 20%) (Tabela 4). Chama atenção o fato de que as variedades selecionadas positivamente, ou seja, aquelas com fenótipos mais apreciados (suümkogu, imbese e tunguĩ), aparecem nas parcelas com uma frequência relativamente alta (F Rel ≥ 50% - Tabela 4), a despeito, uma vez mais, da dominância da categoria lato sensu imbé hekugu. Falta-nos ainda uma análise mais detalhada do sistema local de classificações em comparação com a efetiva diversidade genética dessas populações, para corroborar a hipótese de que o pequi xinguano está em processo avançado de domesticação local. Entretanto, a alta frequência relativa de tunguĩ em populações cultivadas fornece fortes indícios de domesticação, já que se trata de um genótipo recessivo que ocorre em baixíssima frequência na natureza30 30 De acordo com o pesquisador da EMBRAPA Cerrados, Nilton Junqueira, especialista em melhoramento genético de pequi, pequis sem espinho no caroço ocorrem em proporção de 1/1.000 indivíduos (comunicação pessoal, 2013) .
Frequência relativa das variantes morfológicas indicadas nos 12 pequizais amostrados no entorno da aldeia Ipatse. Legenda: F Rel = frequência relativa.
REDE DE CIRCULAÇÃO DE SEMENTES
O plantio de novos pomares de pequi está associado a um período inicial de ocupação de assentamentos permanentes ou temporários, uma vez que se trata de uma espécie perene, cujas unidades de cultivo (pequizais) são repassadas às novas gerações. Desse modo, poucas pessoas da aldeia de Ipatse tiveram a experiência de plantar pequi durante sua história de vida. No censo realizado em junho de 2011, de um total de 308 pessoas, apenas 23 disseram ter sido responsáveis pelo plantio de pequi naquela localidade. Esse número, porém, é mais expressivo do que parece à primeira vista, uma vez que o plantio é feito para as gerações futuras: pais e avôs de sexo masculino plantam para seus filhos e netos. Entre os 37 homens com mais de 40 anos vivendo naquele momento na aldeia, mais de 60% já tinham plantado pequizais para seus descendentes.
O fluxo de sementes de pequi no alto Xingu ocorre por meio de redes sociais que atuam em diferentes níveis e escalas. Na escala temporal, já mencionamos que o plantio de novos pequizais está associado à transmissão intergeracional e aos períodos de ocupação de novos assentamentos. Isso ocorreu em um ciclo de médio prazo, em torno de 20 a 50 anos no último século, em função das realocações pós-contato, mas possivelmente era de mais longo prazo no passado. Como os pequizais se tornam marcas no território, podem ser reutilizados também como forma de reavivar um direito de ocupação. Há velhos pequizais, com pouquíssimos indivíduos sobreviventes, plantados entre o final do século XIX e o começo do século XX, cuja cadeia de transmissão geracional ainda é reconhecida pelos Kuikuro.
Na escala espacial, a rede de circulação de sementes atua em dois níveis principais: um nível interno entre diferentes pequizais de uma mesma aldeia e um nível regional, entre aldeias e povos do alto Xingu. Entre os 12 pequizais amostrados, cinco deles foram plantados com sementes coletadas preferencialmente na localidade do lago de Ipatse (arredores da aldeia atual e da aldeia precedente de Ahangitahagü para a qual se mudaram em 1961, face à exclusão inicial da aldeia de Lahatuá do perímetro do Parque Indígena do Xingu). Nos outros sete pequizais, foram utilizadas sementes trazidas de procedências diversas, mas sempre do território de uso dos povos de língua karib do alto Xingu (Figura 5). O fato de sete, dos 12 pequizais amostrados, possuírem sementes de outras aldeias parece indicar uma preferência dos Kuikuro por obter sementes de várias origens, inclusive fora de seus domínios31 31 Surpreendentemente, pelo menos nesta amostra, não há referência à região de Kuhikugu e Lahatuá, onde permaneceram de 1820 a 1961 (e que atualmente reocuparam). Apenas Haitsehü trouxe semente de um sítio de ocupação provisória nesta região, conhecido como Majene. .
Locais de origem das sementes de pequi plantadas nos doze pequizais amostrados na aldeia Ipatse. A espessura das setas indica quantos, entre os pequizais amostrados, receberam sementes de cada localidade: Naruvoto = 1; Akuku = 2; Uagihütü = 2; Sítio Majene = 1; Ahangitahagü = 4; Ipatse = 2; Matipu = 1; Aiha = 1. Setas vermelhas = circuito local; setas azuis = circuito regional. Note-se que, neste caso específico, as sementes circularam entre aldeias e assentamentos de povos de língua karib altoxinguanos, seja no nível local ou regional (circuito karib).
As principais fontes de sementes para o plantio dos pequizais estão associadas a espaços de referência na história de vida de cada um dos agricultores e/ou de suas famílias. Elementos importantes desta história referem-se aos casamentos, à origem dos pais e aos locais de moradia.
Se observarmos a Tabela 3, podemos rapidamente traçar estas relações no tempo e no espaço. Assim, por exemplo, Haitsehü é um homem de mais de 70 anos de origem kalapalo. As sementes do pequizal que plantou vieram da antiga aldeia de Akuku, onde viviam os Kalapalo na década de 40, no momento da chegada dos irmãos Villas-Boas. O cacique Afukaká também trouxe suas sementes desta mesma aldeia, abandonada desde 1961, pois se casou com duas irmãs, filhas de uma mulher kalapalo. Já Sagiguá, que é casado com uma outra mulher kalapalo e tem cerca de 50 anos, trouxe as sementes da aldeia de Aiha, a aldeia kalapalo que sucedeu a Akuku e onde a sua esposa nasceu. O pai de Ausuki foi um importante chefe matipu e, por isso, trouxe as sementes de lá. Essa cadeia poderia ser multiplicada, pois a circulação ocorre horizontalmente (via casamento) e verticalmente (como herança); ou seja, as sementes trazidas de outras aldeias e plantadas para as gerações futuras foram elas mesmas trazidas de outras aldeias e plantadas para gerações, hoje, passadas.
Um fato notável no caso kuikuro é que a rede horizontal possui uma amplitude restrita, uma vez que os casamentos internos são prevalentes. Se há certa frequência de casamentos interétnicos, a grande maioria ocorre dentro do bloco linguístico karib, com pessoas kalapalo, matipu e nafukwá. São pouco numerosos os casamentos com pessoas falantes de outras línguas do sistema xinguano, sendo as uniões fora da constelação xinguana ainda mais raras32 32 Em 2012, havia um casamento com uma mulher xavante-terena e com uma não indígena. Ambos os maridos não viviam nas aldeias kuikuro. . Assim, a circulação de sementes de pequi em nível regional, pelo menos no caso kuikuro, ocorre preferencialmente no interior do território do bloco linguístico. Isto não é tão marcado no caso de outros povos altoxinguanos e podemos supor que haja também um fluxo ‘interlinguístico’ importante de sementes na região, o que contribui para a diversidade fenotípica observada nos pequizais cultivados.
Os pequizais de Ipatse são resultantes do fluxo de sementes em diversos níveis de abrangência espacial, pois, em um único pequizal, há uma mistura de indivíduos provenientes de sementes coletadas em diferentes aldeias no mesmo período. Aqui as redes de circulação de sementes se configuram de maneira distinta daquela das redes de troca de germoplasma, tal qual descrito em outros estudos sobre sistemas agrícolas tradicionais (Emperaire et al., 2008EMPERAIRE, L.; VAN VELTHEN, L.; OLIVEIRA, A. G. Patrimônio cultural imaterial e sistema agrícola: o manejo da diversidadeagrícola no médio rio negro. In: REUNIÃO BRASILEIRA DE ANTROPOLOGIA, 26., 2008, Porto Seguro. Anais... Porto Seguro: ABA, 2008. p. 1-16.; Seixas, 2008SEIXAS, A. C. P. S. Entre terreiros e roçados: a construção da agrobiodiversidade por moradores do Rio Croa, Vale do Juruá (AC). 2008. 124 f. Dissertação (Mestrado em Desenvolvimento Sustentável: Política e Gestão Ambiental) – Universidade de Brasília, Brasília, 2008.; Machado, 2012MACHADO, J. S. Lugares de gente: mulheres, plantas e redes de troca no delta amazônico (Caviana, PA). 2012. Tese (Doutorado em Antropologia Social) – Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2012.). Nestes estudos, o germoplasma de diferentes espécies circula entre pessoas, caracterizando uma rede de doadores e receptores. Em uma análise sobre todas as espécies e variedades cultivadas no sistema agrícola de uma comunidade ribeirinha do Rio Croa, no Acre, Seixas (2008)SEIXAS, A. C. P. S. Entre terreiros e roçados: a construção da agrobiodiversidade por moradores do Rio Croa, Vale do Juruá (AC). 2008. 124 f. Dissertação (Mestrado em Desenvolvimento Sustentável: Política e Gestão Ambiental) – Universidade de Brasília, Brasília, 2008. observou diferentes amplitudes de redes de troca entre famílias ligadas por relações de parentesco e vizinhança, de comércio e até mesmo institucionais.
A floresta, neste caso, também é uma importante provedora de diversidade agrícola (Seixas, 2008SEIXAS, A. C. P. S. Entre terreiros e roçados: a construção da agrobiodiversidade por moradores do Rio Croa, Vale do Juruá (AC). 2008. 124 f. Dissertação (Mestrado em Desenvolvimento Sustentável: Política e Gestão Ambiental) – Universidade de Brasília, Brasília, 2008.).
Com relação ao pequi, a rede de circulação de sementes caracteriza-se menos pela relação direta entre doadores e receptores – embora as relações sociais de parentesco sejam fundamentais na operação do fluxo – e antes pela relação de pessoas com territórios de referência (Figura 5). No processo de fissão e formação de novas aldeias, bem como por meio do casamento entre povos, permanecem fortes laços das famílias com seus territórios de origem, seja pela existência de parentes nestes locais ou pelo referencial territorial em si mesmo. Lugares de moradia passada, sobretudo ali onde os pequizais ainda florescem, são índices duráveis das relações que os produziram enquanto espaço humano. Essas relações pretéritas estendem-se até o presente, gerando direitos de uso, que podem ser mobilizados pelas gerações contemporâneas.
No sistema agrícola kuikuro, a amplitude da rede social de circulação de sementes de pequi se diferencia da rede de circulação de germoplasma de mandioca. No primeiro caso, o fluxo em nível espacial regional é expressivo na composição de pequizais; já no segundo, o fluxo de estacas (manivas) de mandioca ocorre preferencialmente em nível local (entre parentes da mesma aldeia). Na escala temporal, essa diferença de amplitude é ainda mais significativa: a circulação das estacas de mandioca é feita em ciclo anual, enquanto a de sementes de pequi ocorre em ciclos mais longos (em torno de 20 a 50 anos). Esta constatação é relevante, pois aponta para diferentes lógicas de gestão da agrobiodiversidade no manejo de diferentes tipos de plantas dentro de um mesmo sistema agrícola.
O processo de coleta, seleção e circulação espacial de sementes de pequi no alto Xingu leva-os a misturar as sementes de frutos de diferentes procedências em um mesmo cesto, próprio para guardá-las. No procedimento de plantio, os Kuikuro produzem mudas, enterrando estas sementes todas juntas e depois transferindo aquelas que germinaram para seus locais permanentes na roça de mandioca. Da mesma forma que as roças de mandioca, que geralmente são abertas em áreas contíguas, separadas apenas por uma ou poucas fileiras de variedades marcadoras de limites, os pequizais plantados nas roças mantêm a mesma configuração. Deste modo, árvores de pequi nascidas a partir de sementes de diferentes procedências ficam próximas entre si, favorecendo o fluxo gênico e a fecundação cruzada. Todos estes elementos, desde a seleção de sementes em escalas diferenciadas (temporal e espacial) e níveis espaciais distintos, a circulação social destas sementes (que segue os padrões de circulação local e/ou dos agricultores e de suas famílias), o armazenamento e o plantio destas sementes misturadas e a proximidade entre os pequizais são mecanismos que favorecem a diversidade morfológica de pequis cultivados pelos índios do alto Xingu.
CONSIDERAÇÕES FINAIS
Neste artigo, procuramos traçar a rede de relações tramada a partir do pequi altoxinguano. Esta rede não é formada apenas por horticultures, suas famílias e um objeto biológico, o pequizeiro. Ela é bem mais extensa, coletando em sua malha pessoas e coisas, humanos e não humanos, e se estendendo no espaço e no tempo para além das fronteiras da família, da aldeia, do bloco linguístico e mesmo da humanidade. Para usar os termos de Latour (1991)LATOUR, B. Nous n’avons jamais été modernes: essai d’anthropologie symétrique. Paris: Editions La Découverte, 1991., o pequi xinguano é um híbrido de natureza e cultura e, neste sentido, poderíamos tratá-lo como uma planta biocultural. Não podemos afirmar, ainda, que se trata de uma variedade ‘domesticada’ na acepção mais comum do termo em biologia (i.e. uma população modificada geneticamente por meio da seleção humana ao longo de gerações, de modo a acentuar traços que beneficiem os seus seletores). Devemos, ainda, aguardar as análises genéticas em andamento para confirmá-lo.
Neste artigo, interessou-nos, sobretudo, pensar o pequi não como recurso biológico revestido de aspectos socioculturais, mas como objeto constituído por relações sociais – sem estas, a rigor, o pequi xinguano simplesmente não existe. Isto não quer dizer que a biologia não possa estudar os conhecimentos, práticas e técnicas de cultivo da espécie e manejo dos pequizais e o seu impacto sobre a diversidade intraespecífica. Foi o que fizemos aqui de maneira preliminar, mostrando como certas práticas – a seleção de sementes de procedências diversas, o seu plantio conjunto para a produção de mudas, a proximidade entre pequizais permitindo o fluxo gênico – incidem direta ou indiretamente na diversificação e na manutenção de diversidade morfológica nas populações de pequi cultivadas. Contudo, essas práticas, assim como as redes sociais de circulação de sementes, não são aspectos culturais justapostos às características reprodutivas da espécie, mas parte mesmo de sua reprodução enquanto ‘pequi xinguano’ ou, mais exatamente, daquilo que os Kuikuro denominam imbé hekugu.
Vimos, ainda, que, se a diversidade intraespecífica da mandioca e de outras espécies cultivadas costuma ser descrita e quantificada por meio da noção de variedade – tal como, por exemplo, aquela utilizada por Emperaire (2006)EMPERAIRE, L. Histórias de plantas, histórias de vida: uma abordagem integrada da diversidade agrícola tradicional na Amazônia. Revista da Sociedade Brasileira de Etnobiologia e Etnoecologia, v. 3, p. 189-198, 2006. –, esta se aplica mal para caracterizar a diversidade morfológica de uma espécie alógama e de ciclo de vida longo, como o pequi, fato que dificulta a manutenção de variantes estáveis ao longo de gerações. À parte as oito categorias que correspondem mais de perto a nossa noção de variedade, os Kuikuro reconhecem ainda grande variação morfológica em pequizais cultivados. De fato, todo o sistema de manejo da espécie, o qual se inicia pelo processo de seleção de sementes para o plantio, atua com base nessa variação, independentemente de sua classificação em categorias nominais bem definidas. Esses dados indicam que dentro de um mesmo sistema agrícola podem operar formas distintas de classificação e manejo da diversidade intraespecífica com respeito a diferentes espécies vegetais. Sugerimos, assim, que o conceito de variedade seja utilizado com cautela e adaptado aos contextos locais.
Nosso estudo vai ao encontro de outros trabalhos que ressaltam a importância das redes sociais de circulação de sementes e de conhecimentos na dinâmica de geração e conservação da diversidade varietal de espécies agrícolas nos sistemas amazônicos (Emperaire e Eloy, 2008EMPERAIRE, L.; ELOY, L. A cidade, um foco de diversidade agrícola no Rio Negro (Amazonas, Brasil)? Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, v. 3, n. 2, p. 195-211, 2008.; Seixas, 2008SEIXAS, A. C. P. S. Entre terreiros e roçados: a construção da agrobiodiversidade por moradores do Rio Croa, Vale do Juruá (AC). 2008. 124 f. Dissertação (Mestrado em Desenvolvimento Sustentável: Política e Gestão Ambiental) – Universidade de Brasília, Brasília, 2008.; Lima et al., 2012LIMA, D.; STEWARD, A.; RICHERS, B. T. Trocas, experimentações e preferências: um estudo sobre a dinâmica da diversidade da mandioca no médio Solimões, Amazonas. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, v. 7, n. 2, p. 371396, 2012.). Essas redes operam de formas e em amplitudes espaço-temporais diversas, entre e dentro de cada um desses sistemas. Um dos elementos instigantes da fruticultura do pequi é sua transmissão intergeracional, que lhe dota de uma dinâmica própria na construção de paisagens culturais, algo que deve ser aproximado comparativamente ao manejo de palmeiras na Amazônia ocidental, sobretudo da pupunha (Rival, 1998RIVAL, L. Domestication as a historical and symbolic process: wild gardens and cultivated forests in the Ecuadorian Amazon. In: BALÉE, W. (Org.). Advances in historical ecology. New York: Columbia University Press, 1998. p. 232-250., p. 239-241; Costa, 2007COSTA, L. As faces do jaguar. Parentesco, história e mitologia entre os Kanamari da Amazônia ocidental. 2007. Tese (Doutorado em Antropologia Social) – Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2007., p. 84, 160, 193; Clement et al., 2009bCLEMENT, C. R.; RIVAL, L.; COLE, D. M. Domestication of peach palm (Bactris gasipaes Kunth): the roles of human mobility and migration. In: ALEXIADES, M. N. (Ed.). Shifting spaces, changing times: mobility, migration and displacement in indigenous lowland South America, Oxford: Berghahn Books, 2009b. p. 117-140.). Como mostrou Rival e outros autores, os bosques de palmeiras, com seu lento crescimento e sua perenidade, costumam simbolizar a continuidade das gerações e a memória dos mortos. O cultivo de pomares de pequi em roças de mandioca entre os Kuikuro é mais uma evidência da relevância de espécies arbóreas nos sistemas agrícolas amazônicos, bem como do fato de que o seu cultivo não se restringe aos quintais agroflorestais em torno de casas ou aldeias, podendo constituir agroecossistemas diferenciados no escopo desses sistemas.
A compreensão da impossibilidade de dissociar ‘aspectos biológicos’ e ‘socioculturais’ na dinâmica de produção e conservação da agrobiodiversidade, seja em nível intraespecífico, interespecífico ou ecossistêmico, reforça a necessidade de investir-se em estratégias de conservação on-farm, em consonância com as formas locais de cultivo e de transmissão do conhecimento. É central aqui a percepção de que estamos diante de uma tradição, cujos mecanismos pragmáticos de transmissão e memorização devem ser levados em conta nas estratégias de conservação e salvaguarda. Este é, evidentemente, um problema que só se coloca quando a cadeia de transmissão de um determinado conhecimento esteja em vias de se romper ou quando ele entre em conflito com outras formas de conhecimento e de apropriação, como ocorre hoje com o pequi sem espinhos xinguano. É a possibilidade de ele se tornar um bem com valor monetário, e não mais uma variedade inserida em um sistema agrícola indígena, que torna necessário pensar em reconhecimento de direitos e em ações de conservação e salvaguarda.
Este estudo busca, por fim, ter relevância para as políticas públicas, uma vez que o atual marco legal33 33 Mesmo o novo marco legal brasileiro, instituído por meio da Lei nº 13.123, de maio de 2015, a despeito das mudanças em relação ao marco anterior, mantém uma visão fragmentada da biodiversidade, separando o patrimônio genético dos conhecimentos tradicionais associados. , que regulamenta o acesso ao patrimônio genético e/ou aos conhecimentos tradicionais associados para fins de utilização comercial e repartição de benefícios (sistema ABS34 34 A sigla ABS é resultado da abreviação do termo em inglês access and benefit sharing, utilizado no escopo da Convenção sobre Diversidade Biológica (CDB) em nível internacional. ) no Brasil, baseia-se em uma visão fragmentada das diversas dimensões que compõem a diversidade biológica, sobretudo no que concerne à agrobiodiversidade. Esta visão fragmentada tem prejudicado os povos indígenas e os povos e comunidades locais na luta por seus direitos, enquanto principais detentores de conhecimentos tradicionais associados no sistema ABS.
AGRADECIMENTOS
Esta pesquisa foi desenvolvida no âmbito do projeto de doutorado “Árvores de cultura: cultivo e uso do pequi (Caryocar sp., Caryocaraceae) entre os Kuikuro do alto Xingu, MT” desenvolvido por Maira Smith, no Centro de Desenvolvimento Sustentável da Universidade de Brasília (CDS/UnB) entre 2009 e 2013, sob orientação de Laure Emperaire (Institut de Recherche pour le Développement – IRD) e co-orientação de Carlos Fausto (Museu Nacional do Rio de Janeiro). A autorização de acesso ao conhecimento tradicional associado para fins de pesquisa científica foi concedida pelo Instituto do Patrimônio Histórico e Artístico Nacional/Conselho de Gestão do Patrimônio Genético (IPHAN/CGEN) e publicada no Diário Oficial da União em março de 2012. Smith agradece os apoios concedidos pelo Programa de Documentação de Culturas Indígenas (PRODOCULT) - Museu do Índio/FUNAI; pelo Programa Universidades e Comunidades no Cerrado (UNICOM/Florelos/ISPN/União Europeia); pela Unidade Mista de Pesquisa 208 do Institut de Recherche pour le Développement/Museum National d’Histoire Naturelle (IRD/MNHN); pelo Programa de Cooperação Bilateral Conselho Nacional de Desenvolvimento Científico e Tecnológico-Institut de Recherche pour le Développement (CNPq-IRD), “Populações locais, agrobiodiversidade e conhecimentos tradicionais associados” (PACTA); e pela Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), na forma de bolsa de doutorado. Fausto agradece os recursos de pesquisa concedidos pelo CNPq e pela Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), bem como as bolsas de Iniciação Científica e mestrado concedidas pelo CNPq e pela CAPES.
Agradecemos também aos pesquisadores que fizeram parte do Conselho Científico do projeto: Vânia Rennó de Azevedo (Centro Nacional de Pesquisa de Recursos Genéticos e Biotecnologia – CENARGEN), Charles Clement (Instituto Nacional de Pesquisas da Amazônia – INPA), Fabio Freitas (CENARGEN), Frédéric Mertens (CDS/UnB) e Carlos Saito (CDS/UnB), e aos Kuikuro que participaram diretamente da coleta de dados: Sepé Ragati Kuikuro, Aruiá Kuikuro, Amuneri Kuikuro, Kamasinuá Kuikuro, Kanuta Kuikuro, Rikakumã Kuikuro, Ossó Kuikuro e Mayaya Kuikuro. As etapas de campo do projeto do Museu do Índio contaram com a presença de Hélio Sá Cabral Neto e de Helena Pagliaro Cooper, respectivamente alunos de mestrado e iniciação científica de Carlos Fausto. Eles participaram ativamente nas atividades de coleta de dados, auxiliando Maira Smith. Por fim, agradecemos os iluminadores comentários dos pareceristas e ao apoio de Robert Miller na revisão do abstract.
-
1
Segundo a Organização das Nações Unidas para Agricultura e Alimentação (FAO), a agrobiodiversidade é resultante de processos evolutivos, como a seleção natural e a seleção artificial por agricultores, durante milênios, e engloba todos os recursos e espaços envolvidos na produção agrícola pelas sociedades humanas (FAO, 1999FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS (FAO). Background Papers 1: agricultural biodiversity. In: FAO/NL CONFERENCE ON THE MULTIFUNCTIONAL CHARACTER OF AGRICULTURE AND LAND, 1., Maastricht, 1999. Background Papers... Maastricht: FAO, 1999. Disponível em: <http://www.fao.org/mfcal/pdf/bp_1_agb.pdf>. Acesso em: 10 maio 2014.
http://www.fao.org/mfcal/pdf/bp_1_agb.pd... ). Neste trabalho, estamos restringindo o termo agrobiodiversidade à diversidade específica e intraespecífica dos recursos genéticos vegetais e aos agroecossistemas nos quais eles se reproduzem. -
2
Segundo Thrupp (2000)THRUPP, L. A. Linking agricultural biodiversity and food security: the valuable role of agrobiodiversity for sustainable agriculture. International Affairs, v. 76, n. 2, p. 265-281, 2000., mais de 20% dos recursos alimentares mundiais são provenientes de sistemas agrícolas tradicionais.
-
3
Estamos chamando de ‘conhecimentos tradicionais intrínsecos’ os aspectos socioculturais que se referem não somente ao conhecimento, mas também ao trabalho indígena empregado na seleção de sementes, nas técnicas e práticas de produção de mudas, no transplante das mudas e nos tratos silviculturais envolvidos no processo de domesticação ou melhoramento genético tradicional do pequizeiro.
-
4
Os campos de murundus ocorrem geralmente no cerrado e são constituídos por áreas planas, que sofrem inundação no período de chuvas, e por morrotes (murundus), que não inundam. Os morrotes menores são cobertos por vegetação campestre e os maiores, por vegetação lenhosa do cerrado (Resende et al., 2004RESENDE, I. L. M.; ARAÚJO, G. M.; ASSIS-OLIVEIRA, A. P. A comunidade vegetal e as características abióticas de um campo de murundu em Uberlândia, MG. Acta Botanica Brasilica, v. 18, n. 1, p. 9-17, 2004.).
-
5
Carneiro (1957CARNEIRO, R. L. Subsistence and social structure: an ecological study of the Kuikuro Indians. 1957. Tese (Doutorado em Antropologia) – University of Michigan, Michigan, 1957., 1983)CARNEIRO, R. L. The cultivation of manioc among the Kuikuro of the upper Xingu. In: HAMES, R. B.; VICKERS, W. T. (Eds.). Adaptive responses in native Amazonians. New York: Academic Press, 1983. p. 65-111. levantou 46 variedades cultivadas e nomeadas; Fausto levantou 35 variedades, em trabalho preliminar realizado em 2002 com Morgan Schmidt. Em 2014, um levantamento mais cuidadoso, realizado com Helena Cooper, permitiu corrigir as homonímias na lista original de Carneiro, confirmando o número de variedades reconhecidas a 36. Mais da metade, contudo, não está mais sendo plantada pelos Kuikuro, tornando urgente um projeto de salvaguarda.
-
6
As entrevistas realizadas por Maira Smith foram mediadas pelo professor indígena Sepé Ragati Kuikuro, que ajudou no planejamento das atividades, na elaboração de questões e na tradução.
-
7
A Associação Indígena Kuikuro do alto Xingu (AIKAX) mantém um termo de cooperação com o Museu do Índio (Fundação Nacional do Índio – FUNAI) para acomodar o acervo resultante de atividades de pesquisa e documentação cultural produzidas na aldeia.
-
8
Inicialmente, as mudas de pequi são plantadas na roça consorciadas com as plantas de mandioca. Quando a roça de mandioca deixa de ser produtiva, os pequizeiros em estágio juvenil crescem, formando um pomar, os quais são manejados pelos Kuikuro de modo a desfavorecer a regeneração florestal espontânea nestas áreas.
-
9
O termo geral utilizado para a roça de mandioca é tuhinhaho, mas existem termos mais específicos usados para as diferentes fases da roça, desde o momento de sua abertura, passando pelo plantio, podas sucessivas, replantio até o abandono para pousio.
-
10
Baseada principalmente nas versões contadas por Haitsehü Kalapalo, em 2011, e Tugupé Kuikuro, em 2002, durante as filmagens de “Imbé Gikegü: cheiro de pequi”.
-
11
Dos testículos, nasce a mangaba, que embora não seja plantada, encontra-se no entorno das aldeias e é importante fonte alimentar em novembro, auge da colheita do pequi. Do pênis, nasce um fruto silvestre localmente denominado akaga e dos olhos nascem ugukumi. A associação entre o pequi e esses frutos deve-se à época de frutificação.
-
12
Não cabe aqui desenvolver este aspecto, mas ele é central na construção do filme “Cheiro de pequi” – cheiro este que é o da vagina, a qual é modelada em cera e mostrada às mulheres durante o ritual do Hugagü. Sobre o mesmo ritual entre os povos arawak do Xingu, ver Coelho (1992)COELHO, V. P. A festa do pequi e o zunidor entre os índios Waurá. Bulletin de la Société Suisse des Américanistes, n. 55-56, p. 37-56, 1992. e Gregor (1985)GREGOR, T. Anxious pleasures: the sexual lives of an Amazonian people. Chicago/London: The University of Chicago Press, 1985..
-
13
Esse é outro fato notável e singular da tecnologia xinguana associada à fruticultura do pequi. Eles constroem um cesto onde colocam a polpa, o qual é hermeticamente fechado e depositado em uma lagoa. A bebida feita a partir do pequi, conhecida como imbene, pode ser tomada durante o ritual do Quarup, na estação seca, cerca de seis meses após o final da safra.
-
14
Os Kuikuro costumam traduzir itseke por “bicho”, pois os itseke que povoam o mundo xinguano são, em sua maioria, animais em sua condição de pessoa; i.e., animais dotados de intencionalidade, reflexividade e capacidade comunicativa (ver Fausto, 2012bFAUSTO, C. Sangue de lua: reflexões ameríndias sobre espíritos e eclipses. Journal de la Société des Américanistes, v. 98, n. 1, p 63-80, 2012b.). Para a tradução do sufixo –kuegü como “hiper”, ver Franchetto (2003)FRANCHETTO, B. L’autre du même: parallélisme et grammaire dans l’art verbal des récits Kuikuro (caribe du Haut Xingu, Brésil). Ameríndia, n. 28, p. 213-248, 2003..
-
15
Embora os morcegos sejam os principais polinizadores de C. brasiliense no período noturno, algumas espécies de beija-flor também visitam as flores desta árvore nas primeiras horas do dia para sugar néctar, agindo como polinizadores cruzados e favorecendo a diversidade genética nas populações cultivadas (Gribel e Hay, 1993GRIBEL, R.; HAY, J. D. Pollination ecology of Caryocar brasiliense(Caryocaraceae) in Central Brazil cerrado vegetation. Journal of Tropical Ecology, v. 9, n. 2, p. 199-211, 1993.). Por seu turno, a arara e a cotia possuem papel importante como dispersores naturais das sementes de pequi.
-
16
Para uma análise desse tipo de reza de “batismo”, que ocorre também no caso do milho e da rede de pesca, ver Fausto et al. (2011)FAUSTO, C. Mil años de transformación: la cultura de la tradición entre los del alto Xingú. In: CHAUMEIL, J. P.; ESPINOSA, O.; CORNEJO, M. (Orgs.). Por donde hay soplo: estudios amazónicos en los países andinos. Lima: IFEA-CAAP-PUCP, 2011. (Actes & Mémoires, 29). p. 185-216..
-
17
Para obterem árvores mais produtivas, os Kuikuro utilizam uma madeira chamada kuõ, a fim de cavar o buraco em forma de jacaré. Kuõ é uma árvore que dá muitos frutos e serve, por este motivo, para estimular o pequizeiro a produzir muito também (não foi possível fazer a identificação botânica da espécie). Segundo Tsaná Kuikuro, um dos principais cantores de Ipatse, seu pai, ensinou-lhe cantos que também servem para o mesmo fim.
-
18
Hagatu, hagatu imukugu ipogohoingo higei, “lagarto, lagarto, isto servirá para a furação de seu filho”. A furação de orelha é um ritual de passagem masculino associado à chefia.
-
19
A ‘arranhadeira’ (escarificador) utilizada no Xingu é feita de dentes de uma espécie menor de peixe-cachorro, incrustrados em um pedaço de cabaça.
-
20
Hekugu, assim como o sufixo eté tupi-guarani, é tanto um intensificador como um indicador “de pertencimento perfeito a uma classe ou – para usar a terminologia da teoria dos protótipos – maior representatividade como membro de uma categoria; i.e., maior grau de prototipicalidade” (Fausto, 2001FAUSTO, C. Inimigos fiéis: história, guerra e xamanismo na Amazônia. São Paulo: EDUSP, 2001., p. 262).
-
21
Kapula designa o conjunto das populações silvestres que ocorrem na unidade de paisagem denominada tahutahupe pelos Kuikuro, que corresponde ao cerrado típico. Em outro estudo em andamento, avaliamos que ambos os grupos correspondem a variantes internas dentro da espécie Caryocar brasiliense. O termo kapula refere-se especificamente às populações silvestres de C. brasiliense.
-
22
Mais precisamente, são 15, pois uma delas, eungatagü ("o que foi plantado pelo besouro"), designa qualquer árvore na proximidade de um pequizal que se encontra fora da área plantada. Os Kuikuro não cortam esse pequizeiro e reconhecem os frutos como pertencendo a uma das outras categorias.
-
23
Mesmo quando alguns interlocutores indígenas falam um bom português e um dos pesquisadores possui um domínio razoável da língua indígena, como é o nosso caso.
-
24
Conforme pesquisa realizada por H. Cooper e C. Fausto, em 2014, os Kuikuro utilizam-se conscientemente dos dois mecanismos reprodutivos da mandioca. Assim, ao encontrarem um novo pé gerado por reprodução sexuada em suas roças ou em roças abandonadas, experimentam a variedade plantando as manivas. Se as novas plantas gerarem tubérculos do agrado das mulheres, repetem a ação, ampliando o número de pés até um ponto em que as manivas começam a circular em uma rede social de vizinhança e parentesco. Uma das variedades, hoje dominante nas roças kuikuro, resultou desse processo de experimentação e clonagem.
-
25
A castanha seca é importante no ritual do Quarup, que ocorre cerca de seis meses após o término da safra de pequi. Ao final da festa, as meninas que deixam a reclusão são levadas pelos donos da festa até os chefes das aldeias convidadas, oferecendo-lhe castanhas de pequi.
-
26
Além da preferência alimentar pelo tunguĩ, os Kuikuro separam suas sementes para fazer os chocalhos de fieira, que são usados nos rituais, amarrados ao tornozelo. Isso porque é mais fácil limpá-lo, dada a ausência de espinhos. Assim, é comum ver sementes de tunguĩ separadas das demais durante a safra do pequi.
-
27
Embora não o plantem, no caso do kapula, podem utilizá-lo eventualmente para a produção de óleo, mas jamais para consumo alimentar.
-
28
Termos tais como tükitse hügipingo (‘com a flecha do grilo’), o qual designa um fruto machucado, ou tinaki, que descreve o fruto que, ao cair, o pedúnculo descola inteiramente do fruto.
-
29
Apesar de terem identificado vários indivíduos como tsupügügi (pequenos) nas parcelas, os entrevistados mencionavam-no como hekugu tsupügügi e, por esse motivo, foram todos quantificados na categoria hekugu, tomada aqui em sentido estrito (i.e., como subcategoria do grupo de pequizeiros plantados).
-
30
De acordo com o pesquisador da EMBRAPA Cerrados, Nilton Junqueira, especialista em melhoramento genético de pequi, pequis sem espinho no caroço ocorrem em proporção de 1/1.000 indivíduos (comunicação pessoal, 2013)
-
31
Surpreendentemente, pelo menos nesta amostra, não há referência à região de Kuhikugu e Lahatuá, onde permaneceram de 1820 a 1961 (e que atualmente reocuparam). Apenas Haitsehü trouxe semente de um sítio de ocupação provisória nesta região, conhecido como Majene.
-
32
Em 2012, havia um casamento com uma mulher xavante-terena e com uma não indígena. Ambos os maridos não viviam nas aldeias kuikuro.
-
33
Mesmo o novo marco legal brasileiro, instituído por meio da Lei nº 13.123, de maio de 2015, a despeito das mudanças em relação ao marco anterior, mantém uma visão fragmentada da biodiversidade, separando o patrimônio genético dos conhecimentos tradicionais associados.
-
34
A sigla ABS é resultado da abreviação do termo em inglês access and benefit sharing, utilizado no escopo da Convenção sobre Diversidade Biológica (CDB) em nível internacional.
REFERÊNCIAS
- ANDRELLO, G. Cidade do índio: transformações e cotidiano em Iauaretê. São Paulo: Editora UNESP, 2006.
- BALÉE, W. Biodiversidade e os índios amazônicos. In: CARNEIRO DA CUNHA, M.; VIVEIROS DE CASTRO, E. (Orgs.). Amazônia: etnologia e história indígena. São Paulo, NHII/USP, 1993. p. 383-393.
- BALÉE, W. The culture of Amazonian forests. In: POSEY, D. A.; BALÉE, W. (Eds.). Resource management in Amazonia: indigenous and folk strategies. New York: New York Botanical Garden, 1989. (Advances in Economic Botany, v. 7). p. 1-21.
- BELLON M.R.; PHAM J. L.; JACKSON, M. T. Genetic conservation: a role for rice farmers. In: MAXTED N.; FORD-LLOYD B. V.; HAWKES J. G. Plant genetic conservation: the in situ approach. London: Chapman and Hall, 1997. p. 263-289.
- BOSTER, J. S. Selection for perceptual distinctiveness: evidence from Aguaruna cultivars of Manihot esculenta Economic Botany, v. 39, n. 3, p. 310-325, 1985.
- BOSTER, J. S. Classification, cultivation, and selection of Aguaruna cultivars of Manihot esculenta (Euphorbiaceae). Advances in Economic Botany, v. 1, p. 34-47, 1984.
- CARNEIRO, R. L. The cultivation of manioc among the Kuikuro of the upper Xingu. In: HAMES, R. B.; VICKERS, W. T. (Eds.). Adaptive responses in native Amazonians New York: Academic Press, 1983. p. 65-111.
- CARNEIRO, R. L. Subsistence and social structure: an ecological study of the Kuikuro Indians. 1957. Tese (Doutorado em Antropologia) – University of Michigan, Michigan, 1957.
- CLEMENT, C. R. 1492 and the loss of Amazonian crop genetic resources. I. The relation between domestication and human population decline. Economic Botany, v. 53, p. 188-202, 1999.
- CLEMENT, C. R.; CRISTO-ARAÚJO, M.; D’EECKENBRUGGE, G. C.; PEREIRA, A. A.; PICANÇO-RODRIGUES, D. Origin and domestication of native Amazonian crops. Diversity, v. 2, p. 72-106, 2010.
- CLEMENT, C. R.; SANTOS, RonaldoP.; DESMOULIERE, Sylvain J. M.; FERREIRA, Evandro J. L.; FARIAS NETO, JoãoTomé. Ecological adaptation of wild peach palm, its in situ conservation and deforestation-mediated extinction in southern Brazilian Amazonia. PloS ONE, v. 4, n. 2, p. e4564, 2009a.
- CLEMENT, C. R.; RIVAL, L.; COLE, D. M. Domestication of peach palm (Bactris gasipaes Kunth): the roles of human mobility and migration. In: ALEXIADES, M. N. (Ed.). Shifting spaces, changing times: mobility, migration and displacement in indigenous lowland South America, Oxford: Berghahn Books, 2009b. p. 117-140.
- CLEMENT, C. R.; BORÉM, A.; LOPES, M. T. G. Da domesticação ao melhoramento de plantas. In: BORÉM, A.; LOPES, M. T. G.; CLEMENT, C. R. (Eds.) Domesticação e melhoramento: espécies amazônicas. Viçosa: UFV, 2009c. p. 11-38.
- COELHO, V. P. A festa do pequi e o zunidor entre os índios Waurá. Bulletin de la Société Suisse des Américanistes, n. 55-56, p. 37-56, 1992.
- COSTA, L. As faces do jaguar Parentesco, história e mitologia entre os Kanamari da Amazônia ocidental. 2007. Tese (Doutorado em Antropologia Social) – Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2007.
- DESCOLA, P. Par-delà nature et culture Paris: Gallimard, 2005.
- DESCOLA, P. Ecologia e cosmologia. In: DIEGUES, A. C. S. (Ed.). Etnoconservação: novos rumos para a proteção da natureza nos trópicos. São Paulo: Hucitec, 2000. p. 149-164.
- DESCOLA, P. La nature domestique: symbolisme et praxis dans l’écologie des Achuar. Paris: Maison des Sciences de l’Homme, 1986.
- ELIAS, M.; MCKEY, Doyle; PANAUD, Olivier; ANSTETT, Marie Charlotte; ROBERT, Thierry. Traditional management of cassava morphological and genetic diversity by the Makushi Amerindians (Guyana, South America): perspectives for on-farm conservation of crop genetic resources. Euphytica, v. 120, n. 1, p. 143-157, 2001.
- ELIAS, M.; RIVAL, L.; MCKEY, D. Perception and management of cassava (Manihot esculenta Crantz) diversity among Makushi Amerindians of Guyana (South America). Journal of Ethnobiology, v. 20, n. 2, p. 239-265, 2000.
- EMPERAIRE, L. Histórias de plantas, histórias de vida: uma abordagem integrada da diversidade agrícola tradicional na Amazônia. Revista da Sociedade Brasileira de Etnobiologia e Etnoecologia, v. 3, p. 189-198, 2006.
- EMPERAIRE, L. A biodiversidade agrícola na Amazônia brasileira: recurso e patrimônio. Revista do Patrimônio Histórico e Artístico Nacional, v. 32, p. 23-45, 2005.
- EMPERAIRE, L. A agrobiodiversidade em risco: o exemplo das mandiocas na Amazônia. Ciência Hoje, v. 32, n. 187, p. 28-33, 2002.
- EMPERAIRE, L.; VAN VELTHEN, L.; OLIVEIRA, A. G. Patrimônio cultural imaterial e sistema agrícola: o manejo da diversidadeagrícola no médio rio negro. In: REUNIÃO BRASILEIRA DE ANTROPOLOGIA, 26., 2008, Porto Seguro. Anais.. Porto Seguro: ABA, 2008. p. 1-16.
- EMPERAIRE, L.; ELOY, L. A cidade, um foco de diversidade agrícola no Rio Negro (Amazonas, Brasil)? Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, v. 3, n. 2, p. 195-211, 2008.
- ERICKSON, C. Amazonia: the historical ecology of a domesticated landscape. In: SILVERMAN, H.; ISBELL, W. H. (Eds.). The handbook of South American archaeology New York: Springer, 2008. p. 157-183.
- FAUSTO, C. Too many owners: mastery and ownership in Amazonia. In: BRIGHTMAN, M.; GROTTI, V. E.; ULTURGASHEVA, O. (Orgs.). Animism in forest and tundra: personhood, animals, plants and things in contemporary Amazonia and Siberia. London: Berghahn, 2012a. p. 85-105.
- FAUSTO, C. Sangue de lua: reflexões ameríndias sobre espíritos e eclipses. Journal de la Société des Américanistes, v. 98, n. 1, p 63-80, 2012b.
- FAUSTO, C. Mil años de transformación: la cultura de la tradición entre los del alto Xingú. In: CHAUMEIL, J. P.; ESPINOSA, O.; CORNEJO, M. (Orgs.). Por donde hay soplo: estudios amazónicos en los países andinos. Lima: IFEA-CAAP-PUCP, 2011. (Actes & Mémoires, 29). p. 185-216.
- FAUSTO, C. Donos demais: maestria e domínio na Amazônia. Mana, v. 14, n. 2, p. 329-366, 2008.
- FAUSTO, C. Inimigos fiéis: história, guerra e xamanismo na Amazônia. São Paulo: EDUSP, 2001.
- FAUSTO, C.; FRANCHETTO, B.; MONTAGNANI, T. Les formes de la mémoire: art verbal et musique chez les Kuikuro du Haut-Xingu (Brésil). L’Homme, n. 197, p. 41-70, 2011.
- FAUSTO, C.; FRANCHETTO, B.; HECKENBERGER, M. J. Language, ritual and historical reconstruction. In: HARRISON, K. D.; DAVID, S. R.; DWYER, A. (Eds.). Lessons from documented endangered languages Amsterdam: John Benjamins, 2008. p. 129-157.
- FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS (FAO). Background Papers 1: agricultural biodiversity. In: FAO/NL CONFERENCE ON THE MULTIFUNCTIONAL CHARACTER OF AGRICULTURE AND LAND, 1., Maastricht, 1999. Background Papers.. Maastricht: FAO, 1999. Disponível em: <http://www.fao.org/mfcal/pdf/bp_1_agb.pdf>. Acesso em: 10 maio 2014.
» http://www.fao.org/mfcal/pdf/bp_1_agb.pdf - FRANCHETTO, B. L’autre du même: parallélisme et grammaire dans l’art verbal des récits Kuikuro (caribe du Haut Xingu, Brésil). Ameríndia, n. 28, p. 213-248, 2003.
- FRANCHETTO, B. Línguas e histórias no alto Xingu. In: FRANCHETTO, B.; HECKENBERGER, M. (Orgs.). Os povos do alto Xingu: história e cultura. Rio de Janeiro: UFRJ, 2001. p. 111-156.
- GORDON, C. Economia selvagem: ritual e mercadoria entre os índios Xikrin-Mebêngôkre. São Paulo: Editora UNESP/Instituto Socioambiental/Núcleo de Transformações Indígenas, 2006.
- GREGOR, T. Anxious pleasures: the sexual lives of an Amazonian people. Chicago/London: The University of Chicago Press, 1985.
- GRIBEL, R.; HAY, J. D. Pollination ecology of Caryocar brasiliense(Caryocaraceae) in Central Brazil cerrado vegetation. Journal of Tropical Ecology, v. 9, n. 2, p. 199-211, 1993.
- HECKENBERGER, M. J. The ecology of power New York: Routledge, 2005.
- HECKENBERGER, M. Estrutura, história e transformação: a cultura xinguana na longue durée, 1000-2000 D.C. In: FRANCHETTO, B.; HECKENBERGER, M. (Orgs.). Os povos do alto Xingu: história e cultura. Rio de Janeiro: UFRJ, 2001. p. 21-62.
- HECKENBERGER, M. J.; KUIKURO, A.; KUIKURO, O. T.; RUSSEL, C.; SCHMIDT, M.; FAUSTO, C.; FRANCHETTO, B. Amazonia 1492: pristine forest or cultural parkland? Science, v. 301, n. 5640, p. 1710-1714, 2003.
- HECKLER, S.; ZENT, S. Piaroa manioc varietals: hyperdiversity or social currency? Human Ecology, v. 36, n. 5, p. 679-697, 2008.
- INGOLD, T. The perception of the environment: essays on livelihood, dwelling and skill. London/New York: Routledge, 2000.
- IVANAUSKAS, N. M.; MONTEIRO, R.; RODRIGUES, R. R. Classificação fitogeográfica das florestas do Alto Rio Xingu. Acta Amazonica, v. 38, n. 3, p. 387-402, 2008.
- IVANAUSKAS, N. M.; MONTEIRO, R.; RODRIGUES, R. R. Estrutura de um trecho de floresta Amazônica na bacia do alto rio Xingu. Acta Amazonica, v. 34, n. 2, p. 275-299, 2004.
- KERR, W. Agricultura e seleções genéticas de plantas. In: RIBEIRO, B. (Ed.). Suma etnológica brasileira: etnobiologia. Petrópolis: Vozes, 1986. v. 1, p. 159-172.
- LATOUR, B. Reassembling the social: an introduction to actornetwork-theory. Oxford/New York: Oxford University Press, 2005.
- LATOUR, B. Nous n’avons jamais été modernes: essai d’anthropologie symétrique. Paris: Editions La Découverte, 1991.
- LIMA, D.; STEWARD, A.; RICHERS, B. T. Trocas, experimentações e preferências: um estudo sobre a dinâmica da diversidade da mandioca no médio Solimões, Amazonas. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, v. 7, n. 2, p. 371396, 2012.
- MACHADO, J. S. Lugares de gente: mulheres, plantas e redes de troca no delta amazônico (Caviana, PA). 2012. Tese (Doutorado em Antropologia Social) – Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2012.
- MARTINS, P. S. Dinâmica evolutiva em roças de caboclos amazônicos. Estudos Avançados, v. 19, n. 53, p. 209-220, 2005.
- MEIRA, S.; FRANCHETTO, B. The southern Cariban languages and the Cariban family. International Journal of American Linguistics, v. 71, n. 2, p. 127-192, 2005.
- PEREIRA, A. V.; PEREIRA, E. B. C.; SILVA, D. B.; GOMES, A. C.; SOUSA SILVA, J. C. Quebra da dormência de sementes de pequi. Boletim de Pesquisa e Desenvolvimento, v. 136, p. 1-15, 2004.
- POSEY, D. A. Etnobiologia: teoria e prática. In: RIBEIRO, B. (Ed.). Suma etnológica brasileira: etnobiologia. Petrópolis: Vozes, 1986. v. 1, p. 15-28.
- RESENDE, I. L. M.; ARAÚJO, G. M.; ASSIS-OLIVEIRA, A. P. A comunidade vegetal e as características abióticas de um campo de murundu em Uberlândia, MG. Acta Botanica Brasilica, v. 18, n. 1, p. 9-17, 2004.
- RIVAL, L. Domesticating the landscape, producing crops, and reproducing society in Amazonia. In: PARKIN, D.; ULIJASZEK, S. (Eds.). Convergence and emergence: towards a new holistic anthropology? Oxford: Berghahn Books, 2007. p. 72-90.
- RIVAL, L. Domestication as a historical and symbolic process: wild gardens and cultivated forests in the Ecuadorian Amazon. In: BALÉE, W. (Org.). Advances in historical ecology New York: Columbia University Press, 1998. p. 232-250.
- RIVAL, L.; MCKEY, D. 2008. Domestication and diversity in manioc (Manihot esculenta Crantz ssp. esculenta, Euphorbiaceae). Current Anthropology, v. 49, n. 6, p. 1119-1128, 2008.
- SCHMIDT, M. J. Reconstructing tropical nature: prehistoric and modern anthrosols (Terra Preta) in the Amazon rainforest, Upper Xingu river, Brazil 2010. Tese (Doutorado em Geografia) –University of Florida, Gainesville, 2010.
- SCHMIDT, M. J.; HECKENBERGER, M. J. Formação de Terra Preta na região do alto Xingu: resultados preliminares. In: TEIXEIRA, W.; KERN, D. C.; MADARI, B.; LIMA, H. N. (Eds.). As terras pretas de índio da Amazônia: sua caracterização e uso deste conhecimento na criação de novas áreas. Manaus: EDUA, 2009. p. 103-126.
- SEIXAS, A. C. P. S. Entre terreiros e roçados: a construção da agrobiodiversidade por moradores do Rio Croa, Vale do Juruá (AC). 2008. 124 f. Dissertação (Mestrado em Desenvolvimento Sustentável: Política e Gestão Ambiental) – Universidade de Brasília, Brasília, 2008.
- SHANNON, C. E.; WEAVER, W. The mathematical theory of communication Urbana: Illinois University, 1949. 117 p.
- SMITH, M. Árvores de cultura: cultivo e uso do pequi (Caryocar sp., Caryocaraceae) entre os Kuikuro do alto Xingu, MT. 2013. 221 f. Tese (Doutorado em Desenvolvimento Sustentável: Política e Gestão Ambiental) – Universidade de Brasília, Brasília, 2013.
- TER STEEGE, H.; PITMAN, Nigel C. A.; SABATIER, Daniel; BARALOTO, Christopher; SALOMÃO, Rafael P.; GUEVARA, Juan Ernesto; PHILLIPS, Oliver L.; CASTILHO, Carolina V.; MAGNUSSON, William E.; MOLINO, Jean-François; MONTEAGUDO, Abel; VARGAS, Percy Núñez; MONTERO, Juan Carlos; FELDPAUSCH, Ted R.; CORONADO, Eurídice N. Honorio; KILLEEN, Tim J.; MOSTACEDO, Bonifacio; VASQUEZ, Rodolfo; ASSIS, Rafael L.; TERBORGH, John; WITTMANN, Florian; ANDRADE, Ana; LAURANCE, William F.; LAURANCE, Susan G. W.; MARIMON, Beatriz S.; MARIMON JR., Ben-Hur; VIEIRA, Ima Célia Guimarães; AMARAL, Iêda Leão; BRIENEN, Roel; CASTELLANOS, Hernán; LÓPEZ, Dairon Cárdenas; DUIVENVOORDEN, Joost F.; MOGOLLÓN, Hugo F.; MATOS, Francisca Dionízia de Almeida; DÁVILA, Nállarett; GARCÍA-VILLACORTA, Roosevelt; DIAZ, Pablo Roberto Stevenson; COSTA, Flávia; EMILIO, Thaise; LEVIS, Carolina; SCHIETTI, Juliana; SOUZA, Priscila; ALONSO, Alfonso; DALLMEIER, Francisco; MONTOYA, Alvaro Javier Duque; PIEDADE, Maria Teresa Fernandez; ARAUJO-MURAKAMI, Alejandro; ARROYO, Luzmila; GRIBEL, Rogerio; FINE, Paul V. A.; PERES, Carlos A.; TOLEDO, Marisol; AYMARD C., Gerardo A.; BAKER, Tim R.; CERÓN, Carlos; ENGEL, Julien; HENKEL, Terry W.; MAAS, Paul; PETRONELLI, Pascal; STROPP, Juliana; ZARTMAN, Charles Eugene; DALY, Doug; NEILL, David; SILVEIRA, Marcos; PAREDES, Marcos Ríos; CHAVE, Jerome; LIMA FILHO, Diógenes de Andrade ; JØRGENSEN, Peter Møller; FUENTES, Alfredo; SCHÖNGART, Jochen; VALVERDE, Fernando Cornejo; FIORE, Anthony Di; JIMENEZ, Eliana M.; MORA, Maria Cristina Peñuela; PHILLIPS, Juan Fernando; RIVAS, Gonzalo; VAN ANDEL, Tinde R.; VON HILDEBRAND, Patricio; HOFFMAN, Bruce; ZENT, Eglée L.; MALHI, Yadvinder; PRIETO, Adriana; RUDAS, Agustín; RUSCHELL, Ademir R.; SILVA, Natalino; VOS, Vincent; ZENT, Stanford; OLIVEIRA, Alexandre A.; SCHUTZ, Angela Cano; GONZALES, Therany; NASCIMENTO, Marcelo Trindade; RAMIREZ-ANGULO, Hirma; SIERRA, Rodrigo; TIRADO, Milton; MEDINA, María Natalia Umaña; VAN DER HEIJDEN, Geertje; VELA, César I. A.; TORRE, Emilio Vilanova; VRIESENDORP, Corine; WANG, Ophelia; YOUNG, Kenneth R.; BAIDER, Claudia; BALSLEV, Henrik; FERREIRA, Cid; MESONES, Italo; TORRES-LEZAMA, Armando; GIRALDO, Ligia Estela Urrego; ZAGT, Roderick; ALEXIADES, Miguel N.; HERNANDEZ, Lionel; HUAMANTUPA-CHUQUIMACO, Isau; MILLIKEN, William; CUENCA, Walter Palacios; PAULETTO, Daniela; SANDOVAL, Elvis Valderrama; GAMARRA, Luis Valenzuela; DEXTER, Kyle G.; FEELEY, Ken; LOPEZ-GONZALEZ, Gabriela; SILMAN, Miles R. Hyperdominance in the Amazonian tree flora. Science, v. 342, n. 6156, p. 1243092-1-1243092-9, 2013.
- THRUPP, L. A. Linking agricultural biodiversity and food security: the valuable role of agrobiodiversity for sustainable agriculture. International Affairs, v. 76, n. 2, p. 265-281, 2000.
- VELOSO, H. P.; RANGEL FILHO, A. L. R.; LIMA, J. C. A. Classificação da vegetação brasileira, adaptada a um sistema universal Rio de Janeiro: IBGE, 1991. 112 p.
- VILLAS-BÔAS, A. De olho na bacia do Xingu São Paulo: Instituto Socioambiental, 2012. (Série Cartô Brasil Socioambiental, n. 5). 61 p.
- VIVEIROS DE CASTRO, E. Os pronomes cosmológicos e o perspectivismo ameríndio. Mana, v. 2, n. 2, p. 115-144, 1996.
- WITTENBURG, P.; BRUGMAN, H.; RUSSEL, A.; KLASSMANN, A.; SLOETJES, H. Elan: a professional framework for multimodality research. In: INTERNATIONAL CONFERENCE ON LANGUAGE RESOURCES AND EVALUATION, 5., Genoa, 2006. Proceedings… Genoa: ELRA, 2006. p. 1556-1559. Disponível em: <http://www.lrec-conf.org/proceedings/lrec2006/pdf/153_pdf.pdf>. Acesso em: 10/05/2014
» http://www.lrec-conf.org/proceedings/lrec2006/pdf/153_pdf.pdf - WOOD, D.; LENNÉ, J. M. The conservation of agrobiodiversity on-farm: questioning the emerging paradigm. Biodiversity & Conservation, v. 6, n. 1, p. 109-129, 1997.
Datas de Publicação
-
Publicação nesta coleção
Jan-Apr 2016
Histórico
-
Recebido
29 Nov 2013 -
Aceito
21 Mar 2016