Mouse Lipocalins (MUP, OBP, LCN) Are Co-expressed in Tissues Involved in Chemical Communication
 Romana Stopková1
Romana Stopková1  David Vinkler1
David Vinkler1  Barbora Kuntová1
Barbora Kuntová1  Ondrej Šedo2,3
Ondrej Šedo2,3  Tomáš Albrecht1 Jan Suchan1 Kateřina Dvořáková-Hortová1
Tomáš Albrecht1 Jan Suchan1 Kateřina Dvořáková-Hortová1  Zbyněk Zdráhal2,3
Zbyněk Zdráhal2,3  Pavel Stopka1*
Pavel Stopka1*- 1BIOCEV Group, Department of Zoology, Faculty of Science, Charles University in Prague, Prague, Czech Republic
- 2Research Group Proteomics, Faculty of Science, Central European Institute of Technology, Masaryk University, Brno, Czech Republic
- 3Faculty of Science, National Centre for Biomolecular Research, Masaryk University, Brno, Czech Republic
Chemical communication is mediated by signal production and signal perception and in house mice (Mus musculus), both processes involve lipocalin proteins (OBP, MUP, LCN) that transport volatiles and protect them in tissues where they are produced. However, potential roles of lacrimal, nasal, and salivary lipocalins are still not well known. We aimed to determine the expression of the recently described family of odorant binding proteins (Obp), along with major urinary proteins (Mup) across different tissues in wild mice (Mus musculus) to assess the importance of these proteins based on their quantity in particular expression sites. We performed qPCR analysis of selected Mup, Lcn, Obp genes, and predicted Obp members to study their expression in selected tissues. We identified new members of the mouse odorant binding protein gene family in two subspecies, M. m. musculus and M. m. domesticus. We show that Mup4 and Mup5 from the phylogenetically older group-A are co-expressed with Obps in orofacial tissues. We also identified a sexually dimorphic pattern of female-biased Obp7 and male-biased Mup4 expression in lacrimal glands. OBPs, MUPs, and LCNs are produced in parallel, which may function to widen the spectrum of bound ligands, potentially including the degradation products of olfactory signals and/or toxic compounds. Moreover, our study demonstrates that several pheromone transporters from the lipocalin family are co-expressed in the nasal and lacrimal tissues of mice with the newly detected OBPs that further expand the already diverse mouse lipocalin family.
Background
John Maynard Smith and David Harper defined signal as “…any act or structure which alters the behaviour of other organisms, which evolved because of that effect, and which is effective because the receiver's response has also evolved” (Maynard Smith and Harper, 2003). The house mouse (Mus musculus) uses a system of volatile pheromones (Mucignat-Caretta et al., 2010) and their transporters from the lipocalin protein family that together form a signal (Novotny et al., 1985). Because volatiles degrade in water solutions (Kwak et al., 2013), their life span largely depends on lipocalins that protect them (Hurst et al., 1998; Timm et al., 2001), and transport them in secretory fluids (Flower, 1996) to an outside world. The signals have strong effects on the reproductive success of the signaler (Thonhauser et al., 2013) due to strong effects on reproductive physiology of the receiver (Whitten et al., 1968; Roberts et al., 2004; Stopka et al., 2007; Janotova and Stopka, 2011) through chemosensory receptors of the main olfactory and vomeronasal organs (Moss et al., 1997; Luo and Katz, 2004).
Since the discovery of the structure and function of olfactory receptors GPCRs—G-protein coupled receptors (Buck and Axel, 1991), research on chemical communication has concentrated on signal reception by nasal and vomeronasal chemosensory neuronal receptors, and on lipocalin transporters of pheromones. Lipocalins generally function to sequester hydrophobic volatiles and transport them in their eight-stranded beta barrel structure (Timm et al., 2001; Sharrow et al., 2002). Volatiles specifically bind to receptors of chemosensory neurons when released (Tirindelli et al., 1998; Novotny, 2003). In mice, the functions of lipocalin transporters are not well understood and most studies focused on the major urinary proteins (MUPs), which are expressed in the liver and transport volatile odor/organic compounds (VOCs) to the urine (Shahan and Derman, 1984; Shahan et al., 1987a,b; Stopková et al., 2007). MUPs have also been reported to be expressed in several tissues other than the liver (Shaw et al., 1983; Shahan et al., 1987a; Cavaggioni et al., 1999; Utsumi et al., 1999; Karn and Laukaitis, 2011), though their functions are not understood.
Mup genes have recently duplicated in rodents, and in house mice they form a cluster of 21 coding genes (and a similar number of pseudogenes), which can be divided into two groups, the group-A (ancestral), containing Mup3, Mup4, Mup5, Mup6, Mup20 (or “Darcin”), and Mup21 and the group-B, consisting of 15 other Mups sharing almost 99% sequence identity: Mup1, Mup2, Mup7-Mup19 (Logan et al., 2008; Mudge et al., 2008), reviewed in Janotová and Stopka (2009), Stopková et al. (2009), and Phelan et al. (2014). The level of urinary MUP production is socially regulated in C57BL/6 laboratory mice (Stopka et al., 2007) and wild living M. m. musculus (Janotova and Stopka, 2011) and M. m. domesticus (Cunningham et al., 2013) mice. Furthermore, male M. m. musculus up-regulated urinary MUP production when caged with a female, but down-regulated when caged with a male. Down-regulation of MUPs was more pronounced in males that were defeated in a male–male encounter (Janotova and Stopka, 2011). Furthermore, social experience of parents can regulate MUP expression level in subsequent generations through epigenetic effects (Nelson et al., 2013).
Specific roles limited to a single urinary MUP were attributed only to a major urinary protein MUP20 (or “Darcin”; a group-A MUP) expressed in males, which attracts females and aids spatial learning (Roberts et al., 2010, 2012). Remaining MUPs were supposed to present an individual “barcode” signal due to differences in urinary MUP profiles (Hurst et al., 2001). However, a recent study with sufficient sample sizes shows that MUP profiles of wild male house mice (M. m. musculus) are not individually unique. They are not highly stable but instead are dynamic over time with significant changes after puberty and during adulthood (Thoß et al., 2015), thus challenging the “barcode” hypothesis. Moreover, the variation in pheromone affinities of the urinary MUP isoforms provides low support for the proposal that heterogeneity in MUPs plays a role in regulating profiles of available pheromones (Sharrow et al., 2002).
Another group of lipocalins that is thought to be involved in chemical communication, but less understood, is a cluster of the odorant binding protein genes (Obp). Obp genes have also undergone a series of duplications in mice, and they occur in a cluster of six genes and two pseudogenes on the X chromosome (Stopková et al., 2009, 2014, see Figure 1A). Whilst the Mup genes are abundant only in house mice and rats (Rattus norvegicus) and rarely found in other species of mammals in multiple copies, Obp genes occur as a cluster in various mammalian taxa, e.g., porcupines (Hystrix cristata) (Felicioli et al., 1993), bank voles (Myodes glareolus) (Stopkova et al., 2010), elephants (Lazar et al., 2002), cows (Bos taurus) (Bignetti et al., 1985), boar (Sus scrofa) (Spinelli et al., 1998; Nagnan-Le Meillour et al., 2014), and potentially also mole rats (Fukomys anselli, F. kafuensis) (Hagemeyer et al., 2011). One OBP member (i.e., Aphrodisin) has been shown to be major pheromone transporter in vaginal flushes of hamsters (Cricetus cricetus). Interestingly, pigs have OBPs and SAL. SAL is the major salivary protein in pigs with affinity to steroids and to 2-isobutyl-3-methoxypyrazine, it is phylogenetically close to MUPs and is expressed by the male submaxillary glands (Marchese et al., 1998). Moreover, three of the six predicted OBP members described (Stopková et al., 2009, 2014) were also corroborated with MS techniques in the tear and saliva proteomes of the laboratory mouse C57BL/6 (Karn and Laukaitis, 2015), though the authors did not further specify detected OBP variants. Therefore, one of the aims of our paper was to detect potential expression sites of mRNAs coding OBPs that were found in the mouse saliva.
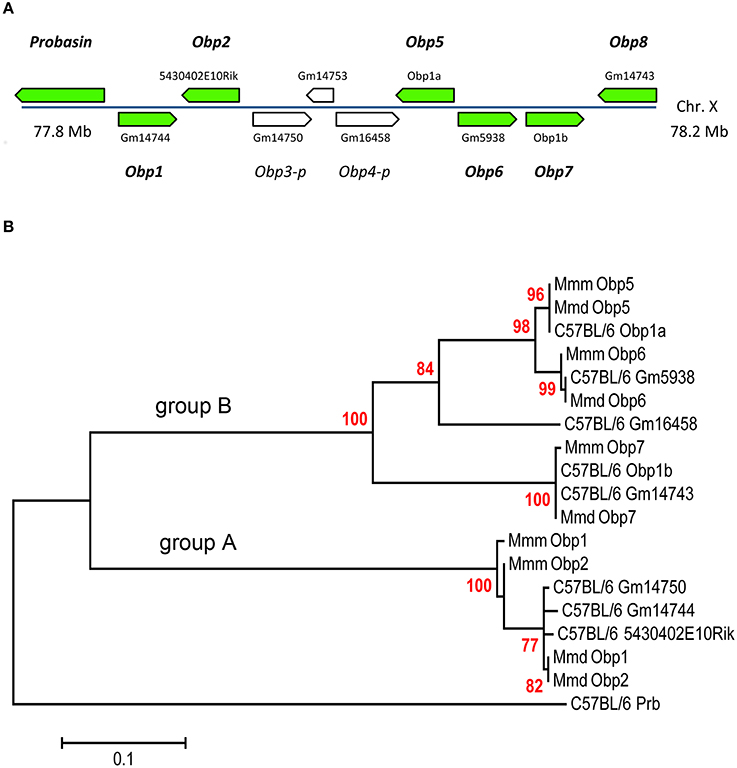
Figure 1. The mouse Obp cluster (A), and Phylogenetic analysis of novel OBPs by Maximum Likelihood method (B). Green arrows (A) indicate position of coding genes mapped on C57BL/6 X-chromosome with specific codes from the mouse genome. Consecutive numbering in italics (Obp1 to Obp8) represents alternative names for corroborated transcripts under this study in wild mice. This nomenclature also involves two pseudogenes (Obp3-p, Obp4-p) with positions indicated by white arrows. Pseudogene Gm14753 is not involved in numbering because it is a processed retroelement similar to actin and not Obp.
Because mice typically begin social interactions by investigating facial and mouth areas (Luo et al., 2003), we may assume that tear and salivary lipocalins secrete chemical signals, whilst nasal and vomeronasal lipocalins activate and/or deactivate chemosensory GPCR receptors. In addition to their function in chemical communication, some lipocalins also have important roles in innate immune responses (Fluckinger et al., 2004; Stopková et al., 2014). We have previously suggested that chemical communication and immunity have been shaped by similar evolutionary forces because the nasal cavity is a place of pathogen recognition via lymphoid tissues and signal perception via chemosensory neurons (Stopková et al., 2009, 2014). Moreover, lipocalins may have as yet another function. The “toxic waste hypothesis” states that various lipocalins are involved in removing toxic waste from the body (Stopková et al., 2009; Kwak et al., 2011) and that some of the compounds might have been constituting a signal under selection (Stopková et al., 2009). The toxic waste disposal role has been experimentally demonstrated in a recent paper (Kwak et al., 2016) where mice loaded with an industrial chemical, 2,4-di-tert-butylphenol (DTBP) used MUPs for a consequent detoxification (Kwak et al., 2016). To conclude, lipocalins are ubiquitous proteins with diverse functions and multiple sites of their expression.
In this study, we investigated potential differences in the expression of selected lipocalins in two sub-species of the house mouse, Mus musculus musculus and M. m. domesticus. This could be a starting point to determine how these proteins evolve through speciation (Hiadlovska et al., 2013), and their potential influence on sub-species recognition (Smadja and Ganem, 2002, 2008) and aggression (Dureje et al., 2011). These two sub-species have been previously found to vary quantitatively in the abundance of male VOCs (Mucignat-Caretta et al., 2010) and MUP expression between sexes (Stopková et al., 2007) with differences in the beta-barrel residues under selection (Karn and Laukaitis, 2012). Thus, we have identified new members of the odorant binding protein family and focused on the level and locations of expression of soluble lipocalins. It is our hope that investigating lipocalins in different tissues, and not only in the urine, will provide a better understanding of this fascinating and complex family of carrier proteins.
Methods
Ethical Standards
All animal procedures were carried out in strict accordance with the law of the Czech Republic paragraph 17 no. 246/1992 and the local ethics committee of the Faculty of Science of Charles University in Prague specifically approved this study in accordance with accreditation no. 27335/2013-17214 valid through 2019. Animals were sacrificed by cervical dislocation.
Samples
The total of 12 individuals (i.e., six males and six females) was studied in this experiment with six individual M m. domesticus from Hattingen (51°20′39.84″N, 7°12′06.38″E) and Ruther (51°23′01.0″N, 6°57′48.9″E) and six individual Mus m. musculus from Jičín (50°28′18.802″N, 15°22′31.667″E). Individual mice were kept for 6 weeks following capture in the accredited mouse facility with food and water provided ad libitum and on a 12:12-h light cycle with lights off at 1900 h. Samples for 2DE were collected from the oral cavity with 100 μl of 0.9% saline water repeatedly flushing in and out with a pipette. Samples were immediately acetone precipitated and used in further steps.
When salivary samples were collected, all specimens were sacrificed by cervical dislocation and tissue samples were collected from each animal. Tissue samples were obtained from preputial/clitorial glands, liver, lacrimal and Harderian glands, submandibular (salivary) glands, olfactory epithelia, vomeronasal organ, and Nasal-associated lymphoid tissue (NALT). NALT is the paired lymphoid organ (Kiyono and Fukuyama, 2004), and it was isolated from the upper mouse jaw by peeling away the palate where NALT was localized bilaterally on the posterior side.
2DE-Page Analysis
Two dimensional polyacrylamide electrophoresis (2DE) was performed with IEF cell (Bio-rad®) and Protean II electrophoresis system. For the first dimension 12 μg of proteins was applied to Bio-Rad 11 cm strips (pI: 3.9–5.1). Isoelectric focusing was performed after passive rehydration at room temperature and run at 50 V for 9 h, 250 V (rapid) for 15 min, 8000 V (rapid) for 1 h, 8000/30,000 V/h, and finished at 500 V until further step. For the second- dimension separation—strips were equilibrated for 10 min in 45 mM Tris base (pH 7.0) containing 6 M urea, 1.6 SDS, 30% glycerol, and 130 mM dithiothreitol, and then re-equilibrated for 10 min in the same buffer containing 135 mM iodoacetamide in place of dithiothreitol. The strips were then placed on Criterion (dodeca) precast 12–20% gels along with unstained molecular standards in a separate well. Second dimension gels were run at constant current—50 mA for 1 h, 100 mA for 1 h and 150 mA for 1.5 h at 10°C. After electrophoresis, the gels were stained with the Colloidal Coomassie G-250 stain (Bio-rad). All spots in the range 15–30 kDa were excised with a Bio-rad Spot Cutter.
MALDI-MS/MS Analysis
The most abundant protein spots were selected for the analysis and excised from 2-DE gels from 12 individuals. Gel pieces were destained by alternative washing steps using 50 mM ammonium bicarbonate and acetonitrile (i.e., provided in detail in Data Sheet 1). After destaining, the proteins in gel pieces were incubated with trypsin (sequencing grade, Promega) at 37°C for 2 h. Digested peptides were extracted from gels using 50% ACN solution with 5% formic acid. MALDI- MS/MS analyses were performed on an Ultraflex III mass spectrometer (Bruker Daltonik, Bremen, Germany). Peptide maps were acquired inreflectron positive mode (25 kV acceleration voltage) with 800 laser shots. Peaks within 700–4000 Da mass range and minimum S/N 10 were picked out for MS/MS analysis employing LID-LIFT arrangement with 600 laser shots for each peptide.
CHCA was used as the matrix in combination with AnchorChip target to enhance measurement sensitivity. Sample (1 μl) was mixed with matrix solution on the target in a 2:1 ratio. Known autoproteolytic products of trypsin were used for internal calibration of digested peptides. In the absence of these products, an external calibration procedure was employed, using a mixture of seven peptide standards (Bruker Daltonik) covering the mass range of 1000–3100 Da. The Flex Analysis 3.0 and MS Biotools 3.1 (Bruker Daltonik) software were used for data processing.
Data Processing
MASCOT 2.2 (MatrixScience, London, UK) search engine was used for processing the MS/MS data under standard settings with significance threshold p < 0.05. Database searches were done against the NCBI protein database (Release 20101113) without taxonomic restriction. Mass tolerances of peptide precursors and MS/MS fragments were set to 60 ppm and 0.7 Da, respectively. Trypsin specificity with possibility of semitryptic cleavage, oxidation (M), carbamidomethylation (C) and pyro-Glu (Q, N-term) as optional modifications and up to two enzyme miscleavages were set for all searches. Protein identifications based on one or more unique peptides with significant score (under the settings—59 or higher) were accepted. See more details in Data Sheet 1.
Real-Time PCR Analysis
Immediately after resection, each tissue sample was placed into Eppendorff tube with a mixture of 1 ml of Trizol (TRIzol Reagent–Invitrogen) and glass pellets, and homogenized using a homogenizer (MM200–Retsch). RNA was isolated using standard Trizol protocol and followed by cDNA synthesis using First strand cDNA synthesis kit (Fermentas). RNA was assessed from the ratio of the optical densities at 260 and 280 nm, and the RNA integrity was assessed with 1% agarose gel containing ethidium bromide. One microgram of total RNA (DNase treated) was used for the synthesis of single-stranded cDNA according to a first-strand cDNA synthesis protocol (Fermentas UAB, Vilnius, Lithuania) with RevertAid™M-MulV Reverse Transcriptase and oligo(dT)18 primer.
Real-time PCR was performed on a Light Cycler 480 (Roche Applied Sciences) using specific dual hydrolysation probe method (Universal Probe–Roche Applied Sciences) with the Probe Master kit (Roche) and protocol according to the manufacturer's instructions. Specific primers and their respective probes were designed by Universal probe library software (Roche) using our newly provided (i.e., Obp) and NCBI reference sequences. Intron-spanning assay and multiplex PCR condition with reference gene (Gapdh) were selected. The resulting primers and probes are provided in Table 1. Moreover, most group-B Mups are almost 99% similar and it is difficult on the level of transcript to find a probe that would differentiate between different group-B Mups. Thus, most urinary Mups including Mup2 is included in Figure 3 within the category MUP-B (detected with universal group-B Mup primers).

Table 1. Primers and probes used in Real-time qPCR analysis.
PCR amplification was performed with the following conditions: initial denaturation at 95°C for 10 min, followed by 40 cycles consisting of denaturation at 95°C for 10 s, annealing at 60°C for 10 s where fluorescence was acquired, and elongation at 72°C for 5 s. Each sample was measured in triplicate. The data used for calculation are the means of Cq (i.e., cycle of quantitation in Roche software) values of triplicate samples. The variation in triplicate values never exceeded 0.5 Cq in our samples.
The level of mRNA of the target gene (Obp etc.,) in each sample was calculated relative to the reference gene (Gapdh) amplified in the same well. A calibration curve was generated for each pair of primers using 10-fold serial dilution of cDNA to assess the value for PCR efficiency (E). In all cases E was not lower than 0.9 (i.e., 90% efficiency of PCR reaction). E-values were than used in the formula (The Efficiency sensitive model Pfaffl, 2001) used for the calculation of relative expression (RE), i.e., normalized mRNA abundance:
Non-template and non-RT reactions were used as controls. For the analysis of expression patterns via hierarchical clustering we used R software. Our hierarchical clustering utilized Euclidian distance metric on log2 transformed data and complete linkage method. The mixed-model approaches, ANOVA, t-test, Shapiro-Wilk's normality test, and Fligner-Killeen test for testing the homogeneity of variances, were also computed and plotted in R (Venables and Smith, 2009). Data for pI values were cross-checked from multiple online resources (Ensembl Genome Browser—www.ensembl.org/, NCBI) and with our recently obtained sequences. Isoelectric point was calculated with ExPASy (http://web.expasy.org/compute_pi/), whilst the index of hydropathy (i.e., GRAVY index) was calculated with Gravy calculator (www.gravy-calculator.de/). Sequence data are provided as additional Data Sheet 2, data for calculation of pI and H are provided in Data Sheet 3.
Sanger Sequencing
Various primer sets were derived from predicted sequences of the genome mouse C57BL/6 (Stopková et al., 2009, 2014) and used to amplify Obp transcripts. Mixed samples from studied orofacial tissues were used for transcript identification. Finally we set up a pair of primers per transcript giving one clear band in the expected area. These sequencing primers covered the whole region from start to stop codon: Obp1- (F—CTCTGAACTCCTTCGGAAGGA, R—AAAAGAATCAGTACCATGGTAGGA), Obp5—(F—CTGTAGAAAAGAAAGTCTTGTACCA, R—CATTCAAAAAAGGAAGATCATGAGA), Obp6—(F—AAGTCTTGTGCCATAATGGCAA, R—TCAAAAAAAGGAATAACAGGTCGTA), Obp7—(F - TGAACATCTCCAGAGGAGCAA, R—GGAAGAAGAGTTTATAGATTAGGCAA). The products were double sequenced (downstream, upstream) with 3130 Genetic Analyzer, Applied Biosystems using either forward or reverse Obp primers and with 5 to 10 technical replicates per transcript. Sequences were analyzed using the Sequence Scanner (Applied Biosystem) software and compared to predicted and known sequences from public database NCBI using BLAST. Novel sequences were deposited in GeneBank with accession numbers provided in the results section.
Phylogenetic Analysis of Obps by Maximum Likelihood Method
The evolutionary history was inferred by using the Maximum Likelihood method based on the JTT matrix-based model (Jones et al., 1992). The tree with the highest log likelihood (−1578.6730) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The analysis involved 19 nucleotide sequences. Codon positions included were 1st + 2nd + 3rd + Noncoding. All positions with less than 95% site coverage were eliminated. That is, fewer than 5% alignment gaps, missing data, and ambiguous bases were allowed at any position. There were a total of 284 positions in the final dataset. Evolutionary analyses were conducted in MEGA5 (Tamura et al., 2011).
Results
Saliva Contains Lipocalins
To detect the expression of lipocalins on the level of proteins, we used the MALDI- MS/MS analyses for protein identification in mouse saliva. In both subspecies, we have identified several abundant lipocalins (Figure 2): OBP5 (Odorant binding protein 1a, gi|1835143), LCN11 (Lipocalin 11, gi|154689678), MUP5 (Major urinary protein 5 precursor, gi|113930708), and highly similar group-B MUPs with the most likely identification provided in Data Sheet 1. We have also identified a fragment of a putative pheromone transporter VM (Vomeromodulin precursor, gi|70909314) which is a 70 kDa glycoprotein expressed in the posterior septal and vomeronasal glands but not in the mucus of the main olfactory neuroepithelium (Khew-Goodall et al., 1991). The presence of VM (and potentially also OBP5) in the mouse saliva suggests that nasal and oral cavities are functionally connected because proteins expressed by nasal/vomeronasal tissues are also found in the oral cavity. We have also identified several proteins below 17 kDa size including PIP [Prolactin Inducible Protein—also highly abundant in saliva of the laboratory mouse (Blanchard et al., 2015)], and several unspecified members of ABP (androgen binding protein) family which were recently described in detail in the tear and saliva proteomes of the genome mouse C57BL/6 (Blanchard et al., 2015; Karn and Laukaitis, 2015).

Figure 2. Proteomic search for acidic proteins with hydrophobic beta barrel. Proteomic 2DE analysis on narrow range strips (pI 3.9–5.1) revealed that mouse saliva from a male (M. m. domesticus) contains proteins depicted in the figure including OBP5 (MMD11 spot in A1 file), group-A MUP5 (spots: MMD15, MMD17), group-B MUP2 (spots: MMD12-14), unspecified ABPs (spots: MMD18) and prolactin-inducible protein, PIP (spot MMD16). In most gels (i.e., from males and females) we also identified VM (vomeromodulin), LCN11 (lipocalin11). See additional file A1 for a MS report (i.e., section Mus musculus domesticus—male).
mRNA Sequencing Corroborated Predicted Obps
In contrast to MUPs, the family of OBPs is rather enigmatic with respect to the expression of their predicted members. In studying the poorly-described OBP1a protein found in mouse saliva, we discovered that it is related to a gene cluster that had been incompletely described (Stopková et al., 2014). Therefore, we have sequenced all Obp predicted transcripts in wild mice from pooled oro-facial tissues using primers generated from C57BL/6 genomic data and provided specific product names based on their chromosomal position (for MUPs see Logan et al., 2008). All Obp transcripts were mapped on the X chromosome of the laboratory mouse C57BL/6 and have been given consecutive names Obp1-Obp8 (Figure 1A). We have also included two pseudogenes (i.e., Obp3-p, Obp4-p; classified by Ensembl as unprocessed pseudogenes without a protein product) in our consecutive numbering of Obps as in other rodent taxa these genes may be intact with ORF (i.e., not truncated) and expressed. Thus, we have provided unique Obp sequences for feral M. m. domesticus (Obp1—KJ605385, Obp2—KJ605386, Obp5—KJ605387, Obp6—KJ605388, and Obp7—KJ605389), and M. m. musculus (Obp1—KJ605390, Obp2—KJ605391, Obp5—KJ605392, Obp6—KJ605393, and Obp7—KJ605394) and submitted them to GenBank (NCBI).
Phylogenetic Analysis of Novel Obp Sequences
All novel OBPs have a feature typical for the entire Obp cluster - a specific disulfide bond (Cys38–Cys42), which represents a strong OBP-diagnostic motif CXXXC (Cys-Xaa-Xaa-Xaa-Cys; reviewed in Stopková et al., 2009, 2014). We used our mRNA (i.e., CDS) sequences along with those from C57BL/6 mice to generate the Maximum Likelihood (MLM) tree (Figure 1B). The MLM algorithm with 2000 permutations identified Prb (Probasin) as the root (i.e., the outgroup to all OBPs). Thus, remaining Obps form two sub-clusters that we decided to name as the group-A and the group-B Obps. Ancestral group-A Obps include Obp1 and Obp2 (bootstrap = 100). The later evolved group-B Obps include Obp5, Obp6, Obp7, and Obp8 (bootstrap = 99). The group-B Obp sequences perfectly match those predicted transcripts that we extracted from the laboratory mouse genome (Figure 1A). However, newly described Obp1 and Obp2 from M. m. musculus cluster together (bootstrap = 90) and seem to be divergent from M. m. domesticus and C57BL/6 (see Data Sheet 2 for Multiple sequence alignment).
The strong CXXXC motif present in all OBP proteins (including Probasin) is represented by CNDDC in OBP1 and OBP2, CDEGC in OBP7 and OBP8, CEKEC in OBP5 and OBP6. Obp3-p pseudogene (if expressed) would belong to the group-A cluster whilst Obp4-p would belong to the group-B, Figure 1B. Novel Obp (transcript) sequences along with the Mup sequences downloaded from NCBI were translated and further used for the calculation of hydropathy and pI properties. We also measured the expression of these newly identified genes in numerous tissues.
Hierarchical Clustering Revealed Differential mRNA Expression Across Tissues
We assessed expression of lipocalins in eight tissues using qRT-PCR. We were primarily interested in the location of expression of the newly described OBPs. So we designed primers for all OBPs and for other lipocalins from the list of identified proteins and the VNO-specific LCN4. In the next step, we used hierarchical clustering in R software as a graphical method to show relationships among expression levels of different genes across tissues. Primarily, we focused on the detection of similarities among expression levels in different tissues and averaged across individuals of the two subspecies to cluster particular tissues on the basis of their similar pattern (Figures 3A,B).
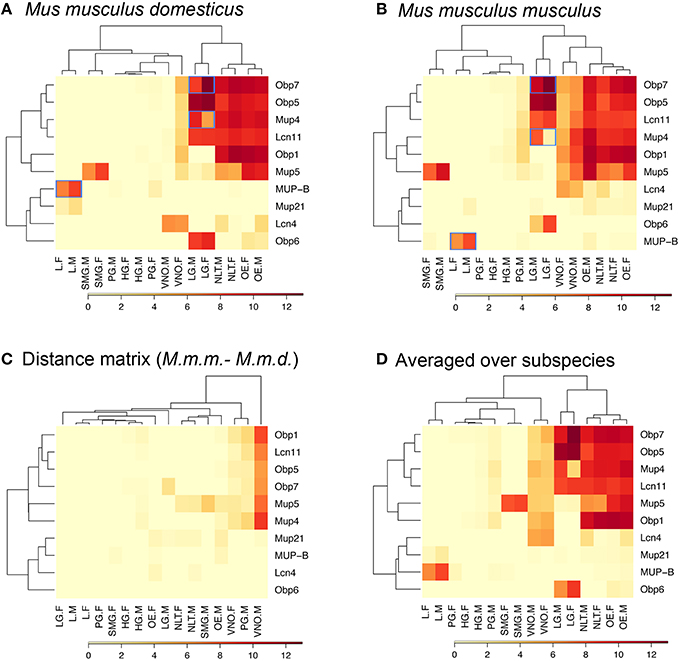
Figure 3. Graphical representation of the qPCR expression pattern. Group-A Mups are co-expressed with Obps and Lcns in sensory tissues and exo-orbital lacrimal glands, whilst the later evolved group-B Mups are an outgroup for their specific expression and excretory functions by the liver. Blue-framed are the significantly sexually dimorphic genes (i.e., male-biased group-B Mups in the liver, male-biased group-A Mup4 in lacrimal glands, and female-biased Obp7 in lacrimal glands). Abbreviations: L, Liver; SMG, submandibular gland; PG, preputial gland; HG, Harderian gland; LG, lacrimal gland; VNO, vomeronasal organ; NLT, NALT / nasal-associated lymphoid tissue; OE, olfactory epithelia; F, females; M, males; Highly expressed genes (~12 fold) are dark red whilst low expression genes are in pale yellow. Heat-maps are provided for each sub-species, (A) M. m. domesticus, (B) M. m. musculus, for the distance between them (C) and their average (D).
Hierarchical clustering separated selected tissues according to their pattern of expression into two groups depicted on (upper) X axis in Figure 3. Interestingly, in M. m. domesticus - olfactory epithelia (OE), lacrimal gland (LG), and nasal-associated lymphoid tissue (NLT) have clustered together in both subspecies (see Figures 3A,B), whilst vomeronasal organ (VNO) and other secretory tissues including liver, Harderian gland, submandibular gland, and preputial gland were located on the other branch in M. m. domesticus (Figure 3A). In M. m. musculus VNO clustered together with OE, LG, and NLT (Figure 3B), thus suggesting higher VNO activity in this subspecies. This difference is shown in Figure 3C where the M. m. domesticus matrix is subtracted from that of M. m. musculus. The average matrix in Figure 3D is a representative matrix with individuals averaged over the two subspecies.
Sexually Dimorphic Mups in the Liver, and Lacrimal Mup4 and Obp7
Mean value of log2 expression levels from the whole data set and standard deviation was 0.759 ± 1.154. Therefore, we opted for a two-fold (~2 sd) filtering procedure to obtain data with elevated sexual dimorphisms. To our surprise and probably due to the limited sample size, we have detected only three sexually dimorphic genes: the male-biased group-B Mups (i.e., a group of highly similar genes amplified with the primers provided in Table 1) in the liver, and unique male-biased lacrimal group-A Mup4, and female-biased lacrimal Obp7. Only these three groups were further tested.
To control for pseudoreplication of the expression data (i.e., three measurements taken from each individual with the total of 36 measurements) we used a mixed-effect model approach (nlme package) assuming normal distribution of the dependent variable, with individuals as random grouping variable (i.e., 12 clusters), and sex, gene, and species as fixed effects. Based on the minimum adequate model (Crawley, 2007) the level of detected sexual dimorphism was highly significant Δdf = 9, L-Ratio = 102.1, p < 0.0001. Post-hoc comparison with Tukey HSD further revealed how each gene contributed to significant sexual dimorphism: Mus m. musculus—Mup-B (p = 0.002), Mup4 (p = 0.0002), Obp7 (p = 0.09 ns); Mus m. domesticus—Mup-B (p = 0.009), Mup4 (p = 0.008), Obp7 (p = 0.005). The data and details of the model are provided in Data Sheets 2, 3. Additionally, we did not detect any significant sex-differences in the expression of lipocalins between M. m. musculus and M. m. domesticus.
Biochemical Properties of Co-Expressed OBPs and MUPs
Our bioinformatics analysis revealed that OBPs and MUPs have different predicted isoelectric points with MUPs being more acidic than OBPs (2-tailed t-test, p = 0.0009; Figure 4), which differentially affects their solubility at different pH. Data are provided in Data Sheet 4.
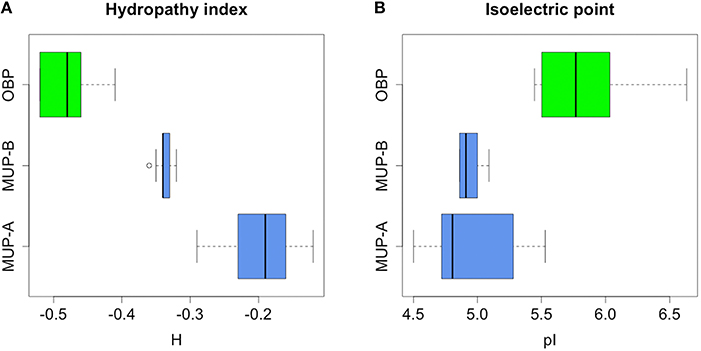
Figure 4. Bioinformatic analysis of hydropathy properties of MUPs and OBPs. Group-A MUPs, and OBPs have non-overlapping hydropathy properties. Interestingly, excretory group-B MUPs adopted a position between more hydropathic group-A MUPs (i.e., with more hydrophobic beta barrel) and less hydrophobic OBPs (A). Furthermore, MUPs have lower pI than OBPs (B).
Instead of looking at particular residues, we searched for a more general parameter that along with the structure directly affects lipocalin-binding properties. As a proxy, we calculated the grand average of hydropathy (i.e., GRAVY values, Xiong et al., 2009) which is defined as the sum of hydropathy values of all amino acids divided by the protein length. The values are negative for all individual members of MUPs and OBPs, however, the spectra of predicted hydrophobicities in OBPs and MUPs are non-overlapping (ANOVA, F = 54.59, p < 0.0001; Fligner-Killeen test of homogeneity of variances (chi-squared = 3.474, df = 1, p-value = 0.06234).
Furthermore, we have separated the group of MUPs based on previous studies (Logan et al., 2008; Mudge et al., 2008) into the ancestral group-A genes and the later duplicated group-B genes. Statistically significant differences in their mean values are graphically represented in Figure 4 by non-overlapping confidence intervals. Furthermore, the group-B MUPs evolved hydropathic properties that are intermediate between the group-A MUPs and OBPs.
Discussion
This study attempts to extend the current knowledge on genes for olfactory signals in feral mice by measuring selected mRNA expression across tissues, thus, revealing which other glands and tissues may be involved in chemical communication. We have corroborated that the group-B Mup transcripts show sexual dimorphisms in the liver in both subspecies (Stopková et al., 2007) whereby males excrete higher levels of Mups than females. The most interesting result of the current study is a sexually dimorphic pattern of the highly expressed ancestral group-A Mup4 and the female-biased Obp7 in lacrimal glands. Tears containing MUP4 and OBP7 are presumably spread onto the fur during a course of facial self-grooming, thus contributing to body odor with hydrophobic ligands that these proteins may transport. Nasal MUP4, however, was suggested to play roles in sequestering pheromones and possibly transporting them to their receptors (Sharrow et al., 2002).
The exocrine roles of lacrimal lipocalins are supported by observations that facial areas elicited strong neuronal activity response in the accessory olfactory bulb (Luo et al., 2003). Similarly Obp5, Obp6, Obp7 are also highly expressed in lacrimal glands (Obp5 and Obp7 originally annotated in inbred C57BL/6 as Obp1a and Obp1b). Recently, OBP proteins have been identified in tears of the laboratory mouse C57BL/6 (Karn and Laukaitis, 2015). In this study, Obp transcripts seem to be co-expressed in combination with other lipocalins (e.g., nasal and lacrimal Obp5, Obp7, Mup4, and Lcn11). OBP proteins (OBP5 and OBP7) were also predicted to form hetero-dimers (Pes et al., 1992) which may explain why Obp5 and Obp7 expression levels cluster together (Figure 3). Additionally, lacrimal expression of Obp7 is female biased and thus, may have female specific roles in chemical communication.
Evaluation of mRNA distribution across tissues also revealed that some lipocalin genes are expressed in just one tissue. Similarly to the group-B Mup genes that are mostly expressed in the liver, Obp6 is expressed only in lacrimal glands, and Lcn4 is expressed almost exclusively in the vomeronasal organ where LCN4 protein is covering the vomeronasal sensory epithelium to enable primary reception of pheromones (Miyawaki et al., 1994). It is therefore likely, that LCN4 together with Vomeromodulin (Khew-Goodall et al., 1991) and MUP4 (Sharrow et al., 2002) participate in the process of pheromone access and detection by VNO. Following the process of pheromone detection, some of these proteins (LCN3, LCN4, VM, OBPs) are presumably transported to the oral cavity where they are often detected in saliva of C57BL/6 mice (Blanchard et al., 2015; Karn and Laukaitis, 2015) but, as we found in this study, their mRNAs are produced elsewhere (i.e., mainly VNO).
Obp and Mup (or Lcn) genes are co-expressed in particular tissues probably because their proteins have non-overlapping ligand-binding properties (Cavaggioni et al., 1990) with MUPs having higher and OBPs lower number of hydrophobic residues. This has been originally reported for two OBPs in inbred mice (Cavaggioni et al., 1987; Pes et al., 1992; Pelosi, 1994) and extended for newly detected OBPs in feral mice in this study. Therefore, we also suggest that co-expressed lipocalins may have complementary functions where MUPs may transport more hydrophobic volatiles to and from the vicinity of olfactory receptors whilst OBPs may transport less hydrophobic ligands or may play roles in the deactivation of partially degraded non-specific (i.e., less hydrophobic = hydroxylated or oxidized) volatiles after the signal transduction (Strotmann and Breer, 2011). Our analysis plots in Figure 4 support such dual functionality. MUPs and OBPs have different pI and therefore may be active under different pH. We have already suggested that this difference in pI may imply that MUPs and OBPs have differential activities during cyclic (de-) acidification of nasal mucosa during ventilation (Stopková et al., 2014) similarly as in the study by Cichy et al. (2015) who provided evidence that extracellular pH regulates excitability of vomeronasal sensory neurons. Also, the acidification balance is maintained by Carbonic anhydrase IV (CA IV) which is secreted by salivary, lacrimal, and nasal glands (Kimoto et al., 2004).
The importance of MUPs and OBPs for general olfaction has previously been reported by Sharrow et al. (2002) who analyzed binding properties of nasal Mup4, and by Utsumi et al. (1999) who provided evidence that the expression of nasal Mup (i.e., most likely Mup5) and Obp genes is high. Furthermore, many species do not have multiple copies of Mup genes and thus MUP products—a major component of chemical signaling and olfaction in mice and rats—but express functional OBPs. This has been shown in many mammals (Singer and Macrides, 1990; Stopkova et al., 2010; Hagemeyer et al., 2011; Nagnan-Le Meillour et al., 2014) and it is our hope that potentially diverse functions—i.e., including the detoxification roles (Stopková et al., 2009; Kwak et al., 2011, 2016)—of these proteins will be further resolved.
Conclusions
We have studied the expression of known and newly described mRNAs coding for nasal, lacrimal, salivary and urinary lipocalins that are characteristic for their unusually high quantities and the capacity to bind pheromones in their beta barrel. Many of these proteins were individually reported in previous studies by various authors. However, we have provided evidence that some proteins found in saliva are produced by multiple tissues with the normalized expression levels being as high or higher as those described for the urinary group-B Mup genes in the liver. For the first time, we have described a novel cluster of odorant binding proteins in feral mice and shown that some of them are differentially expressed in tissues or are sexually dimorphic. Some lipocalins (OBP, MUP, LCN) are co-expressed probably to widen the spectrum of potential ligands that these proteins may sequester and transport. Such expression pattern is almost identical in the two studied subspecies of the house mouse with the exception of VNO, which shows higher lipocalin expression in M. m. musculus males. Moreover, further study with sufficient sample sizes could further reveal the level of variation between different individuals and species.
Author Contributions
RS performed qPCR analyses and wrote the manuscript. DV and JS were involved in transcript sequencing and qPCR analyses. BK did the bioinformatics analysis of OBP properties. OŠ and ZZ did the MS analyses. TA did most of the statistical analyses. KD was involved in all dissections and sample preparations. PS was involved in designing the experiment, in proteomic analyses and in writing the ms. All authors participated in writing the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The research of PS, RS, KD, DV, and JS was supported by the Grant Agency of the Czech Republic GACR No. P506/12/1046, and by the project “BIOCEV—Biotechnology and Biomedicine Centre of the Academy of Sciences and Charles University in Prague” (CZ.1.05/1.1.00/02.0109), from the European Regional Development Fund. BD was supported from a student grant SVV 260 313 / 2016 from Charles University in Prague. The MALDI-MS/MS part of the work (OŠ a ZZ) was carried out with the support of Proteomics Core Facility of CEITEC – Central European Institute of Technology, ID number CZ.1.05/1.1.00/02.0068, financed from European Regional Development Fund and by the Grant Agency of the Czech Republic GACR No. P206/12/G151.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/article/10.3389/fevo.2016.00047
Data Sheet 1. Protein identification and destaining protocol.
Data Sheet 2. Multiple sequence alignment of OBPs.
Data Sheet 3. Original data for statistical analyses.
Data Sheet 4. Details of the statistical model.
References
Bignetti, E., Cavaggioni, A., Pelosi, P., Persaud, K. C., Sorbi, R. T., and Tirindelli, R. (1985). Purification and characterisation of an odorant-binding protein from cow nasal tissue. Eur. J. Biochem. 149, 227–231. doi: 10.1111/j.1432-1033.1985.tb08916.x
Blanchard, A. A., Ezzati, P., Shamshurin, D., Nistor, A. C., Leygue, E., Wilkins, J. A., et al. (2015). Towards further defining the proteome of mouse saliva. Proteome Sci. 13, 10. doi: 10.1186/s12953-015-0068-3
Buck, L., and Axel, R. (1991). A novell multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell 65, 175–187. doi: 10.1016/0092-8674(91)90418-X
Cavaggioni, A., Findlay, J. B., and Tirindelli, R. (1990). Ligand binding characteristics of homologous rat and mouse urinary proteins and pyrazine-binding protein of calf. Comp. Biochem. Physiol. B 96, 513–520. doi: 10.1016/0305-0491(90)90049-y
Cavaggioni, A., Mucignat, C., and Tirindelli, R. (1999). Pheromone signalling in the mouse: role of urinary proteins and vomeronasal organ. Arch. Ital. Biol. 137, 193–200.
Cavaggioni, A., Sorbi, R. T., Keen, J. N., Pappin, D. J. C., and Findlay, J. B. C. (1987). Homology between the pyrazine-binding protein form nasal mucosa and major urinary proteins. FEBS Lett. 212, 225–228. doi: 10.1016/0014-5793(87)81349-2
Cichy, A., Ackels, T., Tsitoura, C., Kahan, A., Gronloh, N., Söchtig, M., et al. (2015). Extracellular pH regulates excitability of vomeronasal sensory neurons. J. Neurosci. 35, 4025–4039. doi: 10.1523/JNEUROSCI.2593-14.2015
Cunningham, C. B., Nelson, A. C., Ruff, J. S., and Potts, W. K. (2013). MUP expression is linked with sociality not competitive ability in male house mice. Integr. Comp. Biol. 53, E46.
Dureje, L., Bímová-Voslajerova, B., and Piálek, J. (2011). No postnatal maternal effect on male aggressiveness in wild-derived strains of house mice. Aggress. Behav. 37, 48–55. doi: 10.1002/ab.20371
Felicioli, A., Ganni, M., Garibotti, M., and Pelosi, P. (1993). Multiple types and forms of odorant-binding proteins in the Old-World porcupine Hystrix cristata. Comp. Biochem. Physiol. B 105, 775–784. doi: 10.1016/0305-0491(93)90119-p
Flower, D. (1996). The lipocalin protein family: structure and function. Biochem. J. 318, 1–14. doi: 10.1042/bj3180001
Fluckinger, M., Haas, H., Merschak, P., Glasgow, B. J., and Redl, B. (2004). Human tear lipocalin exhibits antimicrobial activity by scavenging microbial siderophores. Antimicrob. Agents Chemother. 48, 3367–3372. doi: 10.1128/AAC.48.9.3367-3372.2004
Hagemeyer, P., Begall, S., Janotova, K., Todrank, J., Heth, G., Jedelsky, P. L., et al. (2011). Searching for major urinary proteins (MUPs) as chemosignals in urine of subterranean rodents. J. Chem. Ecol. 37, 687–694. doi: 10.1007/s10886-011-9971-y
Hiadlovska, Z., Bimova, B. V., Mikula, O., Pialek, J., and Macholan, M. (2013). Transgressive segregation in a behavioural trait? Explorative strategies in two house mouse subspecies and their hybrids. Biol. J. Linnean Soc. 108, 225–235. doi: 10.1111/j.1095-8312.2012.01997.x
Hurst, J. L., Payne, C. E., Nevison, C. M., Marie, A. D., Humphries, R. E., Robertson, D. H. L., et al. (2001). Individual recognition in mice mediated by major urinary proteins. Nature 414, 631–634. doi: 10.1038/414631a
Hurst, J. L., Robertson, D. H. L., Tolladay, U., and Beynon, R. J. (1998). Proteins in urine scent marks of male house mice extend the longevity of olfactory signals. Anim. Behav. 55, 1289–1297. doi: 10.1006/anbe.1997.0650
Janotová, K., and Stopka, P. (2009). Mechanisms of chemical communication: the role of major urinary proteins. Folia Zool. 58, 41–55.
Janotova, K., and Stopka, P. (2011). The level of major urinary proteins is socially regulated in wild Mus musculus musculus. J. Chem. Ecol. 37, 647–656. doi: 10.1007/s10886-011-9966-8
Jones, D. T., Taylor, W. R., and Thornton, J. M. (1992). The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 8, 275–282. doi: 10.1093/bioinformatics/8.3.275
Karn, R. C., and Laukaitis, C. M. (2011). Positive selection shaped the convergent evolution of independently expanded kallikrein subfamilies expressed in mouse and rat saliva proteomes. PLoS ONE 6:e20979. doi: 10.1371/journal.pone.0020979
Karn, R. C., and Laukaitis, C. M. (2012). The roles of gene duplication, gene conversion and positive selection in rodent Esp and Mup pheromone gene families with comparison to the Abp family. PLoS ONE 7:e47697. doi: 10.1371/journal.pone.0047697
Karn, R., and Laukaitis, C. (2015). Comparative proteomics of mouse tears and saliva: evidence from large protein families for functional adaptation. Proteomes 3, 283. doi: 10.3390/proteomes3030283
Khew-Goodall, Y., Grillo, M., Getchell, M. L., Danho, W., Getchell, T. V., and Margolis, F. L. (1991). Vomeromodulin, a putative pheromone transporter: cloning, characterization, and cellular localization of a novel glycoprotein of lateral nasal gland. FASEB J. 5, 2976–2982.
Kimoto, M., Iwai, S., Maeda, T., Yura, Y., Fernley, R. T., and Ogawa, Y. (2004). Carbonic anhydrase VI in the mouse nasal gland. J. Histochem. Cytochem. 52, 1057–1062. doi: 10.1369/jhc.3A6243.2004
Kiyono, H., and Fukuyama, S. (2004). NALT- versus Peyer's-patch-mediated mucosal immunity. Nat. Rev. Immunol. 4, 699–710. doi: 10.1038/nri1439
Kwak, J., Grigsby, C. C., Preti, G., Rizki, M. M., Yamazaki, K., and Beauchamp, G. K. (2013). Changes in volatile compounds of mouse urine as it ages: their interactions with water and urinary proteins. Physiol. Behav. 120, 211–219. doi: 10.1016/j.physbeh.2013.08.011
Kwak, J., Josue, J., Faranda, A., Opiekun, M. C., Preti, G., Osada, K., et al. (2011). Butylated hydroxytoluene is a ligand of urinary proteins derived from female mice. Chem. Senses 36, 443–452. doi: 10.1093/chemse/bjr015
Kwak, J., Strasser, E., Luzynski, K., Thoß, M., and Penn, D. J. (2016). Are MUPs a toxic waste disposal system? PLoS ONE 11:e0151474. doi: 10.1371/journal.pone.0151474
Lazar, J., Greenwood, D. R., Rasmussen, L. E. L., and Prestwich, G. D. (2002). Molecular and functional characterization of an odorant binding protein of the asian elephant, elephas maximus: implications for the role of lipocalins in mammalian olfaction. Biochemistry 41, 11786–11794. doi: 10.1021/bi0256734
Logan, D. W., Marton, T. F., and Stowers, L. (2008). Species specificity in major urinary proteins by parallel evolution. PLoS ONE 3:e3280. doi: 10.1371/journal.pone.0003280
Luo, M., Fee, M. S., and Katz, L. C. (2003). Encoding pheromonal signals in the accessory olfactory bulb of behaving mice. Science 299, 1196–1201. doi: 10.1126/science.1082133
Luo, M., and Katz, L. C. (2004). Encoding pheromonal signals in the mammalian vomeronasal system. Curr. Opin. Neurobiol. 2004, 428–434. doi: 10.1016/j.conb.2004.07.001
Marchese, S., Pes, D., Scaloni, A., Carbone, V., and Pelosi, P. (1998). Lipocalins of boar salivary glands binding odours and pheromones. Eur. J. Biochem. 252, 563–568. doi: 10.1046/j.1432-1327.1998.2520563.x
Miyawaki, A., Matsushita, F., Ryo, Y., and Mikoshiba, K. (1994). Possible pheromone-carrier function of two lipocalin proteins in the vomeronasal organ. EMBO J. 13, 5835–5842.
Moss, R. L., Flynn, R. E., Shen, X. M., Dudley, C., Shi, J., and Novotny, M. (1997). Urine-derived compound evokes membrane responses in mouse vomeronasal receptor neurons. J. Neurophysiol. 77, 2856–2862.
Mucignat-Caretta, C., Redaelli, M., Orsetti, A., Perriat-Sanguinet, M., Zagotto, G., and Ganem, G. (2010). Urinary volatile molecules vary in males of the 2 European subspecies of the house mouse and their hybrids. Chem. Senses 35, 647–654. doi: 10.1093/chemse/bjq049
Mudge, J. M., Armstrong, S. D., McLaren, K., Beynon, R. J., Hurst, J. L., Nicholson, C., et al. (2008). Dynamic instability of the major urinary protein gene family revealed by genomic and phenotypic comparisons between C57 and 129 strain mice. Genome Biol. 9, R91. doi: 10.1186/gb-2008-9-5-r91
Nagnan-Le Meillour, P., Vercoutter-Edouart, A. S., Hilliou, F., Le Danvic, C., and Lévy, F. (2014). Proteomic analysis of pig (Sus scrofa) olfactory soluble proteome reveals o-linked-n-acetylglucosaminylation of secreted odorant-binding proteins. Front. Endocrinol. 5:202. doi: 10.3389/fendo.2014.00202
Nelson, A. C., Cauceglia, J. W., Merkley, S. D., Youngson, N. A., Oler, A. J., Nelson, R. J., et al. (2013). Reintroducing domesticated wild mice to sociality induces adaptive transgenerational effects on MUP expression. Proc. Natl. Acad. Sci. U.S.A. 110, 19848–19853. doi: 10.1073/pnas.1310427110
Novotny, M. V. (2003). Pheromones, binding proteins and receptor responses in rodents. Biochem. Soc. 31, 117–122. doi: 10.1042/bst0310117
Novotny, M., Harvey, S., Jemiolo, B., and Alberts, J. (1985). Synthetic pheromones that promote inter-male aggression in mice. Proc. Natl. Acad. Sci. U.S.A. 82, 2059–2061. doi: 10.1073/pnas.82.7.2059
Pelosi, P. (1994). Odorant-binding proteins: structural aspects. Crit. Rev. Biochem. Mol. Biol. 29, 199–228. doi: 10.3109/10409239409086801
Pes, D., Dal Monte, M., Ganni, M., and Pelosi, P. (1992). Isolation of two odorant-binding proteins from mouse nasal tissue. Comp. Biochem. Physiol. 103B, 1011–1017. doi: 10.1016/0305-0491(92)90231-f
Pfaffl, M. W. (2001). A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29, e45. doi: 10.1093/nar/29.9.e45
Phelan, M. M., McLean, L., Hurst, J. L., Beynon, R. J., and Lian, L. Y. (2014). Comparative study of the molecular variation between ‘central’ and ‘peripheral’ MUPs and significance for behavioural signalling. Biochem. Soc. Trans. 42, 866–872. doi: 10.1042/BST20140082
Roberts, S. C., Havlicek, J., Flegr, J., Hruskova, M., Littie, A. C., Jones, B. C., et al. (2004). Female factial attractiveness increases during the fertile phase of the menstrual cycle. Proc. R. Soc. Lond. B 271, S270–S272. doi: 10.1098/rsbl.2004.0174
Roberts, S. A., Davidson, A. J., McLean, L., Beynon, R. J., and Hurst, J. L. (2012). Pheromonal induction of spatial learning in mice. Science 338, 1462–1465. doi: 10.1126/science.1225638
Roberts, S. A., Simpson, D. M., Armstrong, S. D., Davidson, A. J., Robertson, D. H., McLean, L., et al. (2010). Darcin: a male pheromone that stimulates female memory and sexual attraction to an individual male's odour. BMC Biol. 8:75. doi: 10.1186/1741-7007-8-75
Shahan, K., Denaro, M., Gilmartin, M., Shi, Y., and Derman, E. (1987a). Expression of six mouse major urinary protein genes in the mammary, parotid, sublingual, submaxillary, and lachrymal glands and in the liver. Mol. Cell. Biol. 7, 1947–1954.
Shahan, K., and Derman, E. (1984). Tissue-specific expression of major urinary protein (MUP) genes in mice: characterization of MUP mRNAs by restriction mapping of cDNA and by in vitro translation. Mol. Cell. Biol. 4, 2259–2265. doi: 10.1128/MCB.4.11.2259
Shahan, K., Gilmartin, M., and Derman, E. (1987b). Nucleotide sequences of liver, lachrymal, and submaxillary gland mouse major urinary protein mRNAs: mosaic structure and construction of panels of gene-specific synthetic oligonucleotide probes. Mol. Cell. Biol. 7, 1938–1946.
Sharrow, S. D., Vaughn, J. L., Žídek, L., Novotny, M. V., and Stone, M. J. (2002). Pheromone binding by polymorphic mouse major urinary proteins. Protein Sci. 11, 2247–2256. doi: 10.1110/ps.0204202
Shaw, P. H., Held, W. A., and Hastie, N. D. (1983). The gene family for major urinary proteins: expression in several secretory tissues of the mouse. Cell 32, 755–761. doi: 10.1016/0092-8674(83)90061-2
Smadja, C., and Ganem, G. (2002). Subspecies recognition in the house mouse: a study of two populations from the border of a hybrid zone. Behav. Ecol. 13, 312–320. doi: 10.1093/beheco/13.3.312
Smadja, C., and Ganem, G. (2008). Divergence of odorant signals within and between the two European subspecies of the house mouse. Behav. Ecol. 19, 223–230. doi: 10.1093/beheco/arm127
Spinelli, S., Ramoni, R., Grolli, S., Bonicel, J., Cambillau, C., and Tegoni, M. (1998). The structure of the monomeric porcine odorant binding protein sheds light on the domain swapping mechanism. Biochemistry 37, 7913–7918. doi: 10.1021/bi980179e
Stopka, P., Janotova, K., and Heyrovsky, D. (2007). The advertisement role of major urinary proteins in mice. Physiol. Behav. 91, 667–670. doi: 10.1016/j.physbeh.2007.03.030
Stopková, R., Hladovcová, D. J. K., Vyoral, D., and Stopka, P. (2009). Multiple roles of secretory lipocalins (MUP, OBP) in mice. Folia Zool. 58, 29–40.
Stopková, R., Stopka, P., Janotová, K., and Jedelský, P. L. (2007). Species-specific expression of major urinary proteins in the house mice (Mus musculus musculus and Mus musculus domesticus). J. Chem. Ecol. 33, 861–869. doi: 10.1007/s10886-007-9262-9
Stopková, R., Dudková, B., Hájková, P., and Stopka, P. (2014). Complementary roles of mouse lipocalins in chemical communication and immunity. Biochem. Soc. Trans. 42, 893–898. doi: 10.1042/BST20140053
Stopkova, R., Zdrahal, Z., Ryba, S., Šedo, O., Sandera, M., and Stopka, P. (2010). Novel OBP genes similar to hamster Aphrodisin in the bank vole, Myodes glareolus. BMC Genomics 11:45. doi: 10.1186/1471-2164-11-45
Strotmann, J., and Breer, H. (2011). Internalization of odorant-binding proteins into the mouse olfactory epithelium. Histochem. Cell Biol. 136, 357–369. doi: 10.1007/s00418-011-0850-y
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Thoß, M., Luzynski, K., Ante, M., Miller, I., and Penn, D. J. (2015). Major urinary protein (MUP) profiles show dynamic changes rather than individual ‘barcode’ signatures. Front. Evol. 3:71. doi: 10.3389/fevo.2015.00071
Thonhauser, K. E., Raveh, S., Hettyey, A., Beissmann, H., and Penn, D. J. (2013). Scent marking increases male reproductive success in wild house mice. Anim. Behav. 86, 1013–1021. doi: 10.1016/j.anbehav.2013.09.004
Timm, D. E., Baker, L. J., Mueller, H., Zidek, L., and Novotny, M. V. (2001). Structural basis of pheromone binding to mouse major urinary protein (MUP-I). Protein Sci. 10, 997–1004. doi: 10.1110/ps.52201
Tirindelli, R., Mucignat-Caretta, C., and Ryba, N. J. P. (1998). Molecular aspects of pheromonal communication via the vomeronasal organ of mammals. Trends Neurosci. 21, 482–486. doi: 10.1016/S0166-2236(98)01274-0
Utsumi, M., Ohno, K., Kawasaki, Y., Tamura, M., Kubo, T., and Tohyama, M. (1999). Expression of major urinary protein genes in the nasal glands associated with general olfaction. J. Neurobiol. 39, 227–236.
Venables, W. N., and Smith, D. M. (2009). An Introduction to R: A Programming Environment for Data Analysis and Graphics, Version 2.9.0. Bristol, CT: Network Theory.
Whitten, W. K., Bronson, F. H., and Greenstein, J. A. (1968). Estrus-inducing pheromone of male mice: transport by movement of air. Science 161, 584–585. doi: 10.1126/science.161.3841.584
Keywords: lipocalin, odorant, chemical communication, Mus musculus, olfaction
Citation: Stopková R, Vinkler D, Kuntová B, Šedo O, Albrecht T, Suchan J, Dvořáková-Hortová K, Zdráhal Z and Stopka P (2016) Mouse Lipocalins (MUP, OBP, LCN) Are Co-expressed in Tissues Involved in Chemical Communication. Front. Ecol. Evol. 4:47. doi: 10.3389/fevo.2016.00047
Received: 26 February 2016; Accepted: 15 April 2016;
Published: 28 April 2016.
Edited by:
Carla Mucignat, University of Padova, ItalyReviewed by:
Patricia Nagnan-Le Meillour, Institut National de la Recherche Agronomique, FranceDustin J. Penn, Veterinary University Vienna, Austria
Roberto Ramoni, University of Parma, Italy
Copyright © 2016 Stopková, Vinkler, Kuntová, Šedo, Albrecht, Suchan, Dvořáková-Hortová, Zdráhal and Stopka. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pavel Stopka, pstopka@natur.cuni.cz