 Loan T. Nguyen1,2
Loan T. Nguyen1,2 Antonio Reverter3
Antonio Reverter3 Angela Cánovas4
Angela Cánovas4 Bronwyn Venus5
Bronwyn Venus5 Stephen T. Anderson6Alma Islas-Trejo7Marina M. Dias8Natalie F. Crawford9
Stephen T. Anderson6Alma Islas-Trejo7Marina M. Dias8Natalie F. Crawford9 Sigrid A. Lehnert3
Sigrid A. Lehnert3 Juan F. Medrano7
Juan F. Medrano7 Milt G. Thomas9
Milt G. Thomas9 Stephen S. Moore5
Stephen S. Moore5 Marina R. S. Fortes1,5*
Marina R. S. Fortes1,5*- 1School of Chemistry and Molecular Biosciences, The University of Queensland, St. Lucia, QLD, Australia
- 2Faculty of Biotechnology, Vietnam National University of Agriculture, Hanoi, Vietnam
- 3CSIRO Agriculture and Food, Queensland Bioscience Precinct, St. Lucia, QLD, Australia
- 4Centre for Genetic Improvement of Livestock, Department of Animal Biosciences, University of Guelph, Guelph, ON, Canada
- 5Queensland Alliance for Agriculture and Food Innovation, The University of Queensland, St. Lucia, QLD, Australia
- 6School of Biomedical Sciences, The University of Queensland, Brisbane, QLD, Australia
- 7Department of Animal Science, University of California, Davis, Davis, CA, United States
- 8Departamento de Zootecnia, Faculdade de Ciências Agráìrias e Veterináìrias, Universidade Estadual Paulista Júlio de Mesquita Filho, São Paulo, Brazil
- 9Department of Animal Science, Colorado State University, Fort Collins, CO, United States
The liver plays a central role in metabolism and produces important hormones. Hepatic estrogen receptors and the release of insulin-like growth factor 1 (IGF1) are critical links between liver function and the reproductive system. However, the role of liver in pubertal development is not fully understood. To explore this question, we applied transcriptomic analyses to liver samples of pre- and post-pubertal Brahman heifers and identified differentially expressed (DE) genes and genes encoding transcription factors (TFs). Differential expression of genes suggests potential biological mechanisms and pathways linking liver function to puberty. The analyses identified 452 DE genes and 82 TF with significant contribution to differential gene expression by using a regulatory impact factor metric. Brain-derived neurotrophic factor was observed as the most down-regulated gene (P = 0.003) in post-pubertal heifers and we propose this gene influences pubertal development in Brahman heifers. Additionally, co-expression network analysis provided evidence for three TF as key regulators of liver function during pubertal development: the signal transducer and activator of transcription 6, PBX homeobox 2, and polybromo 1. Pathway enrichment analysis identified transforming growth factor-beta and Wnt signaling pathways as significant annotation terms for the list of DE genes and TF in the co-expression network. Molecular information regarding genes and pathways described in this work are important to further our understanding of puberty onset in Brahman heifers.
Introduction
The beef industry in Northern Australia is facing increased demand for improved herd productivity. Brahman cattle, a breed of the Bos indicus sub-species, can withstand hot-humid conditions, but enter puberty at older age in comparison with Bos taurus (Johnston et al., 2009). Late onset of puberty in B. indicus predicts a decrease in lifetime productivity (Lesmeister et al., 1973; Johnston et al., 2013). Therefore, reducing the age at puberty to increase B. indicus cow productivity is a worthwhile goal for management and breeding.
Reproduction is an energy intensive process that is likely to require specific involvement of the liver. The physiological mechanisms controlling energy balance are closely linked to fertility, to minimize the risk that pregnancy and lactation coincide with periods of nutritional stress (Mircea et al., 2007). While in placental mammals, the hypothalamus–pituitary–ovaries axis takes precedence in the integration of metabolic and reproductive status, there are evidences for the involvement of the liver in this process. In the postpartum cow, it is known that the metabolic stress associated with transition is linked to impaired liver function and delayed ovulation (Montagner et al., 2016). In mice, it has been shown that hepatic synthesis of Insulin-like growth factor 1 (IGF1) is regulated by amino acid-dependent activation of ERα in the liver (Della Torre et al., 2011). Additionally, an association between single nucleotide polymorphisms in genes of the IGF1 signaling pathways and age at puberty in Brahman cattle was observed (Fortes et al., 2013).
Several studies investigated the change in hepatic mRNA expression of genes encoding proteins that participate in various processes including growth hormone signaling, liver lipoprotein assembly, ureagenesis, and gluconeogenesis (Loor, 2010). Few studies have utilized microarray technology to evaluate hepatic metabolic adaptations to dairy cow throughout pregnancy, transition period, early lactation, and mid lactation (Herath et al., 2004; Loor et al., 2005; Loor, 2010; McCarthy et al., 2010; Akbar et al., 2013). A microarray study of the effect of pregnancy and diet in liver gene expression revealed specific hepatic adaptations of beef cows to different nutritional environments. For example, the study found clear evidence of gluconeogenesis in the liver of pregnant cows during limited forage availability (Laporta et al., 2014). Using candidate gene approach or transcriptomics investigating genomic peripartal adaptions in dairy cows provided insights into physiological function and genetics of key tissues.
A study has evaluated the liver transcriptome during puberty onset in Brangus heifers (3/8 Brahman; B. indicus × 5/8 Angus; B. taurus) (Cánovas et al., 2014). This study has used RNA sequencing, which is a more sensitive transcriptome profiling method than microarray. Sequencing RNA is capable of detecting not only expression differences in the most highly expressed metabolic genes, but also in regulatory genes (Marioni et al., 2008). Our study used sequencing to evaluated mRNA expression, regulatory factors, and potential biological pathways that occur in the liver related to pubertal development in Brahman heifers; a different population that is predominantly B. indicus. These heifers were used in two studies that reported transcriptomics of the hypothalamus–pituitary–ovarian axis, with no observation of liver function (Fortes et al., 2016; Nguyen et al., 2017). Molecular information of key regulators and pathways in liver may reveal mechanisms involved in puberty onset and energy metabolism. To access IGF1 signaling in this context, we also report hormonal measurements. This information may contribute to future approaches for reducing the age at puberty of B. indicus cattle used in tropical beef production systems.
Materials and Methods
Animals and Samples
Heifers used in this study were managed, handled, and euthanized as per approval of the Animal Ethics Committee of the University of Queensland, Production, and Companion Animal group (certificate number QAAFI/279/12). Heifer used were 12 young animals from commercial Queensland herds with typical phenotypic characteristics of B. indicus cattle. In Australia, the average content of B. indicus in Brahman cattle is about 95% (Porto Neto et al., 2013). They were unrelated heifers of similar age (born during the wet season 2011/2012) and weight <250 kg. They were maintained on pasture at the Gatton Campus facilities of the University of Queensland.
We performed ultrasound observations of pubertal development every fortnight from October 2012 to May 2013. With ultrasound, the observation of the first corpus luteum (CL) was used to define pubertal status (Johnston et al., 2009). Euthanasia occurred 15 days after the observation of the first CL, with samples collected in the next estrous cycle. Six post-pubertal heifers were euthanized during the luteal phase of their second estrous cycle, confirmed by the observation of the second CL at euthanasia. Serum progesterone concentrations were measured to confirm a functional CL in post-pubertal heifers (2.0 ± 0.7 ng/mL, mean ± SE). Pre-puberty heifers were randomly selected from the group that had never ovulated (plasma progesterone concentration 0.4 ± 0.2 ng/mL, mean ± SE) and paired with post-pubertal animals in slaughter day. Therefore, on each slaughter day, two heifers were euthanatized, one pre- and one post-puberty.
Serum progesterone concentrations were measured in hexane extracts by RIA (Curlewis et al., 1985) at the Laboratory for Animal Endocrinology at the University of Queensland. The extraction efficiency was 75% and reported values were not corrected for these losses. The assay sensitivity was 0.1 ng/mL and the within-assay coefficient of variation was 5.0%.
Circulating IGF1 concentrations were measured using a commercial radioimmunoassay kit (10IGF100 Kit; Bioclone, Sydney, NSW, Australia). The method included an acid–ethanol extraction to remove IGF1-binding proteins and measure total IGF1. All samples were analyzed within a single assay kit as previously described (Dahlanuddin et al., 2014). The assay sensitivity was 0.2 ng/mL and the within-assay coefficient of variation was 2.5%.
Body weight (BW, kg) and body condition scores (BCSs, 5-point scale) were also measured before tissue harvesting, as previously described (Fortes et al., 2016). BWs were 338 ± 54 and 363 ± 39 kg and BCSs were 3.5 ± 0.4 and 3.8 ± 0.4 for pre- and post-pubertal heifers, respectively. There was no statistical difference in an unpaired t-test in either BW (P = 0.38) or BCS (P = 0.18) between the heifer groups (Fortes et al., 2016).
After slaughter, tissue harvest was conducted as fast as possible to preserve RNA integrity. The entire liver was removed from the animal and three samples of 1 cm3 were dissected from the liver and snap frozen in liquid nitrogen. Samples were stored at -80°C until RNA extraction. In total, 12 liver samples were processed separately for RNA extraction and sequencing.
Ribonucleic Acid Extraction
Total RNA was isolated from fragmented frozen liver tissue (∼100 mg) as previously described by Nguyen et al. (2017). Quality of the total RNA was evaluated using the RNA integrity number (RIN) measured with an Agilent Bioanalyzer 2100 (Agilent Technologies). Values of RIN range from 7.3 to 8.5, which indicated good quality of the RNA samples, which were sent to the University of California, Davis, for library preparation and sequencing.
Library Preparation and Sequencing
mRNA was purified, fragmented, and used to synthesize cDNA, as described in (Cánovas et al., 2010). Briefly, after ligation of the adapters to the ends of double-stranded cDNA fragments, PCR was conducted to create the final cDNA library. Sequencing libraries were constructed with the TruSeq RNA sample preparation kit (Illumina, San Diego, CA, United States). RNA sequencing was conducted with a HiSeq 2000 Sequencer Analyzer (Illumina, San Diego, CA, United States). Quality control was performed using procedures described previously (Cánovas et al., 2013) using the application NGS quality control of CLC Bio Genomic workbench software (CLC Bio, Aarhus, Denmark). All samples passed all the parameters indicating a very good quality.
Sequence pair-end reads (100 bp) were assembled against the annotated bovine genome (release 771). The “reads per kilo base per million mapped reads” (RPKM = total exon reads/mapped reads in millions × exon length in kb) was calculated for data normalization (Mortazavi et al., 2008). A threshold of RPKM ≥ 0.2 was utilized to annotated expressed genes (Wickramasinghe et al., 2012). Normalization and transformation data were performed using CLCBio Genomic workbench software (CLC Bio, Aarhus, Denmark) to transform the expression data from negative binomial distribution to normal distribution.
Identification of Differentially Expressed Genes
Because genes with low counts can be easily biased without transformation, the base-2 log-transformed RPKM values were used. Mixed model equations are an optimal method for data normalization in gene expression studies (Reverter et al., 2005). We normalize the log-transformed RPKM values using mixed model equations to increase the sensitivity to detect differential expression and co-expression. This normalization approach for transformed RPKM values was previously described (Reverter et al., 2005; Cánovas et al., 2014). In more detail, differential gene expression after puberty was calculated using a mixed model: Y ijkpt = μ + Li + Gj + GAPTjkpt + eijkpt, where log 2-transformed RPKM (Yijkpt) was modeled as a function of the fixed effect of the i library (Li) and of the random effects of gene (Gj), and the interaction of gene × animal × physiological state × tissue (GAPTjkpt) for the i library (with 72 levels) and the j gene (with 16,978 levels) of the k animal (12 levels) in the p physiological state (with two levels) from the t tissue (with five levels). Finally, eijkpt represents the random residual term. Our liver study was part of a larger experiment where five tissues were sampled per animal (hypothalamus, pituitary, liver, ovaries, and uterus), which allows fitting for tissue in the model. The VCE6 software2 was used to assemble and solve the mixed model equations and to estimate variance components associated with random effects. For each gene, the normalized mean expression was obtained based on adding the solutions G + GAPT. A t-test was used to test the hypothesis that the differential expression in post- vs. pre-pubertal heifers was significant. With the strict normalization performed, we then used P < 0.05 as the threshold to determine DE genes. This seemingly not very stringent nominal P was used in context with the strict normalization performed and the subsequent analyses, for which differential expression is one of many criteria under scrutiny.
Identification of Key Gene Regulators
To determine gene regulators from the data, we mined the AnimalTFDB bovine database3, which comprises the classification and annotation of animal genomes for transcription factors (TFs), chromatin remodeling factors, and transcription co-factors. Among the annotated TF for B. taurus, 1,085 were expressed in the liver and further filtered for significance in terms of co-expression with DE genes, using regulatory impact factor (RIF) metrics (Hudson et al., 2009; Reverter et al., 2010). The RIF metric was explored in two measures: RIF1 and RIF2, calculated from the number of DE genes and the predicted interactions between TF and target DE genes (Reverter et al., 2010). In brief, RIF1 captured those TF that showed a large differential connectivity to highly abundant DE genes, whereas RIF2 focused on TF showing evidence as predictors of change in abundance of DE genes. A TF was considered as a key regulator if either of the two RIF scores was higher than 1.96 of the standard deviation, equivalent to a P-value level of at least 0.05.
Gene Network Prediction
The partial correlation and information theory (PCIT) algorithm was utilized to detect the association between genes in a co-expression gene network (Reverter and Chan, 2008). In brief, PCIT explores all pair-wise direct and partial correlations among all possible trios of genes before identifying significant correlations that will establish edges during network reconstruction. The co-expression network predicted for the liver data was then visualized with Cytoscape (Shannon et al., 2003). From the large predicted network, we explored the subnetwork deemed to have biological significance for puberty trait. The subnetwork was used to identify the best trio TF that spanned most of the network topology with minimum redundancy. Specifically, an information lossless approach (Reverter and Fortes, 2013) that explored the 59,640 possible trios among 82 available TFs in the network was used to identify the best TF trio.
Functional Enrichment Analysis
For enriched pathways and gene expression patterns, the Database for Annotation, Visualization, and Integrated Discovery (DAVID4) was utilized (Dennis et al., 2003; Huang da et al., 2009). In our study, the queried gene lists included genes and TF that formed the predicted gene network. This list of genes was utilized as a target gene list in comparison with a background gene list formed by all genes expressed in liver. Functional annotation chart revealed the most relevant (overrepresented) gene ontology terms and pathways associated with these gene lists, reporting an enrichment P-value for each annotation term. Significant results after Benjamini–Hochberg correction for multiple testing are reported.
Results
RNA-Seq Data and Normalization
The liver transcriptome data passed quality control performed with CLC Genomics workbench (CLC Bio, Aarhus, Denmark). Each individual sample had about 63 million sequence reads. Previous studies demonstrated that approximately 30 million reads are sufficient to detect more than 90% of annotated genes in mammalian genomes (Wang et al., 2011; Lee et al., 2013; Singh et al., 2017). The relatively high number of sequence reads generated in this transcriptome study indicates that our data are adequate for identification of DE genes. The number of unique reads and RPKM of each gene per physiological state are provided (Supplementary Table S1). Sequence data are available through the Functional Annotation of Animal Genomes project5.
Identification of Differentially Expressed Genes
A total of 16,978 transcripts were detected in both groups (pre- and post-puberty). A t-test of log-transformed data identified 452 DE genes (including 57 novel genes), of which 253 were up-regulated and 199 were down-regulated post-puberty (P < 0.05). Ten genes showed a threefold change (FC) difference in expression levels and P < 0.01 between pre- and post-puberty heifers (Table 1). Figure 1 shows a volcano plot of log 2 FC vs. –log10 P-values for pre- vs. post-puberty gene expression. The gene annotation, FC, and P-value of 452 DE genes are presented in Supplementary Table S2. Significant DE genes were useful for understanding the biological mechanisms in the liver underlying puberty onset in Brahman cattle.
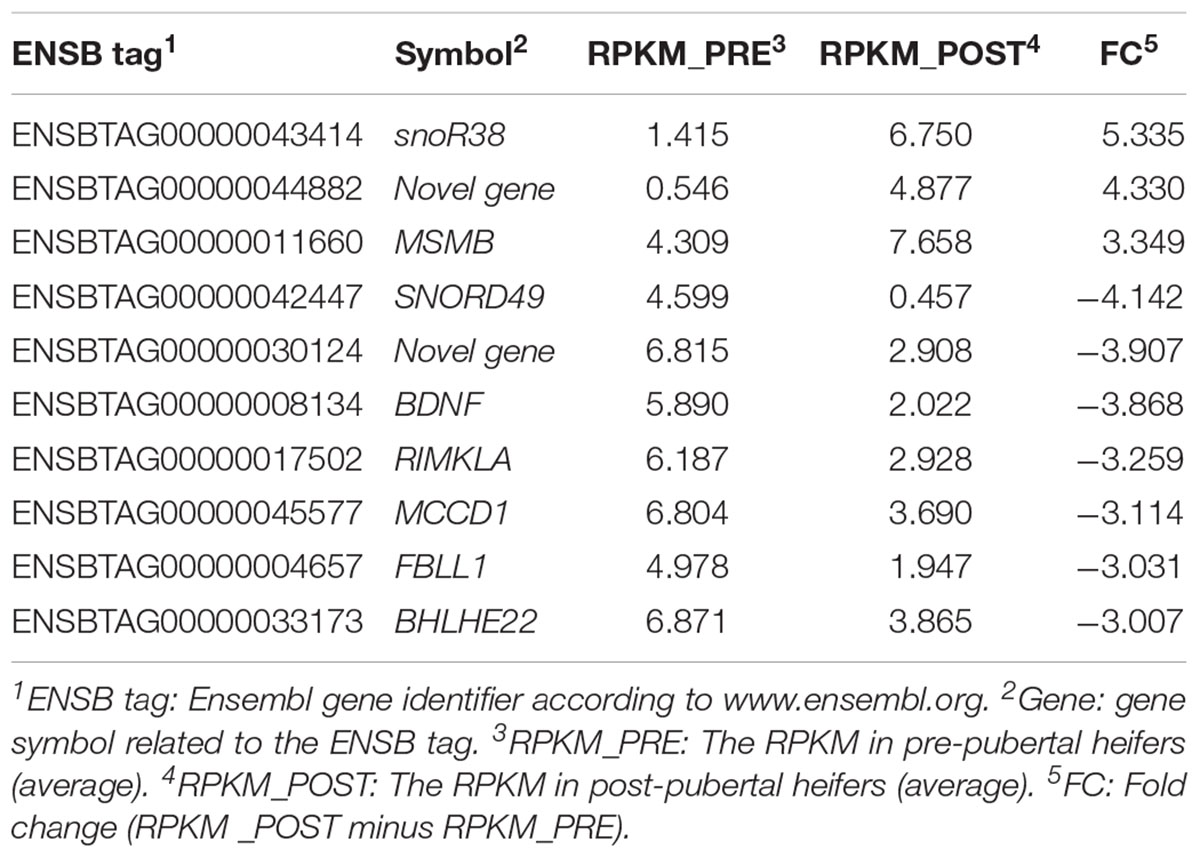
TABLE 1. The reads per kilobase per million (RPKM) mapped read values for genes that significantly differ in expression in liver between pre- vs. post-pubertal Brahman heifers (|FC|≥ 3, P ≤ 0.01).
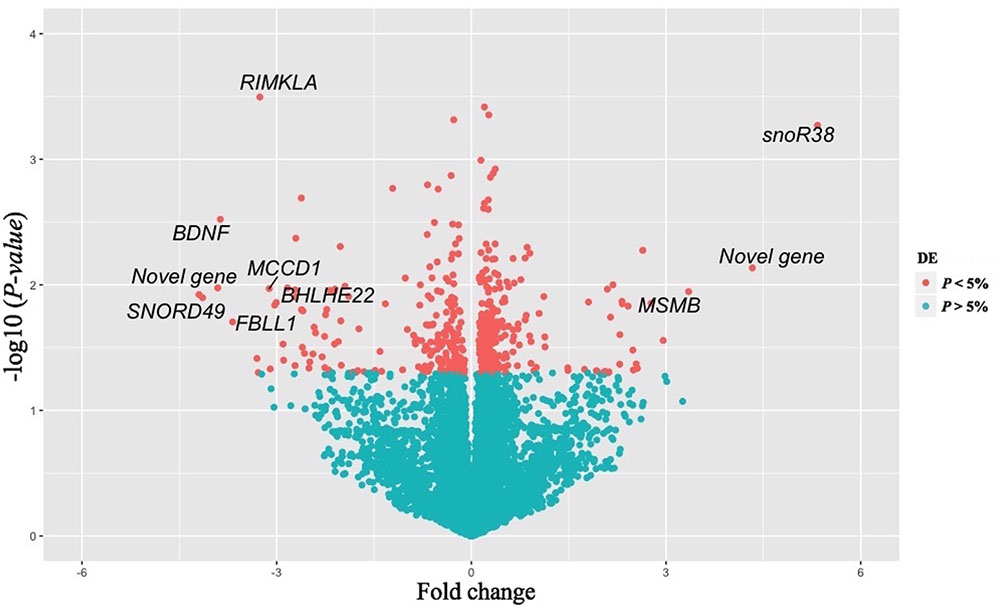
FIGURE 1. Volcano plot of differentially expressed genes (N = 452) in liver between pre- vs post-pubertal heifers. The x-axis represents the FC while the y-axis represents statistical significance for each gene. Red dots indicate genes that differ significantly (P < 0.05) between the two groups. Genes plotted in the left portion of the graph were expressed at a lower level in post-pubertal liver, and gene in the right-hand portion had higher expression levels post-puberty. Gene symbols are provided for genes with a |FC|≥ 3 and P ≤ 0.01.
Insulin-like growth factor 1 is the major hormone secreted by the liver and is known to increase during puberty. In our study, the circulating IGF1 concentrations differed between pre- and post-pubertal heifers at euthanasia (P = 0.008) with the average serum IGF1 levels were 159.3 ± 25.5 ng/mL at pre-puberty and 203.2 ± 31.1 ng/mL at post-pubertal heifers. Although, RNA-seq analysis showed increase in IGF-1 mRNA levels (2.01 ± 0.17 vs. 2.35 ± 0.19) after puberty in the liver, the result was not significant (P = 0.222).
Identification of Key Gene Regulators
From AnimalTFDB Bovine database, we retrieved 1,085 TF that were expressed in the liver. Using RIF metrics, these known TFs were filtered for those most consistently associated with DE genes from this study, identifying 82 TF (P < 0.05). Supplementary Table S3 summarizes relevant data for these TF:RIF results, expression levels, and its description. Of the 82 TF, 19 genes (23%) coded for TF of the zinc finger family (ZNF). Further, out of the 82 TF, 2 TF (SOX13 and BHLHE22) were themselves identified as DE genes.
Predicted Gene Co-expression Network and Sub-network
Partial correlation and information theory algorithm determined significant partial correlations between DE and TF. A predicted gene co-expression network with 1,408 nodes representing genes and a total of 8,330 edges which account for the predicted interactions was constructed (Figure 2). In order to identify potential regulators of the predicted gene co-expression network, we focused on 82 TF contained in the network. After exploring all the TF trios, the top trio which spanned most of the network topology with highest connectivity (a total of 59,640 possible connections) and minimum redundancy was identified, including the signal transducer and activator of transcription 6 (STAT6), PBX homeobox 2 (PBX2), and polybromo 1 (PBRM1). Figure 3 presents the connections between the top trio of TF and their potential targets.
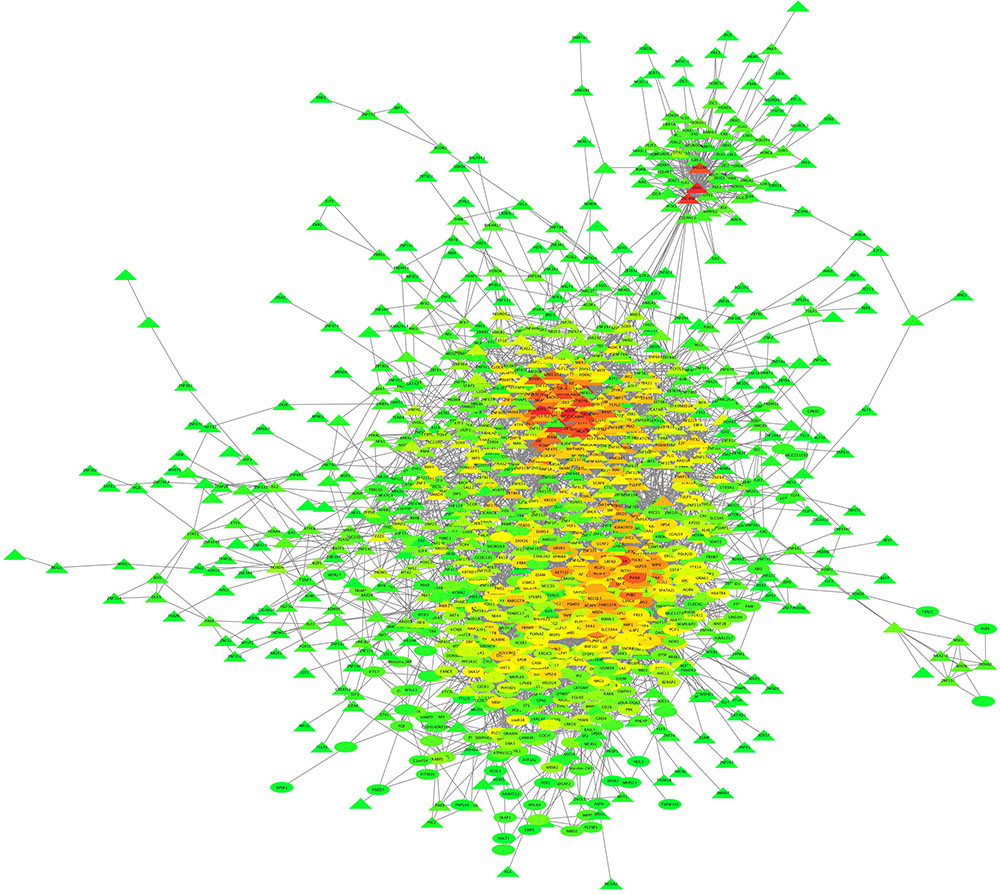
FIGURE 2. Liver gene co-expression network constructed by PCIT in pre- and post-puberty Brahman heifers. The entire network comprises 1,408 nodes (or genes) and 8,330 interactions. The color spectrum ranges from green to red for low and high number of connection, respectively.
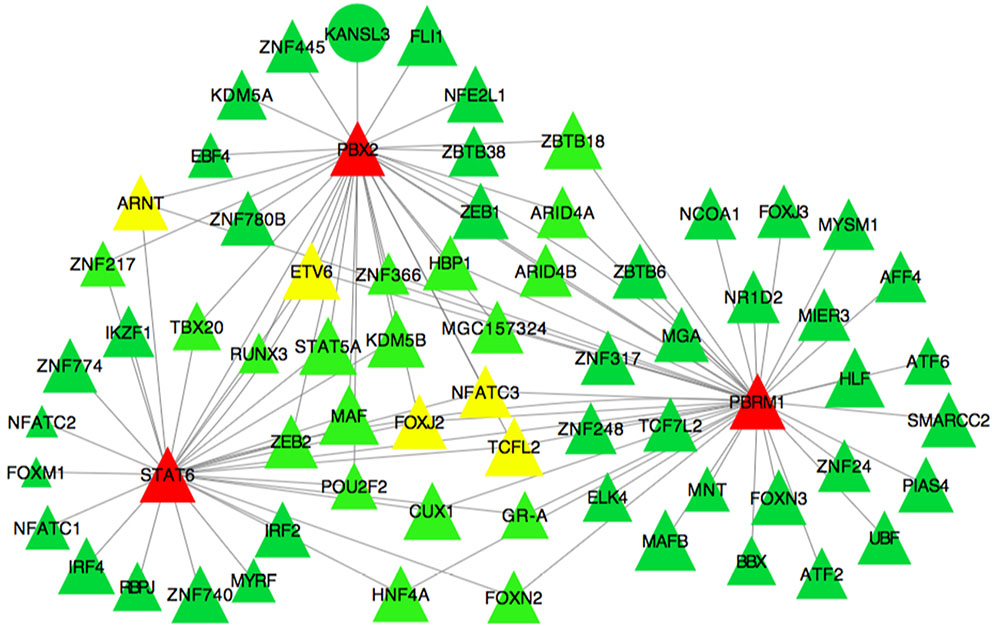
FIGURE 3. Subset of the liver co-expression network showing the best trio of TF and its predicted target genes in pre- and post-puberty Brahman heifers. Each node represents a gene. Nodes represented as triangles are TF and other coding sequences are represented as ellipses. Edges represent significant interaction between nodes. Node color indicates the number of connections of a specific node in the network. The color spectrum ranges from green to red for low and high number of connections.
Functional Enrichment Analysis of Target Genes Involved in Gene Co-expression Network
Functional analysis using DAVID (Dennis et al., 2003; Huang da et al., 2009) allowed identification of biological functions overrepresented in our nodes in the co-expression network. Results showed that 91 GO terms (49.7%, 91/183) were significantly enriched in the biological process category, 8 GO terms (53.3%, 8/15) were significantly enriched in the molecular function category, and 21 GO terms (75%, 21/28) were significantly enriched in the cellular component category. In the biological process category, GO terms were related to liver development, gonad development, immune system development, and muscle organ development. Most importantly, many of the enriched GO terms were closely related to reproduction, including reproductive developmental process, reproductive structure development, and response to protein stimulus. In addition, the molecular function term associated with steroid hormone receptor activity was also enriched in our co-expression network. Pathway analyses revealed 10 significantly enriched pathways (47.6%, 10/21). Among these overrepresented pathways identified, we observed TGF-β signaling (adjusted P = 1.7 × 10-4) and Wnt signaling (adjusted P = 7.4 × 10-4) pathways. Supplementary Table S4 provides the full list of enriched GO terms and pathways, discovered using all genes in the network as the target dataset.
Discussion
Reducing the age at puberty to increase cattle productivity is a major aim for B. indicus breeders. Although the hypothalamus–pituitary–ovarian axis is central to reproduction, the involment of the liver in controlling energy balance and affecting reproduction was reported before (Fontana and Della Torre, 2016; Montagner et al., 2016). IGF1 seems to be an important link between liver function and puberty onset (Akers et al., 2005). Although the post-pubertal liver samples were collected from animals with significantly higher progesterone levels, we found no direct evidence of increased synthesis of liver enzymes involved in the metabolism of steroid hormones. IGF1 transcripts were not among the list of DE genes, although this endocrine signal from the liver is known to increase leading up to puberty and serum IGF1 was increased at post-pubertal Brahman heifers (current study). The bioavailability of and circulating half-life of IGF1 are determined by IGF-binding proteins (IGFBPs) and these may be an important consideration in puberty. It should be noted that most assays measure total IGF1, after extraction procedures that remove IGFBPs, like our results in the current study. Very few studies determine the small (<1% total), free bioactive fraction of IGF1 and/or concentrations of IGFBPs. Our results suggest that circulating IGF1 concentrations are influenced by multiple factors beyond IGF1 gene expression.
In liver, 452 genes were DE between pre- and post-pubertal Brahman heifers (this study). Previously, 288 genes were DE between pre- and post-pubertal Brangus heifers (Cánovas et al., 2014). In Brangus heifers, liver DE genes contributed an abundant number of connections to the co-expression network and had the largest disappearance of connections after puberty. In short, network topology suggests that the liver warrants further scrutiny in Brangus heifers that was beyond the scope of the original publication (Cánovas et al., 2014). Here, we performed gene ontology and pathway enrichment analyses for both lists of DE genes, from the Brangus study and the current Brahman data. No ontologies were the same across breeds. Only 10 DE genes were the same across breeds and these are discussed further below. In short, biological differences between Brangus and Brahman heifers seem evident from the contrasting results in these transcriptomics studies.
The most DE genes (|FC| > 3 and P ≤ 0.01) in the liver of Brahman heifers were snoR38, snorD49, MSMB, RIMKLA, MCCD1, FBLL1, BHLHE22, brain-derived neurotrophic factor (BDNF), and two uncharacterized proteins (ENSBTAG00000044882 and ENSBTAG00000030124). These emerging candidate genes are discussed in the following paragraphs.
Small nucleolar RNA R38 (snoR38) and small nucleolar RNA SNORD49 (snorD49) are non-coding RNAs functioning in modifications of other small nuclear RNA (Matera et al., 2007). The snoRNA families are essential for major biological processes such as mRNA splicing and protein translation (Matera et al., 2007). There is limited evidence for the involvement of these snoRNA with puberty. The first deletion animal model of other snoRNA gene (snorD116) in mice revealed a role in growth and feeding regulation for the snoRNA family (Ding et al., 2008). The highest and lowest mRNA levels after puberty of snoR38 (FC = 5.33) and snorD49 (FC = -4.12) warrant further studies to understand the role that these non-coding RNAs play in liver function and puberty.
The gene β-microseminoprotein (MSMB) plays an important role in semen quality and fertilization (Anahí Franchi et al., 2008). Not restricted to male tissues, MSMB protein was also identified in porcine CL (Tanaka et al., 1995) and its gene expression was identified in human female reproductive tissues (Baijal-Gupta et al., 2000). Importantly, MSMB influences FSH secretion from pituitary gland, impacting on ovarian function (Thakur et al., 1981; Sheth et al., 1984; Frankenberg et al., 2011). It remains to be explored if liver production of MSMB achieves the pituitary signaling in growing heifers.
Very little is known about mitochondrial coiled-coil domain 1 (MCCD1) and fibrillarin-like 1 (FBLL1) function in the liver or with relation to puberty onset. One study in humans identified high expression levels of MCCD1 in fetal liver (Semple et al., 2003). In cattle, MCCD1 was DE in both RNA sequencing studies of puberty, ours, and the study by Cánovas et al. (2014). Its liver function merits further investigation.
The gene RIMKLA is involved in alanine, aspartate, and glutamate metabolism; as per KEEG pathway annotation (Kanehisa et al., 2017). Notably, glutamate and aspartate are major metabolic fuels for nutrient metabolism and oxidative defense (Yao et al., 2008; Brasse-Lagnel et al., 2009). It seems coherent that RIMKLA would be relevant for the liver metabolic function and perhaps it is another link between energy metabolism and reproduction to be explored.
The gene BHLHE22 (FC = -3.00 and P ≤ 0.01) was revealed as the most down-regulated DE gene after puberty. This gene is also a significant TF (RIF2 score of -2.88). The BHLHE22 gene belongs to basic helix–loop–helix (bHLH) family and plays significant role in cell proliferation and differentiation of several developmental pathways as well as cell fate determination (Lee et al., 1995; Ma et al., 1996; Farah et al., 2000; Xu et al., 2002). Further, an in vitro transfection assay used in mice showed that BHLHB5 (an alias of BHLHE22) strongly inhibits the expression of the human PAX6 promoter (Xu et al., 2002). The PAX6 promoter is known as a TF involved in embryonic development and neurulation (Callaerts et al., 1997). A PAX6 mutation was associated with isolated GH deficiency (Guerra-Junior et al., 2008). BHLHE22 has been described as a transcriptional repressor of insulin expression in pancreatic β cells (Peyton et al., 1996; Melkman-Zehavi et al., 2011). Insulin can mediate follicular growth in cattle (Webb et al., 2004), stimulate GnRH release from the hypothalamus in combination with glucose (Arias et al., 1992), and may also facilitate IGF1 synthesis and secretion by the liver (Keisler and Lucy, 1996; Webb et al., 2004). The role of insulin in the regulation of lipid, glucose, protein homeostasis, and energy balance (Saltiel and Kahn, 2001; Liu and Barrett, 2002; Obici and Rossetti, 2003) suggests a link between insulin and the reproductive axis. In our study, BHLHE22 was the most down-regulated gene, with lower expression in post-pubertal heifers (FC = -3.00 and P = 0.01). Lower expression of BHLHE22 could mean decreased repression of insulin expression leading to increased GH and GnRH stimulus via IGF1 signaling. Therefore, liver produced BHLHE22 could impact on animal growth and pubertal development.
The BDNF gene is related to neural development and peripheral metabolism (Binder and Scharfman, 2004; Pedersen et al., 2009). In the brain, BDNF can suppress GABAergic synaptic transmission by acute down-regulation of GABAA receptors and thus can affect GnRH release (Henneberger et al., 2002). Previous studies suggested BDNF as a key component of the hypothalamic pathway controlling energy homeostasis and BW (Wisse and Schwartz, 2003; Xu et al., 2003; Jo and Chua, 2013). A genome-wide association studies (GWASs) in humans found BDNF to be related to timing of puberty and body mass index (Perry et al., 2014). Further, estrogen–BDNF–NPY has been noted as important tri-molecular cascade in understanding the hormonal regulation in hippocampus (Scharfman and MacLusky, 2006). It is unclear whether BDNF is able to cross the blood–brain barrier. Some researchers have found evidence for a link between central BDNF and peripheral BDNF (Poduslo and Curran, 1996; Pan et al., 1998; Rasmussen et al., 2009; Seifert et al., 2010), whereas others have argued that it does not cross the blood–brain barrier (Pardridge et al., 1998; Kyeremanteng et al., 2012). Hence, if further research can prove BDNF delivery across blood–brain barrier, it is possible that BDNF produced in the liver may have endocrine effects in the brain.
Brain-derived neurotrophic factor in the liver, similarly to skeletal muscle, results in increase of AMP-activated protein kinase (AMPK) and its downstream target acetyl coenzyme A carboxylase (ACC), inhibiting fatty acid synthesis and enhancing fatty acid oxidation (Matthews et al., 2009; Pedersen et al., 2009; Genzer et al., 2017). A study in humans suggested that fatty acid oxidation is higher in children than adults (Kostyak et al., 2007). Estrogen was also cited to regulate hepatic fatty acid oxidation (O’Sullivan, 2012). In liver, there is little information of precise mechanisms in which estrogen reduces fatty acid oxidation. Our study results led us to hypothesize that the interaction between estrogen and BDNF in fatty acid oxidation in liver, contributing to metabolic changes that can regulate puberty onset.
Comparing the liver transcriptional profile between our Brahman heifer study and a study by Cánovas et al. (2014) in Brangus heifers, we found 10 genes (including a novel gene) that were DE in both populations (P < 0.05). Five genes, MCCD1, ADGRF2, brain expressed X-linked 2 (BEX2), PDZD7, and LRRC46, had a | FC| > 1 in both breeds. The expression of these genes was up-regulated in Brangus heifers and down-regulated in Brahman heifers. The Brangus study involved eight heifers greatly differing in age and weight whereas our Brahman study used 12 heifers that were age and weight matched. Further, in the absence of a reference genome of B. indicus, we have utilized B. taurus reference genome for sequence assembly, and so the divergence between B. taurus and B. indicus genomes can impact our results. The significant difference in expression levels and patterns of these five DE genes between two breeds warrants further studies. The candidate gene MCCD1 and its limited literature were discussed above. Similarly, PDZD7 and LRRC46 roles in puberty and liver function cannot be speculated from current knowledge. The remaining two genes, ADGRF2 and BEX2, are discussed below.
The expression of adhesion G protein-coupled receptor F2 (ADGRF2, alias GPR111), in reproductive tissues and lung was reported (Fredriksson et al., 2002). Our study was the first to report mRNA expression of ADGRF2 in the liver of B. indicus heifers. It is intriguing to suggest that ADGRF2 could be another link between liver function and puberty, because G protein-coupled receptors have been associated with GnRH regulation (Noel and Kaiser, 2011).
The BEX2 was observed as a DE gene in the ovary of pre- and post-pubertal Brahman heifers (Nguyen et al., 2017) and in the adipose tissue of pre- and post-pubertal Brangus heifers (Cánovas et al., 2014). The BEX2 gene is highly expressed in the human embryonic brain and have a regulatory role in embryonic development (Han et al., 2005). A study of mice liver gene expression revealed a strong expression of BEX2 in stem/progenitor cells (Ito et al., 2014). Further, BEX2 is a downstream molecule of the mammalian target of rapamycin (mTOR) signaling pathway (Hu et al., 2015) that can regulate lipogenesis and ketogenesis in liver (Laplante and Sabatini, 2012). The mTOR pathway is also a known regulator of ovarian activity (Lu et al., 2017). In short, BEX2 was DE in two studies of pubertal heifers, two different breeds, and thus it merits further investigation. It is possible that this is a liver signal with impact on ovarian activity.
Transcription factors play a key role in controlling gene expression, but their expression levels are often low and not detected as DE (Vaquerizas et al., 2009). The interactions between TF are important for tissue remodeling and temporal changes in gene expression (Ravasi et al., 2010). Differential expression analyses overlook vital changes in regulatory information. Adding an analysis focused on identifying key TF could help to understand the gene regulation processes under investigation (i.e., puberty). Previously, we found that TFs in the ZNF were DE and/or important TF in the transcriptomic profile of hypothalamus, pituitary gland, and ovaries in Brahman heifers undergoing puberty (Fortes et al., 2016; Nguyen et al., 2017). These studies noted that 26% of top ranking TF from hypothalamus, 28% from ovaries, and 22% from the pituitary gland top ranking TF coded for ZNF members in the same Brahman heifers (Fortes et al., 2016; Nguyen et al., 2017). Likewise, this current study revealed that 23% of TF identified by RIF analysis of liver transcriptome data belong to the ZNF.
The potential role of ZNF genes in the puberty process was suggested by several studies. A mouse study found that a mutation in regulator of sex-limitation (RSL), one of the Kruppel-associated box zinc finger proteins (KRAB-ZFP) genes, can impact reproduction by regulating expression patterns of target genes in liver (Krebs and Robins, 2010). In addition, ZNF genes have been implicated in the epigenetic control of transcription in the female primate hypothalamus around puberty (Lomniczi et al., 2015). Previous GWASs in women reported the association between single-nucleotide polymorphism located near ZNF462 and ZNF483 and age of menarche, which is the age of puberty in girls (Perry et al., 2009; Elks et al., 2010; Chen et al., 2012; Demerath et al., 2013). Expression of ZNF127 was increased pre-puberty and decreased immediately before puberty (Abreu et al., 2013). Study of female monkeys also reported decrease of ZNF573 mRNA levels in peripubertal animals (Lomniczi et al., 2015). Our study contributes to the growing body of evidence that support ZNF genes can influence puberty onset, a developmental role which may extend to tissues and organs outside of the reproductive axis.
In the sub-network, the trio of TF that spanned most of network topology with minimum redundancy and highest connectivity was STAT6, PBX2, and PBRM1. Previous evidence suggested these TF have important roles in liver and reproductive function. Specifically, the STAT6 locus on BTA5 has been described as a QTL associated with reproduction (Kappes et al., 2000; Allan et al., 2009; Kim et al., 2009; Luna-Nevarez et al., 2011; Hawken et al., 2012). Further, this gene was identified as a key TF in a gene network constructed using GWAS results of first service conception in Brangus heifers (Fortes et al., 2012). Other studies noted the association between STAT6 gene and age at first CL, an indicator of puberty onset in Brahman and Tropical Composite heifers (Fortes et al., 2010, 2011). Our study supported the potential role of STAT6 in puberty onset in Brahman heifers.
The PBX2 gene has a role in the development of ovarian follicles (Ota et al., 2008). Pbx2–Prep1 complexes repress HNF1α-mediated activation of the UDP glucuronosyltransferase family 2 member B17 (UGT2B17) promoter in liver cells (Gregory and Mackenzie, 2002). The UGT2B17 gene, a sex steroid-metabolizing gene, has been associated with male infertility and impaired spermatogenesis (Plaseska-Karanfilska et al., 2012). Hepatocyte nuclear factor-1α (HNF-1α) is a homeodomain-containing TF that regulates liver-specific gene transcription (Mendel and Crabtree, 1991) and was suggested to control development and metabolism in a HNF-1α-null mouse study (Pontoglio et al., 1996). In liver, HNF-1α regulates the expression of glucocorticoid receptor (GR), IGF1, STAT5, and other GH-responsive genes (Lee et al., 1998; Lin et al., 2008). In our sub-network of predicted gene co-expression, PBX2 and STAT6 was also connected to STAT5 (RIF2 score of 2.13) suggesting that the interaction between these TF could contribute to the regulation of growth, liver metabolism, and puberty onset.
Finally, the gene PBRM1 seems to play a role in metabolic and immune system regulation, pertinent to liver expression. The gene PBMR1 was described as a repressor of interleukin 10 (IL-10) transcription; an anti-inflammatory cytokine involved in metabolic syndrome (Mallat et al., 1999; Calcaterra et al., 2009; Wurster et al., 2012). Calcaterra et al. (2009) study showed high levels of IL-10 in serum samples of obese children. Further, IL-10 was proposed to be involved in the inflammatory network of metabolic syndrome in correlation with adiponectin (Böttner et al., 2004; Nishida et al., 2007). Of note, adiponectin plays a significant role in energy homeostasis (Lee and Shao, 2014). In liver, adiponectin can activate glucose transport as well as enhances insulin sensitivity (Berg et al., 2001; Combs et al., 2001; Ye and Scherer, 2013). A study of Holstein cows reported an association between follicular growth and the change in adiponectin and its receptor expression (Tabandeh et al., 2010). The role of PBRM1 as a regulator of heifer puberty needs further investigation, but it is possible that it acts through adiponectin signaling.
After identification of DE genes and TF, GO and pathway analysis was performed to better understand the biological function of these genes in the context of puberty. Information about gene co-expression, enriched GO, and pathways facilitates the interpretation of RNA-Seq results. Based on GO analysis of 1,408 nodes from the liver co-expression network, multiple biological processes were affected. GO terms “reproductive developmental process” and “reproductive structure development” were significantly enriched and are logical in the context of puberty. Steroid hormone receptor activity and steroid binding were expected GO terms as liver is the principal site of steroid hormone metabolism.
We observed TGF-β signaling (P = 6.4 × 10-6) and Wnt signaling (P = 3.8 × 10-5) pathways among the enriched pathways. Both pathways were also enriched in pre- vs. post-pubertal results from the pituitary gland of Brahman heifers (Nguyen et al., 2017). Of note, transforming growth factor-beta (TGF-β) superfamily signaling plays a pivotal role in the regulation of cell differentiation, growth, morphogenesis, tissue homeostasis, and regeneration (Massague, 2012). In neural tissue, TGF-β1 one member of the TGF-β superfamily can increase GnRH gene expression as well as GnRH release (Prevot, 2002; Mahesh et al., 2006). Expression and release of GnRH are pivotal for puberty. The Wnt signaling pathway is an important physiological regulator of embryonic and liver development as well as mammalian hepatic metabolism (McLin et al., 2007; Marfil et al., 2010; Sethi and Vidal-Puig, 2010; Liu et al., 2011). Results from functional enrichment analyses provide evidence of pathways that are relevant for both liver metabolism and reproductive function. These pathways may point to some of the links between liver and reproductive function in B. indicus cattle.
We successfully exploited RNA-Seq data to identify the transcriptomic differences in liver between pre- and post-pubertal Brahman heifers. Previously, liver transcriptomics in B. indicus bulls and steers identified DE genes related to feed efficiency (Alexandre et al., 2015; Tizioto et al., 2015). This paper is the first attempt to demonstrate molecular mechanisms of puberty in liver of Brahman heifers. In the study, 452 DE genes were identified, many of which are closely related to reproductive developmental process, reproductive structure development, steroid hormone receptor activity, and steroid binding. In liver, TGF-β signaling and Wnt signaling genes may play a role in reproductive function. Moreover, the genes, BDNF, STAT6, PBX2, and PBRM1, might impact on the regulation of growth, liver metabolism, and puberty onset. As BDNF and estrogen can regulate fatty acid oxidation, we reasoned that BDNF and estrogen signaling may interact. This interaction can contribute to metabolic changes that can regulate the occurrence of puberty in Brahman heifers. Further studies are warranted to determine the function of these candidate genes. Our findings provide useful information for understanding molecular mechanisms in liver that may influence puberty onset of Brahman heifers.
Author Contributions
LN performed RNA extraction, data analyses, interpretation of results and wrote the first draft. AR performed statistical analyses using mixed models. AC assemble, annotation and count of RNA sequencing data. MD and NC performed quality control of raw data. BV performed laboratory work. SA measured hormones. AI-T performed laboratory work. SL designed the experiment and interpreted the results. JM supervised library preparation and RNA sequencing. MT designed the experiment and interpreted the results. SM obtained funds for the research and supervised the project. MF performed the field trial, sample collection, experimental design, drafting of the manuscript, and interpretation of results.
Funding
The project was funded by a grant to Professor SM from University of Queensland.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors acknowledge the contributions to the field experiment and sample collection of Dr. Gry Boe-Hansen, Dr. Laercio R. Porto-Neto, Dr. Lisa Kidd, and Dr. Joao Paulo A. do Rego.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fgene.2018.00087/full#supplementary-material
Footnotes
- ^ftp://ftp.ensembl.org/pub/release-77/genbank/bos_taurus/
- ^ftp://ftp.tzv.fal.de/pub/vce6
- ^http://bioinfo.life.hust.edu.cn/AnimalTFDB/download_index?tr=Bos_taurus
- ^http://david.abcc.ncifcrf.gov
- ^http://data.faang.org/home
References
Abreu, A. P., Dauber, A., Macedo, D. B., Noel, S. D., Brito, V. N., Gill, J. C., et al. (2013). Central precocious puberty caused by mutations in the imprinted gene MKRN3. N. Engl. J. Med. 368, 2467–2475. doi: 10.1056/NEJMoa1302160
Akbar, H., Bionaz, M., Carlson, D. B., Rodriguez-Za, S. L., Everts, R. E., Lewin, H. A., et al. (2013). Feed restriction, but not l-carnitine infusion, alters the liver transcriptome by inhibiting sterol synthesis and mitochondrial oxidative phosphorylation and increasing gluconeogenesis in mid-lactation dairy cows. J. Dairy Sci. 96, 2201–2213. doi: 10.3168/jds.2012-6036
Akers, R. M., Ellis, S. E., and Berry, S. D. (2005). Ovarian and IGF-I axis control of mammary development in prepubertal heifers. Domest. Anim. Endocrinol. 29, 259–267. doi: 10.1016/j.domaniend.2005.02.037
Alexandre, P. A., Kogelman, L. J., Santana, M. H., Passarelli, D., Pulz, L. H., Fantinato-Neto, P., et al. (2015). Liver transcriptomic networks reveal main biological processes associated with feed efficiency in beef cattle. BMC Genomics 16:1073. doi: 10.1186/s12864-015-2292-8
Allan, M. F., Kuehn, L. A., Cushman, R. A., Snelling, W. M., Echternkamp, S. E., and Thallman, R. M. (2009). Confirmation of quantitative trait loci using a low-density single nucleotide polymorphism map for twinning and ovulation rate on bovine chromosome 5. J. Anim. Sci. 87, 46–56. doi: 10.2527/jas.2008-0959
Anahí Franchi, N., Avendaño, C., Molina, R. I., Tissera, A. D., Maldonado, C. A., Oehninger, S., et al. (2008). beta-Microseminoprotein in human spermatozoa and its potential role in male fertility. Reproduction 136, 157–166. doi: 10.1530/REP-08-0032
Arias, P., Rodriguez, M., Szwarcfarb, B., Sinay, I. R., and Moguilevsky, J. A. (1992). Effect of insulin on LHRH release by perifused hypothalamic fragments. Neuroendocrinology 56, 415–418. doi: 10.1159/000126257
Baijal-Gupta, M., Clarke, M. W., Finkelman, M. A., McLachlin, C. M., and Han, V. K. (2000). Prostatic secretory protein (PSP94) expression in human female reproductive tissues, breast and in endometrial cancer cell lines. J. Endocrinol. 165, 425–433. doi: 10.1677/joe.0.1650425
Berg, A. H., Combs, T. P., Du, X., Brownlee, M., and Scherer, P. E. (2001). The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat. Med. 7, 947–953. doi: 10.1038/90992
Binder, D. K., and Scharfman, H. E. (2004). Brain-derived neurotrophic factor. Growth Factors 22, 123–131. doi: 10.1080/08977190410001723308
Böttner, A., Kratzsch, J., Müller, G., Kapellen, T. M., Blüher, S., Keller, E., et al. (2004). Gender differences of adiponectin levels develop during the progression of puberty and are related to serum androgen levels. J. Clin. Endocrinol. Metab. 89, 4053–4061. doi: 10.1210/jc.2004-0303
Brasse-Lagnel, C., Lavoinne, A., and Husson, A. (2009). Control of mammalian gene expression by amino acids, especially glutamine. FEBS J. 276, 1826–1844. doi: 10.1111/j.1742-4658.2009.06920.x
Calcaterra, V., De Amici, M., Klersy, C., Torre, C., Brizzi, V., Scaglia, F., et al. (2009). Adiponectin, IL-10 and metabolic syndrome in obese children and adolescents. Acta Biomed. 80, 117–123.
Callaerts, P., Halder, G., and Gehring, W. J. (1997). PAX-6 in development and evolution. Annu. Rev. Neurosci. 20, 483–532. doi: 10.1146/annurev.neuro.20.1.483
Cánovas, A., Reverter, A., DeAtley, K. L., Ashley, R. L., Colgrave, M. L., Fortes, M. R., et al. (2014). Multi-tissue omics analyses reveal molecular regulatory networks for puberty in composite beef cattle. PLoS One 9:e102551. doi: 10.1371/journal.pone.0102551
Cánovas, A., Rincón, G., Islas-Trejo, A., Jimenez-Flores, R., Laubscher, A., and Medrano, J. F. (2013). RNA sequencing to study gene expression and single nucleotide polymorphism variation associated with citrate content in cow milk. J. Dairy Sci. 96, 2637–2648. doi: 10.3168/jds.2012-6213
Cánovas, A., Rincon, G., Islas-Trejo, A., Wickramasinghe, S., and Medrano, J. F. (2010). SNP discovery in the bovine milk transcriptome using RNA-Seq technology. Mamm. Genome 21, 592–598. doi: 10.1007/s00335-010-9297-z
Chen, C. T., Fernández-Rhodes, L., Brzyski, R. G., Carlson, C. S., Chen, Z., Heiss, G., et al. (2012). Replication of loci influencing ages at menarche and menopause in Hispanic women: the Women’s Health Initiative SHARe Study. Hum. Mol. Genet. 21, 1419–1432. doi: 10.1093/hmg/ddr570
Combs, T. P., Berg, A. H., Obici, S., Scherer, P. E., and Rossetti, L. (2001). Endogenous glucose production is inhibited by the adipose-derived protein Acrp30. J. Clin. Invest. 108, 1875–1881. doi: 10.1172/JCI14120
Curlewis, J. D., Axelson, M., and Stone, G. M. (1985). Identification of the major steroids in ovarian and adrenal venous plasma of the brush-tail possum (Trichosurus vulpecula) and changes in the peripheral plasma levels of oestradiol and progesterone during the reproductive cycle. J. Endocrinol. 105, 53–62. doi: 10.1677/joe.0.1050053
Dahlanuddin, D., Ningsih, B. S., Poppi, D. P., Anderson, S. T., and Quigley, S. P. (2014). Long-term growth of male and female Bali cattle fed Sesbania grandiflora. Anim. Prod. Sci. 54, 1615–1619.
Della Torre, S., Rando, G., Meda, C., Stell, A., Chambon, P., Krust, A., et al. (2011). Amino acid-dependent activation of liver estrogen receptor alpha integrates metabolic and reproductive functions via IGF-1. Cell Metab. 13, 205–214. doi: 10.1016/j.cmet.2011.01.002
Demerath, E. W., Liu, C. T., Franceschini, N., Chen, G., Palmer, J. R., Smith, E. N., et al. (2013). Genome-wide association study of age at menarche in African-American women. Hum. Mol. Genet. 22, 3329–3346. doi: 10.1093/hmg/ddt181
Dennis, G. Jr., Sherman, B. T., and Hosack, D. A. (2003). DAVID: database for annotation, visualization, and integrated discovery. Genome Biol. 4:R60. doi: 10.1186/gb-2003-4-9-r60
Ding, F., Li, H. H., Zhang, S., Solomon, N. M., Camper, S. A., Cohen, P., et al. (2008). SnoRNA Snord116 (Pwcr1/MBII-85) deletion causes growth deficiency and hyperphagia in mice. PLoS One 3:e1709. doi: 10.1371/journal.pone.0001709
Elks, C. E., Perry, J. R., Sulem, P., Chasman, D. I., Franceschini, N., He, C., et al. (2010). Thirty new loci for age at menarche identified by a meta-analysis of genome-wide association studies. Nat. Genet. 42, 1077–1085. doi: 10.1038/ng.714
Farah, M. H., Olson, J. M., Sucic, H. B., Hume, R. I., Tapscott, S. J., and Turner, D. L. (2000). Generation of neurons by transient expression of neural bHLH proteins in mammalian cells. Development 127, 693–702.
Fontana, R., and Della Torre, S. (2016). The deep correlation between energy metabolism and reproduction: a view on the effects of nutrition for women fertility. Nutrients 8:87. doi: 10.3390/nu8020087
Fortes, M. R., Li, Y., Collis, E., Zhang, Y., and Hawken, R. J. (2013). The IGF1 pathway genes and their association with age of puberty in cattle. Anim. Genet. 44, 91–95. doi: 10.1111/j.1365-2052.2012.02367.x
Fortes, M. R., Nguyen, L. T., Weller, M. M., Cánovas, A., Islas-Trejo, A., Porto-Neto, L. R., et al. (2016). Transcriptome analyses identify five transcription factors differentially expressed in the hypothalamus of post- versus prepubertal Brahman heifers. J. Anim. Sci. 94, 3693–3702. doi: 10.2527/jas.2016-0471
Fortes, M. R., Reverter, A., Nagaraj, S. H., Zhang, Y., Jonsson, N. N., Barris, W., et al. (2011). A single nucleotide polymorphism-derived regulatory gene network underlying puberty in 2 tropical breeds of beef cattle. J. Anim. Sci. 89, 1669–1683. doi: 10.2527/jas.2010-3681
Fortes, M. R., Reverter, A., Zhang, Y., Collis, E., Nagaraj, S. H., Jonsson, N. N., et al. (2010). Association weight matrix for the genetic dissection of puberty in beef cattle. Proc. Natl. Acad. Sci. U.S.A. 107, 13642–13647. doi: 10.1073/pnas.1002044107
Fortes, M. R., Snelling, W. M., Reverter, A., Nagaraj, S. H., Lehnert, S. A., Hawken, R. J., et al. (2012). Gene network analyses of first service conception in Brangus heifers: use of genome and trait associations, hypothalamic-transcriptome information, and transcription factors. J. Anim. Sci. 90, 2894–2906. doi: 10.2527/jas.2011-4601
Frankenberg, S., Fenelon, J., Dopheide, B., Shaw, G., and Renfree, M. B. (2011). A novel MSMB-related microprotein in the postovulatory egg coats of marsupials. BMC Evol. Biol. 11:373. doi: 10.1186/1471-2148-11-373
Fredriksson, R., Lagerstr, M. C., Höglund, P. J., and Schiöth, H. B. (2002). Novel human G protein-coupled receptors with long N-terminals containing GPS domains and Ser/Thr-rich regions. FEBS Lett. 531, 407–414. doi: 10.1016/S0014-5793(02)03574-3
Genzer, Y., Chapnik, N., and Froy, O. (2017). Effect of brain-derived neurotrophic factor (BDNF) on hepatocyte metabolism. Int. J. Biochem. Cell Biol. 88(Suppl. C), 69–74. doi: 10.1016/j.biocel.2017.05.008
Gregory, P. A., and Mackenzie, P. I. (2002). The homeodomain Pbx2-Prep1 complex modulates hepatocyte nuclear factor 1alpha-mediated activation of the UDP-glucuronosyltransferase 2B17 gene. Mol. Pharmacol. 62, 154–161. doi: 10.1124/mol.62.1.154
Guerra-Junior, G., Spinola-Castro, A. M., Siviero-Miachon, A. A., Nogueira, R. G., Lemos-Marini, S. H., D’Souza-Li, L. F., et al. (2008). Absence of mutations in Pax6 gene in three cases of morning glory syndrome associated with isolated growth hormone deficiency. Arq. Bras. Endocrinol. Metabol. 52, 1221–1227. doi: 10.1590/S0004-27302008000800004
Han, C., Liu, H., Liu, J., Yin, K., Xie, Y., Shen, X., et al. (2005). Human Bex2 interacts with LMO2 and regulates the transcriptional activity of a novel DNA-binding complex. Nucleic Acids Res. 33, 6555–6565. doi: 10.1093/nar/gki964
Hawken, R. J., Zhang, Y. D., Fortes, M. R., Collis, E., Barris, W. C., Corbet, N. J., et al. (2012). Genome-wide association studies of female reproduction in tropically adapted beef cattle. J. Anim. Sci. 90, 1398–1410. doi: 10.2527/jas.2011-4410
Henneberger, C., Juttner, R., Rothe, T., and Grantyn, R. (2002). Postsynaptic action of BDNF on GABAergic synaptic transmission in the superficial layers of the mouse superior colliculus. J. Neurophysiol. 88, 595–603. doi: 10.1152/jn.2002.88.2.595
Herath, C. B., Shiojima, S., Ishiwata, H., Katsuma, S., Kadowaki, T., Ushizawa, K., et al. (2004). Pregnancy-associated changes in genome-wide gene expression profiles in the liver of cow throughout pregnancy. Biochem. Biophys. Res. Commun. 313, 666–680. doi: 10.1016/j.bbrc.2003.11.151
Hu, Z., Wang, Y., Huang, F., Chen, R., Li, C., Wang, F., et al. (2015). Brain-expressed X-linked 2 is pivotal for hyperactive mechanistic target of rapamycin (mTOR)-mediated tumorigenesis. J. Biol. Chem. 290, 25756–25765. doi: 10.1074/jbc.M115.665208
Huang da, W., Sherman, B. T., and Lempicki, R. A. (2009). Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 4, 44–57. doi: 10.1038/nprot.2008.211
Hudson, N. J., Reverter, A., Wang, Y., Greenwood, P. L., and Dalrymple, B. P. (2009). Inferring the transcriptional landscape of bovine skeletal muscle by integrating co-expression networks. PLoS One 4:e7249. doi: 10.1371/journal.pone.0007249
Ito, K., Yamazaki, S., Yamamoto, R., Tajima, Y., Yanagida, A., Kobayashi, T., et al. (2014). Gene targeting study reveals unexpected expression of Brain-expressed X-linked 2 in endocrine and tissue Stem/Progenitor cells in Mice. J. Biol. Chem. 289, 29892–29911. doi: 10.1074/jbc.M114.580084
Jo, Y.-H., and Chua, S. C. (2013). The brain–liver connection between BDNF and glucose control. Diabetes Metab. Res. Rev. 62, 1367–1368. doi: 10.2337/db12-1824
Johnston, D. J., Barwick, S. A., Corbet, N. J., et al. (2009). Genetics of heifer puberty in two tropical beef genotypes in northern Australia and associations with heifer- and steer-production traits. Anim. Prod. Sci. 49, 399–412. doi: 10.1071/EA08276
Johnston, D. J., Barwick, S. A., Fordyce, G., Holroyd, R. G., Williams, P. J., Corbet, N. J., et al. (2013). Genetics of early and lifetime annual reproductive performance in cows of two tropical beef genotypes in northern Australia. Anim. Prod. Sci. 54, 1–15. doi: 10.1071/AN13043
Kanehisa, M., Furumichi, M., Tanabe, M., Sato, Y., and Morishima, K. (2017). KEGG: new perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 45, D353–D361. doi: 10.1093/nar/gkw1092
Kappes, S. M., Bennett, G. L., Keele, J. W., Echternkamp, S. E., Gregory, K. E., and Thallman, R. M. (2000). Initial results of genomic scans for ovulation rate in a cattle population selected for increased twinning rate. J. Anim. Sci. 78, 3053–3059. doi: 10.2527/2000.78123053x
Keisler, D. H., and Lucy, M. C. (1996). Perception and interpretation of the effects of undernutrition on reproduction. J. Anim. Sci. 74(Suppl._3), 1–17. doi: 10.2527/1996.74suppl_31x
Kim, E. S., Shi, X., Cobanoglu, O., Weigel, K., Berger, P. J., and Kirkpatrick, B. W. (2009). Refined mapping of twinning-rate quantitative trait loci on bovine chromosome 5 and analysis of insulin-like growth factor-1 as a positional candidate gene. J. Anim. Sci. 87, 835–843. doi: 10.2527/jas.2008-1252
Kostyak, J. C., Kris-Etherton, P., Bagshaw, D., DeLany, J. P., and Farrell, P. A. (2007). Relative fat oxidation is higher in children than adults. Nutr. J. 6, 19–19. doi: 10.1186/1475-2891-6-19
Krebs, C. J., and Robins, D. M. (2010). A pair of mouse KRAB Zinc finger proteins modulates multiple indicators of female reproduction. Biol. Reprod. 82, 662–668. doi: 10.1095/biolreprod.109.080846
Kyeremanteng, C., James, J., Mackay, J., and Merali, Z. (2012). A study of brain and serum brain-derived neurotrophic factor protein in Wistar and Wistar-Kyoto rat strains after electroconvulsive stimulus. Pharmacopsychiatry 45, 244–249. doi: 10.1055/s-0032-1306278
Laplante, M., and Sabatini, D. M. (2012). mTOR signaling in growth control and disease. Cell 149, 274–293. doi: 10.1016/j.cell.2012.03.017
Laporta, J., Rosa, G. J., Naya, H., and Carriquiry, M. (2014). Liver functional genomics in beef cows on grazing systems: novel genes and pathways revealed. Physiol. Genomics 46, 138–147. doi: 10.1152/physiolgenomics.00120.2013
Lee, B., and Shao, J. (2014). Adiponectin and energy homeostasis. Rev. Endocr. Metab. Disord. 15, 149–156. doi: 10.1007/s11154-013-9283-3
Lee, H. J., Jang, M., Kim, H., Kwak, W., Park, W., Hwang, J. Y., et al. (2013). Comparative transcriptome analysis of adipose tissues reveals that ECM-receptor interaction is involved in the depot-specific adipogenesis in cattle. PLoS One 8:e66267. doi: 10.1371/journal.pone.0066267
Lee, J. E., Hollenberg, S. M., Snider, L., Turner, D. L., Lipnick, N., and Weintraub, H. (1995). Conversion of Xenopus ectoderm into neurons by neuroD, a basic helix-loop-helix protein. Science 268, 836–844. doi: 10.1126/science.7754368
Lee, Y. H., Sauer, B., and Gonzalez, F. J. (1998). Laron dwarfism and non-insulin-dependent diabetes mellitus in the Hnf-1alpha knockout mouse. Mol. Cell. Biol. 18, 3059–3068. doi: 10.1128/MCB.18.5.3059
Lesmeister, J. L., Burfening, P. J., and Blackwell, R. L. (1973). Date of first calving in beef cows and subsequent calf production. J. Anim. Sci. 36, 1–6. doi: 10.2527/jas1973.3611
Lin, W.-Y., Yu-Jie, H., and Lee, Y.-H. (2008). Hepatocyte nuclear factor-1α regulates glucocorticoid receptor expression to control postnatal body growth. Am. J. Physiol. Gastrointest. Liver Physiol. 295, G542–G551. doi: 10.1152/ajpgi.00081.2008
Liu, H., Fergusson, M. M., Wu, J. J., Rovira, I. I., Liu, J., Gavrilova, O., et al. (2011). Wnt signaling regulates hepatic metabolism. Sci. Signal. 4:ra6. doi: 10.1126/scisignal.2001249
Liu, Z., and Barrett, E. J. (2002). Human protein metabolism: its measurement and regulation. Am. J. Physiol. Endocrinol. Metab. 283, E1105–E1112. doi: 10.1152/ajpendo.00337.2002
Lomniczi, A., Wright, H., Castellano, J. M., Matagne, V., Toro, C. A., Ramaswamy, S., et al. (2015). Epigenetic regulation of puberty via Zinc finger protein-mediated transcriptional repression. Nat. Commun. 6:10195. doi: 10.1038/ncomms10195
Loor, J. J. (2010). Genomics of metabolic adaptations in the peripartal cow. Animal 4, 1110–1139. doi: 10.1017/S1751731110000960
Loor, J. J., Dann, H. M., Everts, R. E., Oliveira, R., Green, C. A., Guretzky, N. A., et al. (2005). Temporal gene expression profiling of liver from periparturient dairy cows reveals complex adaptive mechanisms in hepatic function. Physiol. Genomics 23, 217–226. doi: 10.1152/physiolgenomics.00132.2005
Lu, X., Guo, S., Cheng, Y., Kim, J. H., Feng, Y., and Feng, Y. (2017). Stimulation of ovarian follicle growth after AMPK inhibition. Reproduction 153, 683–694. doi: 10.1530/REP-16-0577
Luna-Nevarez, P., Rincon, G., Medrano, J. F., Riley, D. G., Chase, C. C. Jr., Coleman, S. W., et al. (2011). Single nucleotide polymorphisms in the growth hormone-insulin-like growth factor axis in straightbred and crossbred Angus, Brahman, and Romosinuano heifers: population genetic analyses and association of genotypes with reproductive phenotypes. J. Anim. Sci. 89, 926–934. doi: 10.2527/jas.2010-3483
Ma, Q., Kintner, C., and Anderson, D. J. (1996). Identification of neurogenin, a vertebrate neuronal determination gene. Cell 87, 43–52. doi: 10.1016/S0092-8674(00)81321-5
Mahesh, V. B., Dhandapani, K. M., and Brann, D. W. (2006). Role of astrocytes in reproduction and neuroprotection. Mol. Cell. Endocrinol. 246, 1–9. doi: 10.1016/j.mce.2005.11.017
Mallat, Z., Heymes, C., Ohan, J., Faggin, E., Lesèche, G., and Tedgui, A. (1999). Expression of interleukin-10 in advanced human atherosclerotic plaques: relation to inducible nitric oxide synthase expression and cell death. Arterioscler. Thromb. Vasc. Biol. 19, 611–616. doi: 10.1161/01.ATV.19.3.611
Marfil, V., Moya, M., Pierreux, C. E., Castell, J. V., Lemaigre, F. P., Real, F. X., et al. (2010). interaction between Hhex and SOX13 modulates Wnt/TCF activity. J. Biol. Chem. 285, 5726–5737. doi: 10.1074/jbc.M109.046649
Marioni, J. C., Mason, C. E., Mane, S. M., Stephens, M., and Gilad, Y. (2008). RNA-seq: an assessment of technical reproducibility and comparison with gene expression arrays. Genome Res. 18, 1509–1517. doi: 10.1101/gr.079558.108
Massague, J. (2012). TGFβ signalling in context. Nat. Rev. Mol. Cell Biol. 13, 616–630. doi: 10.1038/nrm3434
Matera, A. G., Terns, R. M., and Terns, M. P. (2007). Non-coding RNAs: lessons from the small nuclear and small nucleolar RNAs. Nat. Rev. Mol. Cell Biol. 8, 209–220. doi: 10.1038/nrm2124
Matthews, V. B., Aström, M. B., Chan, M. H., Bruce, C. R., Krabbe, K. S., Prelovsek, O., et al. (2009). Brain-derived neurotrophic factor is produced by skeletal muscle cells in response to contraction and enhances fat oxidation via activation of AMP-activated protein kinase. Diabetologia 52, 1409–1418. doi: 10.1007/s00125-009-1364-1
McCarthy, S. D., Waters, S. M., Kenny, D. A., Diskin, M. G., Fitzpatrick, R., Patton, J., et al. (2010). Negative energy balance and hepatic gene expression patterns in high-yielding dairy cows during the early postpartum period: a global approach. Physiol. Genomics 42A, 188–199. doi: 10.1152/physiolgenomics.00118.2010
McLin, V. A., Rankin, S. A., and Zorn, A. M. (2007). Repression of Wnt/β-catenin signaling in the anterior endoderm is essential for liver and pancreas development. Development 134, 2207–2217. doi: 10.1242/dev.001230
Melkman-Zehavi, T., Oren, R., Kredo-Russo, S., Shapira, T., Mandelbaum, A. D., Rivkin, N., et al. (2011). miRNAs control insulin content in pancreatic β-cells via downregulation of transcriptional repressors. EMBO J. 30, 835–845. doi: 10.1038/emboj.2010.361
Mendel, D. B., and Crabtree, G. R. (1991). HNF-1, a member of a novel class of dimerizing homeodomain proteins. J. Biol. Chem. 266, 677–680.
Mircea, C. N., Lujan, M. E., and Pierson, R. A. (2007). Metabolic fuel and clinical implications for female reproduction. J. Obstet. Gynaecol. Can. 29, 887–902. doi: 10.1016/S1701-2163(16)32661-5
Montagner, P., Krause, A. R., Schwegler, E., Weschenfelder, M. M., Rabassa, V. R., Schneider, A., et al. (2016). Reduction of liver function delays resumption of postpartum ovarian activity and alters the synthesis of acute phase proteins in dairy cows. Res. Vet. Sci. 106, 84–88. doi: 10.1016/j.rvsc.2016.02.015
Mortazavi, A., Williams, B. A., McCue, K., Schaeffer, L., and Wold, B. (2008). Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 5, 621–628. doi: 10.1038/nmeth.1226
Nguyen, L. T., Reverter, A., Cánovas, A., Venus, B., Islas-Trejo, A., Porto-Neto, L. R., et al. (2017). Global differential gene expression in the pituitary gland and the ovaries of pre- and post-pubertal Brahman heifers. J. Anim. Sci. 95, 599–615. doi: 10.2527/jas.2016.0921
Nishida, M., Moriyama, T., Sugita, Y., and Yamauchi-Takihara, K. (2007). Interleukin-10 associates with adiponectin predominantly in subjects with metabolic syndrome. Circ. J. 71, 1234–1238. doi: 10.1253/circj.71.1234
Noel, S. D., and Kaiser, U. B. (2011). G protein-coupled receptors involved in GnRH regulation: molecular insights from human disease. Mol. Cell. Endocrinol. 346, 91–101. doi: 10.1016/j.mce.2011.06.022
Obici, S., and Rossetti, L. (2003). Minireview: nutrient sensing and the regulation of insulin action and energy balance. Endocrinology 144, 5172–5178. doi: 10.1210/en.2003-0999
O’Sullivan, A. J. (2012). “Fat storage in women: from puberty to the menopause,” in Handbook of Growth and Growth Monitoring in Health and Disease, ed. V. R. Preedy (New York, NY: Springer), 1087–1101. doi: 10.1007/978-1-4419-1795-9_65
Ota, T., Asahina, H., Park, S. H., Huang, Q., Minegishi, T., Auersperg, N., et al. (2008). HOX cofactors expression and regulation in the human ovary. Reprod. Biol. Endocrinol. 6:49. doi: 10.1186/1477-7827-6-49
Pan, W., Banks, W. A., Fasold, M. B., Bluth, J., et al. (1998). Transport of brain-derived neurotrophic factor across the blood-brain barrier. Neuropharmacology 37, 1553–1561. doi: 10.1016/S0028-3908(98)00141-5
Pardridge, W. M., Wu, D., and Sakane, T. (1998). Combined use of carboxyl-directed protein pegylation and vector-mediated blood-brain barrier drug delivery system optimizes brain uptake of brain-derived neurotrophic factor following intravenous administration. Pharm. Res. 15, 576–582. doi: 10.1023/A:1011981927620
Pedersen, B. K., Pedersen, M., Krabbe, K. S., Bruunsgaard, H., Matthews, V. B., and Febbraio, M. A. (2009). Role of exercise-induced brain-derived neurotrophic factor production in the regulation of energy homeostasis in mammals. Exp. Physiol. 94, 1153–1160. doi: 10.1113/expphysiol.2009.048561
Perry, J. R., Day, F., Elks, C. E., Sulem, P., Thompson, D. J., Ferreira, T., et al. (2014). Parent-of-origin-specific allelic associations among 106 genomic loci for age at menarche. Nature 514, 92–97. doi: 10.1038/nature13545
Perry, J. R., Stolk, L., Franceschini, N., Lunetta, K. L., Zhai, G., McArdle, P. F., et al. (2009). Meta-analysis of genome-wide association data identifies two loci influencing age at menarche. Nat. Genet. 41, 648–650. doi: 10.1038/ng.386
Peyton, M., Stellrecht, C. M., Naya, F. J., Huang, H. P., Samora, P. J., and Tsai, M. J. (1996). BETA3, a novel helix-loop-helix protein, can act as a negative regulator of BETA2 and MyoD-responsive genes. Mol. Cell. Biol. 16, 626–633. doi: 10.1128/MCB.16.2.626
Plaseska-Karanfilska, D., Noveski, P., Plaseski, T., Maleva, I., Madjunkova, S., and Moneva, Z. (2012). Genetic causes of male infertility. Balkan J. Med. Genet. 15, 31–34.
Poduslo, J. F., and Curran, G. L. (1996). Permeability at the blood-brain and blood-nerve barriers of the neurotrophic factors: NGF, CNTF, NT-3, BDNF. Mol. Brain Res. 36, 280–286. doi: 10.1016/0169-328X(95)00250-V
Pontoglio, M., Barra, J., Hadchouel, M., Doyen, A., Kress, C., Bach, J. P., et al. (1996). Hepatocyte nuclear factor 1 inactivation results in hepatic dysfunction, phenylketonuria, and renal Fanconi syndrome. Cell 84, 575–585. doi: 10.1016/S0092-8674(00)81033-8
Porto Neto, L. R., Lehnert, S. A., Fortes, M. R. S., Kelly, M., and Reverter, A. (2013). “Population stratification and breed composition of australian tropically adapted cattle,” in Proceedings of the Association for the Advancement of Animal Breeding and Genetics, Napier, 147–150.
Prevot, V. (2002). Glial-neuronal-endothelial interactions are involved in the control of GnRH secretion. J. Neuroendocrinol. 14, 247–255. doi: 10.1046/j.0007-1331.2001.00772.x
Rasmussen, P., Brassard, P., Adser, H., Pedersen, M. V., Leick, L., Hart, E., et al. (2009). Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Exp. Physiol. 94, 1062–1069. doi: 10.1113/expphysiol.2009.048512
Ravasi, T., Suzuki, H., Cannistraci, C. V., Katayama, S., Bajic, V. B., Tan, K., et al. (2010). An atlas of combinatorial transcriptional regulation in mouse and man. Cell 140, 744–752. doi: 10.1016/j.cell.2010.01.044
Reverter, A., Barris, W., McWilliam, S., Byrne, K. A., Wang, Y. H., Tan, S. H., et al. (2005). Validation of alternative methods of data normalization in gene co-expression studies. Bioinformatics 21, 1112–1120. doi: 10.1093/bioinformatics/bti124
Reverter, A., and Chan, E. K. (2008). Combining partial correlation and an information theory approach to the reversed engineering of gene co-expression networks. Bioinformatics 24, 2491–2497. doi: 10.1093/bioinformatics/btn482
Reverter, A., and Fortes, M. R. (2013). Breeding and genetics symposium: building single nucleotide polymorphism-derived gene regulatory networks: towards functional genomewide association studies. J. Anim. Sci. 91, 530–536. doi: 10.2527/jas.2012-5780
Reverter, A., Hudson, N. J., Nagaraj, S. H., Perez-Enciso, M., and Dalrymple, B. P. (2010). Regulatory impact factors: unraveling the transcriptional regulation of complex traits from expression data. Bioinformatics 26, 896–904. doi: 10.1093/bioinformatics/btq051
Saltiel, A. R., and Kahn, C. R. (2001). Insulin signalling and the regulation of glucose and lipid metabolism. Nature 414, 799–806. doi: 10.1038/414799a
Scharfman, H. E., and MacLusky, N. J. (2006). Estrogen and brain-derived neurotrophic factor (BDNF) in hippocampus: complexity of steroid hormone-growth factor interactions in the adult CNS. Front. Neuroendocrinol. 27, 415–435. doi: 10.1016/j.yfrne.2006.09.004
Seifert, T., Brassard, P., Wissenberg, M., Rasmussen, P., Nordby, P., Stallknecht, B., et al. (2010). Endurance training enhances BDNF release from the human brain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 298, R372–R377. doi: 10.1152/ajpregu.00525.2009
Semple, J. I., Ribas, G., Hillyard, G., Brown, S. E., Sanderson, C. M., and Campbell, R. D. (2003). A novel gene encoding a coiled-coil mitochondrial protein located at the telomeric end of the human MHC Class III region. Gene 314, 41–54. doi: 10.1016/S0378-1119(03)00735-2
Sethi, J. K., and Vidal-Puig, A. (2010). Wnt signalling and the control of cellular metabolism. Biochem. J. 427, 1–17. doi: 10.1042/BJ20091866
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Sheth, A. R., Arabatti, N., Carlquist, M., and Jornvall, H. (1984). Characterization of a polypeptide from human seminal plasma with inhibin (inhibition of FSH secretion)-like activity. FEBS Lett. 165, 11–15. doi: 10.1016/0014-5793(84)80004-6
Singh, P., Singh, G., Bhandawat, A., Singh, G., Parmar, R., Seth, R., et al. (2017). Spatial transcriptome analysis provides insights of key gene(s) involved in steroidal saponin biosynthesis in medicinally important herb Trillium govanianum. Sci. Rep. 7:45295. doi: 10.1038/srep45295
Tabandeh, M. R., Hosseini, A., Saeb, M., Kafi, M., and Saeb, S. (2010). Changes in the gene expression of adiponectin and adiponectin receptors (AdipoR1 and AdipoR2) in ovarian follicular cells of dairy cow at different stages of development. Theriogenology 73, 659–669. doi: 10.1016/j.theriogenology.2009.11.006
Tanaka, T., Itahana, K., Andoh, N., Takeya, T., and Sato, E. (1995). Expression of prostatic secretory protein (PSP)-like protein in porcine corpus luteum: isolation and characterization of a new gene encoding PSP94-like protein. Mol. Reprod. Dev. 42, 149–156. doi: 10.1002/mrd.1080420204
Thakur, A. N., Vaze, A. Y., Dattatreyamurthy, B., and Sheth, A. R. (1981). Isolation & characterization of inhibin from human seminal plasma. Indian J. Exp. Biol. 19, 307–313.
Tizioto, P. C., Coutinho, L. L., Decker, J. E., Schnabel, R. D., Rosa, K. O., Oliveira, P. S., et al. (2015). Global liver gene expression differences in Nelore steers with divergent residual feed intake phenotypes. BMC Genomics 16:242. doi: 10.1186/s12864-015-1464-x
Vaquerizas, J. M., Kummerfeld, S. K., Teichmann, S. A., and Luscombe, N. M. (2009). A census of human transcription factors: function, expression and evolution. Nat. Rev. Genet. 10, 252–263. doi: 10.1038/nrg2538
Wang, Y., Ghaffari, N., Johnson, C. D., Braga-Neto, U. M., Wang, H., Chen, R., et al. (2011). Evaluation of the coverage and depth of transcriptome by RNA-Seq in chickens. BMC Bioinformatics 12(Suppl. 10):S5. doi: 10.1186/1471-2105-12-S10-S5
Webb, R., Garnsworthy, P. C., Gong, J. G., and Armstrong, D. G. (2004). Control of follicular growth: local interactions and nutritional influences. J. Anim. Sci. 82(E–Suppl.), E63–E74.
Wickramasinghe, S., Rincon, G., Islas-Trejo, A., and Medrano, J. F. (2012). Transcriptional profiling of bovine milk using RNA sequencing. BMC Genomics 13:45. doi: 10.1186/1471-2164-13-45
Wisse, B. E., and Schwartz, M. W. (2003). The skinny on neurotrophins. Nat. Neurosci. 6, 655–656. doi: 10.1038/nn0703-655
Wurster, A. L., Precht, P., Becker, K. G., Wood, W. H. III, Zhang, Y., Wang, Z., et al. (2012). IL-10 transcription is negatively regulated by BAF180, a component of the SWI/SNF chromatin remodeling enzyme. BMC Immunol. 13:9. doi: 10.1186/1471-2172-13-9
Xu, B., Goulding, E. H., Zang, K., Cepoi, D., Cone, R. D., Jones, K. R., et al. (2003). Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nat. Neurosci. 6, 736–742. doi: 10.1038/nn1073
Xu, Z. P., Dutra, A., Stellrecht, C. M., Wu, C., Piatigorsky, J., and Saunders, G. F. (2002). Functional and structural characterization of the human gene BHLHB5, encoding a basic helix-loop-helix transcription factor. Genomics 80, 311–318. doi: 10.1006/geno.2002.6833
Yao, K., Yin, Y. L., Chu, W., Liu, Z., Deng, D., Li, T., et al. (2008). Dietary arginine supplementation increases mTOR signaling activity in skeletal muscle of neonatal pigs. J. Nutr. 138, 867–872. doi: 10.1093/jn/138.5.867
Keywords: Bos indicus, puberty, gene expression, RNA sequencing, gene network, liver
Citation: Nguyen LT, Reverter A, Cánovas A, Venus B, Anderson ST, Islas-Trejo A, Dias MM, Crawford NF, Lehnert SA, Medrano JF, Thomas MG, Moore SS and Fortes MRS (2018) STAT6, PBX2, and PBRM1 Emerge as Predicted Regulators of 452 Differentially Expressed Genes Associated With Puberty in Brahman Heifers. Front. Genet. 9:87. doi: 10.3389/fgene.2018.00087
Received: 08 December 2017; Accepted: 02 March 2018;
Published: 20 March 2018.
Edited by:
Haja N. Kadarmideen, Technical University of Denmark, DenmarkReviewed by:
Shikai Liu, Ocean University of China, ChinaKieran G. Meade, Teagasc, The Irish Agriculture and Food Development Authority, Ireland
Copyright © 2018 Nguyen, Reverter, Cánovas, Venus, Anderson, Islas-Trejo, Dias, Crawford, Lehnert, Medrano, Thomas, Moore and Fortes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marina R. S. Fortes, m.fortes@uq.edu.au