 Jing Ouyang1,2,3†
Jing Ouyang1,2,3† John Lin1,2†
John Lin1,2† Stéphane Isnard1,2
Stéphane Isnard1,2 Brandon Fombuena1,2,4Xiaorong Peng1,2,5
Brandon Fombuena1,2,4Xiaorong Peng1,2,5 André Marette6,7Bertrand Routy8,9Meriem Messaoudene8Yaokai Chen3*
André Marette6,7Bertrand Routy8,9Meriem Messaoudene8Yaokai Chen3* Jean-Pierre Routy1,2,10*
Jean-Pierre Routy1,2,10*- 1Infectious Diseases and Immunity in Global Health Program, Research Institute, McGill University Health Centre, Montréal, QC, Canada
- 2Chronic Viral Illness Service, McGill University Health Centre, Montréal, QC, Canada
- 3Chongqing Public Health Medical Center, Chongqing, China
- 4Department of Microbiology and Immunology, McGill University, Montréal, QC, Canada
- 5State Key Laboratory for Diagnosis and Treatment of Infectious Diseases, National Clinical Research Center for Infectious Diseases, Collaborative Innovation Center for Diagnosis and Treatment of Infectious Diseases, The First Affiliated Hospital, College of Medicine, Zhejiang University, Hangzhou, China
- 6Department of Medicine, Faculty of Medicine, Cardiology Axis of the Québec Heart and Lung Institute, Laval University, Laval, QC, Canada
- 7Institute of Nutrition and Functional Foods, Laval University, Laval, QC, Canada
- 8Centre de Recherche du Centre Hospitalier de l’Université de Montréal (CRCHUM), Montréal, QC, Canada
- 9Hematology-Oncology Division, Department of Medicine, Centre Hospitalier de l’Université de Montréal (CHUM), Montréal, QC, Canada
- 10Division of Hematology, McGill University Health Centre, Montréal, QC, Canada
Gut dysbiosis, namely dysregulation of the intestinal microbiota, and increased gut permeability lead to enhanced inflammation and are commonly seen in chronic conditions such as obesity and aging. In people living with HIV (PLWH), several lines of evidence suggest that a depletion of gut CD4 T-cells is associated with gut dysbiosis, microbial translocation and systemic inflammation. Antiretroviral therapy (ART) rapidly controls viral replication, which leads to CD4 T-cell recovery and control of the disease. However, gut dysbiosis, epithelial damage and microbial translocation persist despite ART, increasing risk of developing inflammatory non-AIDS comorbidities such as cardiovascular disease, diabetes mellitus, liver steatosis and cancer. In addition to ART, an emerging research priority is to discover strategies to improve the gut microbial composition and intestinal barrier function. Probiotic interventions have been extensively used with controversial benefits in humans. Encouragingly, within the last decade, the intestinal symbiotic bacterium Akkermansia muciniphila has emerged as the “sentinel of the gut.” A lower abundance of A. muciniphila has been shown in diabetic and obese people as well as in PLWH. Interventions with high levels of polyphenols such as tea or diets rich in fruit, the antibiotic vancomycin and the antidiabetic drug metformin have been shown to increase A. muciniphila abundance, contributing to improved metabolic function in diabetic and obese individuals. We hypothesize that gut microbiota rich in A. muciniphila can reduce microbial translocation and inflammation, preventing occurrences of non-AIDS comorbidities in PLWH. To this aim, we will discuss the protective effect of A. muciniphila and its potential applications, paving the way toward novel therapeutic strategies to improve gut health in PLWH.
Introduction
Gut microbiota is composed of a community of microorganisms gathered in the gastrointestinal (GI) tract. The number of micro-organisms is 1–10 times greater in the GI tract than the number of host cells in humans. Additionally, the number of microbial genes is 100 times greater than that of the human genome (1). In normal, healthy conditions, a state of eubiosis is attained when the composition of the gut microbiota is balanced. The gut microbiota is emerging as a prominent player in maintaining health through several metabolic and immune pathways. Dysregulation of gut microbiota composition, also known as dysbiosis, can be associated with gut barrier dysfunction and intestinal homeostasis disruption through translocation of microbial products and proinflammatory factors (2). Increasing evidence has put a spotlight on the contribution of gut dysbiosis and its related inflammation in obesity, diabetes mellitus (DM), cancer, aging and more recently, human immunodeficiency virus (HIV) infection (3–7).
In people living with HIV (PLWH), intestinal CD4 T-cells are a preferential target of the virus due to their high expression of CCR5, a chemokine co-receptor allowing for the entry of HIV, leading to their massive depletion during early infection (8, 9). This disruption in gut homeostasis results in dysbiosis, microbial translocation and systemic inflammation (10, 11). Antiretroviral therapy (ART) has transformed the lives of PLWH by rapidly controlling viral replication and allowing CD4 recovery, reducing morbidity and mortality. However, despite controlling viral load and CD4 T-cell count, long-term ART reduces but does not normalize gut dysbiosis, microbial translocation, immune activation and inflammation (12–14). In addition to HIV itself, coinfection with cytomegalovirus or viral hepatitis, leaky gut and microbial translocation also lead to inflammation which has been associated with the risk of non-AIDS comorbidities (13, 15–18). The direct influence of dysbiosis, microbiota by-products, epithelial barrier and local immune response will need further studies to define their distinctive role on systemic inflammation and subsequent development of non-AIDS comorbidities. Cardiovascular disease, DM, liver steatosis, neurocognitive disorders and cancer represent the most frequent manifestations of non-AIDS comorbidities, which represent a new frontier in the management of PLWH in today’s medical practice (19–21). Thus, in addition to ART, strategies to improve the gut microbial composition and intestinal barrier function are emerging as a research priority.
Converging evidence has recently demonstrated the key role of commensal bacteria harbored in the GI tract. Interestingly, the bacterium Akkermansia muciniphila has been described as a protective ally against the development of metabolic diseases and colitis (22). A. muciniphila of the phylum Verrucomicrobia, was first isolated and characterized in 2004. This Gram-negative, anaerobic, non-motile, non-spore-forming bacterium has been considered to be a next-generation beneficial microbe (23). In humans, A. muciniphila colonizes the intestinal tract in infanthood and will reach 1–4% of the fecal microbiota by adulthood (24–26). Furthermore, studies have shown a link between low A. muciniphila abundance and increased occurrence of inflammatory metabolic diseases such as diabetes, obesity, ulcerative colitis (UC) and Crohn’s disease (CD), all of which are associated with epithelial gut damage and high permeability (27–35). On the other hand, supplementation with A. muciniphila can help protect from specific metabolic disorders, inflammatory diseases and increase response to cancer immunotherapy (4, 36–43). Moreover, increasing A. muciniphila abundance with the antidiabetic drug metformin or with high polyphenol interventions such as tea or diets rich in fruit further improves metabolic function in diabetic and obese individuals (42, 44–50). The causal or consequential role of A. muciniphila in protection from various diseases in humans remains under debate. Some evidence points toward this symbiotic intestinal bacterium as an emerging “gatekeeper of the gut”, associated with gut barrier integrity and the regulation of inflammation (22, 51, 52). Herein, we discuss recent advances in the understanding of the protective effects of A. muciniphila and its potential relevance in HIV infection.
The Multifunctional Properties of A. Muciniphila
Akkermansia muciniphila encodes a particularly wide repertoire of mucin-degrading enzymes in its relatively small genome, uses mucin as its sole source of carbon and nitrogen, and its downstream glycan byproducts can cross-feed other gut bacteria (23, 53, 54). Based on its unique properties, the bacterium was named after the Dutch microbial ecologist Antoon DL Akkermans for his contributions to the field (55). Additionally, this bacterium exhibits multiple biological functions, including promoting gut barrier integrity, modulating immune response, inhibiting inflammation and cross-feeding, called syntrophy, with other microbiota species.
The gut barrier is organized as a multi-layered and complex system which allows nutrient absorption while preventing the translocation of microbes and their products. Disruption of the gut barrier leads to the transit of luminal contents into the bloodstream, activating the immune response and inducing inflammation (56). Mucus covers the outer intestinal epithelial cell layer and serves as physical protection from penetration of micro-organisms and harmful compounds (57). In addition to degrading mucins, A. muciniphila was also found to stimulate mucin production (42, 52). In animal models, A. muciniphila supplementation increased the thickness of the colonic mucus layer approximately 3-fold, significantly more than the thickness increased induced by the beneficial bacterium Lactobacillus plantarum (52). Furthermore, in vitro, A. muciniphila was found to improve enterocyte monolayer integrity by binding directly to the enterocytes (51). Ottman et al. also showed that the outer membrane protein Amuc_1100 of A. muciniphila improved epithelial cell monolayer integrity in an in vitro culture after 24 h (58).
There is evidence to show that A. muciniphila may regulate inflammation. Supplementation of this bacterium attenuated inflammation in an accelerated aging mouse model (52). Other studies have also shown the anti-inflammatory properties of A. muciniphila in different mouse models including germ-free, liver injury and obesity models (59–64). Huck et al. (62) reported that A. muciniphila could reduce inflammation induced by Porphyromonas gingivalis in lean or obese mice. Ansaldoi et al. (59) demonstrated that A. muciniphila plays a context-dependent role in the induction of gut-resident T-cells during homeostasis in mice. Sessa et al. reported in a cross-sectional study of perinatally HIV-infected children and adolescents that A. muciniphila abundance was associated with elevated IL-6 and soluble CD14 (65).
Additionally, it should be noted that there are also other microbes which are commonly found in the mucus layer aside from A. muciniphila. These microbes include bacteria such as Faecalibacterium prausnitzii, Eubacterium rectale, Roseburia intestinalis, and Anaerostipes caccae which produce the anti-inflammatory short-chain fatty acid (SCFA) butyrate (66–69). Butyrate-producing bacteria do not have the ability to degrade mucus, but use carbon and nitrogen degraded by mucin-degraded species such as A. muciniphila (53). Belzer et al. (66) reported that coculturing A. muciniphila with non-mucus-degrading butyrate-producing bacteria F. prausnitzii, A. caccae, and Eubacterium hallii resulted in syntrophic growth and production of butyrate. Thus, not only does A. muciniphila play an important role by itself in protecting the gut epithelium, but also supports anti-inflammatory intestinal microbiota.
Due to this, and considering its relatively high abundance at all stages of life, A. muciniphila is considered a promising beneficial microbe for some diseases, including metabolic disorders and cancers.
Supplementation of A. Muciniphila in the Context of Metabolic Disorders and Cancers
As a strictly anaerobic bacterium, culture of A. muciniphila needs to be conducted under strict conditions. Advances in the culture and preparation of A. muciniphila have made it feasible for study as a beneficial microbe (36, 70). Supplements of this promising bacterium include live A. muciniphila, pasteurized (killed) A. muciniphila and A. muciniphila-derived extracellular vesicles (AmEVs) (4, 36, 38).
Obesity and metabolic disorders including DM are closely associated with low-grade inflammation and intestinal dysbiosis (71). Everard et al. reported that the abundance of A. muciniphila was 3,300-fold lower in obese mice than in their lean littermates. A 4-week oral gavage of live A. muciniphila in mice reversed high-fat diet-induced metabolic disorders, including fat-mass gain, metabolic endotoxemia, adipose tissue inflammation, and insulin resistance, and increased intestinal levels of endocannabinoids that controlled inflammation, increased gut mucus, and increased expression of gut antimicrobial peptides such as regenerating islet-derived 3-gamma (Reg3γ) for innate immunity (40). In addition, even when A. muciniphila is killed through pasteurization, supplementation demonstrated beneficial effects by protecting from ovariectomy-induced fat mass gain (72). In overweight insulin-resistant humans, a randomized, double-blind, placebo-controlled pilot study showed that daily oral supplementation of 1010 live or pasteurized A. muciniphila bacteria for 3 months was safe and well tolerated, and improved insulin sensitivity, reduced insulinemia, plasma total cholesterol, body weight, fat mass and hip circumference, without great changes in the overall gut microbiota composition (43). Furthermore, AmEV administration was reported to enhance tight junction function, reduce body weight gain and improve glucose tolerance in high-fat diet (HFD)-induced diabetic mice, suggesting that derivatives of the bacterium are sufficient to induce a protective response (38). These findings suggest the direct benefit of this bacterium on the gut barrier and the host metabolism.
Remarkably, the influence of the gut microbiota composition in modulating tumor responses to immunotherapy has also been reported in various cancers such as melanoma, lung and kidney cancer. This effect was observed in different geographic regions where microbiota might differ (North America, Europe, East Asia) (4, 73–75). Reconstitution of germ-free mice with fecal material from lung cancer immunotherapy responders led to increased T-cell responses, and greater efficacy of anti-PD-1 therapy (4). Oral supplementation with live A. muciniphila after fecal microbiota transplantation (FMT) with non-responder feces restored the efficacy of PD-1 blockade in murine models (4).
Although the long term effects of A. muciniphila supplementation are unknown with concerns over the translocation of probiotics (76), this bacterium may play a crucial role in increasing the efficacy of metabolic and cancer therapies and provide strong scientific rationale to launch microbiota-based clinical trials.
Strategies to Increase the Abundance of A. Muciniphila
Supplementation of A. muciniphila may be difficult or costly, however, strategies to indirectly increase the abundance of A. muciniphila exist through dietary interventions, the antidiabetic drug metformin, selective antibiotics and FMT.
Dietary polyphenols are natural antioxidants, which may help protect obligate anaerobes by scavenging oxygen radicals. Gurley et al. reported that administration of green tea to mice, with comparable levels of polyphenols to those consumed by humans, resulted in significant modulation of gut microflora, with the greatest increases observed in A. muciniphila (47). Concord grape, cranberry and the Amazonian fruit Camu Camu have been reported to increase the abundance of A. muciniphila in the intestinal tract approximately 7-fold, 15-fold, and 5-fold, respectively, reduced inflammation and body weight gain, and increased gut barrier integrity in obese mouse models (44, 45, 49). Although a currently unpopular option, caloric restriction such as intermittent fasting has shown increases in A. muciniphila abundance (77). To scale up this approach, diet-mimetic medications are under intense scrutiny. Among the most commonly used in both animals and humans is the anti-diabetic drug metformin.
Metformin is the most commonly used drug to treat DM2 and recently has been shown to reduce inflammation, exert anti-aging effects and modify the gut microbiota composition (78). Although metformin acts primarily as a glucose mediator in the liver by inhibiting hepatic gluconeogenesis, accumulating evidence suggests that metformin also mediates changes in gut microbiota composition (79–81). Convergent reports showed that metformin significantly increased A. muciniphila abundance in animal models (42, 46, 82). The nitrogen-rich structure of metformin may also play a role in the nurturing of A. muciniphila, which requires nitrogen for proliferation and survival (80). Thus, the use of metformin is a strategy to enrich the abundance of A. muciniphila in the gut, among its other metabolic benefits as seen in DM2 (83).
A. muciniphila is resistant to vancomycin, metronidazole, and penicillin (84). Selective antibiotic treatment with vancomycin was shown to dramastically increase A. muciniphila abundance in young non-obese diabetic (NOD) mice, reducing their glucose levels and the diabetes incidence when compared with untreated NOD mice (85). In two patients from the intensive care unit of Marseille, France, broad spectrum antibiotics increased A. muciniphila abundance to more than 40% in stools, without inducing gastrointestinal disorders (84). Furthermore, Uribe-Herranz et al. reported that in pre-clinical models to study the immune-based off-target (abscopal) effect of radiotherapy, oral supplementation with vancomycin increased A. muciniphila which was associated with tumor growth inhibition in mouse models (86). Although further explorations are required in humans, vancomycin treatment appears safe and able to increase A. muciniphila abundance in the gut microbiota.
Fecal microbiota transplantation is also effective in restoring eubiosis in colitis and metabolic diseases. Zhang et al. showed that transplanting fecal bacteria from people with normal glucose tolerance into DM2 mice downregulated levels of fasting blood glucose, postprandial glucose, total cholesterol, triglyceride, and low-density lipoprotein-cholesterol and increased the abundance of A. muciniphila (87). Huang et al. reported that FMT improved gastrointestinal symptoms and alleviated depression and anxiety in irritable bowel syndrome (IBS) patients. Further, gut microbiota analyses revealed that Methanobrevibacter and A. muciniphila were the most abundant fecal microbiota a month after compared to before FMT (88).
These animal models and human epidemiological studies suggest methods to increase A. muciniphila abundance in humans, but efforts to scale up its abundance in PLWH, and in turn improving their gut health and various metabolic factors, are yet unexplored (89).
Leaky Gut and Dysbiosis in PLWH
HIV infection is characterized by a rapid decline in CD4 T-cell count, early gut mucosal damage, and subsequent translocation of microbial products through the now more permeable epithelium (10, 90). Circulating levels of lipopolysaccharide (LPS) and (1→3)-β-D-Glucan (BDG) are two clinically significant markers that assess the level of bacterial and fungal translocation, respectively, of which high levels lead to metabolic endotoxemia (89). Our group and others have shown that LPS and BDG translocation are correlated with immune dysfunction in PLWH and increased risk of non-AIDS comorbidities (91–94). Moreover, we and others have evaluated circulating intestinal fatty acid binding protein (I-FABP) and regenerating islet-derived protein-3α (REG3α) as two gut damage markers in PLWH (14, 95). I-FABP, an intracellular protein constitutively expressed in enterocytes, is released upon cell death and subsequently detected in the blood (96, 97). REG3α is an antimicrobial peptide secreted by intestinal Paneth cells into the gut lumen and upon gut damage, translocates into the blood (14). We observed that these two gut damage markers were correlated with HIV disease progression, microbial translocation and immune activation in PLWH (14). These findings point to the leaky gut as a significant contributor to chronic inflammation and non-AIDS comorbidities in PLWH.
Recently, accumulating evidence has suggested that the gut microbiota is emerging as a prominent player in the regulation of host metabolism and chronic inflammation (98, 99). Bacterial communities residing in the intestine of HIV-infected individuals have been shown to differ from those of individuals not infected with HIV, independently of age, sex and sexual practice (6). Dysbiosis is associated with impaired intestinal barrier activity, impaired mucosal immunity function and worse clinical outcome in PLWH (6, 16, 100, 101). Moreover, A. muciniphila was significantly depleted in ART-naïve and ART-treated PLWH, compared to uninfected controls (101, 102). In one study, Mutlu et al. demonstrated that PLWH had significantly less A. muciniphila abundance regardless of ART, CD4 count or viral load, compared to healthy controls (102). Rocafort et al. confirmed and expanded these results by showing that A. muciniphila abundance was significantly higher in 49 recently infected PLWH and 55 healthy controls compared to 71 chronically infected untreated PLWH. Furthermore, in 27 chronically infected ART-treated PLWH, A. muciniphila abundance was similar to healthy controls (101). These findings suggest that chronic HIV infection leads to progressive depletion of A. muciniphila abundance, and following ART initiation, A. muciniphila abundance returns to levels similar to those of healthy controls. The causative role of A. muciniphila abundance in HIV infection with respect to gut integrity and inflammation needs to be further elucidated.
Hypothesis: A. Muciniphila as a Sentinel for Gut Permeability in PLWH
HIV infection, metabolic disorders and cancer share common features such as chronic inflammation and dysbiosis, which includes the decreased abundance of A. muciniphila in the gut microbiota (4, 40, 71, 101–104). Given this decreased abundance of A. muciniphila in PLWH, and considering the benefits of increasing A. muciniphila abundance in obesity, we hypothesize that A. muciniphila can act as a shield for gut permeability, preventing microbial translocation and reducing inflammation, with the aim toward decreasing risks of developing non-AIDS comorbidities in PLWH. Potential interventions that may increase A. muciniphila abundance in people living with HIV are shown in Figure 1.
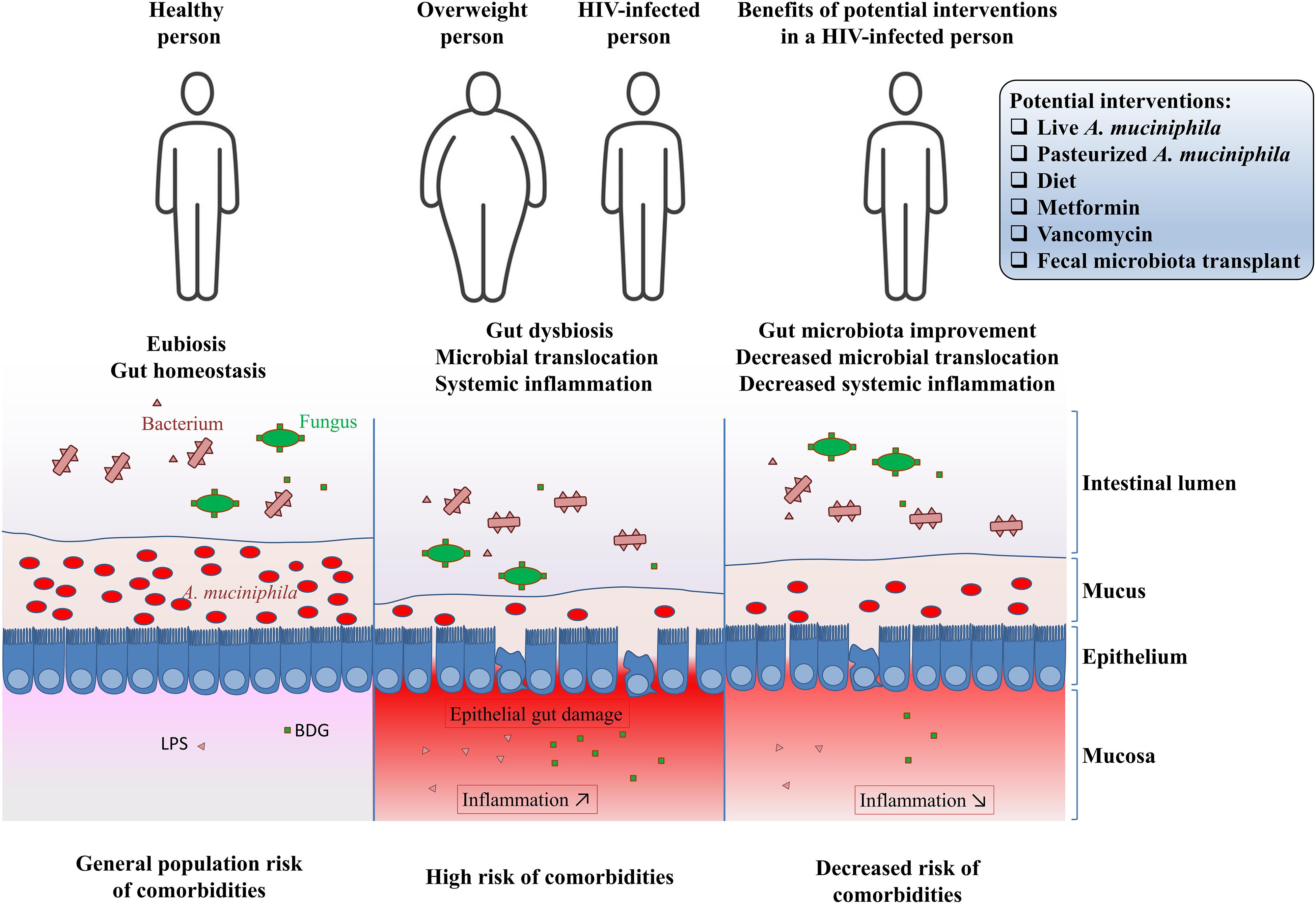
Figure 1. Potential interventions to increase Akkermansia muciniphila abundance in people living with HIV.
Leaky gut has been considered one of the most important factors for microbial translocation and increased inflammation in PLWH (15). In three in vitro human cell line models, Caco-2, HT-29, and TIGK, A. muciniphila was reported to improve enterocyte monolayer integrity and increase the expression of cell–cell adhesion and tight junction molecules (51, 62). Furthermore, in an accelerated aging mouse model, the thickness of the colonic mucus layer increased approximately 3-fold after long-term A. muciniphila supplementation (52). Therefore, we propose that A. muciniphila might decrease inflammation by preserving gut barrier integrity and subsequently preventing microbial translocation in PLWH.
Furthermore, in PLWH, there is a lower abundance of butyrate-producing bacteria (105, 106). Butyrate plays an important role as an energy source for colonic epithelial cells and epithelial barrier integrity, T-cell activation, colonic regulatory T cell differentiation, gut and blood antigen presenting cell (APC) modulation (105–109). Lower abundance of butyrate-producing bacteria has been associated with poor clinical outcome in Crohn’s disease, ulcerative colitis and colon cancer (110, 111). Interestingly, A. muciniphila could promote butyrate-producing bacteria growth and butyrate production (66). We therefore suggest that A. muciniphila, by supporting butyrate-producing bacteria, may also decrease inflammation in PLWH through this method (106, 109).
Moreover, antimicrobial peptides in the gut play a prominent role as host defense effector molecules. Specifically, the C-type lectin REG3α secreted by human Paneth cells and its mouse ortholog REG3γ can bind peptidoglycan and serve as bactericidal agents against Gram-positive species (112). Live A. muciniphila supplementation showed an increased expression of the murine homolog REG3γ in an obese mouse model (40). Moreover, A. muciniphila was reported to induce immunoglobulin G1 (IgG1) antibodies, antigen-specific T-cell responses and intestinal adaptive immune responses (59). Therefore, A. muciniphila may improve intestinal homeostasis through the increased expression of REG3α in Paneth cells and inducing intestinal adaptive immune responses in PLWH.
Conclusion
Epithelial gut damage, microbial translocation and inflammation are considered common determinant mediators of inflammatory non-AIDS comorbidities in PLWH. A. muciniphila has emerged as the “sentinel of the gut” and has been shown to promote gut barrier integrity, modulate immune response, inhibit inflammation and enrich butyrate-producing bacteria. Supplementation of A. muciniphila and other strategies promoting the abundance of A. muciniphila have been proven to be effective in some metabolic disorders and cancer. Recently, clinical trials involving metformin (113), prebiotics (CIHR/CTN NCT04058392) or FMT to increase A. muciniphila abundance have come into fruition, and we suggest that a gut microbiota enriched in A. muciniphila can reduce microbial translocation and inflammation, lowering the risk of developing non-AIDS comorbidities and improving quality-of-life in PLWH.
Author Contributions
JO and JL wrote the first draft of the manuscript. SI, BF, XP, AM, BR, and MM provided critical revision of the manuscript. YC and J-PR conceived and designed the manuscript. All authors approved it for publication.
Funding
This work was funded by the Canadian Institutes of Health Research (CIHR; grants MOP 103230 and PTJ 166049), the Vaccines & Immunotherapy Core of the CIHR Canadian HIV Trials Network (CTN, grant CTN 257 and CTN PT027), the CIHR-funded Canadian HIV Cure Enterprise (CanCURE) Team Grant HB2-164064, and the Chinese National Science and Technology Major Project during the 13th Five-Year Plan (No. 2018ZX10302104). AM holds a CIHR/Pfizer Research Chair in the pathogenesis of insulin resistance and cardiovascular complications was supported by a CIHR Foundation grant.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are highly grateful to Angie Massicotte, Josée Girouard, and Cezar Iovi for coordination and assistance.
References
1. Cornejo-Pareja I, Munoz-Garach A, Clemente-Postigo M, Tinahones FJ. Importance of gut microbiota in obesity. Eur J Clin Nutr. (2019) 72:26–37. doi: 10.1038/s41430-018-0306-8
2. Vinolo MA, Rodrigues HG, Nachbar RT, Curi R. Regulation of inflammation by short chain fatty acids. Nutrients. (2011) 3:858–76. doi: 10.3390/nu3100858
3. Larsen N, Vogensen FK, van den Berg FW, Nielsen DS, Andreasen AS, Pedersen BK, et al. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS One. (2010) 5:e9085. doi: 10.1371/journal.pone.0009085
4. Routy B, Le Chatelier E, Derosa L, Duong CPM, Alou MT, Daillere R, et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science. (2018) 359:91–7. doi: 10.1126/science.aan3706
5. Choi J, Hur TY, Hong Y. Influence of altered gut microbiota composition on aging and aging-related diseases. J Lifestyle Med. (2018) 8:1–7. doi: 10.15280/jlm.2018.8.1.1
6. Vujkovic-Cvijin I, Somsouk M. HIV and the gut microbiota: composition, consequences, and avenues for amelioration. Curr HIV/AIDS Rep. (2019) 16:204–13. doi: 10.1007/s11904-019-00441-w
7. Serino M. Molecular paths linking metabolic diseases, gut microbiota dysbiosis and enterobacteria infections. J Mol Biol. (2018) 430:581–90. doi: 10.1016/j.jmb.2018.01.010
8. Mehandru S, Tenner-Racz K, Racz P, Markowitz M. The gastrointestinal tract is critical to the pathogenesis of acute HIV-1 infection. J Allergy Clin Immunol. (2005) 116:419–22. doi: 10.1016/j.jaci.2005.05.040
9. Brenchley JM, Douek DC. HIV infection and the gastrointestinal immune system. Mucosal Immunol. (2008) 1:23–30. doi: 10.1038/mi.2007.1
10. Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. (2006) 12:1365–71. doi: 10.1038/nm1511
11. Mehraj V, Ramendra R, Isnard S, Dupuy FP, Ponte R, Chen J, et al. Circulating (1–>3)-beta-D-Glucan is associated with immune activation during HIV infection. Clin Infect Dis. (2019) 70:232–41. doi: 10.1093/cid/ciz212
12. Zicari S, Sessa L, Cotugno N, Ruggiero A, Morrocchi E, Concato C, et al. Immune activation, inflammation, and non-AIDS co-morbidities in HIV-infected patients under long-term ART. Viruses. (2019) 11:200. doi: 10.3390/v11030200
13. Ramendra R, Isnard S, Lin J, Fombuena B, Ouyang J, Mehraj V, et al. CMV seropositivity is associated with increased microbial translocation in people living with HIV and uninfected controls. Clin Infect Dis. (2019). doi: 10.1093/cid/ciz1001 [Epub ahead of print].
14. Isnard S, Ramendra R, Dupuy FP, Lin J, Fombuena B, Kokinov N, et al. Plasma levels of C-type lectin REG3alpha and gut damage in people with human immunodeficiency virus. J Infect Dis. (2020) 221:110–21. doi: 10.1093/infdis/jiz423
15. Douek D. HIV disease progression: immune activation, microbes, and a leaky gut. Top HIV Med. (2007) 15:114–7.
16. Vujkovic-Cvijin I, Dunham RM, Iwai S, Maher MC, Albright RG, Broadhurst MJ, et al. Dysbiosis of the gut microbiota is associated with HIV disease progression and tryptophan catabolism. Sci Transl Med. (2013) 5:193ra91. doi: 10.1126/scitranslmed.3006438
17. Gogokhia L, Taur Y, Juluru K, Yagan N, Zhu YS, Pamer E, et al. Intestinal dysbiosis and markers of systemic inflammation in viscerally and generally obese persons living with HIV. J Acquir Immune Defic Syndr. (2020) 83:81–9. doi: 10.1097/qai.0000000000002229
18. Duffau P, Ozanne A, Bonnet F, Lazaro E, Cazanave C, Blanco P, et al. Multimorbidity, age-related comorbidities and mortality: association of activation, senescence and inflammation markers in HIV adults. AIDS (London, England). (2018) 32:1651–60. doi: 10.1097/qad.0000000000001875
19. Hsu DC, Sereti I. Serious Non-AIDS events: therapeutic targets of immune activation and chronic inflammation in HIV infection. Drugs. (2016) 76:533–49. doi: 10.1007/s40265-016-0546-7
20. Brown TT, Cole SR, Li X, Kingsley LA, Palella FJ, Riddler SA, et al. Antiretroviral therapy and the prevalence and incidence of diabetes mellitus in the multicenter AIDS cohort study. Arch Intern Med. (2005) 165:1179–84. doi: 10.1001/archinte.165.10.1179
21. Krahn T, Sebastiani G. Nonalcoholic fatty liver disease in human immunodeficiency virus: the (Not So) new kid on the block? Clin Infect Dis. (2019). doi: 10.1093/cid/ciz929 [Epub ahead of print].
22. de Vos WM. Microbe profile: Akkermansia muciniphila: a conserved intestinal symbiont that acts as the gatekeeper of our mucosa. Microbiology. (2017) 163:646–8. doi: 10.1099/mic.0.000444
23. Derrien M, Vaughan EE, Plugge CM, de Vos WM. Akkermansia muciniphila gen. nov., sp. nov., a human intestinal mucin-degrading bacterium. Int J Syst Evol Microbiol. (2004) 54:1469–76. doi: 10.1099/ijs.0.02873-0
24. Collado MC, Derrien M, Isolauri E, de Vos WM, Salminen S. Intestinal integrity and Akkermansia muciniphila, a mucin-degrading member of the intestinal microbiota present in infants, adults, and the elderly. App Environ Microbiol. (2007) 73:7767–70. doi: 10.1128/AEM.01477-07
25. Cheng J, Ringel-Kulka T, Heikamp-de Jong I, Ringel Y, Carroll I, de Vos WM, et al. Discordant temporal development of bacterial phyla and the emergence of core in the fecal microbiota of young children. ISME J. (2016) 10:1002–14. doi: 10.1038/ismej.2015.177
26. Derrien M, Collado MC, Ben-Amor K, Salminen S, de Vos WM. The Mucin degrader Akkermansia muciniphila is an abundant resident of the human intestinal tract. Appl Environ Microbiol. (2008) 74:1646–8. doi: 10.1128/AEM.01226-07
27. Thomas S, Hoxha K, Alexander W, Gilligan J, Dilbarova R, Whittaker K, et al. Intestinal barrier tightening by a cell-penetrating antibody to Bin1, a candidate target for immunotherapy of ulcerative colitis. J Cell Biochem. (2019) 120:4225–37. doi: 10.1002/jcb.27716
28. Scharl M, Paul G, Weber A, Jung BC, Docherty MJ, Hausmann M, et al. Protection of epithelial barrier function by the Crohn’s disease associated gene protein tyrosine phosphatase n2. Gastroenterology. (2009) 137:2030–40.e5. doi: 10.1053/j.gastro.2009.07.078
29. Png CW, Linden SK, Gilshenan KS, Zoetendal EG, McSweeney CS, Sly LI, et al. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am J Gastroenterol. (2010) 105:2420–8. doi: 10.1038/ajg.2010.281
30. Rajilic-Stojanovic M, Shanahan F, Guarner F, de Vos WM. Phylogenetic analysis of dysbiosis in ulcerative colitis during remission. Inflamm Bowel Dis. (2013) 19:481–8. doi: 10.1097/MIB.0b013e31827fec6d
31. Brahe LK, Le Chatelier E, Prifti E, Pons N, Kennedy S, Hansen T, et al. Specific gut microbiota features and metabolic markers in postmenopausal women with obesity. Nutr Diabetes. (2015) 5:e159. doi: 10.1038/nutd.2015.9
32. Yassour M, Lim MY, Yun HS, Tickle TL, Sung J, Song YM, et al. Sub-clinical detection of gut microbial biomarkers of obesity and type 2 diabetes. Genome Med. (2016) 8:17. doi: 10.1186/s13073-016-0271-6
33. Liu R, Hong J, Xu X, Feng Q, Zhang D, Gu Y, et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat Med. (2017) 23:859–68. doi: 10.1038/nm.4358
34. Cani PD, Everard A. [Akkermansia muciniphila: a novel target controlling obesity, type 2 diabetes and inflammation?]. Med Sci (Paris). (2014) 30:125–7. doi: 10.1051/medsci/20143002003
35. Anhe FF, Pilon G, Roy D, Desjardins Y, Levy E, Marette A. Triggering Akkermansia with dietary polyphenols: a new weapon to combat the metabolic syndrome? Gut Microbes. (2016) 7:146–53. doi: 10.1080/19490976.2016.1142036
36. Plovier H, Everard A, Druart C, Depommier C, Van Hul M, Geurts L, et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat Med. (2017) 23:107–13. doi: 10.1038/nm.4236
37. Hanninen A, Toivonen R, Poysti S, Belzer C, Plovier H, Ouwerkerk JP, et al. Akkermansia muciniphila induces gut microbiota remodelling and controls islet autoimmunity in NOD mice. Gut. (2018) 67:1445–53. doi: 10.1136/gutjnl-2017-314508
38. Chelakkot C, Choi Y, Kim DK, Park HT, Ghim J, Kwon Y, et al. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp Mol Med. (2018) 50:e450. doi: 10.1038/emm.2017.282
39. Li J, Lin S, Vanhoutte PM, Woo CW, Xu A. Akkermansia Muciniphila protects against atherosclerosis by preventing metabolic endotoxemia-induced inflammation in Apoe-/- Mice. Circulation. (2016) 133:2434–46. doi: 10.1161/CIRCULATIONAHA.115.019645
40. Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA. (2013) 110:9066–71. doi: 10.1073/pnas.1219451110
41. Kang CS, Ban M, Choi EJ, Moon HG, Jeon JS, Kim DK, et al. Extracellular vesicles derived from gut microbiota, especially Akkermansia muciniphila, protect the progression of dextran sulfate sodium-induced colitis. PLoS One. (2013) 8:e76520. doi: 10.1371/journal.pone.0076520
42. Shin NR, Lee JC, Lee HY, Kim MS, Whon TW, Lee MS, et al. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut. (2014) 63:727–35. doi: 10.1136/gutjnl-2012-303839
43. Depommier C, Everard A, Druart C, Plovier H, Van Hul M, Vieira-Silva S, et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat Med. (2019) 25:1096–103. doi: 10.1038/s41591-019-0495-2
44. Anhe FF, Roy D, Pilon G, Dudonne S, Matamoros S, Varin TV, et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut. (2015) 64:872–83. doi: 10.1136/gutjnl-2014-307142
45. Roopchand DE, Carmody RN, Kuhn P, Moskal K, Rojas-Silva P, Turnbaugh PJ, et al. Dietary polyphenols promote growth of the gut bacterium Akkermansia muciniphila and attenuate high-fat diet-induced metabolic syndrome. Diabetes. (2015) 64:2847–58. doi: 10.2337/db14-1916
46. Lee H, Ko G. Effect of metformin on metabolic improvement and gut microbiota. Appl Environ Microbiol. (2014) 80:5935–43. doi: 10.1128/AEM.01357-14
47. Gurley BJ, Miousse IR, Nookaew I, Ewing LE, Skinner CM, Jenjaroenpun P, et al. Decaffeinated green tea extract does not elicit hepatotoxic effects and modulates the gut microbiome in lean B6C3F(1) mice. Nutrients. (2019) 11:776. doi: 10.3390/nu11040776
48. Ushiroda C, Naito Y, Takagi T, Uchiyama K, Mizushima K, Higashimura Y, et al. Green tea polyphenol (epigallocatechin-3-gallate) improves gut dysbiosis and serum bile acids dysregulation in high-fat diet-fed mice. J Clin Biochem Nutr. (2019) 65:34–46. doi: 10.3164/jcbn.18-116
49. Anhe FF, Nachbar RT, Varin TV, Trottier J, Dudonne S, Le Barz M, et al. Treatment with camu camu (Myrciaria dubia) prevents obesity by altering the gut microbiota and increasing energy expenditure in diet-induced obese mice. Gut. (2019). 68:453–64. doi: 10.1136/gutjnl-2017-315565
50. Ouyang J, Isnard S, Lin J, Fombuena B, Marette A, Routy B, et al. Metformin effect on gut microbiota: insights for HIV-related inflammation. AIDS Res Ther. (2020) 17:10. doi: 10.1186/s12981-020-00267-2
51. Reunanen J, Kainulainen V, Huuskonen L, Ottman N, Belzer C, Huhtinen H, et al. Akkermansia muciniphila adheres to enterocytes and strengthens the integrity of the epithelial cell layer. Appl Environ Microbiol. (2015) 81:3655–62. doi: 10.1128/AEM.04050-14
52. van der Lugt B, van Beek AA, Aalvink S, Meijer B, Sovran B, Vermeij WP, et al. Akkermansia muciniphila ameliorates the age-related decline in colonic mucus thickness and attenuates immune activation in accelerated aging Ercc1. (-/Delta7) mice. Immun Ageing. (2019) 16:6. doi: 10.1186/s12979-019-0145-z
53. van Passel MW, Kant R, Zoetendal EG, Plugge CM, Derrien M, Malfatti SA, et al. The genome of Akkermansia muciniphila, a dedicated intestinal mucin degrader, and its use in exploring intestinal metagenomes. PLoS One. (2011) 6:e16876. doi: 10.1371/journal.pone.0016876
54. Kosciow K, Deppenmeier U. Characterization of three novel beta-galactosidases from Akkermansia muciniphila involved in mucin degradation. Int J Biol Macromol. (2020) 149:331-340. doi: 10.1016/j.ijbiomac.2020.01.246
55. Naito Y, Uchiyama K, Takagi T. A next-generation beneficial microbe: Akkermansia muciniphila. J Clin Biochem Nutr. (2018) 63:33–5. doi: 10.3164/jcbn.18-57
56. Mu J, Chen Q, Zhu L, Wu Y, Liu S, Zhao Y, et al. Influence of gut microbiota and intestinal barrier on enterogenic infection after liver transplantation. Curr Med Res Opin. (2019) 35:241–8. doi: 10.1080/03007995.2018.1470085
57. McGuckin MA, Linden SK, Sutton P, Florin TH. Mucin dynamics and enteric pathogens. Nat Rev Microbiol. (2011) 9:265–78. doi: 10.1038/nrmicro2538
58. Ottman N, Reunanen J, Meijerink M, Pietila TE, Kainulainen V, Klievink J, et al. Pili-like proteins of Akkermansia muciniphila modulate host immune responses and gut barrier function. PLoS One. (2017) 12:e0173004. doi: 10.1371/journal.pone.0173004
59. Ansaldo E, Slayden LC, Ching KL, Koch MA, Wolf NK, Plichta DR, et al. Akkermansia muciniphila induces intestinal adaptive immune responses during homeostasis. Science. (2019) 364:1179–84. doi: 10.1126/science.aaw7479
60. Wu W, Lv L, Shi D, Ye J, Fang D, Guo F, et al. Protective effect of Akkermansia muciniphila against immune-mediated liver injury in a mouse model. Front Microbiol. (2017) 8:1804. doi: 10.3389/fmicb.2017.01804
61. Derrien M, Van Baarlen P, Hooiveld G, Norin E, Muller M, de Vos WM. Modulation of mucosal immune response, tolerance, and proliferation in mice colonized by the mucin-degrader Akkermansia muciniphila. Front Microbiol. (2011) 2:166. doi: 10.3389/fmicb.2011.00166
62. Huck O, Mulhall H, Rubin G, Kizelnik Z, Iyer R, Perpich JD, et al. Akkermansia muciniphila reduces Porphyromonas gingivalis-induced inflammation and periodontal bone destruction. J Clin Periodontol. (2019) 47:202–12 doi: 10.1111/jcpe.13214
63. Schneeberger M, Everard A, Gomez-Valades AG, Matamoros S, Ramirez S, Delzenne NM, et al. Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci Rep. (2015) 5:16643. doi: 10.1038/srep16643
64. Katiraei S, de Vries MR, Costain AH, Thiem K, Hoving LR, van Diepen JA, et al. Akkermansia muciniphila exerts lipid-lowering and immunomodulatory effects without affecting neointima formation in hyperlipidemic APOE∗3-leiden. CETP Mice. Mol Nutr Food Res. (2019):e1900732. doi: 10.1002/mnfr.201900732 [Epub ahead of print].
65. Sessa L, Reddel S, Manno E, Quagliariello A, Cotugno N, Del Chierico F, et al. Distinct gut microbiota profile in antiretroviral therapy-treated perinatally HIV-infected patients associated with cardiac and inflammatory biomarkers. AIDS (London, England). (2019) 33:1001–11. doi: 10.1097/qad.0000000000002131
66. Belzer C, Chia LW, Aalvink S, Chamlagain B, Piironen V, Knol J, et al. Microbial metabolic networks at the mucus layer lead to diet-independent butyrate and vitamin B12 production by intestinal symbionts. mBio. (2017) 8:e00770-17. doi: 10.1128/mBio.00770-17
67. Van den Abbeele P, Belzer C, Goossens M, Kleerebezem M, De Vos WM, Thas O, et al. Butyrate-producing Clostridium cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. (2013) 7:949–61. doi: 10.1038/ismej.2012.158
68. Lopez-Siles M, Khan TM, Duncan SH, Harmsen HJ, Garcia-Gil LJ, Flint HJ. Cultured representatives of two major phylogroups of human colonic Faecalibacterium prausnitzii can utilize pectin, uronic acids, and host-derived substrates for growth. Appl Environ Microbiol. (2012) 78:420–8. doi: 10.1128/AEM.06858-11
69. Chen L, Wang W, Zhou R, Ng SC, Li J, Huang M, et al. Characteristics of fecal and mucosa-associated microbiota in Chinese patients with inflammatory bowel disease. Medicine (Baltimore). (2014) 93:e51. doi: 10.1097/MD.0000000000000051
70. Ouwerkerk JP, Aalvink S, Belzer C, De Vos WM. Preparation and preservation of viable Akkermansia muciniphila cells for therapeutic interventions. Benef Microbes. (2017) 8:163–9. doi: 10.3920/BM2016.0096
71. Everard A, Lazarevic V, Derrien M, Girard M, Muccioli GG, Neyrinck AM, et al. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes. (2011) 60:2775–86. doi: 10.2337/db11-0227
72. Lawenius L, Scheffler JM, Gustafsson KL, Henning P, Nilsson KH, Collden H, et al. Pasteurized Akkermansia muciniphila protects from fat mass gain but not from bone loss. Am J Physiol Endocrinol Metab. (2020) 318:E480–91. doi: 10.1152/ajpendo.00425.2019
73. Gopalakrishnan V, Spencer CN, Nezi L, Reuben A, Andrews MC, Karpinets TV, et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science. (2018) 359:97–103. doi: 10.1126/science.aan4236
74. Matson V, Fessler J, Bao R, Chongsuwat T, Zha Y, Alegre ML, et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science. (2018) 359:104–8. doi: 10.1126/science.aao3290
75. Zheng Y, Wang T, Tu X, Huang Y, Zhang H, Tan D, et al. Gut microbiome affects the response to anti-PD-1 immunotherapy in patients with hepatocellular carcinoma. J Immunother Cancer. (2019) 7:193. doi: 10.1186/s40425-019-0650-9
76. Yelin I, Flett KB, Merakou C, Mehrotra P, Stam J, Snesrud E, et al. Genomic and epidemiological evidence of bacterial transmission from probiotic capsule to blood in ICU patients. Nat Med. (2019) 25:1728–32. doi: 10.1038/s41591-019-0626-9
77. Ozkul C, Yalinay M, Karakan T. Islamic fasting leads to an increased abundance of Akkermansia muciniphila and Bacteroides fragilis group: a preliminary study on intermittent fasting. Turk J Gastroenterol. (2019) 30:1030–5. doi: 10.5152/tjg.2019.19185
78. Pollak M. The effects of metformin on gut microbiota and the immune system as research frontiers. Diabetologia. (2017) 60:1662–7. doi: 10.1007/s00125-017-4352-x
79. Pryor R, Norvaisas P, Marinos G, Best L, Thingholm LB, Quintaneiro LM, et al. Host-microbe-drug-nutrient screen identifies bacterial effectors of metformin therapy. Cell. (2019) 178:1299–312.e29. doi: 10.1016/j.cell.2019.08.003
80. Rena G, Hardie DG, Pearson ER. The mechanisms of action of metformin. Diabetologia. (2017) 60:1577–85. doi: 10.1007/s00125-017-4342-z
81. Madiraju AK, Qiu Y, Perry RJ, Rahimi Y, Zhang XM, Zhang D, et al. Metformin inhibits gluconeogenesis via a redox-dependent mechanism in vivo. Nat Med. (2018) 24:1384–94. doi: 10.1038/s41591-018-0125-4
82. Zhou ZY, Ren LW, Zhan P, Yang HY, Chai DD, Yu ZW. Metformin exerts glucose-lowering action in high-fat fed mice via attenuating endotoxemia and enhancing insulin signaling. Acta Pharmacol Sin. (2016) 37:1063–75. doi: 10.1038/aps.2016.21
83. Derrien M, Belzer C, de Vos WM. Akkermansia muciniphila and its role in regulating host functions. Microb Pathog. (2017) 106:171–81. doi: 10.1016/j.micpath.2016.02.005
84. Dubourg G, Lagier JC, Armougom F, Robert C, Audoly G, Papazian L, et al. High-level colonisation of the human gut by Verrucomicrobia following broad-spectrum antibiotic treatment. Int J Antimicrob Agents. (2013) 41:149–55. doi: 10.1016/j.ijantimicag.2012.10.012
85. Hansen CH, Krych L, Nielsen DS, Vogensen FK, Hansen LH, Sorensen SJ, et al. Early life treatment with vancomycin propagates Akkermansia muciniphila and reduces diabetes incidence in the NOD mouse. Diabetologia. (2012) 55:2285–94. doi: 10.1007/s00125-012-2564-7
86. Uribe-Herranz M, Rafail S, Beghi S, Gil-de-Gomez L, Verginadis I, Bittinger K, et al. Gut microbiota modulate dendritic cell antigen presentation and radiotherapy-induced antitumor immune response. J Clin Invest. (2020) 130:466–79. doi: 10.1172/JCI124332
87. Zhang PP, Li LL, Han X, Li QW, Zhang XH, Liu JJ, et al. Fecal microbiota transplantation improves metabolism and gut microbiome composition in db/db mice. Acta Pharmacol Sin. (2020): doi: 10.1038/s41401-019-0330-9
88. Huang HL, Chen HT, Luo QL, Xu HM, He J, Li YQ, et al. Relief of irritable bowel syndrome by fecal microbiota transplantation is associated with changes in diversity and composition of the gut microbiota. J Digest Dis. (2019) 20:401–8. doi: 10.1111/1751-2980.12756
89. Anhê FF, Schertzer JD, Marette A. Bacteria to alleviate metabolic syndrome. Nat Med. (2019) 25:1031–3. doi: 10.1038/s41591-019-0516-1
90. Tincati C, Douek DC, Marchetti G. Gut barrier structure, mucosal immunity and intestinal microbiota in the pathogenesis and treatment of HIV infection. AIDS Res Ther. (2016) 13:19. doi: 10.1186/s12981-016-0103-1
91. Jenabian MA, El-Far M, Vyboh K, Kema I, Costiniuk CT, Thomas R, et al. Immunosuppressive tryptophan catabolism and gut mucosal dysfunction following early HIV infection. J Infect Dis. (2015) 212:355–66. doi: 10.1093/infdis/jiv037
92. Ramendra R, Isnard S, Mehraj V, Chen J, Zhang Y, Finkelman M, et al. Circulating LPS and (1–>3)-beta-D-Glucan: a folie a deux contributing to HIV-associated immune activation. Front Immunol. (2019) 10:465. doi: 10.3389/fimmu.2019.00465
93. Hoenigl M. Fungal translocation: a driving force behind the occurrence of Non-AIDS events? Clin Infect Dis. (2020) 70:242–4. doi: 10.1093/cid/ciz215
94. Morris A, Hillenbrand M, Finkelman M, George MP, Singh V, Kessinger C, et al. Serum (1–>3)-beta-D-glucan levels in HIV-infected individuals are associated with immunosuppression, inflammation, and cardiopulmonary function. J Acquir Immune Defic Syndr. (2012) 61:462–8. doi: 10.1097/QAI.0b013e318271799b
95. Cheru LT, Park EA, Saylor CF, Burdo TH, Fitch KV, Looby S, et al. I-FABP is higher in people with chronic HIV than elite controllers, related to sugar and fatty acid intake and inversely related to body fat in people with HIV. Open Forum Infect Dis. (2018) 5:ofy288. doi: 10.1093/ofid/ofy288
96. Al-Saffar AK, Meijer CH, Gannavarapu VR, Hall G, Li Y, Diaz Tartera HO, et al. Parallel changes in harvey-bradshaw index, TNFalpha, and intestinal fatty acid binding protein in response to infliximab in Crohn’s disease. Gastroenterol Res Pract. (2017) 2017:1745918. doi: 10.1155/2017/1745918
97. Adriaanse MP, Tack GJ, Passos VL, Damoiseaux JG, Schreurs MW, van Wijck K, et al. Serum I-FABP as marker for enterocyte damage in coeliac disease and its relation to villous atrophy and circulating autoantibodies. Aliment Pharmacol Ther. (2013) 37:482–90. doi: 10.1111/apt.12194
98. Schretter CE. Links between the gut microbiota, metabolism, and host behavior. Gut Microbes. (2019) 11:245–8. doi: 10.1080/19490976.2019.1643674
99. Tibbs TN, Lopez LR, Arthur JC. The influence of the microbiota on immune development, chronic inflammation, and cancer in the context of aging. Microb Cell. (2019) 6:324–34. doi: 10.15698/mic2019.08.685
100. Ceccarelli G, Statzu M, Santinelli L, Pinacchio C, Bitossi C, Cavallari EN, et al. Challenges in the management of HIV infection: update on the role of probiotic supplementation as a possible complementary therapeutic strategy for cART treated people living with HIV/AIDS. Expert Opin Biol Ther. (2019) 19:949–65. doi: 10.1080/14712598.2019.1638907
101. Rocafort M, Noguera-Julian M, Rivera J, Pastor L, Guillen Y, Langhorst J, et al. Evolution of the gut microbiome following acute HIV-1 infection. Microbiome. (2019) 7:73. doi: 10.1186/s40168-019-0687-5
102. Mutlu EA, Keshavarzian A, Losurdo J, Swanson G, Siewe B, Forsyth C, et al. A compositional look at the human gastrointestinal microbiome and immune activation parameters in HIV infected subjects. PLoS Pathog. (2014) 10:e1003829. doi: 10.1371/journal.ppat.1003829
103. Estrada V, Gonzalez N. Gut microbiota in diabetes and HIV: inflammation is the link. EBioMedicine. (2018) 38:17–8. doi: 10.1016/j.ebiom.2018.11.019
104. Wortelboer K, Nieuwdorp M, Herrema H. Fecal microbiota transplantation beyond Clostridioides difficile infections. EBioMedicine. (2019) 44:716–29. doi: 10.1016/j.ebiom.2019.05.066
105. Gonzalez-Hernandez LA, Ruiz-Briseno MDR, Sanchez-Reyes K, Alvarez-Zavala M, Vega-Magana N, Lopez-Iniguez A, et al. Alterations in bacterial communities, SCFA and biomarkers in an elderly HIV-positive and HIV-negative population in western Mexico. BMC Infect Dis. (2019) 19:234. doi: 10.1186/s12879-019-3867-9
106. Dillon SM, Kibbie J, Lee EJ, Guo K, Santiago ML, Austin GL, et al. Low abundance of colonic butyrate-producing bacteria in HIV infection is associated with microbial translocation and immune activation. AIDS (London, England). (2017) 31:511–21. doi: 10.1097/QAD.0000000000001366
107. Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y, et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. (2011) 331:337–41. doi: 10.1126/science.1198469
108. Hague A, Butt AJ, Paraskeva C. The role of butyrate in human colonic epithelial cells: an energy source or inducer of differentiation and apoptosis? Proc Nutr Soc. (1996) 55:937–43.
109. Wang HB, Wang PY, Wang X, Wan YL, Liu YC. Butyrate enhances intestinal epithelial barrier function via up-regulation of tight junction protein Claudin-1 transcription. Dig Dis Sci. (2012) 57:3126–35. doi: 10.1007/s10620-012-2259-4
110. Segain JP, Raingeard de la Bletiere D, Bourreille A, Leray V, Gervois N, Rosales C, et al. Butyrate inhibits inflammatory responses through NFkappaB inhibition: implications for Crohn’s disease. Gut. (2000) 47:397–403. doi: 10.1136/gut.47.3.397
111. Frank DN, Robertson CE, Hamm CM, Kpadeh Z, Zhang T, Chen H, et al. Disease phenotype and genotype are associated with shifts in intestinal-associated microbiota in inflammatory bowel diseases. Inflamm Bowel Dis. (2011) 17:179–84. doi: 10.1002/ibd.21339
112. Bevins CL, Salzman NH. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Na Rev Microbiol. (2011) 9:356–68. doi: 10.1038/nrmicro2546
Keywords: Akkermansia muciniphila, epithelial gut damage, inflammation, microbial translocation, HIV
Citation: Ouyang J, Lin J, Isnard S, Fombuena B, Peng X, Marette A, Routy B, Messaoudene M, Chen Y and Routy J-P (2020) The Bacterium Akkermansia muciniphila: A Sentinel for Gut Permeability and Its Relevance to HIV-Related Inflammation. Front. Immunol. 11:645. doi: 10.3389/fimmu.2020.00645
Received: 05 February 2020; Accepted: 20 March 2020;
Published: 09 April 2020.
Edited by:
Cristina Ribeiro De Barros Cardoso, University of São Paulo, BrazilReviewed by:
Andy Wullaert, Ghent University, BelgiumZhengxiang He, Icahn School of Medicine at Mount Sinai, United States
Johan Van Weyenbergh, KU Leuven, Belgium
Copyright © 2020 Ouyang, Lin, Isnard, Fombuena, Peng, Marette, Routy, Messaoudene, Chen and Routy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yaokai Chen, yaokaichen@hotmail.com; Jean-Pierre Routy, jean-pierre.routy@mcgill.ca
†These authors have contributed equally to this work