 Annelies Post1*†‡
Annelies Post1*†‡ Berenger Kaboré1,2†‡
Berenger Kaboré1,2†‡ Mike Berendsen1,3†
Mike Berendsen1,3† Salou Diallo2†
Salou Diallo2† Ousmane Traore2†
Ousmane Traore2† Rob J. W. Arts1†
Rob J. W. Arts1† Mihai G. Netea1,4
Mihai G. Netea1,4 Leo A. B. Joosten1Halidou Tinto2,5†
Leo A. B. Joosten1Halidou Tinto2,5† Jan Jacobs6,7†
Jan Jacobs6,7† Quirijn de Mast1†
Quirijn de Mast1† André van der Ven1*†
André van der Ven1*†- 1Department of Internal Medicine, Radboud Centre for Infectious Diseases, Radboud University Medical Centre, Nijmegen, Netherlands
- 2IRSS/Clinical Research Unit of Nanoro (CRUN), Nanoro, Burkina Faso
- 3Bandim Health Project, Institute of Clinical Research, University of Southern Denmark and Odense University Hospital, Odense, Denmark
- 4Department for Genomics & Immunoregulation, Life and Medical Sciences Institute (LIMES), University of Bonn, Bonn, Germany
- 5Institut Supérieur des Sciences de la Santé, Université Nazi Boni de Bobo-Dioulasso, Bobo-Dioulasso, Burkina Faso
- 6Department of Clinical Sciences, Institute of Tropical Medicine, Antwerp, Belgium
- 7Department of Microbiology, Immunology and Transplantation, KU Leuven, Leuven, Belgium
Introduction: Patients with clinical malaria have an increased risk for bacterial bloodstream infections. We hypothesized that asymptomatic malaria parasitemia increases susceptibility for bacterial infections through an effect on the innate immune system. We measured circulating cytokine levels and ex-vivo cytokine production capacity in asymptomatic malaria and compared with controls.
Methods: Data were collected from asymptomatic participants <5 years old with and without positive malaria microscopy, as well as from hospitalized patients <5 years old with clinical malaria, bacteremia, or malaria/bacteremia co-infections in a malaria endemic region of Burkina Faso. Circulating cytokines (TNF-α, IFN-γ, IL-6, IL-10) were measured using multiplex assays. Whole blood from asymptomatic participants with and without positive malaria microscopy were ex-vivo stimulated with S. aureus, E. coli LPS and Salmonella Typhimurium; cytokine concentrations (TNF-α, IFN-γ, IL-1β, IL-6, IL-10) were measured on supernatants using ELISA.
Results: Included were children with clinical malaria (n=118), bacteremia (n=22), malaria and bacteremia co-infection (n=9), asymptomatic malaria (n=125), and asymptomatic controls (n=237). Children with either clinical or asymptomatic malaria had higher plasma cytokine concentrations than controls. Cytokine concentrations correlated positively with malaria parasite density with the strongest correlation for IL-10 in both asymptomatic (r=0.63) and clinical malaria (r=0.53). Patients with bacteremia had lower circulating IL-10, TNF-α and IFN-γ and higher IL-6 concentrations, compared to clinical malaria. Ex-vivo whole blood cytokine production to LPS and S. aureus was significantly lower in asymptomatic malaria compared to controls. Whole blood IFN-γ and IL-10 production in response to Salmonella was also lower in asymptomatic malaria.
Interpretation: In children with asymptomatic malaria, cytokine responses upon ex-vivo bacterial stimulation are downregulated. Further studies are needed to explore if the suggested impaired innate immune response to bacterial pathogens also translates into impaired control of pathogens such as Salmonella spp.
Introduction
Plasmodium falciparum infections are a risk factor for bacterial bloodstream infections (bBSI) (1, 2), especially those caused by gram negative bacteria. Non-typhoidal Salmonellae are among the most commonly found isolates in blood cultures among children in sub-Saharan Africa (SSA) (3–5), both among patients with and without co-occurring malaria infection. The case fatality rate of invasive non-Typhoidal Salmonella (iNTS) bloodstream infection is estimated at 20% (6). Patients with a co-infection malaria and iNTS have an increased risk of mortality compared to patients with only malaria or iNTS infection.
Pediatric iNTS and malaria overlap in terms of geographical distribution, age distribution, and seasonality. Several studies showed that a decreased malaria incidence in a particular region was associated with a decreased iNTS incidence (7, 8). Different mechanisms may underlie the epidemiologic association between malaria and iNTS infections (9). First, the intestinal mucosal barrier function is disturbed during malaria, facilitating invasion and subsequently infection. Second, malaria is reported to cause temporary neutrophil (10) and macrophage dysfunction, and influences the cytokine production capacity of immune cells which could alter susceptibility to iNTS (11). Third, changes in iron homeostasis with increased storage of iron within macrophages contributes to a suitable growing environment for Salmonella infection (12).
So far, studies investigating the association between bBSI and malaria are mostly limited to clinical malaria and animal models. In many malaria-endemic areas, asymptomatic malaria parasitemia is highly prevalent, but it is unknown whether asymptomatic malaria increases the risk for bBSI by changes in the host immune responses (9, 13). In the present study, we analyzed circulating cytokine concentrations in asymptomatic healthy children below the age of five with or without parasitemia and children with acute febrile illness diagnosed with malaria, bBSI or co-infections. Furthermore, we compared cytokine production capacity in asymptomatic healthy children with or without parasitemia as an indicator of the presence of immune tolerance.
Methods
Study Site and Study Objectives
Samples were obtained from two studies conducted between March 2016 and September 2017 at the Clinical Research Unit of Nanoro (CRUN), a rural research facility situated in the central-west region of Burkina Faso, at approximately 85km from the capital city Ouagadougou (14). With an estimated 7.9 million cases per year in 2017, Burkina Faso has one of the highest incidences of malaria in SSA (15). Peak incidences coincide with the rainy season (July-October), but cases occur year round. Most cases are caused by Plasmodium falciparum, but Plasmodium ovale and Plasmodium malariae are also sporadically found (3, 16). Seasonal Malaria Chemoprophylaxis (SMC) for children under 5 years was introduced in 2016.
The objective of this study was to assess the effect of (asymptomatic) malaria on cytokine responses to bacterial pathogens among children below the age of five, since children of this age group are most likely to suffer from severe malaria and iNTS infection. We hypothesized that patients with clinical malaria and asymptomatic malaria have a lower cytokine response to bacterial pathogens compared to patients who are malaria microscopy slide negative. To assess our hypothesis, we aggregated data from two studies simultaneously being carried out at CRUN on respectively a healthy population (ClinicalTrials.gov identifier: NCT03176719) and a population presenting with acute febrile illness (AFI) (ClinicalTrials.gov identifier: NCT02669823). For the current study, all participants from either study who were less than 5 years old, were selected.
The first study was an explorative cross-sectional study among healthy volunteers of 1 year and older living in the Health and Demographic Surveillance System (HDSS) area of Nanoro. The HDSS area of Nanoro covers a population of approximately 65.000 inhabitants distributed over 24 villages. Study methods are described in detail elsewhere (Kaboré, submitted; ClinicalTrials.gov identifier: NCT03176719). Briefly, healthy participants of 1 year and older were randomly selected from the 24 villages of the HDSS catchment area. Informed consent was obtained from their parents or legal guardians. After inclusion, an extensive electronic questionnaire concerning health and demographic data was obtained. Finally, EDTA anti-coagulated and heparinized blood was collected and transported back to CRUN in a cool box for hemocytometry, malaria diagnostics and ex-vivo whole blood stimulation. Supernatants were stored at -80°C and shipped to the Netherlands for cytokine analyses.
The second study was a phase-three diagnostic accuracy study to assess the diagnostic performance of two novel technologies to differentiate malaria, bacterial infections, and viral infections. Study methods are described in detail elsewhere (17). In summary, participants of three months and older presenting with AFI at the referral hospital of Nanoro were asked to participate in the study. After consent was obtained basic clinical and demographic data were registered. After inclusion, EDTA anti-coagulated blood and an aerobic blood culture were obtained from each patient and transported to the laboratory. Malaria diagnostics and blood culture were performed on site. One milliliter whole-blood and plasma samples were stored at -80°C and shipped to the Netherlands for retrospective malaria- and bacterial PCRs, as well as assessment of circulating cytokines.
Laboratory Procedures
Upon arrival at the clinical research laboratory of CRUN, the EDTA blood samples were analyzed using an XN-1000 automated hematology analyzer (Sysmex Corporation, Kobe, Japan). Complete blood counts (CBC) and leukocyte differential were recorded.
Malaria Diagnostics
Thick and thin blood films were made using 10μL EDTA-anticoagulated blood. Slides were stained with 3% Giemsa solution and examined for presence of Plasmodium parasites according to the World Health Organization (WHO) procedures (18). Results were expressed as asexual parasites/µl using the White Blood Cell (WBC) count as measured by hematology analyzer. At least 200 white blood cells were counted. Slides were viewed by two independent microscopists. In case of discrepancies between the readers’ results (e.g. discrepancy between positive and negative slides, a more than 1 log difference in parasite density, or discrepancy in Plasmodium species) the results of a third blinded microscopist was decisive. Malaria diagnostics and hematology analysis were done within 6 hours after sample collection.
Ex-Vivo Whole Blood Stimulations
A heparinized whole blood sample was collected from participants in the cross-sectional study for ex-vivo whole blood stimulations. The whole blood stimulations were done at the research laboratory of CRUN within 12 hours after blood collection. Three stimuli (10ng/ml) Escherichia coli LPS (serotype 055:B5, Sigma-Aldrich), 1x10ˆ6/ml heat killed Staphylococcus aureus (clinical isolate) and 1x10ˆ6/ml heat-killed Salmonella Typhimurium (ST-313 wildtype, obtained from blood culture in the Democratic Republic of the Congo) and a negative control medium (RPMI-1640 Dutch modification (Life Technologies, Carlsbad, California, USA) supplemented with gentamicin 50μg/ml, glutamax 2mM, pyruvate 1mM) were used for each participant.
Blood was diluted at 1/5 in RPMI and distributed over four wells per participant in a final concentration of 500 µl. Each well was stimulated with one of four stimuli at an end concentration of 1x106/ml and incubated at 37°C. After 48 hours, plates were centrifuged at 400g for 5 minutes. Supernatant aliquots of 300 µl were collected and stored in Micronics tubes (Lelystad, The Netherlands) at -80°C. After completion of the trial, all supernatants were transported to Radboudumc Nijmegen on dry ice for batch analysis of cytokine levels. Participants with an IL-1β and TNF-α response in the RPMI control sample were considered contaminated and were therefore excluded from analyses.
Cytokine Measurements in Supernatants After Whole Blood Stimulation
Cytokines were assessed using commercial ELISA kits for human TNF-α, IFN-γ, IL-1β, IL-10 (R&D systems, MN, USA) and human IL-6 (Sanquin, Amsterdam, the Netherlands) according to manufacturer’s instructions. All assays were performed in a timespan of two consecutive days: RPMI and Staphylococcus aureus were measured on day one, Salmonella Typhimurium and Escherichia coli LPS were measured on day two. Samples were allowed to thaw at 4°C one day prior to analysis. Plate set-up was randomized in terms of participant age, presence of asymptomatic malaria and month of storage to decrease batch-effect. Reported cytokine concentrations are based off standard curves which were ran on each plate.
Circulating Cytokines Measurements
Circulating cytokines of patients participating to both studies were batch-tested, using stored EDTA plasma samples. Plate set-up was randomized as previously described. The pro-inflammatory cytokines IL-6, TNF-α, IFN-γ and anti-inflammatory cytokine IL-10 were quantified using the MAGPIX technology (Luminex Corporation, Austin, Texas, USA) with the Human High Sensitivity Cytokine Premixed Magnetic Luminex Performance Assay kits from Bio-Techne (R&D systems). Manufacturer’s recommendations were followed during the assay.
Case Definitions
Asymptomatic malaria was defined as one or more parasites in thick smear malaria microscopy without fever or other acute symptoms. Clinical malaria was defined as febrile patients with one or more malaria parasites in thick smear microscopy and without other possible explanations for the fever. Bacteremia was defined as a blood culture grown with a pathogenic bacterium. Combined malaria and bacteremia was recorded in case of simultaneous blood culture grown with a pathogenic bacterium and one or more malaria parasites in malaria thick smear microscopy. Patients with a combined malaria and bacteremia are hereafter referred to as “co-infection” or “Salmonella co-infection” in case of combined malaria and Salmonella. Anemia was defined by age and sex according to WHO classification. Malnutrition was recorded as patients with a low weight-for-height (wasting) or height-for-age (stunting) according to WHO guidelines.
Statistical Analyses
As appropriate, means or medians were compared using a two-sample t-test, Mann-Whitney U test or Kruskal-Wallis. Differences in ex-vivo stimulated cytokine production between participants with and without asymptomatic malaria were corrected for using a multivariable regression, multicollinearity was assessed using a correlation matrix. Data were log-transformed for analysis in case of non-normal distribution. For normally distributed cytokine measurements a linear regression model was used. A Tobit regression was used for analyses with results outside of the upper or lower quantification limits. The association between cytokine production and parasite density was measured using a Pearson’s correlation or Spearman’s correlation as appropriate. Graphs and figures were created using Graphpad (Graphpad Prism, version 7.00, La Jolla, California, USA). A p-value of.05 was considered statistically significant. Results were corrected for multiple testing using a Bonferroni correction.
Ethical Considerations
Both study protocols were approved by the national health ethics committee of Burkina Faso (ref 2016-01-006 and 2015-01-006 respectively). The diagnostic accuracy study was furthermore approved by the internal review board of IRSS (ref A03-2016/CEIRES) the ethical committee of the Antwerp University Hospital (ref 15/47/492) and the institutional review board of the Institute of Tropical Medicine Antwerp (ref 1029/15).
Results
Baseline Characteristics
A total of 510 participants were included in this study: 361 from the cross-sectional study among healthy participants, and 149 from the clinical diagnostic accuracy study (Figure 1).
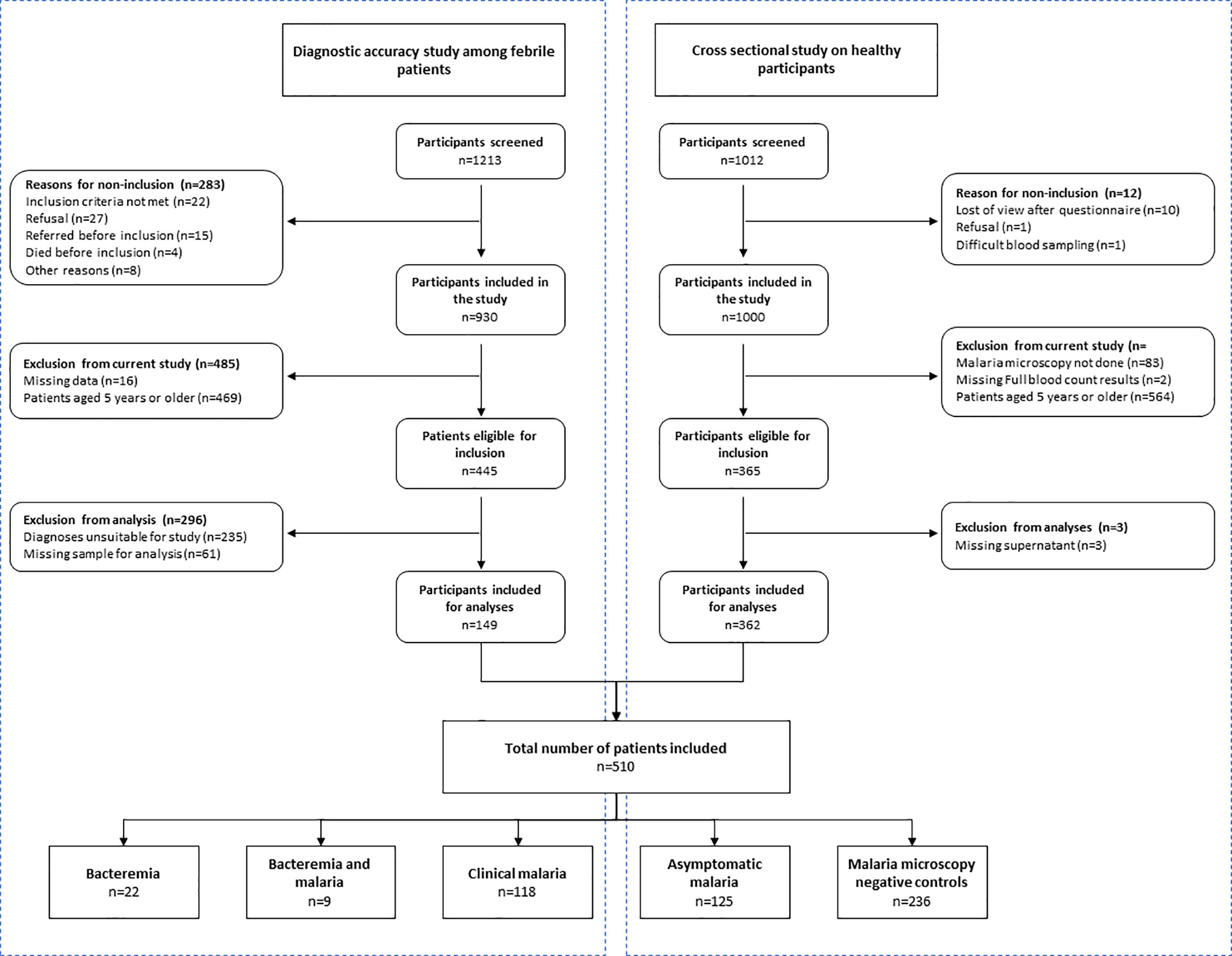
Figure 1 Baseline characteristics of participants included in the current study; derived from participants to the diagnostic accuracy study among febrile patients and the cross-sectional study on healthy participants.
The cross-sectional study enrolled 1000 participants between June and October 2016, of whom 449 (44.6%) had asymptomatic malaria. Out of 399 eligible participants, a total of 361 (90.5%) participants were included in analysis; reasons for exclusion were missing malaria microscopy data (n=33), missing demographic data (n=2) and missing supernatant samples (n=3). Among included participants, 125 (34.6%) had asymptomatic malaria.
The diagnostic accuracy study enrolled 930 patients between March 2015 and July 2016, of whom 461 (49.6%) were between three months and five years old. In total 171 of them were eligible for inclusion in the current study: 171 patients had clinical malaria, 30 patients had bacteremia (Salmonella spp n=18, streptococci n=5, Escherichia coli n=3, Haemophilus influenzae n=2, Neisseria meningitidis n=1 and one double infection) and 9 patients had a co-infection (Salmonella spp. n=6, and one of each Streptococcus pneumoniae, Neisseria meningitidis and Acinetobacter baumannii). Residual blood was available from 149/171 (87.1%) patients; n=118 (69.0%) with clinical malaria, n=22 (73.3%) with bacteremia and n=9 (69.2%) with a co-infection.
Baseline characteristics are presented in Table 1. Among participants from the cross-sectional study, baseline characteristics between participants with asymptomatic malaria and slide negative participants were mostly similar. The number of males was slightly higher among participants with asymptomatic malaria, and participants with asymptomatic malaria had a significantly higher monocyte count (1.1x103 cells versus 0.9x103 cells; p<.0001, lower Hb (10.3 g/dL versus 9.5 g/dL; p<.0001), Reticulocyte count (7.6 versus 10.8; p<.0001) and lower platelet count (382 x103 cells versus 263 x103 cells; p<.0001) compared to slide negative participants, but we found no significant difference in age, height, weight, middle upper arm circumference (MUAC), axillary temperature, white blood cell count or neutrophil count between both groups.
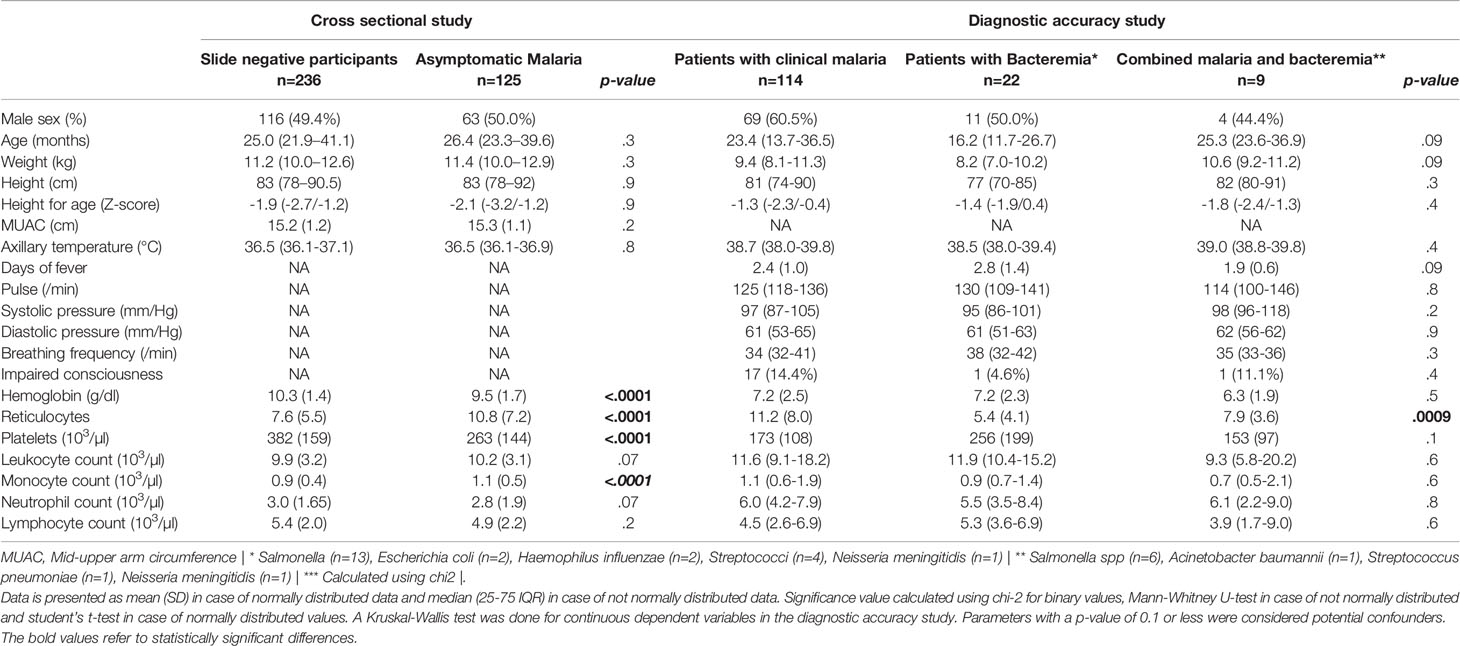
Table 1 Baseline characteristics.
Among participants in the diagnostic accuracy study, differences between patients with clinical malaria, bacteremia and co-infection were more pronounced, though this was also more difficult to interpret due the variability in numbers of cases. Patients with bacteremia were in median less than 1.5 years old, whereas patients with malaria and those with co-infection were approximately 2 years old. The height-for-age among patients with co-infection was considerably lower compared to patients with malaria and patients with bacteremia, suggesting a correlation with chronic malnutrition. Patients with co-infection typically presented at the hospital earlier (1.9 day after onset of fever) compared to patients with clinical malaria (mean 2.4 days) and bacteremia (mean 2.8 days) and had a slightly higher temperature at presentation. As expected, patients from the diagnostic accuracy study had higher neutrophil counts compared to participants to the cross-sectional study. The median numbers of white blood cells were comparable between both groups.
Circulating Cytokines and Correlation to Parasite Density
We first assessed circulating cytokine (IL-6, IL-10, TNF-α, IFN-3γ) concentrations among healthy participants with- and without asymptomatic malaria and patients with clinical malaria. All cytokines were highest among patients with clinical malaria (Table 2) (19, 20) Participants with asymptomatic malaria also had significantly higher circulating cytokines compared to healthy participants without asymptomatic malaria. The median parasite density among patients with clinical malaria was 50,882 parasites/µL (IQR 7,587-104,497) compared to 3,517 parasites/µL (IQR 745-19,282) among participants with asymptomatic malaria (p<.0001). Circulating cytokine concentrations and parasite density significantly correlated for both asymptomatic and clinical malaria (Figure 2). The correlation was strongest for IL 10 (r=0.63 p>.0001 and r=0.53, p<.0001 respectively), followed by IL6 (r=0.59, p<.0001) in asymptomatic malaria, and TNF-α in clinical malaria (r=0.48, p<0.001). The ratio between IL-10 levels and parasite density was 0.008 and 0.01 for clinical and asymptomatic malaria respectively, whereas for IL-6 the ratio was 0.0014 versus 0.00088, and for TNF-α 0.0014 versus 0.010.

Table 2 Circulating cytokine concentrations among malaria slide negative participants, participants with asymptomatic malaria and patient with clinical malaria.
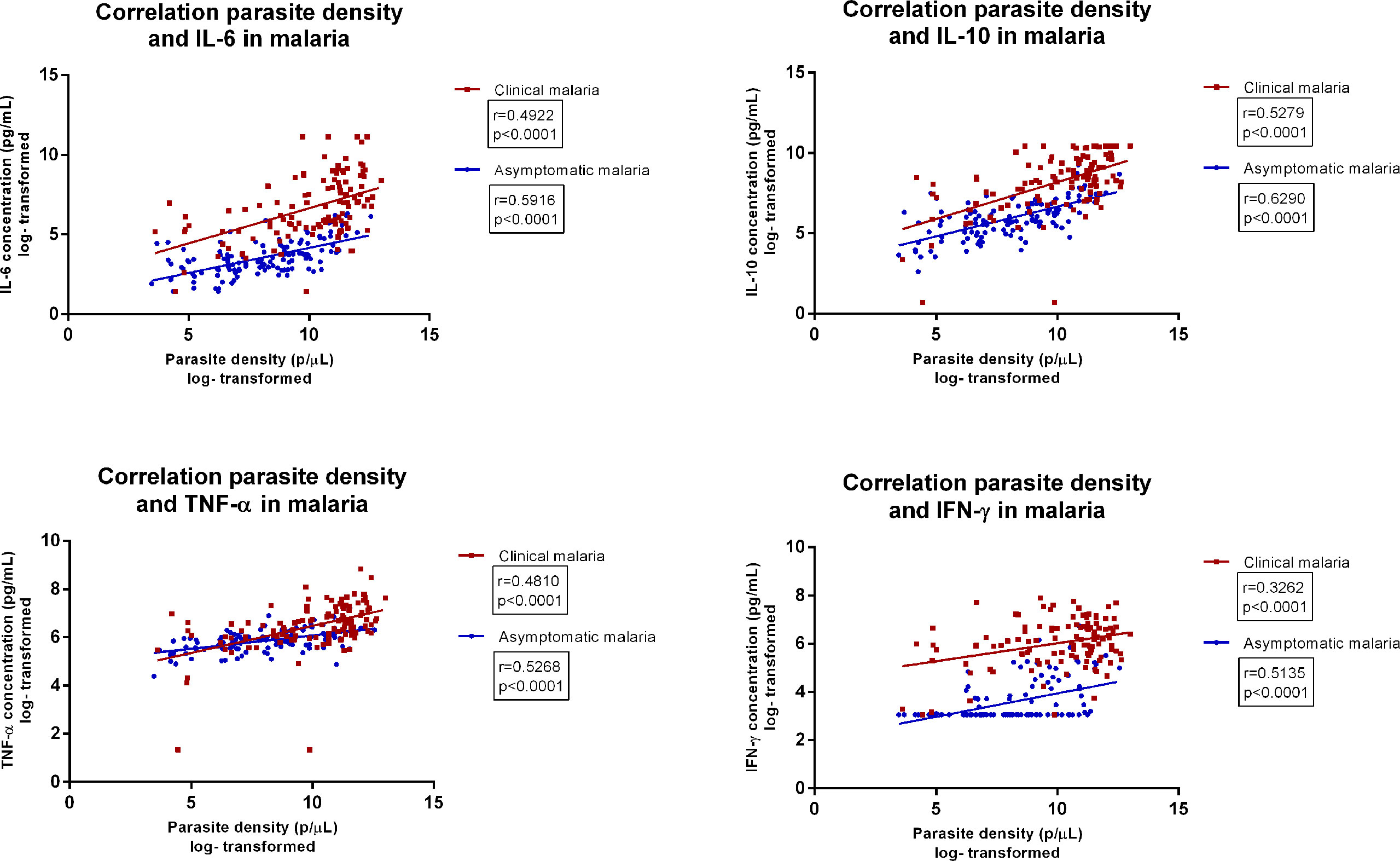
Figure 2 Correlation between parasite density and concentrations of various circulating cytokines in asymptomatic malaria and clinical malaria, among patients below the age of five. Legend | parasite densities reported in parasites/µL. Cytokine concentrations are reported in pg/mL.
These results indicate that apart from IFN-γ, all other tested cytokine concentrations are upregulated in asymptomatic malaria, as in clinical malaria. Cytokine concentrations were significantly higher in clinical malaria, whereas the IL-10 and TNF-α over parasite density ratio were higher in asymptomatic malaria.
We next assessed circulating cytokine concentrations in acute febrile patients below the age of five diagnosed as malaria, bacteremia, or malarial bacteremia co-infection. Results are presented in Table 3 and Figure 3. Patients with bacteremia had significantly lower IL-10 concentrations (p=0.02) compared to those with malaria parasitemia with or without bacteremia co-infection. The other circulating cytokines were not statistically different between groups.

Figure 3 Circulating cytokine concentrations among patients below five years old with blood culture confirmed bacteremia (Gram positive, Gram negative, Salmonella) and malaria-bacteremia co-infection. Legend | Cytokine concentrations reported in pg/mL.
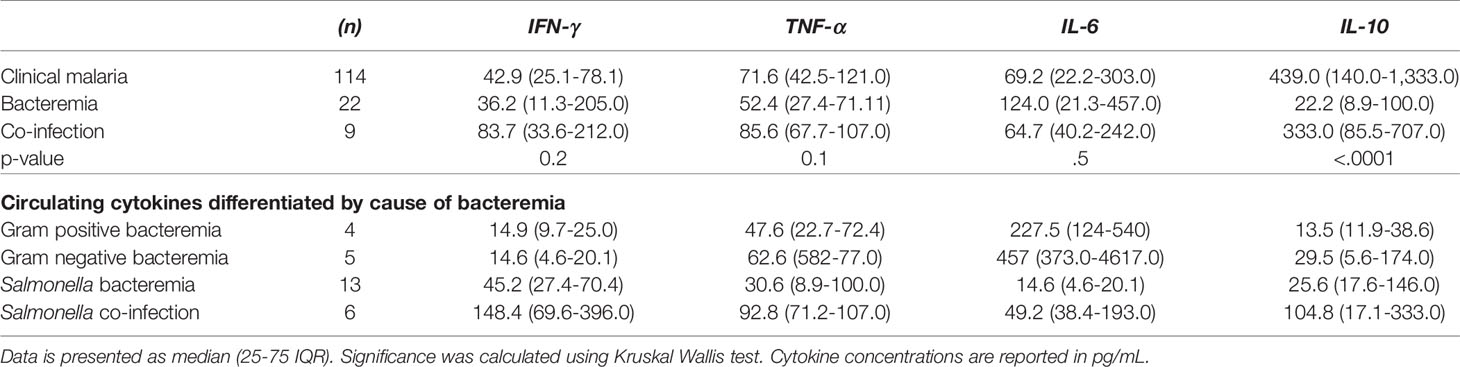
Table 3 Circulating cytokines among patients with clinical malaria, bacteremia, and combined malaria-bacteremia infection.
Further analysis of the different pathogens involved in bacteremia (i.e. Gram positive [Streptococcus pneumoniae, other Streptococci], Gram negative [Escherichia coli, Haemophilus influenzae, Neisseria meningitidis], Salmonella spp.) showed a lower IL-6 concentration among patients with Salmonella bacteremia compared to the patients suffering from infections with other pathogens. Patients with Salmonella bacteremia also had the lowest circulating concentrations of IL-10, but the highest concentration of IFN-γ. There was no significant correlation between parasite density and cytokine levels among patients with co-infection, or Salmonella co-infection. These results suggest that the anti-inflammatory response (IL-10) is upregulated during clinical malaria infections while pro-inflammatory IL-6 (and TNF-α) are particularly downregulated during Salmonella bacteremia.
Ex-vivo Cytokine Production in Participants With Asymptomatic Malaria Versus Slide Negative Controls
To explore whether circulating immune cells of asymptomatic patients with or without malaria parasitemia differed in their capacity to produce cytokines, whole blood was stimulated with various antigens, including heat killed Staphylococcus aureus (S. aureus), heat-killed Salmonella Typhimurium and Escherichia coli LPS (LPS). Samples from 180 (49.7%) participants were excluded due to contamination of the sample during the stimulation assay. The excluded participants are further detailed in Supplementary Tables 1 and 2: there was no significant difference in number of participants with asymptomatic malaria between included and excluded samples (n=67 versus n=59, respectively p=.38).
Table 4 and Figure 4 present the cytokine concentrations of asymptomatic children with or without malaria parasitemia after whole-blood stimulation with S. aureus, Salmonella Typhimurium and LPS, after correction for age, sex, weight, MUAC, leukocyte count, monocyte count, platelet count, hemoglobin levels and ELISA-plate layout to correct for batch effect. Height caused multicollinearity and was therefore not corrected for.
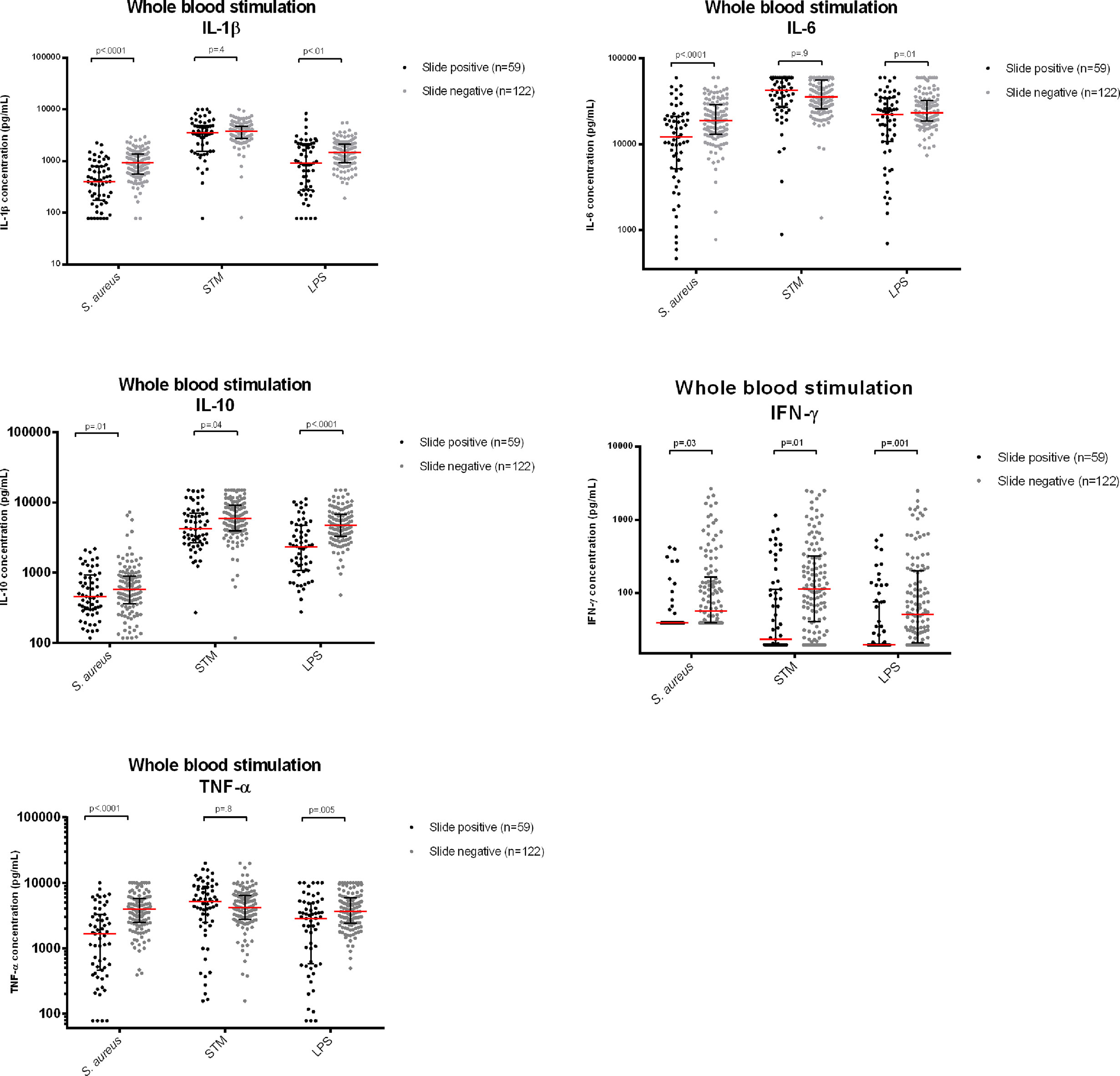
Figure 4 Cytokine concentrations measured in supernatants after whole blood stimulation with Staphylococcus aureus, Salmonella Typhimurium (STM) and Escherichia coli LPS (LPS), comparing participants with asymptomatic Malaria with thick film negative healthy controls. Legend | Cytokine concentrations reported in pg/mL. For comparison, cytokine concentrations were log-converted. P-values corrected for multiple testing using a Bonferroni test.
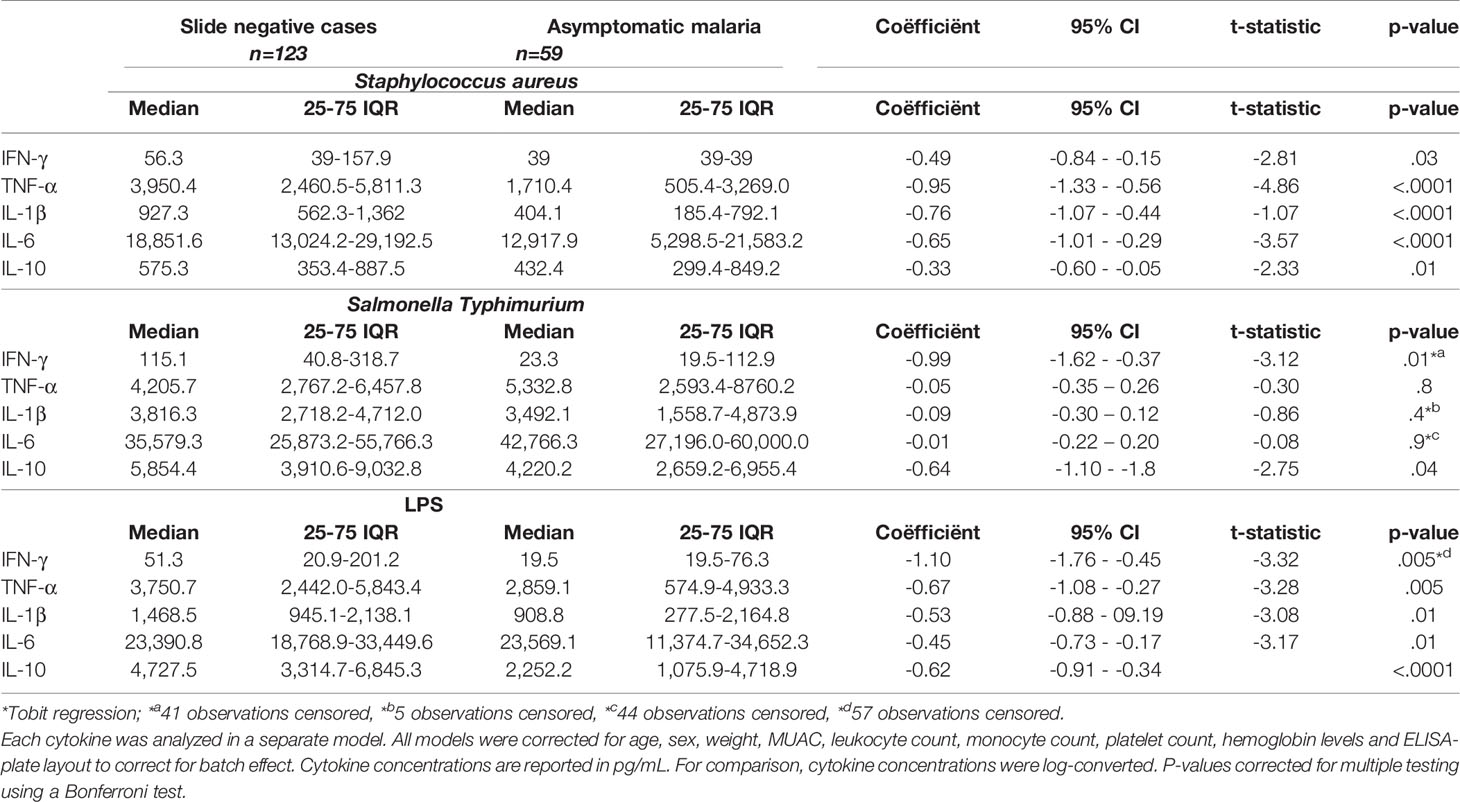
Table 4 Multivariable regression analyses of whole blood stimulated samples among slide negative participants and participants with asymptomatic malaria.
Significantly lower cytokine responses (IL-1β, IL-6, IL-10, TNF-α and IFN-γ) to LPS and Staphylococcus aureus were observed among participants with asymptomatic malaria compared to controls. Participants with asymptomatic malaria had also lower IFN-γ and IL-10 responses after Salmonella Typhimurium stimulation, while monocyte cytokines (IL-1β, IL-6, TNF-α) were not statistically different between slide negative participants and participants with asymptomatic malaria. Likewise, we found that cytokine responses to Salmonella Typhimurium were generally considerably higher compared to the other two stimuli.
In the whole-blood stimulations, parasite density was strongly negatively correlated to production of most cytokines (Figure 5). IL-6, IL-10 and IL-1β in samples exposed to Salmonella Typhimurium did not significantly correlate with parasite densities, which corresponds to the ex-vivo cytokine stimulation results.
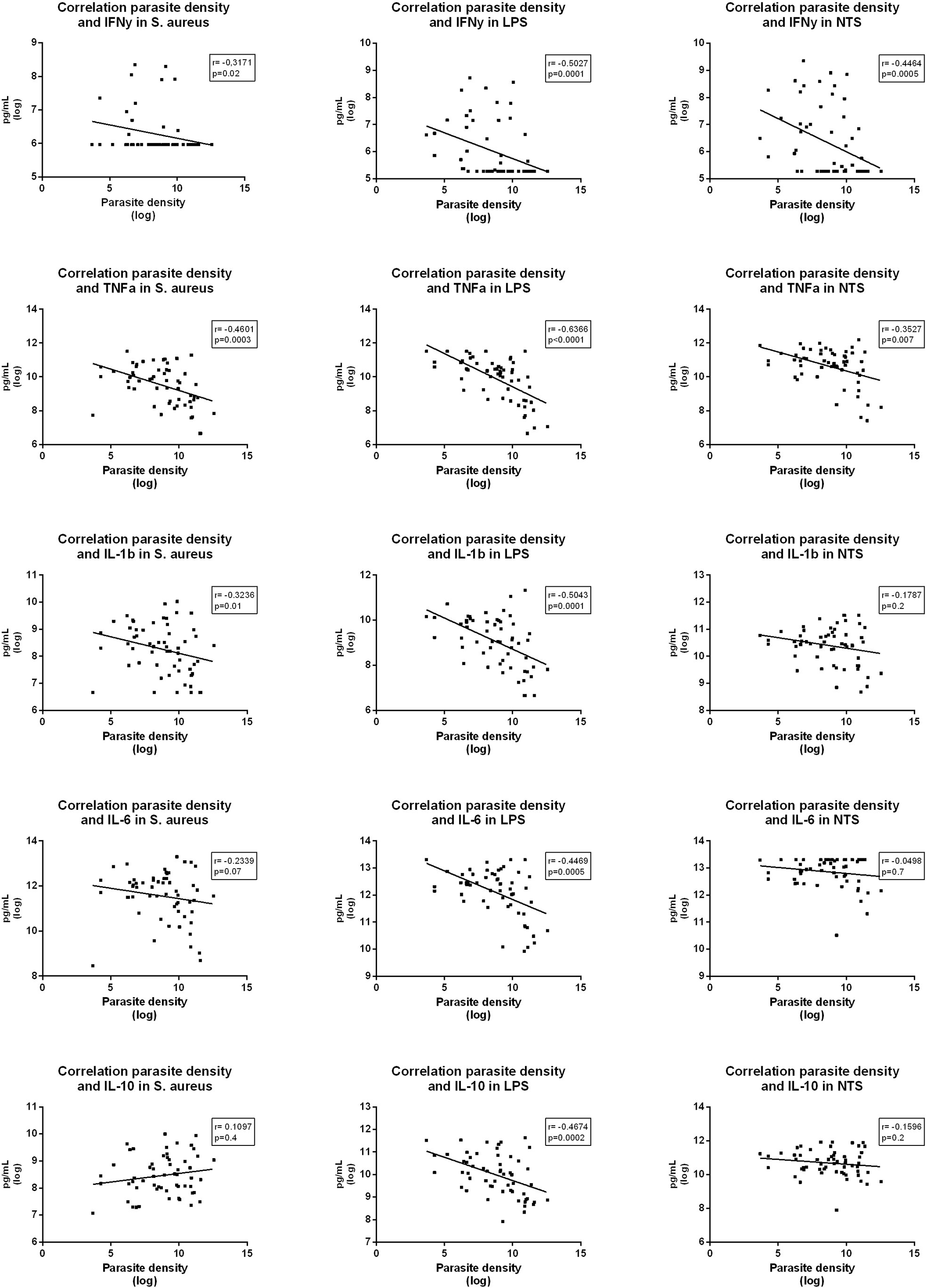
Figure 5 Correlations between parasite density and various cytokine levels measured in supernatants after whole blood stimulation with Staphylococcus aureus, Salmonella Typhimurium (STM) and Escherichia coli LPS (LPS) among participants with asymptomatic malaria.
These results suggest that asymptomatic malaria downregulates the cytokine production capacity of circulating immune cells after stimulation with LPS and heat killed S. aureus, while Salmonella Typhimurium downregulates the mostly lymphocyte-derived IFN-γ but had limited effect on monocyte-derived cytokines.
Discussion
Our findings demonstrate that plasma cytokine concentrations and cytokine production capacity of circulating immune cells upon ex-vivo bacterial stimulation in children with asymptomatic malaria are altered compared to children without asymptomatic malaria. Further studies should be done to explore whether the observed downregulation of pro-inflammatory cytokines impairs the control of bacterial infections, in particular intracellular infections such as invasive Salmonella, confirming the non-benign nature of chronic malaria parasitemia (21, 22).
The study has several strengths and weaknesses. A large number of clinical cases and asymptomatic participants with or without microscopic malaria were included, allowing for comparison of different clinical phenotypes among individuals living in a highly endemic malaria area. A limitation is the number of samples that had to be excluded from ex-vivo cytokine production analyses due to possible contamination. The large number of contaminated cases may be explained by traces op endotoxin in heparinized collecting tubes (23) or blood sampling procedures during the field studies. The effect of excluding these samples on our data was minimal; all analyses were performed while including and excluding these isolates and results were not significantly different, but it is possible that the high number of excluded samples introduced a bias.
The study was carried out in a high malaria endemic area which is both a strength - high number of asymptomatic cases, and a weakness - submicroscopic malaria cases are likely to have been included in the “slide negative” group. The number of these sub-microscopic cases may be significant, as there is evidence of missing on average half of all P. falciparum infections with microscopy in endemic areas compared with PCR (24). On the other hand, excluding the submicroscopic malaria cases is likely to increase the observed differences in cytokine production capacity between asymptomatic malaria and PCR negative asymptomatic cases. A comparison with clinical cases would have been interesting, however ex-vivo stimulation was not done in symptomatic cases.
The introduction of seasonal malaria prophylaxis in 2016 (during our study period) may have influenced results, seeing as part of the clinical malaria samples were collected prior to the introduction of seasonal prophylaxis whereas the asymptomatic cases were collected after introduction.
The amount of blood sampled was too low to perform all analyses in duplicate. Since anemia is common among children in the study site, to sample more blood would have been unethical. However, the fact that we did not use duplicate measurements may have introduced a bias.
The Salmonella serotype used in the stimulation experiments was obtained from DR Congo (4) and not from the study site. However, the strain is the same invasive clade (ST-313 Lineage II) which is also most common in Burkina Faso (25) and is therefore representative for the wild types prevalent at our study site. For the detection of invasive Salmonella infections, blood cultures were used in the clinical cases, which only have an approximate sensitivity of 50% (26).
Several of our findings are in line with existing literature. Parasite densities among patients with clinical malaria were significantly higher compared to those with asymptomatic malaria. Likewise, patients with clinical malaria infection had significantly higher circulating cytokine concentrations compared to participants with asymptomatic malaria. We found a significant positive relation between parasite densities and circulating cytokine levels (IL-10), both in asymptomatic (27) and clinical malaria cases (28, 29). Patients with combined malaria-bacteremia generally had the highest circulating cytokine concentrations compared to patients with malaria or bacteremia alone. Among these patients, cytokine concentrations did not statistically correlate with parasite density, juxtaposed to all other analyses in which cytokine concentrations either positively or negatively correlated to parasite density. Together, these results suggest that once co-infection is established, bacteremia is the primary determinant of cytokine concentrations, and the downregulatory effect of malaria is no longer clinically relevant.
Comparing circulating cytokine concentrations among asymptomatic participants, we also found significantly higher pro- and anti-inflammatory cytokine concentrations in malaria microscopy positive participants compared to microscopy negative controls (30). The largest difference was found for IL-10, confirming the presence and importance of an anti-inflammatory response in asymptomatic malaria to dampen the pro-inflammatory response and prevent the development of clinical disease (31, 32). The cytokine/parasite density ratios also pointed in this direction as the IL-10/parasite density ratio was highest in asymptomatic malaria cases while the pro-inflammatory IL-6/parasite density ratio was highest in patients with clinical malaria. Previous studies demonstrate that high levels of IL-6 as well as TNF-α contribute to complications of malaria such as cerebral malaria (20).
Cytokine concentrations in the supernatant of stimulated whole blood of asymptomatic participants indicate a downregulation of cytokine production capacity of immune cells in asymptomatic participants with positive malaria microscopy compared to those with negative microscopy, consistent with the hypothesis that Plasmodium induces immune hyporesponsivity to bacterial antigenic stimulation. Metenou et al (33) studying filarial infected patients with and without asymptomatic malaria also found evidence using flow cytometry of a lower cytokine production capacity of immune cells in case of concurrent malaria parasitemia. Other previous research focused on clinical malaria, demonstrating that circulating blood samples of clinical malaria patients produced a lower cytokine response (34–36) and contained less TNF-α and IL-6 producing monocytes compared to healthy controls upon stimulation with E. coli LPS (37). These cytokines play a crucial role in host defense and an impaired cytokine production capacity may therefore contribute to the clinically observed increased susceptibility to bBSI in asymptomatic malaria (34, 37), an effect we found to be inversely related to parasite density. While the current study also found a downregulation in cytokine responses, it is difficult to interpret how this would affect clinical outcome without a clinical study. On one hand a downregulation of the early pro-inflammatory cytokines could impair the host response leading to an increased susceptibility to bacterial infections. Alternatively, the lower cytokine concentrations could be sufficient to contain bacterial infection but prevent further damage due to septicemia.
Specific cytokines are known to play a crucial role in the host defense against intracellular pathogens like Salmonella. The early immune response to Salmonella is driven by TNF-α, with the purpose to contain a localized infection (9, 37). IFN-γ, produced by antigen-specific helper T cells, activates macrophages thereby inducing intracellular killing mechanisms against Salmonella (38–40). Simultaneously, Salmonella upregulates the IL-10 production of monocytes, compromising the production of reactive oxygen radicals and intracellular killing capacity of monocytes (41, 42). The relatively low level of IL-6 among patients with iNTS as observed in this study may partially explain why patients with iNTS often present with relatively mild symptoms, as IL-6, together with IL-1 are considered the major endogenous pyrogens (43, 44).
Numerous studies explored the effect of malaria on immune cells, among others the effect of the malaria pigment hemozoin (45). Hemozoin causes release of peroxidation derivatives in mononuclear cells thereby impairing various functions of these cells (46). Hemozoin is detected in mononuclear cells of clinical and asymptomatic malaria cases reflecting the level of parasitemia (47). Mononuclear cells are of special importance to Salmonella as it invades and survives within these cells. It is therefore remarkable that that stimulation with Salmonella Typhimurium among participants with asymptomatic malaria resulted in less IFN-γ and IL-10 production compared to LPS and S. aureus, while no significant difference was observed among TNF-α, IL-1β and IL-6 production capacity. The latter cytokines are primarily produced by monocyte/macrophages while T cells mostly produce IFN-γ. It must be noted that we did not assess which cells were responsible for cytokine production in the ex-vivo experiments and we can therefore not rule out that T-cells may have contributed to the cytokine concentrations in supernatants. Our findings suggest that the downregulation of the IFN-γ axis contributes to the increased risk for iNTS among patients with asymptomatic malaria. The importance of the IFN-γ axis was also observed in other studies assessing genetic risk profile for iNTS (48). A previous study (37) suggests an important role for TNF-α as a risk factor for iNTS in malaria. That study was however done using E. coli LPS instead of heat-killed Salmonella, which may account for the difference in results.
Clinical Implications
Asymptomatic malaria is common in sub-Saharan Africa, with a reported prevalence up to 75% (13). Asymptomatic malaria has long been associated with immunity against malaria while its possible negative consequences are only more recently recognized (21, 22). The observed increase in circulating cytokine concentrations among patients with asymptomatic malaria supports the presence of a pro-inflammatory state with possible consequences on erythropoiesis and decreased hemoglobin levels. Our present data demonstrate that asymptomatic malaria modulates cytokine production capacity of immune cells in response to stimulation with bacterial pathogens that are common in SSA, which may influence the early immune response to bacterial pathogens. A next step in proving this theory may be to compare ex-vivo cytokine concentrations among patients with asymptomatic malaria before and after treatment for malaria. Conversely, the presence of bacteremia may also affect asymptomatic malaria infections: our findings of relatively low circulating levels of IL-10 and IFN-γ in patients with bacteremia indicate a compromised host defense against malaria and thus a less restricted malaria infection. Treatment of asymptomatic malaria may therefore not only reduce the malaria reservoir and malaria transmission, but also protect against the effects of malaria parasitemia on the immune system.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics Statement
Both study protocols were approved by the National Health Ethics Committee of Burkina Faso (ref 2016-01-006 and 2015-01-006, respectively). The diagnostic accuracy study was furthermore approved by the Internal Review Board of IRSS (ref A03-2016/CEIRES), the Ethical Committee of the Antwerp University Hospital (ref 15/47/492), and the Institutional Review Board of the Institute of Tropical Medicine Antwerp (ref 1029/15). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author Contributions
BK, AP, MB, QM, and AV were responsible for study design and execution of the cross-sectional study. AP, BK, SD, HT, JJ, QM, and AV were involved in study design of the diagnostic accuracy study. BK, SD, and OT were responsible for study management in Burkina Faso. RA, MN, and LJ were involved in preparation of laboratory techniques and quality control. Analyses and writing were done by AP, BK, MB, QM, and AV. All authors contributed to the article and approved the submitted version.
Funding
AV and QM have a non-restricted research grant from SYSMEX, which funded the current study. Both the diagnostic accuracy study and the cross-sectional study were funded by a non-restricted research grant from SYSMEX company.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank all study participants for their participation. In addition, we would like to thank the CRUN outreach team, the nurses from CMA, the laboratory technicians from CRUN, the team of data managers from CRUN, and the study nurses from CRUN for their dedication to the study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2021.614817/full#supplementary-material
Supplementary Table 1 | Baseline characteristics of participants with cytokine readout on non-stimulated samples.
Supplementary Table 2 | Multivariable regression analyses of whole blood stimulated samples among slide negative participants and participants with asymptomatic malaria, after correction for monocyte population and plate lay-out (including all cases, also the ones with possible contamination).
References
1. Church J, Maitland K. Invasive Bacterial Co-Infection in African Children With Plasmodium Falciparum Malaria: A Systematic Review. BMC Med (2014) 12:31–1. doi: 10.1186/1741-7015-12-31
2. Were T, Davenport GC, Hittner JB, Ouma C, Vulule JM, Ong’echa JM, et al. Bacteremia in Kenyan Children Presenting With Malaria. J Clin Microbiol (2011) 49:671–6. doi: 10.1128/JCM.01864-10
3. Guiraud I, Post A, Diallo SN, Lompo P, Maltha J, Thriemer K, et al. Population-Based Incidence, Seasonality and Serotype Distribution of Invasive Salmonellosis Among Children in Nanoro, Rural Burkina Faso. PloS One (2017) 12:e0178577. doi: 10.1371/journal.pone.0178577
4. Kalonji LM, Post A, Phoba M-F, Falay D, Ngbonda D, Muyembe J-J, et al. Invasive Salmonella Infections at Multiple Surveillance Sites in the Democratic Republic of the Congo, 2011–2014. Clin Infect Dis (2015) 61:S346–53. doi: 10.1093/cid/civ713
5. Feasey NA, Dougan G, Kingsley RA, Heyderman RS, Gordon MA. Invasive non-Typhoidal Salmonella Disease: An Emerging and Neglected Tropical Disease in Africa. Lancet (2012) 379:2489–99. doi: 10.1016/S0140-6736(11)61752-2
6. Ao TT, Feasey NA, Gordon MA, Keddy KH, Angulo FJ, Crump JA. Global Burden of Invasive Nontyphoidal Salmonella Disease, 2010. Emerg Infect Dis (2015) 21(6):941–9. doi: 10.3201/eid2106.140999
7. Biggs HM, Lester R, Nadjm B, Mtove G, Todd JE, Kinabo GD, et al. Invasive Salmonella Infections in Areas of High and Low Malaria Transmission Intensity in Tanzania. Clin Infect Dis an Off Publ Infect Dis Soc A (2014) 58:638–47. doi: 10.1093/cid/cit798
8. Mtove G, Amos B, Nadjm B, Hendriksen IC, Dondorp AM, Mwambuli A, et al. Decreasing Incidence of Severe Malaria and Community-Acquired Bacteraemia Among Hospitalized Children in Muheza, North-Eastern Tanzania, 2006-2010. Malar J (2011) 10:320. doi: 10.1186/1475-2875-10-320
9. Takem EN, Roca A, Cunnington A. The Association Between Malaria and Non-Typhoid Salmonella Bacteraemia in Children in Sub-Saharan Africa: A Literature Review. Malar J (2014) 13:400. doi: 10.1186/1475-2875-13-400
10. Cunnington AJ, Njie M, Correa S, Takem EN, Riley EM, Walther M. Prolonged Neutrophil Dysfunction Following Plasmodium Falciparum Malaria Is Related to Hemolysis and Heme Oxygenase-1 Induction(). J Immunol (Baltimore Md (1950) (2012) 189:5336–46. doi: 10.4049/jimmunol.1201028
11. Roux CM, Butler BP, Chau JY, Paixao TA, Cheung KW, Santos RL, et al. Both Hemolytic Anemia and Malaria Parasite-Specific Factors Increase Susceptibility to Nontyphoidal Salmonella Enterica Serovar Typhimurium Infection in Mice. Infect Immun (2010) 78:1520–7. doi: 10.1128/IAI.00887-09
12. van Santen S, de Mast Q, Swinkels DW, van der Ven AJ. The Iron Link Between Malaria and Invasive Non-Typhoid Salmonella Infections. Trends Parasitol (2013) 29:220–7. doi: 10.1016/j.pt.2013.03.006
13. Diallo A, Sié A, Sirima S, Sylla K, Ndiaye M, Bountogo M, et al. An Epidemiological Study to Assess Plasmodium Falciparum Parasite Prevalence and Malaria Control Measures in Burkina Faso and Senegal. Malaria J (2017) 16:63. doi: 10.1186/s12936-017-1715-1
14. Derra K, Rouamba E, Kazienga A, Ouedraogo S, Tahita MC, Sorgho H, et al. Profile: Nanoro Health and Demographic Surveillance System. Int J Epidemiol (2012) 41:1293–301. doi: 10.1093/ije/dys159
16. Maltha J, Guiraud I, Kabore B, Lompo P, Ley B, Bottieau E, et al. Frequency of Severe Malaria and Invasive Bacterial Infections Among Children Admitted to a Rural Hospital in Burkina Faso. PloS One (2014) 9:e89103. doi: 10.1371/journal.pone.0089103
17. Post A, Kaboré B, Reuling IJ, Bognini J, van der Heijden W, Diallo S, et al. The XN-30 Hematology Analyzer for Rapid Sensitive Detection of Malaria: A Diagnostic Accuracy Study. BMC Med (2019) 17:103. doi: 10.1186/s12916-019-1334-5
18. World Health Organisation. Universal Access to Malaria Diagnostic Testing – An Operational Manual World Health Organisation. (2013).
19. Artavanis-Tsakonas K, Tongren JE, Riley EM. The War Between the Malaria Parasite and the Immune System: Immunity, Immunoregulation and Immunopathology. Clin Exp Immunol (2003) 133:145–52. doi: 10.1046/j.1365-2249.2003.02174.x
20. Mandala WL, Msefula CL, Gondwe EN, Drayson MT, Molyneux ME, MacLennan CA. Cytokine Profiles in Malawian Children Presenting With Uncomplicated Malaria, Severe Malarial Anemia, and Cerebral Malaria. Clin Vaccine Immunol CVI (2017) 24:e00533–16. doi: 10.1128/CVI.00533-16
21. Chen I, Clarke SE, Gosling R, Hamainza B, Killeen G, Magill A, et al. “Asymptomatic” Malaria: A Chronic and Debilitating Infection That Should Be Treated. PloS Med (2016) 13:e1001942. doi: 10.1371/journal.pmed.1001942
22. de Mast Q, Brouwers J, Syafruddin D, Bousema T, Baidjoe AY, de Groot PG, et al. Is Asymptomatic Malaria Really Asymptomatic? Hematological, Vascular and Inflammatory Effects of Asymptomatic Malaria Parasitemia. J Infect (2015) 71:587–96. doi: 10.1016/j.jinf.2015.08.005
23. van Laarhoven A, Dian S, van Dorp S, Purnama F, Koeken VACM, Diandini E, et al. Immune Cell Characteristics and Cytokine Responses in Adult HIV-Negative Tuberculous Meningitis: An Observational Cohort Study. Sci Rep (2019) 9:884. doi: 10.1038/s41598-018-36696-3
24. Okell LC, Ghani AC, Lyons E, Drakeley CJ. Submicroscopic Infection in Plasmodium Falciparum-Endemic Populations: A Systematic Review and Meta-Analysis. J Infect Dis (2009) 200:1509–17. doi: 10.1086/644781
25. Post AS, Diallo SN, Guiraud I, Lompo P, Tahita MC, Maltha J, et al. Supporting Evidence for a Human Reservoir of Invasive Non-Typhoidal Salmonella From Household Samples in Burkina Faso. PloS Neglect Trop Dis (2019) 13:e0007782. doi: 10.1371/journal.pntd.0007782
26. Dellinger RP, Levy MM, Carlet JM, Bion J, Parker MM, Jaeschke R, et al. Surviving Sepsis Campaign: International Guidelines for Management of Severe Sepsis and Septic Shock: 2008. Crit Care Med (2008) 36:296–327. doi: 10.1097/01.CCM.0000298158.12101.41
27. Ruizendaal E, Schallig HDFH, Bradley J, Traore-Coulibaly M, Lompo P, d’Alessandro U, et al. Interleukin-10 and Soluble Tumor Necrosis Factor Receptor II Are Potential Biomarkers of Plasmodium Falciparum Infections in Pregnant Women: A Case-Control Study From Nanoro, Burkina Faso. Biomark Res (2017) 5:34–4. doi: 10.1186/s40364-017-0114-7
28. Ademolue TW, Aniweh Y, Kusi KA, Awandare GA. Patterns of Inflammatory Responses and Parasite Tolerance Vary With Malaria Transmission Intensity. Malaria J (2017) 16:145. doi: 10.1186/s12936-017-1796-x
29. Oyegue-Liabagui SL, Bouopda-Tuedom AG, Kouna LC, Maghendji-Nzondo S, Nzoughe H, Tchitoula-Makaya N, et al. Pro- and Anti-Inflammatory Cytokines in Children With Malaria in Franceville, Gabon. Am J Clin Exp Immunol (2017) 6:9–20.
30. Portugal S, Moebius J, Skinner J, Doumbo S, Doumtabe D, Kone Y, et al. Exposure-Dependent Control of Malaria-Induced Inflammation in Children. PloS Pathog (2014) 10:e1004079. doi: 10.1371/journal.ppat.1004079
31. Mendonca VR, Barral-Netto M. Immunoregulation in Human Malaria: The Challenge of Understanding Asymptomatic Infection. Mem Inst Oswaldo Cruz (2015) 110:945–55. doi: 10.1590/0074-02760150241
32. Jagannathan P, Eccles-James I, Bowen K, Nankya F, Auma A, Wamala S, et al. Ifnγ/Il-10 Co-Producing Cells Dominate the CD4 Response to Malaria in Highly Exposed Children. PloS Pathog (2014) 10:e1003864. doi: 10.1371/journal.ppat.1003864
33. Metenou S, Dembele B, Konate S, Dolo H, Coulibaly YI, Diallo AA, et al. Filarial Infection Suppresses Malaria-Specific Multifunctional Th1 and Th17 Responses in Malaria and Filarial Coinfections. J Immunol (Baltimore Md1950) (2011) 186:4725–33. doi: 10.4049/jimmunol.1003778
34. Linssen J, Aderhold S, Nierhaus A, Frings D, Kaltschmidt C, Zanker K. Automation and Validation of a Rapid Method to Assess Neutrophil and Monocyte Activation by Routine Fluorescence Flow Cytometry In Vitro. Cytometry Part B Clin Cytometry (2008) 74:295–309. doi: 10.1002/cyto.b.20422
35. Dobaño C, Nhabomba AJ, Manaca MN, Berthoud T, Aguilar R, Quintó L, et al. And Regulatory Cytokine Signature in Young African Children Is Associated With Lower Risk of Clinical Malaria. Clin Infect Dis an Off Publ Infect Dis Soc A (2019) 69:820–8. doi: 10.1093/cid/ciy934
36. Rhee MS, Akanmori BD, Waterfall M, Riley EM. Changes in Cytokine Production Associated With Acquired Immunity to Plasmodium Falciparum Malaria. Clin Exp Immunol (2001) 126:503–10. doi: 10.1046/j.1365-2249.2001.01681.x
37. Mandala WL, Msefula CL, Gondwe EN, Drayson MT, Molyneux ME, MacLennan CA. Monocyte Activation and Cytokine Production in Malawian Children Presenting With P. Falciparum Malaria. Parasite Immunol (2016) 38:317–25. doi: 10.1111/pim.12319
38. Dougan G, John V, Palmer S, Mastroeni P. Immunity to Salmonellosis. Immunol Rev (2011) 240:196–210. doi: 10.1111/j.1600-065X.2010.00999.x
39. MacLennan CA, Gondwe EN, Msefula CL, Kingsley RA, Thomson NR, White SA, et al. The Neglected Role of Antibody in Protection Against Bacteremia Caused by Nontyphoidal Strains of Salmonella in African Children. J Clin Invest (2008) 118:1553–62. doi: 10.1172/JCI33998
40. Gordon MA, Jack DL, Dockrell DH, Lee ME, Read RC. Gamma Interferon Enhances Internalization and Early Nonoxidative Killing of Salmonella Enterica Serovar Typhimurium by Human Macrophages and Modifies Cytokine Responses. Infect Immun (2005) 73:3445–52. doi: 10.1128/IAI.73.6.3445-3452.2005
41. Lokken KL, Mooney JP, Butler BP, Xavier MN, Chau JY, Schaltenberg N, et al. Malaria Parasite Infection Compromises Control of Concurrent Systemic Non-Typhoidal Salmonella Infection Via IL-10-Mediated Alteration of Myeloid Cell Function. PloS Pathog (2014) 10:e1004049. doi: 10.1371/journal.ppat.1004049
42. Jaslow SL, Gibbs KD, Fricke WF, Wang L, Pittman KJ, Mammel MK, et al. Salmonella Activation of STAT3 Signaling by SarA Effector Promotes Intracellular Replication and Production of IL-10. Cell Rep (2018) 23:3525–36. doi: 10.1016/j.celrep.2018.05.072
43. Crump JA, Sjolund-Karlsson M, Gordon MA, Parry CM. Epidemiology, Clinical Presentation, Laboratory Diagnosis, Antimicrobial Resistance, and Antimicrobial Management of Invasive Salmonella Infections. Clin Microbiol Rev (2015) 28:901–37. doi: 10.1128/CMR.00002-15
44. Gilchrist JJ, Heath JN, Msefula CL, Gondwe EN, Naranbhai V, Mandala W, et al. Cytokine Profiles During Invasive Nontyphoidal Salmonella Disease Predict Outcome in African Children. Clin Vaccine Immunol CVI (2016) 23:601–9. doi: 10.1128/CVI.00128-16
45. Taramelli D, Recalcati S, Basilico N, Olliaro P, Cairo G. Macrophage Preconditioning With Synthetic Malaria Pigment Reduces Cytokine Production Via Heme Iron-Dependent Oxidative Stress. Lab Invest (2000) 80:1781–8. doi: 10.1038/labinvest.3780189
46. Bobade D, Khandare AV, Deval M, Shastry P, Deshpande P. Hemozoin-Induced Activation of Human Monocytes Toward M2-Like Phenotype Is Partially Reversed by Antimalarial Drugs-Chloroquine and Artemisinin. MicrobiologyOpen (2019) 8:e00651. doi: 10.1002/mbo3.651
47. Ortega-Pajares A, Rogerson SJ. The Rough Guide to Monocytes in Malaria Infection. Front Immunol (2018) 9:2888–8. doi: 10.3389/fimmu.2018.02888
Keywords: asymptomatic malaria, bacteraemia, Salmonella, iNTS, bloodstream infection
Citation: Post A, Kaboré B, Berendsen M, Diallo S, Traore O, Arts RJW, Netea MG, Joosten LAB, Tinto H, Jacobs J, de Mast Q and van der Ven A (2021) Altered Ex-Vivo Cytokine Responses in Children With Asymptomatic Plasmodium falciparum Infection in Burkina Faso: An Additional Argument to Treat Asymptomatic Malaria? Front. Immunol. 12:614817. doi: 10.3389/fimmu.2021.614817
Received: 07 October 2020; Accepted: 17 May 2021;
Published: 09 June 2021.
Edited by:
Ann M. Moormann, University of Massachusetts Medical School, United StatesReviewed by:
Prasanna Jagannathan, Stanford University, United StatesTuan M. Tran, Indiana University Bloomington, United States
Copyright © 2021 Post, Kaboré, Berendsen, Diallo, Traore, Arts, Netea, Joosten, Tinto, Jacobs, de Mast and van der Ven. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Annelies Post, annelies.post@gmail.com; André van der Ven, andre.vanderven@radboudumc.nl
†ORCID:Annelies Post, orcid.org/0000-0002-0471-8250 Berenger Kaboré, orcid.org/0000-0002-3719-7033 Mike Berendsen, orcid.org/0000-0001-9516-6190 Salou Diallo, orcid.org/0000-0002-1253-4726 Ousmane Traore, orcid.org/0000-0003-1873-6473 Rob J. W. Arts, orcid.org/0000-0002-6649-7190 Halidou Tinto, orcid.org/0000-0003-4653-7041 Jan Jacobs, orcid.org/0000-0002-8657-6777 Quirijn de Mast, orcid.org/0000-0001-6056-157 André van der Ven, orcid.org/0000-0003-1833-3391
‡These authors have contributed equally to this work