 Xing Wang
Xing Wang Qian Liu
Qian Liu He Zhang
He Zhang Xia Li
Xia Li Weichun Huang
Weichun Huang Qihua Fu
Qihua Fu Min Li
Min Li- 1Department of Laboratory Medicine, Shanghai Children’s Medical Center, Shanghai Jiao Tong University School of Medicine, Shanghai, China
- 2Department of Laboratory Medicine, Renji Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, China
- 3Department of Emergency, Hebei Provincial Hospital of Traditional Chinese Medicine, Hebei, China
- 4Department of Medical Microbiology and Immunology, University of California, Davis, Davis, CA, United States
Staphylococcus aureus is known as an invasive human pathogen, resulting in significant morbidity and mortality worldwide; however, information on community-associated S. aureus (CA-SA) from bloodstream infections (BSI) in children in China remains scarce. This study aimed to investigate the molecular characteristics of 78 CA-SA isolates recovered from pediatric patients with BSI between 2012 and 2017 in Shanghai. All isolates including 51 (65.4%) methicillin-susceptible S. aureus (MSSA) and 27 (34.6%) methicillin-resistant S. aureus (MRSA) isolates were characterized based on antimicrobial resistance, virulence genes, multilocus sequence typing (MLST), spa, and SCCmec typing. A total of 18 distinct sequence types (STs) and 44 spa types were identified. ST188 and ST7 were the predominant MSSA clones and ST59-MRSA-SCCmecIV/V was the most common MRSA clone. Spa t189 (9.0%, 7/78) was the most common spa type. SCCmec types IV and V were observed at frequencies of 59.3 and 40.7%, respectively. Notably, 40 (51.3%) S. aureus BSI strains were multidrug resistant (MDR), and these were mostly resistant to penicillin, erythromycin, and clindamycin. MRSA strains were associated with substantially higher rates of resistance to multiple antibiotics than MSSA strains. Fifty (64.1%, 50/78) isolates, including 19 (70.3%) MRSA isolates, harbored ≥ 10 tested virulence genes, as evaluated in this study. Ten (37.0%) MRSA isolates and four (7.8%) MSSA isolates harbored the gene encoding Panton–Valentine leukocidin (PVL). Virulence genes analysis showed diversity in different clones; the seb-sek-seq genes were present in all ST59 strains, whereas the seg-sei-sem-sen-seo genes were present in different clones including ST5, ST20, ST22, ST25, ST26, ST30, ST121, and ST487 strains. In conclusion, this study revealed that community-associated S. aureus strains from BSI in children demonstrated considerable genetic diversity, and identified major genotypes of CA-MRSA and CA-MSSA, with a high prevalence of CA-MRSA. Furthermore, major genotypes were frequently associated with specific antimicrobial resistance and toxin gene profiles. Understanding the molecular characteristics of those strains might provide further insights regarding the spread of BSI S. aureus among children between communities in China.
Introduction
Staphylococcus aureus can cause a wide variety of diseases ranging from mild skin and soft-tissue infections to severe systemic infections in human and animals (Lowy, 1998). Serious S. aureus infections such as bacteremia are generally associated with high morbidity and mortality, and the acquisition of methicillin resistance further limits therapeutic options (Cosgrove et al., 2003; Wang et al., 2008; Bassetti et al., 2012). Since methicillin-resistant S aureus was first identified at a hospital in the United Kingdom in 1961, it quickly became an important pathogen globally (referred to as hospital-associated MRSA; HA-MRSA). Until the late 1980s and early 1990s, some cases of MRSA in young and otherwise healthy patients without hospital-related risk factors were reported. These isolates were called community-associated MRSA (CA-MRSA) and their emergence and spread increased the risk to public health.
Compared to traditional HA-MRSA strains, CA-MRSA isolates harbor different types of SCCmec elements encoding methicillin(mec) resistance genes. To date, 13 SCCmec types (indicated by roman numerals I to XIII) and three mec(mecA/B/C) genes have been identified among S. aureus in the world (Hiramatsu et al., 2013; Kaya et al., 2018). The majority of CA-MRSA isolates have SCCmec type IV or V, do not exhibit resistance to multiple antibiotics (except to β-lactams), and possess different exotoxin gene profiles (Dinges et al., 2000). Within the last decade, rates of CA-MRSA infection have been increasing, while HA-MRSA infection rates have generally declined (David et al., 2014). The epidemiological impact of CA-MRSA strains is believed to stem from a combination of methicillin resistance and extraordinary virulence, allowing these strains to infect otherwise healthy individuals and spread rapidly throughout the population. As a result, there is an urgent need to understand the molecular characteristics of CA-MRSA isolates to achieve more effective infection control.
Community-associated-Methicillin-resistant S. aureus strains are considered more virulent than HA-MRSA strains because they possess specific virulence factors. The increased expression of various genes has also been associated with increased virulence in CA-MRSA (Otto, 2013). For example, Panton–Valentine leukocidin (pvl), a bicomponent leukotoxin virulence factor, has been linked to severe skin and soft tissue infections and necrotizing pneumonia caused by CA-MRSA strains (Lina et al., 1999), but has also been found in HA-MRSA and CA-MSSA isolates. The superantigen exotoxin (TSST-1), exfoliatin A (ETA) and B (ETB), staphylococcal enterotoxins (SEs) found to be closely related to different types of staphylococcal infections (Hanakawa et al., 2002; Jarraud et al., 2002; Warner and Onderdonk, 2004). However, the significance of these factors in CA-SA bacteremia is not well understood.
It has been found that various CA-SA clones, and especially CA-MRSA, circulate in different countries or regions. For example, ST1-IV and ST8-IV clones are mainly found in the United States and Canada, whereas ST80-IV clones are more prevalent in Europe and ST59-IV/V are the most common CA-MRSA clones in China and several other Asian countries. The prevalence of CA-SA varies with age, geography, disease, and time, and the occurrence rate of CA-MRSA varies substantially worldwide, ranging from < 1% to > 50% in different countries (Deurenberg and Stobberingh, 2008; Chen and Huang, 2014). The incidence of CA-SA infections is higher in children (Huang and Chen, 2011). Although CA-SA infection and transmission has become a serious public health problem worldwide (David et al., 2015; McMullan et al., 2016; Pena Amaya et al., 2017; Kang et al., 2017; Roediger et al., 2017), the information regarding CA-SA associated with bloodstream infections (BSIs) in children in China is still very limited. The aim of this study was to investigate the molecular profile, antimicrobial resistance, and virulence genes associated with 78 CA-SA isolates recovered from pediatric patients with BSIs between 2012 and 2017 in a hospital in China.
Materials and Methods
Bacterial Isolates
From July 2012 to February 2017, 78 non-duplicate community-associated S. aureus isolates were collected from pediatric patients (< 18 years old) with BSIs in a university hospital (Shanghai Children’s Medical Center) in Shanghai. Shanghai Children’s Medical Center (SCMC), affiliated with Shanghai Jiao Tong University, is one of the largest pediatric hospitals in China with 604 beds and approximately 6,000 hospital admissions per day.
Staphylococcus aureus isolates were confirmed by classic microbiological methods including Gram stain and catalase and coagulase activity with rabbit plasma. They were further identified by biochemical characterization using the API-Staph test (bioMérieux, Lyon, France). MRSA isolates were initially identified using cefoxitin screening and the presence of the mecA gene was confirmed by PCR (Kondo et al., 2007). CA-MRSA was defined as an MRSA isolate that was obtained either from an outpatient or an inpatient within 48 h of hospitalization, and without the patient having a medical history of MRSA infection or colonization, admission to a healthcare facility, dialysis, surgery or the insertion of indwelling devices in the past year (Skov et al., 2012). These isolates were processed in Class II Biological Safety Cabinets. All strains were stored at -70°C and grown overnight on sheep blood agar plates at 37°C.
This study was approved by the Ethics Committee of SCMC, and the Review Board exempted the need for written informed consent because it mainly focused on bacteria and patient intervention did not occur.
Antimicrobial Susceptibility Testing
The antibiotic susceptibility profiles of all S. aureus isolates in the current study were performed using the bioMérieux VITEK2 system following manufacturer’s instructions. Results were interpreted according to the recommendations and definitions of the Clinical and Laboratory Standards Institute [CLSI] (2015). The following 16 drugs were tested: cefoxitin (FOX), linezolid (LZD), ciprofloxacin (CIP), clindamycin (DA), erythromycin (E), trimethoprim-sulfamethoxazole (SXT), moxifloxacin (MOF), vancomycin (V), tetracycline (TET), penicillin (P), rifampicin (RF), levofloxacin (LVX), oxacillin (OXA), gentamicin (GM), quinupristin/dalfopristin (Q/D), and tigecycline (TGC). S. aureus ATCC 29213 was used for quality control.
Multilocus Sequence Typing (MLST) Analysis
All S. aureus isolates were screened according to the protocol described on the S. aureus MLST website1 (Enright and Spratt, 1999; Aanensen and Spratt, 2005). PCR amplicons of seven S. aureus housekeeping genes were obtained from chromosomal DNA. DNA was extracted as previously described (Hartmann et al., 1997). The sequences of the PCR products were compared to those of the existing alleles available from the MLST website, and the allelic number (sequence type, ST) was determined for each sequence. Clustering of related STs that were defined as cloned complexes (CCs) was performed using the eBURST (Based Upon Related Sequence types) algorithm.
SCCmec Typing
Methicillin-resistant S. aureus isolates were subjected to SCCmec typing as described by Kondo et al. (2007), which is based on a set of multiplex PCR reactions with 14 primers. SCCmec types I–V were assigned according to the combination of the cassette chromosome recombinase (ccr) type and mec class. MRSA isolates that could not be assigned to any expected type were defined as non-typable (NT).
spa Typing
In S. aureus, the polymorphic X region of the staphylococcal protein A-encoding (spa) gene was amplified and sequenced as described previously (Shopsin et al., 1999; Koreen et al., 2004). spa typing was assigned by submitting the data to the S. aureus spa type database2.
Detection of Virulence Genes
All S. aureus isolates were screened for the following 33 staphylococcal virulence genes: staphylococcal enterotoxin genes (sea, seb, sec, sed, see, seg, seh, sei, sej, sel, sem, sen, seo, sep, seq, sek), toxic shock syndrome toxin (tsst1), arginine catabolic mobile gene (arcA), exfoliative toxin genes (eta, etb), leukocidin (lukF/S-PV, lukE, lukM) (Lina et al., 1999), bacteriocin (bsaA), hemolysin genes (hla, hlb, hlg, hlg2), and adhesin genes (clfA, icaA, sdrC, sdrD, and sdrE), as previously described (Arvidson and Tegmark, 2001; Peacock et al., 2002; Wardenburg et al., 2007).
The amplification was carried out on a GeneAmp 9700 thermal cycler (Applied Biosystems, NY, United States) under the following conditions: an initial 5 min denaturation at 94°C, followed by 35 cycles of 30 s at 94°C, 30 s at 55°C, and 30 s at 72°C, with a final extension at 72°C for 7 min. In each PCR, S. aureus isolates harboring virulence genes determined by our previous study were used as positive control strains (Wang et al., 2016) and distilled water was used for a negative control. The PCR fragments were visualized by agarose gel electrophoresis and ethidium bromide staining.
Statistical Analysis
Statistical analyses were performed using Stata software (version 10.1/SE, Stata Corp, College Station, TX, United States), using χ2 and Fisher’s exact tests, as appropriate for the analysis of categorical data. Statistical significance was set at P ≤ 0.05.
Results
Prevalence of CA-SA From Pediatric Patients With BSIs
From blood culture samples during 2012–2017, we found positive rate of CA-SA to be 0.2076% (13/6261) in 2013, 0.1223% (10/8174) in 2014, 0.1696% (15/8845) in 2015, and 0.2135% (21/9834) in 2016 among pediatric patients. Laboratory-based surveillance indicated a relatively stable number of CA-SA BSI infections in our hospital, with the number of cases increasing from 2014 to 2016 over time.
MLST, SCCmec, and spa Typing
The evolutionary and genetic diversity of 78 S. aureus isolates from BSIs in children was analyzed by MLST (Table 1). There were 18 distinct STs identified within the 78 isolates, of which the most frequently represented were ST59 (15%, 12/78) and ST188 (15%, 12/78), accounting for nearly one third of all S. aureus isolates, followed by ST7 (14%, 11/78), ST398 (12%, 9/78), ST88 (7.7%, 6/78), ST5 (7.7%, 6/78), ST6 (6.4%, 5/78), and ST1 (5.1%, 4/78). Other STs accounted for one or two isolates, respectively.
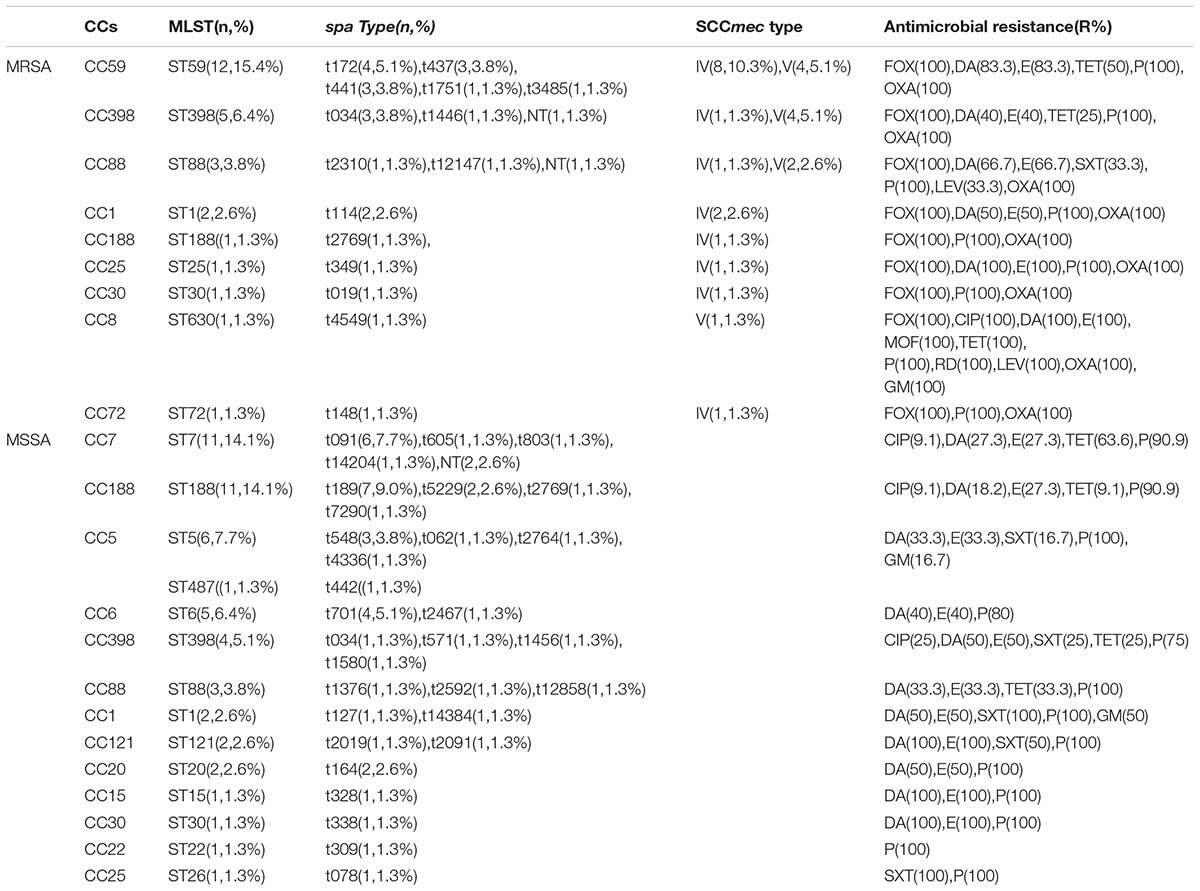
TABLE 1. Molecular characteristics and antibiotic resistance profiles of 78 BSI isolates from pediatric patients.
The eBURST analysis of S. aureus using all STs available in the MLST database is shown. These strains were clustered by eBURST into 16 CCs (CC59, CC188, CC7, CC398, CC88, CC5, CC6, CC1, CC20, CC30, CC121, CC25, CC72, CC8, CC15, and CC22). The largest clusters were CC59 and CC188 each with 12 isolates, followed by CC7 with 11 isolates, CC398 with nine isolates, CC5 with seven isolates, CC88 with six isolates, CC6 with five isolates and CC1 with four isolates. The remaining CCs harbored one or two strains (Supplementary Table S1).
The genetic diversity of these isolates was confirmed by spa typing. Forty-four spa types were observed, spa t189 (9.0%, 7/78) was the most predominant type, followed by t091(7.7%, 6/78), t172(5.1%, 4/78), t034(5.1%, 4/78), t701(5.1%, 4/78), t437(3.8%, 3/78), and t441(3.8%, 3/78). Each of the remaining spa types was represented by fewer than three isolates (Table 1).
By SCCmec typing, only two types (types IV and V) were found among 27 MRSA isolates. The most common was type IV, which comprised 16 isolates (59.3%, 16/27), whereas type V comprised 11 isolates (40.7%, 11/27; Table 1).
Antimicrobial Susceptibility Testing
The antimicrobial resistance profiles of 78 S. aureus BSI isolates according to MLST are listed in Table 1. All the strains were susceptible to linezolid, vancomycin, quinupristin/dalfopristin, and tigecycline. The majority were resistant to penicillin (93.6%), erythromycin (46.2%), and clindamycin (44.9%); however, they were susceptible to most of the antibiotics tested. The resistance rates to other antibiotics tested were 34.6% to cefoxitin or oxacillin, 23.1% to tetracycline, 9% to trimethoprim-sulfamethoxazole, 5.1% to fluoroquinolones (5.1% to ciprofloxacin, 2.6% to levofloxacin, 1.3% to moxifloxacin), 3.8% to gentamicin, and 1.3% to rifampicin (Supplementary Table S2).
Among the 78 S. aureus isolates, 40 (51.3%) strains were resistant to ≥ 3 distinct classes of antibiotics, including 23 (29.5%) MRSA and 17 (21.8%) MSSA strains. Twenty (25.6%) isolates were resistant to five or more antibiotics, six (7.7%) were resistant to four antibiotics, and eighteen (23.1%) were resistant to three antibiotics. Among MSSA strains, 17 (21.8%) were resistant to ≥ 3 antibiotics, eight (10.3%) showed resistance to ≥ 4 antibiotics, and four (5.1%) were resistant to ≥ 5 antibiotics, However, all MRSA strains were found to be resistant to at least three tested antibiotics.
Virulence Gene Profiles
The distribution of 33 putative virulence genes varied among the 78 BSI strains according to ST (Table 2). All of these virulence genes except lukM, etb, and arcA genes were identified within multiple isolates, and all isolates simultaneously harbored at least six virulence genes. Fifty (64.1%, 50/78) isolates harbored ≥ 10 tested virulence genes, among which one isolate harbored 17 genes, five isolates had 15 genes, seven isolates contained 14 genes, 14 isolates harbored 13 genes, eight isolates had 12 genes, 10 isolates contained 11 genes, and five isolates harbored 10 genes.

TABLE 2. Frequencies of virulence and enterotoxin genes among the molecular types of 78 bloodstream infection Staphylococcus aureus isolates from pediatric patients.
Adhesion genes were present in most S. aureus isolates; 100% carried the icaA and clfA genes, 85.9% harbored sdrC, and 69.2% carried sdrE.
The most prevalent toxin-encoding genes detected were hla (100%), lukE (89.8%), hlg2 (84.6%), and hlb (57.7%). The positivity rates for tsst1 (5.1%) and eta (3.8%) among all BSI isolates were low. The pvl gene was detected in 14 strains, which represented six different STs, with ST59 being the most common. The presence of staphylococcal enterotoxin genes was strongly associated with the MLST profile. Thirteen classical enterotoxin genes (sea, seb, sec, sed, see, seg, seh, sei, sem, sen, seo, seq, and sek) were detected within the 78 BSI strains (Table 2). Overall, each enterotoxin gene was found in multiple S. aureus isolates, ranging from 5.1% to 23.1%. No enterotoxin gene was found in ST398 and ST630 isolates. see-sep genes were present in ST7 and ST88 strains, whereas sed-sej genes were present in ST5 and ST487 strains. All ST5, ST20, ST22, ST25, ST26, ST30, ST121, and ST487 strains harbored seg-sei-sem-sen-seo genes, but all ST59 isolates carried seb-sek-seq genes.
Molecular Characteristics of MSSA and MRSA
Both MSSA and MRSA showed considerable genetic diversity. Fourteen distinct STs and 31 spa types were identified among MSSA isolates, whereas nine STs and 15 spa types were found with MRSA strains. Among the MSSA isolates, ST7-MSSA and ST188-MSSA (14.1% each) represented the most predominant clone, followed by ST5-MSSA (7.7%) and ST6-MSSA (6.4%). Among MRSA isolates, ST59-MRSA- SCCmecIV/V (15.4%), the predominant CA-MRSA clone in China, still represented the most common clone. ST398, ST88, ST188, ST1, and ST30 were identified among both MSSA and MRSA isolates.
All MRSA isolates were resistant to oxacillin, penicillin, and cefoxitin. All MSSA isolates were susceptible to cefoxitin, oxacillin, moxifloxacin, levofloxacin, and rifampicin. In general, MRSA strains showed much higher resistance rates to the tested antibiotics than MSSA strains except in the cases of ciprofloxacin, trimethoprim-sulfamethoxazole, and gentamicin (Table 3).
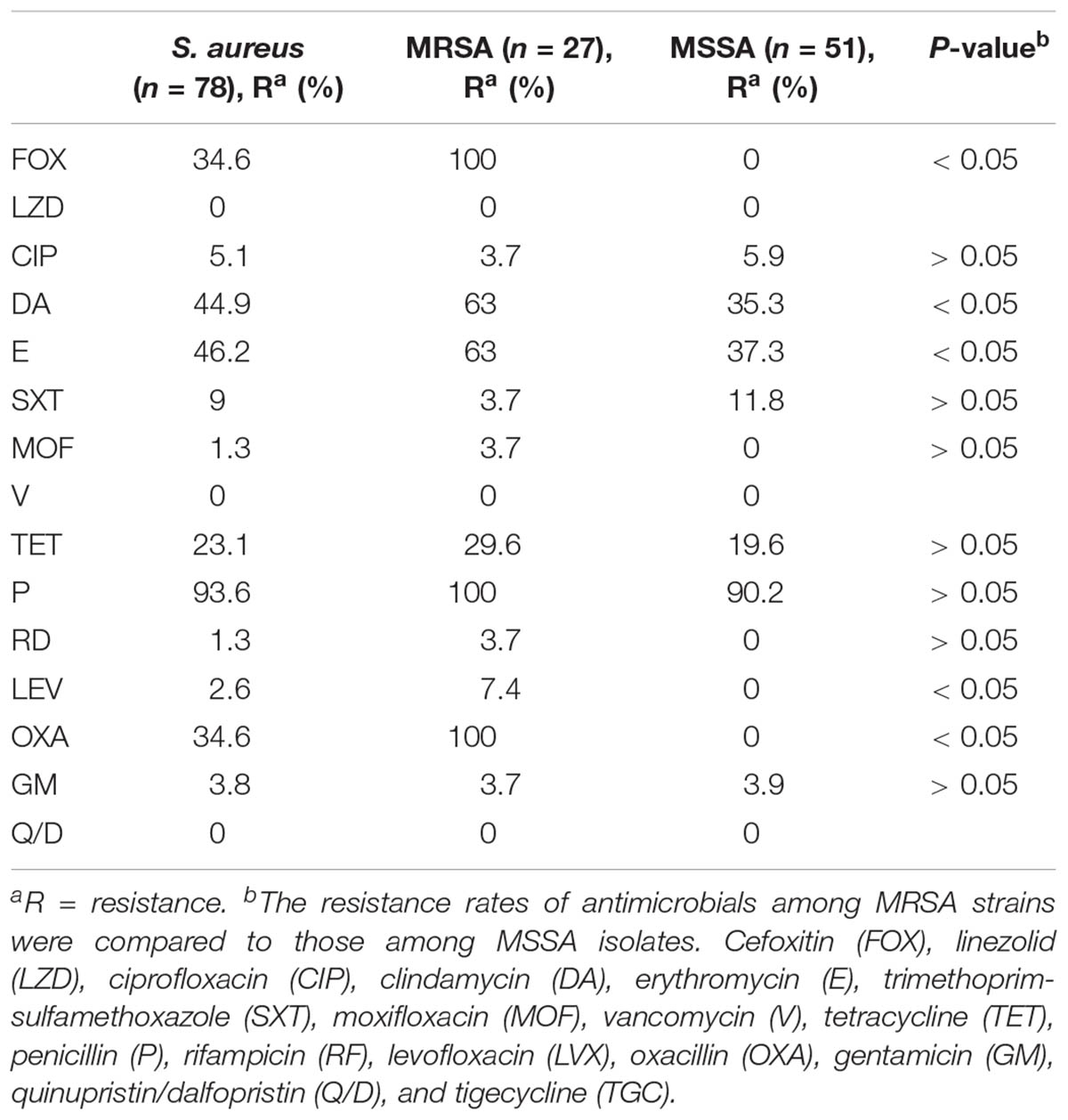
TABLE 3. Antimicrobial susceptibility profiles of methicillin-susceptible Staphylococcus aureus (MSSA) and methicillin-resistant S. aureus (MRSA) isolates.
All isolates harbored icaA, clfA, and hla genes. Fifty (64.1%, 50/78) isolates, including 31 (60.8%) MSSA and 19 (70.4%) MRSA strains, harbored ≥ 10 of the tested virulence genes evaluated in this study. The positivity rates for pvl among MRSA and MSSA isolates were 37.0% (10/27) and 7.8% (4/51), respectively. The virulence genes pvl, hlb, sdrC, lukE, tsst, seb, sek, and seq were significantly more common in MRSA strains, whereas only sdrE was more prevalent in MSSA strains (Table 4). MSSA isolates consistently lacked sek and seq, and these genes were only present in MRSA isolates.
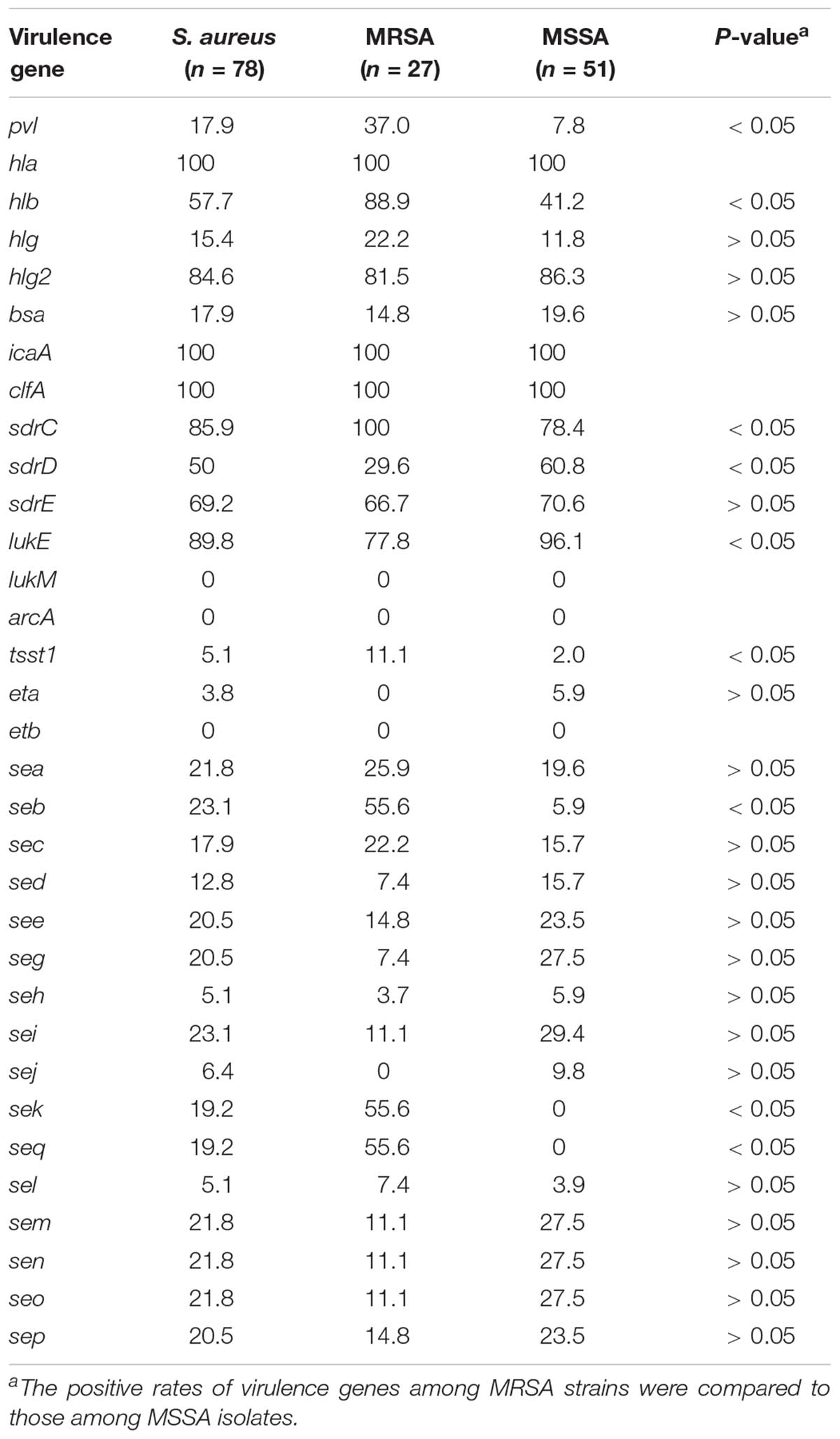
TABLE 4. Frequencies of virulence genes among methicillin-susceptible Staphylococcus aureus (MSSA) and methicillin-resistant S. aureus (MRSA) isolates.
Discussion
Recently, the incidence of community-associated MRSA infection has been increasing. CA-MRSA typically presents as skin and soft tissue infection (SSTI), but invasive infection such as bacteremia can occur, which can lead to serious or even fatal consequences, especially in children and immunocompromised patients. Given this dangerous consequence of CA-MRSA infection in pediatric patients, there is an urgent need to understand the prevalence, molecular characteristics, and virulence profiles of CA-MRSA strains isolated from BSIs to initiate measure to control infection and transmission in local communities.
The occurrence rate of CA-MRSA varies substantially worldwide, ranging from < 1% to > 50% in different countries (Deurenberg and Stobberingh, 2008; Chen and Huang, 2014). In an ANSORP study conducted in 17 hospitals in eight Asian countries, namely Korea, Taiwan, Hong Kong, Thailand, the Philippines, Vietnam, India, and Sri Lanka, the rate of MRSA among CA-SA infections ranged from 2.5 to 39% (Chen and Huang, 2014). In addition, the rate of CA-MRSA among childhood infections in Taiwan increased significantly from 9.8% in 1999–2000 to 56% in 2004–2005. In China, based on a study performed in Beijing Children’s Hospital, 14(4%) of 351 CA-SA infections were caused by MRSA (Wu et al., 2010). These studies focused on CA-MRSA infection, mainly from SSTIs. Few data describing the rate of CA-MRSA isolated from BSI are available. As a result, we characterized 78 CA-SA isolates recovered from pediatric patients with BSIs between 2012 and 2017 in Shanghai and identified 34.6% as MRSA strains. This indicates that MRSA is an important pathogenic bacterium associated with severe, life-threatening infections in children, and suggests the urgent need for active surveillance of such infections and transmission in children.
Community-associated-methicillin-resistant S. aureus infections have been reported globally and the major pandemic clones are frequently related to specific geographic areas (Mediavilla et al., 2012; Li et al., 2016). clones with ST59 are mostly found in the Asia-Pacific region including Taiwan and Australia (Chen and Huang, 2014). In China, ST59-SCCmecIV/V-t437 is always identified as the major epidemic CA-MRSA clone (Geng et al., 2010). Our current data confirms previous findings with respect to the prevalence of ST59, but minor differences were noted. ST59-SCCmecIV-t172 was present with increasing prevalence and became the most common CA-MRSA clone in Shanghai. Although there is considerable genetic diversity among MSSA clones observed in China, ST188 and ST7 have been reported to be the most prevalent clones from adult BSI cases. Our result was consistent with these data, with these strains accounting for 29.5% of all isolates. Recently, ST188 and ST7 were identified as the predominant types among MSSA strains from childhood pneumonia between January 2014 and June 2015 (Song et al., 2017). Furthermore, Our previous study also found these were the common types that caused bovine mastitis from 2014 to 2015 in Shanghai and Zhejiang areas (Li et al., 2017), implying that among the species, some prevailing clones have arisen and spread throughout China.
In the current study, ST398, ST188, ST88, and ST1 were similarly identified among both MSSA and MRSA strains, which suggested that these MSSA lineages probably provide a stable genetic environment for the integration of SCCmec to facilitate MRSA infection and transmission in both healthcare facilities and communities. Of note, ST398 was originally found to be a MSSA strain in China, and increasing numbers of ST398-MRSA isolates have emerged in many regions. Historically, ST398-MRSA, first referred to as livestock-associated MRSA (LA-MRSA), has been reported among pigs and pig farmers in France and the Netherlands (Graveland et al., 2011; Fluit, 2012). After, it has become the overwhelmingly dominant lineage in Europe and North America. In the current study, we identified nine ST398 isolates (Tables 3, 4) including five MRSA and four MSSA strains. It is very difficult to speculate on the origins of these isolates because of the absence of epidemiological data linking these to animals. However, LA-SA usually harbored an intact beta-toxin gene (hlb) and no lysogenic prophages encoding the immune evasion complex genes (sea, sep, sak, scn, and chp genes) (van Wamel et al., 2006). Among ST398 isolates in our study, they all had an intact hlb gene and didn’t contain sea and sep genes. Three strains lacked all the immune evasion complex genes, and others harbored one, two or three of sak, scn and chp genes. This was important evidence that these strains were of animal origin. Therefore, it is very important to carefully monitor the animal-to-human transmissibility of LA-SA.
The pathogenicity for S. aureus mainly relies on the presence of a variety of virulence factors that mediate adhesion, invasion, persistence, tissue invasion, the evasion/destruction of host defenses, and toxin-related disease (Dinges et al., 2000; Wardenburg et al., 2007; Diep and Otto, 2008). These include microbial surface protein (Josefsson et al., 1998), hemolysin, toxic shock syndrome toxin-1, exfoliative toxins, and SEs. In this study, adhesion genes were present in most S. aureus isolates; 100% carried the icaA and clfA genes, 85.9% harbored sdrC, and 69.2% carried sdrE. The high prevalence of adhesion genes in clinical strains is consistent with the notion that the adherence of S. aureus to host cells is the first vital step for bacterial pathogenicity. The presence of pvl has previously been strongly associated with CA-MRSA infections in many studies (Lina et al., 1999). We found ten MRSA isolates and four MSSA isolates that harbored pvl. The prevalence of pvl in MRSA isolates (37.0%) was significantly higher than that in MSSA isolates (7.8%), supporting the hypothesis that pvl is a possible marker for CA-MRSA. In addition, the distribution of some virulence genes, especially SEs, was reported to be closely associated with specific molecular types (He et al., 2013; Wang et al., 2016). A similar result was observed in the current study. The seb-sek-seq genes were present in all ST59 strains, whereas the seg-sei-sem-sen-seo genes were found to be present in different clones, including ST5, ST20, ST22, ST25, ST26, ST30, ST121, and ST487 strains. In addition, ST5 and ST59 isolates harbored more enterotoxin-encoding genes than ST188 and ST398 strains, implying that different molecular types are associated with different virulence profiles.
The current work has several limitations. Most importantly, the relatively small sample size from a single center limited the representative significance of the research. Second, it is not known whether different genotypes are associated with clinical outcomes because of the absence of clinical information. In addition, as retrospective research, it is possible that patients were misclassified as CA due to incomplete data. However, this study revealed that community-associated S. aureus strains from BSIs in children demonstrated considerable genetic diversity and identified major genotypes of CA-MRSA and CA-MSSA, with a high prevalence of MRSA.
Author Contributions
ML and XW designed the studies and obtained funding. XW, QL, HZ, WH, and QF performed the experiments and/or analyzed the data. ML, XW, and XL wrote the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (grant 81301392) and the Training Program for Outstanding Young Teachers in Higher Education Institutions (ZZjdyx13132), the Training Program for Clinical Medical Young Talents in Shanghai (HYWJ201605), and the Cultivated Fund of the National Natural Science Foundation of China (YJG-SCMC2017-5) to XW.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank all the patients who contributed their specimens and clinical data for this study. They also thank the microbiologists and technical staff of Shanghai Children’s Medical Center for collecting the bacterial isolates and laboratory testing.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01211/full#supplementary-material
Footnotes
References
Aanensen, D. M., and Spratt, B. G. (2005). The multilocus sequence typing network: mlst.net. Nucleic Acids Res. 33, W728–W733. doi: 10.1093/nar/gki415
Arvidson, S., and Tegmark, K. (2001). Regulation of virulence determinants in Staphylococcus aureus. Int. J. Med. Microbiol. 291, 159–170. doi: 10.1078/1438-4221-00112
Bassetti, M., Trecarichi, E. M., Mesini, A., Spanu, T., Giacobbe, D. R., and Tumbarello, M. (2012). Risk factors and mortality of healthcare-associated and community-acquired Staphylococcus aureus bacteraemia. Clin. Microbiol. Infect. 18, 862–869. doi: 10.1111/j.1469-0691.2011.03679.x
Chen, C. J., and Huang, Y. C. (2014). New epidemiology of Staphylococcus aureus infection in Asia. Clin. Microbiol. Infect. 20, 605–623. doi: 10.1111/1469-0691.12705
Clinical and Laboratory Standards Institute [CLSI] (2015). Performance Standards. Antimicrobial Susceptibility Testing[S]: Twenty-fourth Informational Supplement. Wayne, PA: Clinical and Laboratory Standards Institute.
Cosgrove, S. E., Sakoulas, G., Perencevich, E. N., Schwaber, M. J., Karchmer, A. W., and Carmeli, Y. (2003). Comparison of mortality associated with methicillin-resistant and methicillin-susceptible Staphylococcus aureus bacteremia: a meta-analysis. Clin. Infect. Dis. 36, 53–59. doi: 10.1086/345476
David, M. Z., Acree, M. E., Sieth, J. J., Boxrud, D. J., Dobbins, G., Lynfield, R., et al. (2015). Pediatric Staphylococcus aureus Isolate genotypes and infections from the dawn of the community-associated methicillin-resistant S. aureus Epidemic Era in Chicago, 1994 to 1997. J. Clin. Microbiol. 53, 2486–2491. doi: 10.1128/JCM.00096-15
David, M. Z., Daum, R. S., Bayer, A. S., Chambers, H. F., Fowler, V. G. Jr., and Spellberg, B. (2014). Staphylococcus aureus bacteremia at 5 US academic medical centers, 2008-2011: significant geographic variation in community-onset infections. Clin. Infect. Dis. 59, 798–807. doi: 10.1093/cid/ciu410
Deurenberg, R. H., and Stobberingh, E. E. (2008). The evolution of Staphylococcus aureus. Infect. Genet. Evol. 8, 747–763. doi: 10.1016/j.meegid.2008.07.007
Diep, B. A., and Otto, M. (2008). The role of virulence determinants in community-associated MRSA pathogenesis. Trends Microbiol. 16, 361–369. doi: 10.1016/j.tim.2008.05.002
Dinges, M. M., Orwin, P. M., and Schlievert, P. M. (2000). Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 13, 16–34. doi: 10.1128/CMR.13.1.16-34.2000
Enright, M. C., and Spratt, B. G. (1999). Multilocus sequence typing. Trends Microbiol. 7, 482–487. doi: 10.1016/S0966-842X(99)01609-1
Fluit, A. C. (2012). Livestock-associated Staphylococcus aureus. Clin. Microbiol. Infect. 18, 735–744. doi: 10.1111/j.1469-0691.2012.03846.x
Geng, W., Yang, Y., Wu, D., Huang, G., Wang, C., and Shen, X. (2010). Molecular characteristics of community-acquired, methicillin-resistant Staphylococcus aureus isolated from Chinese children. FEMS Immunol. Med. Microbiol. 58, 356–362. doi: 10.1111/j.1574-695X.2010.00648.x
Graveland, H., Duim, B., van Duijkeren, E., Heederik, D., and Wagenaar, J. A. (2011). Livestock-associated methicillin-resistant Staphylococcus aureus in animals and humans. Int. J. Med. Microbiol. 301, 630–634. doi: 10.1016/j.ijmm.2011.09.004
Hanakawa, Y., Schechter, N. M., Lin, C., Garza, L., Li, H., and Yamaguchi, T. (2002). Molecular mechanisms of blister formation in bullous impetigo and staphylococcal scalded skin syndrome. J. Clin. Invest. 110, 53–60. doi: 10.1172/JCI0215766
Hartmann, F. A., Trostle, S. S., and Klohnen, A. A. (1997). Isolation of methicillin resistant Staphylococcus aureus from a postoperative wound infection in a horse. J. Am. Vet. Med. Assoc. 211, 590–592.
He, W., Chen, H., Zhao, C., Zhang, F., Li, H., Wang, Q., et al. (2013). Population structure and characterisation of Staphylococcus aureus from bacteraemia at multiple hospitals in China: association between antimicrobial resistance, toxin genes and genotypes. Int. J. Antimicrob. Agents 42, 211–219. doi: 10.1016/j.ijantimicag.2013.04.031
Hiramatsu, K., Ito, T., Tsubakishita, S., Sasaki, T., Takeuchi, F., Morimoto, Y., et al. (2013). Genomic basis for methicillin resistance in Staphylococcus aureus. Infect. Chemother. 45, 117–136. doi: 10.3947/ic.2013.45.2.117
Huang, Y. C., and Chen, C. J. (2011). Community-associated meticillin-resistant Staphylococcus aureus in children in Taiwan, 2000s. Int. J. Antimicrob. Agents 38, 2–8. doi: 10.1016/j.ijantimicag.2011.01.011
Jarraud, S., Mougel, C., Thioulouse, J., Lina, G., Meugnier, H., and Forey, F. (2002). Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect. Immun. 70, 631–641. doi: 10.1128/IAI.70.2.631-641.2002
Josefsson, E., McCrea, K. W., Ni Eidhin, D., O’Connell, D., Cox, J., Hook, M., et al. (1998). Three new members of the serine-aspartate repeat protein multigene family of Staphylococcus aureus. Microbiology 144, 3387–3395. doi: 10.1099/00221287-144-12-3387
Kang, S., Lee, J., and Kim, M. (2017). The association between Staphylococcus aureus nasal colonization and symptomatic infection in children in Korea where ST72 is the major genotype: a prospective observational study. Medicine 96:e7838. doi: 10.1097/MD.0000000000007838
Kaya, H., Hasman, H., Larsen, J., Stegger, M., Johannesen, T. B., Allesoe, R. L., et al. (2018). SCCmecFinder, a web-based tool for typing of staphylococcal cassette chromosome mec in Staphylococcus aureus using whole-genome sequence data. mSphere 3:e006612-17. doi: 10.1128/mSphere.00612-17
Kondo, Y., Ito, T., Ma, X. X., Watanabe, S., Kreiswirth, B. N., Etienne, J., et al. (2007). Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob. Agents Chemother. 51, 264–274. doi: 10.1128/AAC.00165-06
Koreen, L., Ramaswamy, S. V., Graviss, E. A., Naidich, S., Musser, J. M., and Kreiswirth, B. N. (2004). spa typing method for discriminating among Staphylococcus aureus isolates: implications for use of a single marker to detect genetic micro- and macrovariation. J. Clin. Microbiol. 42, 792–799. doi: 10.1128/JCM.42.2.792-799.2004
Li, M., Dai, Y., Zhu, Y., Fu, C. L., Tan, V. Y., and Otto, M. (2016). Virulence determinants associated with the Asian community-associated methicillin-resistant Staphylococcus aureus lineage ST59. Sci. Rep. 6:27899. doi: 10.1038/srep27899
Li, T., Lu, H., Wang, X., Gao, Q., Dai, Y., Shang, J., et al. (2017). Molecular characteristics of Staphylococcus aureus causing bovine mastitis between 2014 and 2015. Front. Cell Infect. Microbiol. 7:127. doi: 10.3389/fcimb.2017.00127
Lina, G., Piemont, Y., Godail-Gamot, F., Bes, M., Peter, M. O., and Etienne, J. (1999). Involvement of panton-valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin. Infect. Dis. 29, 1128–1132. doi: 10.1086/313461
Lowy, F. D. (1998). Staphylococcus aureus infections. N. Engl. J. Med. 339, 520–532. doi: 10.1056/NEJM199808203390806
McMullan, B. J., Bowen, A., Blyth, C. C., Van Hal, S., Korman, T. M., Buttery, J., et al. (2016). Epidemiology and mortality of Staphylococcus aureus bacteremia in Australian and new Zealand children. JAMA Pediatr. 170, 979–986. doi: 10.1001/jamapediatrics.2016.1477
Mediavilla, J. R., Chen, L., Mathema, B., and Kreiswirth, B. N. (2012). Global epidemiology of community-associated methicillin resistant Staphylococcus aureus (CA-MRSA). Curr. Opin. Microbiol. 15, 588–595. doi: 10.1016/j.mib.2012.08.003
Otto, M. (2013). Community-associated MRSA: what makes them special? Int. J. Med. Microbiol. 303, 324–330. doi: 10.1016/j.ijmm.2013.02.007
Peacock, S. J., Moore, C. E., Justice, A., Kantzanou, M., Story, L., and Day, N. P. (2002). Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect. Immun. 70, 4987–4996. doi: 10.1128/IAI.70.9.4987-4996.2002
Pena Amaya, P., Haim, M. S., Fernandez, S., Di Gregorio, S., Teper, A., Vazquez, M., et al. (2017). Molecular epidemiology of methicillin-resistant Staphylococcus aureus in cystic fibrosis patients from Argentina. Microb. Drug Resist. doi: 10.1089/mdr.2017.0162 [Epub ahead of print].
Roediger, J. C., Outhred, A. C., Shadbolt, B., and Britton, P. N. (2017). Paediatric Staphylococcus aureus bacteraemia: a single-centre retrospective cohort. J. Paediatr. Child Health 53, 180–186. doi: 10.1111/jpc.13329
Shopsin, B., Gomez, M., Montgomery, S. O., Smith, D. H., Waddington, M., and Kreiswirth, B. N. (1999). Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J. Clin. Microbiol. 37, 3556–3563.
Skov, R., Christiansen, K., Dancer, S. J., Daum, R. S., Dryden, M., Huang, Y. C., et al. (2012). Update on the prevention and control of community-acquired meticillin-resistant Staphylococcus aureus (CA-MRSA). Int. J. Antimicrob. Agents 39, 193–200. doi: 10.1016/j.ijantimicag.2011.09.029
Song, Z., Gu, F. F., Guo, X. K., Ni, Y. X., He, P., and Han, L. Z. (2017). Antimicrobial resistance and molecular characterization of Staphylococcus aureus causing childhood pneumonia in Shanghai. Front. Microbiol. 8:455. doi: 10.3389/fmicb.2017.00455
van Wamel, W. J., Rooijakkers, S. H., Ruyken, M., van Kessel, K. P., and van Strijp, J. A. (2006). The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J. Bacteriol. 188, 1310–1315. doi: 10.1128/JB.188.4.1310-1315.2006
Wang, J. L., Chen, S. Y., Wang, J. T., Wu, G. H., Chiang, W. C., and Chang, S. C. (2008). Comparison of both clinical features and mortality risk associated with bacteremia due to community-acquired methicillin-resistant Staphylococcus aureus and methicillin-susceptible S. aureus. Clin. Infect. Dis. 46, 799–806. doi: 10.1086/527389
Wang, X., Li, X., Liu, W., Huang, W., Fu, Q., and Li, M. (2016). Molecular characteristic and virulence gene profiles of community-associated methicillin-resistant Staphylococcus aureus isolates from pediatric patients in Shanghai, China. Front. Microbiol. 7:1818. doi: 10.3389/fmicb.2016.01818
Wardenburg, J. B., Patel, R. J., and Schneewind, O. (2007). Surface Proteins and exotoxins are required for the pathogenesis of Staphylococcus aureus Pneumonia. Infect. Immun. 75, 1040–1044. doi: 10.1128/IAI.01313-06
Warner, J. E., and Onderdonk, A. B. (2004). Diversity of toxic shock syndrome toxin 1-positive Staphylococcus aureus isolates. Appl. Environ. Microbiol. 70, 6931–6935. doi: 10.1128/AEM.70.11.6931-6935.2004
Wu, D., Wang, Q., Yang, Y., Geng, W., Wang, Q., and Shen, X. (2010). Epidemiology and molecular characteristics of community-associated methicillin-resistant and methicillin-susceptible Staphylococcus aureus from skin/soft tissue infections in a children’s hospital in Beijing, China. Diagn. Microbiol. Infect. Dis. 67, 1–8. doi: 10.1016/j.diagmicrobio.2009.12.006
Keywords: bloodstream infections, methicillin-resistance S. aureus, methicillin-susceptible S. aureus, multidrug resistance, sequence typing, virulence genes
Citation: Wang X, Liu Q, Zhang H, Li X, Huang W, Fu Q and Li M (2018) Molecular Characteristics of Community-Associated Staphylococcus aureus Isolates From Pediatric Patients With Bloodstream Infections Between 2012 and 2017 in Shanghai, China. Front. Microbiol. 9:1211. doi: 10.3389/fmicb.2018.01211
Received: 14 October 2017; Accepted: 17 May 2018;
Published: 06 June 2018.
Edited by:
Leonard Peruski, Centers for Disease Control and Prevention (CDC), United StatesCopyright © 2018 Wang, Liu, Zhang, Li, Huang, Fu and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xing Wang, wx_5166@163.com Min Li, ruth_limin@126.com
†These authors have contributed equally to this work.