Cdk5 Is Essential for Amphetamine to Increase Dendritic Spine Density in Hippocampal Pyramidal Neurons
 Soledad Ferreras1,2†
Soledad Ferreras1,2†  Guillermo Fernández1† Víctor Danelon3
Guillermo Fernández1† Víctor Danelon3  María V. Pisano1 Luján Masseroni4 Christopher A. Chapleau2 Favio A. Krapacher1 Estela C. Mlewski1 Daniel H. Mascó3 Carlos Arias1
María V. Pisano1 Luján Masseroni4 Christopher A. Chapleau2 Favio A. Krapacher1 Estela C. Mlewski1 Daniel H. Mascó3 Carlos Arias1  Lucas Pozzo-Miller2
Lucas Pozzo-Miller2  María G. Paglini1,5*
María G. Paglini1,5*- 1Laboratory of Neurophysiology, Instituto de Investigación Médica Mercedes y Martín Ferreyra, INIMEC-CONICET, Universidad Nacional de Córdoba, Córdoba, Argentina
- 2Department of Neurobiology, Civitan International Research Center, University of Alabama at Birmingham, Birmingham, AL, United States
- 3Centro de Biología Celular y Molecular, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, IIBYT-CONICET, Córdoba, Argentina
- 4Laboratory of Neurobiology, Instituto de Investigación Médica Mercedes y Martín Ferreyra, INIMEC-CONICET, Universidad Nacional de Córdoba, Córdoba, Argentina
- 5Virology Institute “Dr. J. M. Vanella”, Facultad de Ciencias Médicas, Universidad Nacional de Córdoba, Córdoba, Argentina
Psychostimulant drugs of abuse increase dendritic spine density in reward centers of the brain. However, little is known about their effects in the hippocampus, where activity-dependent changes in the density of dendritic spine are associated with learning and memory. Recent reports suggest that Cdk5 plays an important role in drug addiction, but its role in psychostimulant’s effects on dendritic spines in hippocampus remain unknown. We used in vivo and in vitro approaches to demonstrate that amphetamine increases dendritic spine density in pyramidal neurons of the hippocampus. Primary cultures and organotypic slice cultures were used for cellular, molecular, pharmacological and biochemical analyses of the role of Cdk5/p25 in amphetamine-induced dendritic spine formation. Amphetamine (two-injection protocol) increased dendritic spine density in hippocampal neurons of thy1-green fluorescent protein (GFP) mice, as well as in hippocampal cultured neurons and organotypic slice cultures. Either genetic or pharmacological inhibition of Cdk5 activity prevented the amphetamine–induced increase in dendritic spine density. Amphetamine also increased spine density in neurons overexpressing the strong Cdk5 activator p25. Finally, inhibition of calpain, the protease necessary for the conversion of p35 to p25, prevented amphetamine’s effect on dendritic spine density. We demonstrate, for the first time, that amphetamine increases the density of dendritic spine in hippocampal pyramidal neurons in vivo and in vitro. Moreover, we show that the Cdk5/p25 signaling and calpain activity are both necessary for the effect of amphetamine on dendritic spine density. The identification of molecular mechanisms underlying psychostimulant effects provides novel and promising therapeutic approaches for the treatment of drug addiction.
Introduction
Changes in behavior that occur as a function of experience, like environmental enrichment (Engmann et al., 2015), drug addition (Robinson and Kolb, 1997; Rasakham et al., 2014), injuries (Tseng and Hu, 1996; Chen et al., 2003) and throughout aging (Scheibel et al., 1975; Wang et al., 2009), have their basis in the reorganization of excitatory synaptic connections, also known as structural plasticity of dendritic spines. Structural plasticity involves a variety of proteins controlling the organization of the actin cytoskeleton (Schubert and Dotti, 2007; Cingolani and Goda, 2008) and changes in dendritic spine shape, size and number are determined by local actin dynamics (Fischer et al., 2000; Tada and Sheng, 2006; Lai and Ip, 2013). Activity-dependent dynamic changes in spines of hippocampal pyramidal neurons are essential for learning and memory (Yuste and Bonhoeffer, 2001; Kasai et al., 2003).
The Cdk5/p35 signaling complex has been involved in multiple processes during typical brain development, including neuronal migration and the formation of axons and dendrites (Nikolic et al., 1996; Chae et al., 1997; Paglini et al., 1998, 2001; Kwon et al., 1999; Smith and Tsai, 2002), as well as in associative learning and memory in adult rodents (Fischer et al., 2002, 2005; Angelo et al., 2006; Bignante et al., 2008). On the other hand, Cdk5 plays a critical role in the pathophysiology of several neurodegenerative disorders. Deregulation of Cdk5 results in neuronal loss in Parkinson’s, Alzheimer’s and Huntington’s diseases (Cheung and Ip, 2012; Shah and Lahiri, 2014), which underscores the importance of the complex regulation of Cdk5 activity necessary to maintain its activity within physiological levels. Cdk5 kinase activity is regulated in many different ways. The main regulators of its kinase activity are the activators p35 and p39, which are highly expressed in adult brain (Tsai et al., 1994; Tang et al., 1995). Other Cdk5 regulators in postmitotic neurons are the Cyclin family proteins, such as Cyclin D1, Cyclin E and Cyclin I (De Falco et al., 2004; Brinkkoetter et al., 2010; Odajima et al., 2011). In addition, glutathione-S-transferase P (GSTP1) binds to Cdk5 and modulates its activity in neurons (Sun et al., 2011). Furthermore, Cdk5 can be regulated by posttraslational modifications, such as phosphorylation and S-nitrosylation (reviewed in Shah and Lahiri, 2014). The N-terminal domain of the Cdk5 activator p35 is cleaved by the Ca+-dependent cysteine protease calpain, which generates p25, a C-terminal truncation peptide that contains all necessary elements for Cdk5 activation (Kusakawa et al., 2000; Lee et al., 2000; Nath et al., 2000; Dhavan and Tsai, 2001; Patzke et al., 2003). Since p25 is resistant to ubiquitin-mediated proteolysis, it substantially extends the activation period of Cdk5 (Patrick et al., 1999). Relevant to our studies, transient expression of p25 increased the number of synapses and dendritic spine density, enhancing hippocampal LTP and facilitating learning and memory (Fischer et al., 2005).
Notably, exposure to either amphetamine or cocaine increases the dendritic spine density in reward centers of the brain, suggesting that the molecular mechanisms involved in other forms of synaptic plasticity are hijacked by drugs of abuse (Berke and Hyman, 2000; Hyman and Malenka, 2001; Kolb et al., 2003; Robinson and Kolb, 2004). Although the dopaminergic system is the focus of intense research of motivational and reward-related learning (Li et al., 2003; Chen et al., 2010), the neurobiological basis of natural reward learning and memory cannot be fully explained without the contribution of the hippocampus, a brain area that sends a main output to the dopaminergic reward system. In addition, the hippocampus is a principal site of activity-dependent process during learning and memory (Grace et al., 2007). Little is known, however, about the effects of drugs of abuse on hippocampus.
Interestingly, recent reports have demonstrated that Cdk5 participates in addiction to drugs of abuse (Benavides and Bibb, 2004; Mlewski et al., 2008, 2016). The inhibition of Cdk5 activity with roscovitine decreases cocaine-induced dendritic spine formation in the nucleus accumbens, suggesting that Cdk5 is involved in cocaine-induced dendritic spine morphogenesis (Norrholm et al., 2003). Nevertheless, the detailed mechanism/s by which this or other signaling modules participate in dendritic spine formation induced by psychostimulants in the hippocampus or other brain areas remains to be determined.
In the present study, we demonstrate that amphetamine increases dendritic spine density in pyramidal neurons of the hippocampus. Using a two-amphetamine-injections protocol in a novel context, we observe an increase in dendritic spines density in hippocampal CA1 pyramidal neurons of young mice. In addition, amphetamine increases spine density in primary hippocampal cultures and organotypic slice cultures. Lastly, we provide evidence of the necessity of the Cdk5/p25 signaling complex and of calpain activity for amphetamine to increase spine density.
Materials and Methods
Animals
Mice harboring the thy1-green fluorescent protein (GFP) transgene (Feng et al., 2000) were generated by breeding male heterozygous B6.Cg-Tg(Thy1-EGFP)MJrs/J mice (Jackson Laboratory, Bar Harbor, ME, USA) with female C57BL/6J wild type mice. Animals were weaned at postnatal day (P) 21 and housed 4–5 per cage under a 12 h light/12 h dark cycle at constant temperature (22°C) with free access to food and water. All experiments were performed with P33–40 mice in an isolated behavioral room during the light cycle, between 10:00 AM and 3:00 PM. Embryonic Wistar rats were used for in vitro primary cultures. All animals used in the present study were born and reared at the vivarium of the INIMEC-CONICET-UNC (Cordoba, Argentina). All procedures and care performed in the animals were approved by National Department of Animal Care and Health (SENASA, Argentina) and were in accordance with National Institute of Health general guidelines for the Care and Use of Laboratory Animals. Best efforts were made to reduce the number of animals used and to minimize their suffering. The experimental protocol was reviewed and approved by Institutional Animal Care and Use Committee (CICUAL protocol No. 2016-0001) and complied with the regulations of the Guide for Care and Use of Laboratory Animals (National-Research-Council, 1996).
Drugs
D-Amphetamine (Sigma-Aldrich, St. Louis, MO, USA and Parafarm, Buenos Aires, Argentina), N-Acetyl-L-leucyl-L-norleucinal (ALLN; Sigma-Aldrich, St. Louis, MO, USA) and roscovitine (Calbiochem, San Diego, CA, USA) were used in this study.
Drug Administration
We performed the two-injection protocol of sensitization (TIPS), which consist of two phases: in the first day (phase 1) mice were treated with either an i.p. injection of 4 mg/kg amphetamine or an equivalent volume of saline solution (sal, 0.9% NaCl) and immediately placed in a novel context (open field) for 1 h. After 1 day without receiving any treatment, mice received either amphetamine or sal, and were placed for 1 h in the novel context (phase 2). Male mice were randomly divided into four treatment groups: (1) receiving two vehicle saline injections (sal/sal); (2) receiving saline first and then amphetamine injections (sal/amph); (3) receiving amphetamine first and then saline injections (amph/sal); and (4) receiving two amphetamine injections (amph/amph).
Brain Fixation
Four hours after drug treatments, mice were anesthetized with an i.p. injection of chloral hydrate (0.1 mL/100 g) and transcardially perfused as described (Krapacher et al., 2010). The brains were cut coronally at 100 μm with a freezing microtome (Reicher-Jung Hn40, Leica Microsystems, Wetzlar, Germany) and the collected sections were mounted with Vectashield (Vector Laboratories, Burlingame, CA, USA).
Primary Cultures of Dissociated Neurons
Cultures of dissociated hippocampal pyramidal cells from embryonic rats were prepared as described (Kunda et al., 2001). Neurons at 17–19 days in vitro (DIV) with long axons and well-developed dendritic arbors were used for all experiments.
Organotypic Slice Cultures
Hippocampal slice cultures were prepared from P7 to P10 Wistar rats and maintained in vitro as described (Stoppini et al., 1991; Tyler and Pozzo-Miller, 2001). Briefly, rats were quickly decapitated and their brains aseptically dissected and immersed in ice-cold dissecting solution, consisting of Hanks’ Balanced Salt Solution (HBSS; ThermoFisher Scientific, Waltham, MA, USA) supplemented with glucose (41.55 mM) and antibiotics (1:100; penicillin/streptomycin). Hippocampi were then dissected and transversely sectioned into 400~500 μm slices using a custom-made tissue slicer (Katz, 1987) strung with 20 μm-thick tungsten wire (California Fine Wire Company, Grover Beach, CA, USA). Slices were incubated at 4°C for 30 min, and then plated on tissue culture inserts (0.4 μm pore size, Millicell-CM, Millipore Corporation, Billerica, MA, USA). Culture media contained Neurobasal-A (ThermoFisher Scientific, Waltham, MA, USA), heat-inactivated horse serum (20%) and L-glutamine (1 mM; ThermoFisher Scientific, Waltham, MA, USA). Organotypic cultures were maintained in incubators at 36°C, 5% CO2, 98% relative humidity. The concentration of horse serum in the culture medium was reduced from 20% to 10% at 4 DIV and again reduced to 5% 24 h later. After 24 h in medium containing 5% horse serum, slices were placed in serum-free medium (Neurobasal-A plus B-27 supplement, ThermoFisher Scientific, Waltham, MA, USA) until used for experiments.
DNA Constructs
The following constructs were used: expression cDNA plasmid coding for enhanced yellow fluorescent protein (YFP) obtained from Clontech Inc (Mountain View, CA, USA); expression cDNA plasmid pcDNA3-dnCDK5 was generous gift of Dr. Philip W. Hinds; plasmid DNA coding for p35-full-length was generous gift of Dr. Christopher C. J. Miller. To generate the fusion protein of human p25 with GFP (p25-GFP), a DNA fragment encoding the p25 sequence was obtained by PCR using Pfu polymerase (Stratagene Cloning Systems, La Jolla, CA, USA), with a BamH1 restriction site introduced into the 5′-end and a SAL site into the 3′-end, and cloned into the pEGFP-N1 vector (Clontech Inc, Mountain View, CA, USA). The Cdk5 short hairpin RNA (shRNA) plasmid was constructed in pSilencer U6 1.0 vector (Ambion Inc, Austin, TX, USA). 5′-GGG AGATCTGTCTACTCAAAGAA and 5′-GGG ATT CTG TCA CAG CCG TAA CG were used as targeting sequences following the procedures described (Chuang et al., 2005). The DNA fragments containing U6-Cdk5-sh and U6-control-sh were inserted into pCAG-HcRed vector under the control of chick actin-minimal (CAG) promoter; the resulting plasmids were referred to as shCdk5. Western immunoblots of Cdk5-expressing N2a cells (brain neuroblastoma) confirmed that shCdk5 reduced Cdk5 protein levels (data not shown).
Transfection Methods
For primary neuronal cultures, transient transfections were performed at 16 DIV on 30 mm dishes, using Lipofectamine 2000 (Invitrogen, San Diego, CA, USA) according to the manufacture’s recommendations. For experiments with organotypic cultures, slices were biolistically-transfected at 7 DIV, as described (Alonso et al., 2004). Briefly, prior to transfection, a mixture of penicillin/streptomycin/amphotericin B (ThermoFisher Scientific, Waltham, MA, USA) was added to the culture media in order to prevent contamination during biolistic transfection; this mixture was removed after 24 h. The plasmid (cDNAs of interest) were precipitated onto 25 mg of 1.6 μm-diameter colloidal gold at a ratio of 63.5 μg of YFP to 113.5 μg of interest plasmid and then coated onto Tefzel tubing. The success rate of co-transfection of the same neuron with two cDNA plasmids using the gene-gun has been demonstrated to be >90% (Boda et al., 2004; Moore et al., 2007), significantly reducing the likelihood of occurrence of YFP-positive and dnCDK5/p35/p25-negative neurons.
Culture Fixation
Fifteen hours after transfection, hippocampal slice cultures were exposed to amphetamine (50 μM) and/or roscovitine (10 μM) for 48 h and then were fixed by immersion in 4% paraformaldehyde in 100 mM phosphate buffer (overnight at 4°C), and washed in PBS. Filter membranes around each slice were trimmed, and each slice was individually mounted on glass slides and cover slipped using Vectashield (Vector Laboratories, Burlingame, CA, USA). Cultured hippocampal neurons received the same treatment and then coverslips with neurons were fixed for 20 min in 4% paraformaldehyde in 100 mM phosphate buffer at room temperature and mounted on glass slides using FluorSave (Calbiochem® Merck KGaA, Darmstadt, Germany).
Laser-Scanning Confocal Microscopy
High-resolution images of tertiary and quaternary branches of apical dendrites of pyramidal neurons, displaying either GFP or YFP fluorescence throughout the whole dendritic tree, were acquired in Fluoview FV300 and FV1000 laser-scanning confocal microscopes (Olympus, Tokyo, Japan) using oil immersion 40× (NA 1.4) and 63× (NA 1.4) objectives (PlanApo, Olympus) and 3× digital zoom. GFP and YFP were excited using an Ar laser (488 nm), and detected using standard FITC filters. Series of optical sections were acquired in the z-axis at 0.12 μm intervals through individual apical dendritic branches.
Quantitative Analyses of Dendritic Spine Density and Morphology
Small protrusions that extended ≤3 μm from the parent dendrite were identified as dendritic spine. Spines were counted in maximum-intensity projections of the z-stacks using ImageJ software (National Institute of Health, Bethesda, MD, USA), only if they appeared continuous with the parent dendrite. Spine density was calculated by quantifying the number of spines per dendritic segment, and normalized to 10 μm of dendrite length. The classification of morphological spine types was performed as described (Tyler and Pozzo-Miller, 2003). Spine types were grouped as immature-shaped thin (type-III) spines, mature-shaped spines, which included type-I (stubby) and type II (mushroom) spines, according published criteria (Boda et al., 2004). Images of pyramidal neurons in primary cultures were deconvolved in Fiji (Schindelin et al., 2012), followed by NeuronStudio processing in order to categorize dendritic spines into three morphological types based on their length and the dimensions of the spine head and neck: stubby, mushroom and thin (Rodriguez et al., 2008).
The total length of secondary and tertiary apical dendrites analyzed for Figure 1 was: (i) sal/sal: 1244.58 μm from 20 dendritic segments of 9 cells; (ii) sal/amph: 1041.50 μm from 18 dendritic segments of 8 cells; (iii) amph/sal: 1078.29 μm from 20 dendritic segments of 8 cells; (iv) amph/amph: 1321.83 μm from 23 dendritic segments of 9 cells. For Figure 2, dissociated culture: (i) vehicle: 1432.03 μm from 34 dendritic segments of 10 cells; (ii) amph: 2084.79 μm from 40 dendritic segments of 14 cells; organotypic culture: (iii) vehicle: 840.93 μm from 26 dendritic segments of 12 cells; (iv) amph: 1204.28 μm from 34 dendritic segments of 15 cells. For Figure 3B: (i) eYFP-vehicle: 840.93 μm from 26 dendritic segments of 13 cells; (ii) dnCdk5-vehicle: 1528.945 μm from 34 dendritic segments of 14 cells; (iii) rosco-vehicle: 1227.58 μm from 16 dendritic segments of 6 cells; (iv) shCd5-vehicle: 689.22 μm from 18 dendritic segments of 3 cells; (v) eYFP-amph: 1204.28 μm from 34 dendritic segments of 12 cells; (vi) dnCdk5-amph: 1433.04 μm from 31 dendritic segments of 16 cells; (vii) rosco-amph: 1784.51 μm from 27 dendritic segments of 9 cells; (viii) shCd5-amph: 427.9 μm from 18 dendritic segments of 4 cells. For Figure 3E; (ix) eYFP-vehicle: 840.93 μm from 26 dendritic segments of 13 cells; (x) amph-vehicle: 1528.945 μm from 12 dendritic segments of 12 cells; (xi) p25-vehicle: 1241.19 μm from 26 dendritic segments of 7 cells; (xii) p25-amph: 1918.05 μm from 43 dendritic segments of 13 cells; (xiii) p35-vehicle: 581.81 μm from 13 dendritic segments of 8 cells; (xiv) p35-amph: 855.81 μm from 21 dendritic segments of 13 cells. For Figure 4B: (i) vehicle: 642.49 μm from 11 dendritic segments of 3 cells; (ii) 15 min amph: 717.48 μm from 11 dendritic segments of 3 cells; (iii) 30 min amph: 474.65 μm from six dendritic segments of 3 cells; (iv) 60 min amph: 749.2 μm from 20 dendritic segments of 4 cells. For Figure 4E; (v) vehicle-control: 792 μm from 19 dendritic segments of 7 cells; (vi) vehicle-ALLN: 304.6 μm from 8 dendritic segments of 4 cells; (vii) amph-control: 750 μm from 20 dendritic segments of 7 cells; (viii) amph-ALLN: 350.89 μm from 9 dendritic segments of 6 cells.
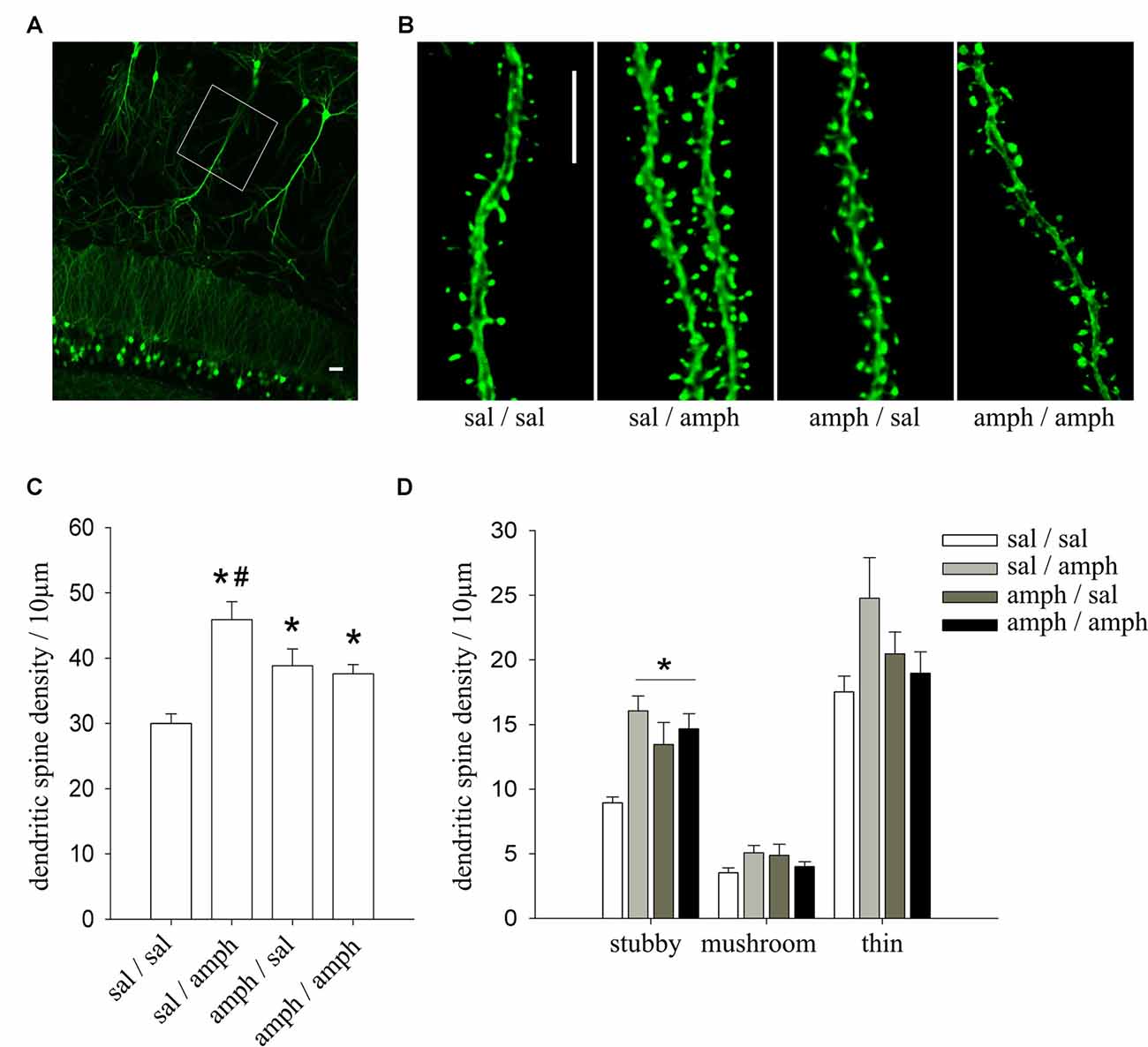
Figure 1. In vivo exposure to amphetamine increased dendritic spine density in hippocampal CA1 pyramidal neurons. (A) Representative image of the CA1 region of a thy1-green fluorescent protein (GFP) mouse showing GFP-expressing pyramidal neurons. White box indicates the region where dendritic segments were imaged at high magnification; scale bar = 20 μm. (B) Representative dendritic segments of CA1 pyramidal neurons of thy1-GFP mice; scale bar = 5 μm. (C) Dendritic spine density expressed as the number of spines normalized to 10 μm of dendritic length; data are mean ± SEM. *Indicates statistically significant differences vs. sal/sal, #vs. all treatment groups (one way ANOVA, *p = 0.0001). (D) Density of the three different morphological spine types (stubby, mushroom and thin). *Indicates statistically significant differences (one way ANOVA, *p = 0.001). N = 4 mice; n = 3 dendritic segments per mouse.

Figure 2. In vitro amphetamine exposure increased dendritic spine density in hippocampal neurons maintained in either dissociated or organotypic slice culture. (A) Representative images of hippocampal pyramidal neurons maintained in dissociated cultures or organotypic slice cultures (scale bar = 20 μm). (B) Representative examples of secondary or tertiary dendritic segments of cultured pyramidal neurons (17–19 DIV) expressing yellow fluorescent protein (YFP) and exposed to 50 μM amphetamine (amph) or vehicle for 48 h (scale bar = 5 μm). (C) Dendritic spine density (one-way ANOVA, *p = 0.001). (D) Representative maximum-intensity projections of confocal z-stacks of dendritic segments of YFP-expressing CA1 pyramidal neurons in slice culture (10 DIV), exposed to either 50 μM amphetamine or vehicle for 48 h (scale bar = 5 μm). (E) Dendritic spine density (one-way ANOVA, *p = 0.005). (F) Density of three different morphological spine types (stubby, mushroom and thin; one-way ANOVA, *p = 0.05). Samples of dissociated cultures (four different cultures): vehicle n = 10 neurons, amph n = 14. Samples of organotypic slice cultures (three different cultures): vehicle n = 13, amph n = 13.
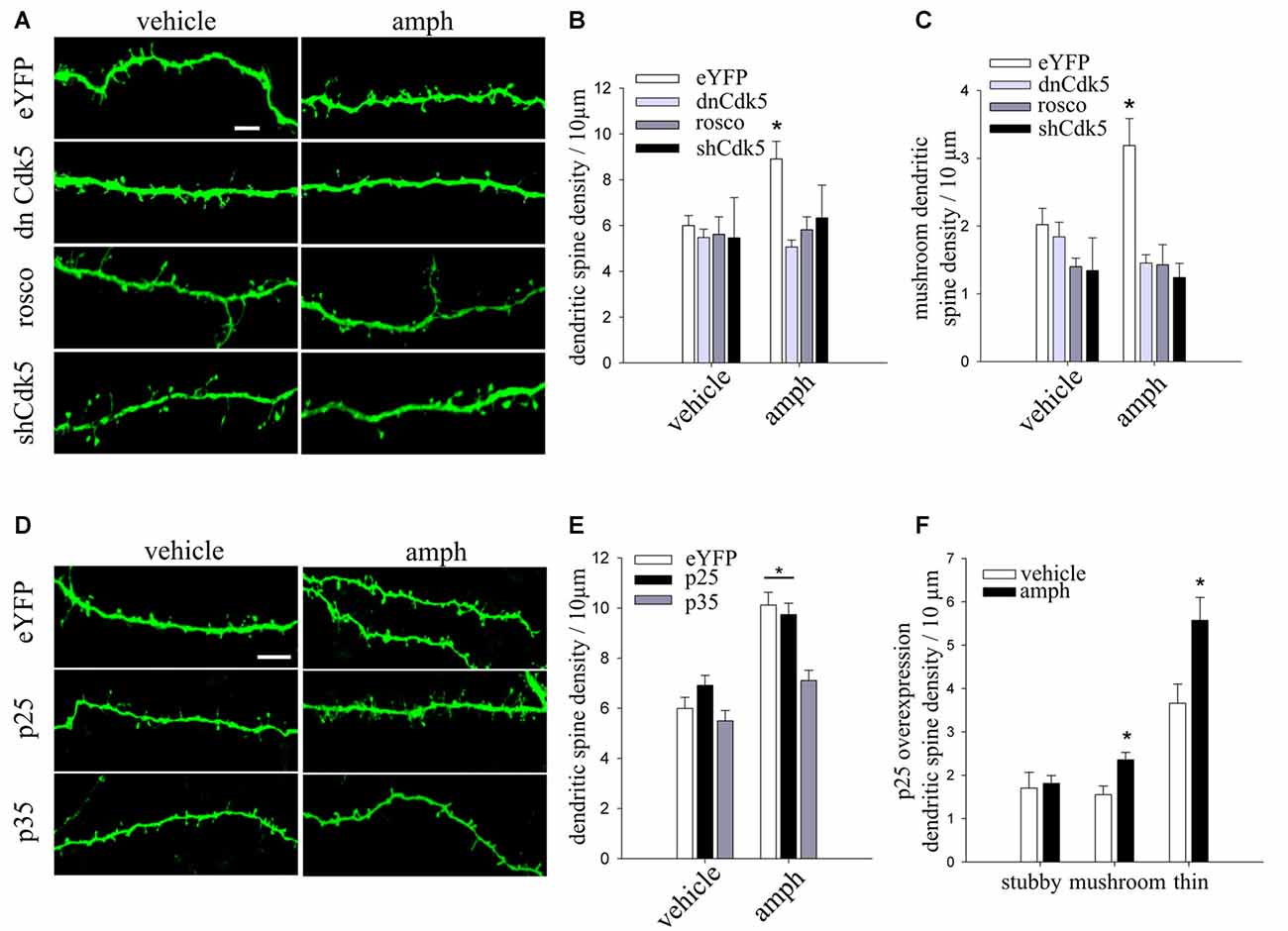
Figure 3. Cdk5 knockdown or pharmacological inhibition prevent amphetamine-evoked increase in dendritic spine density. (A) Representative maximum-intensity projections of confocal z-stacks of dendritic segments of YFP-expressing CA1 pyramidal neurons in slice culture (10 DIV), biolistically co-transfected with either dnCdk5 or a short hairpin RNA (shRNA) against Cdk5, and exposed to 50 μM amphetamine (amph), vehicle, or roscovitine (10 μM) for 48 h (scale bar = 5 μm). (B) Dendritic spine density in each condition (two-way ANOVA, *p = 0.01). (C) Density of mushroom spine type (two-way ANOVA, *p = 0.0001). (D) Representative maximum-intensity projections of confocal z-stacks of dendritic segments of CA1 pyramidal neurons in slice culture (10 DIV), biolistically co-transfected with either p35 or p25, and exposed to 50 μM amphetamine (amph) or vehicle for 48 h (scale bar = 5 μm). (E) Dendritic spine density for each condition (two-way ANOVA, *p = 0.005). (F) Density of three different morphological spine types (stubby, mushroom and thin; one-way ANOVA, *p = 0.01 and *p = 0.05 respectively). N = an average of nine neurons for each condition from three to eight slices of three different cultures.
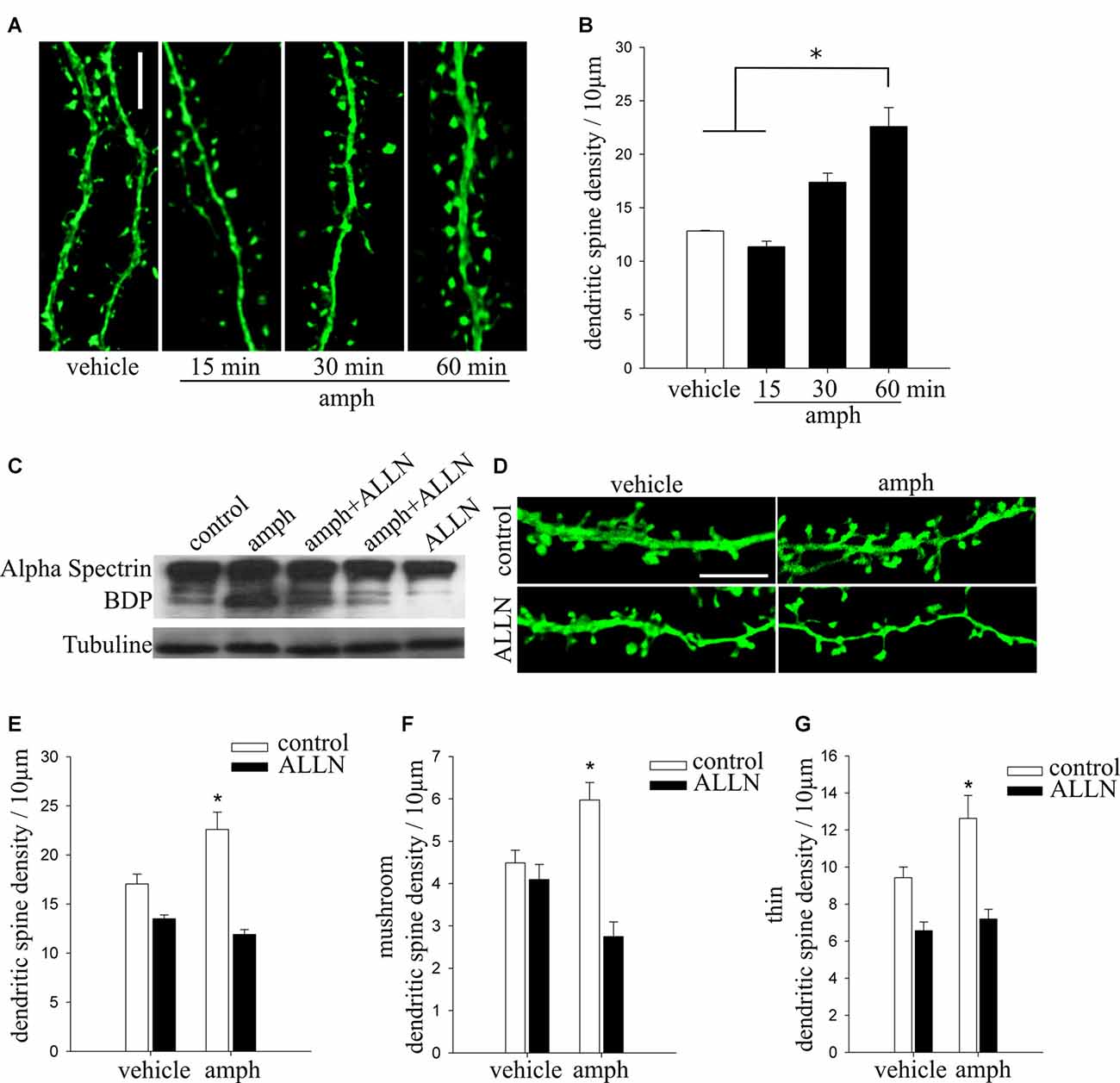
Figure 4. Calpain activity is necessary for amphetamine to increase dendritic spine density in hippocampal pyramidal neurons. (A) Representative dendritic segments of cultured pyramidal neurons (17–19 DIV) expressing YFP and exposed to either vehicle or amphetamine 50 μM (amph) for 15, 30 and 60 min (scale bar = 5 μm). (B) Dendritic spine density (one-way ANOVA, *p = 0.05). (C) Representative Western immunoblot showing that amphetamine increases calpain proteolytic activity. Total homogenate of hippocampal neuronal cultures (17–19 DIV) from control (line 1) or amphetamine-treated for 60 min (line 2) or pretreated with N-Acetyl-L-leucyl-L-norleucinal (ALLN; 10 μM) for 30 min and then exposed to amphetamine for 60 min (lines 3 and 4) or treated with ALLN alone as a control (line 5). (D) Representative maximum-intensity projections of confocal z-stacks of secondary or tertiary dendritic segments of cultured pyramidal neurons (17–19 DIV) expressing YFP and exposed to vehicle, 50 μM amph for 60 min, ALLN for 30 min followed by amph for 60 min, or ALLN alone (scale bar = 5 μm). (E) Dendritic spine density for all treatments (two way ANOVA, *p = 0.000001). (F,G) Density of mushroom (mush) (F) and thin (G) spine types (one way ANOVA, *p = 0.0001 and *p = 0.01, respectively).
Western Immunoblots
Crude synaptosomal fractions were obtained as described (Mlewski et al., 2008). All samples were valued for protein concentration (DC Protein Assay Kit, Bio-Rad Laboratory, Hercules, CA, USA) and equal protein amounts (25 μg) were separated into 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred to Nitrocellulose membranes (Bio-Rad Laboratory, Hercules, CA, USA) as previously described (Krapacher et al., 2010). Membranes were incubated overnight at 4°C with their primary antibodies diluted in 1% skim milk in TBST. The antibodies used were anti-α-tubulin (1:3000, DM1A; Sigma-Aldrich, St. Louis, MO, USA) and anti-spectrin (1:1000, Millipore Corporation, Billerica, MA, USA). Membranes were then washed three times in TBST and incubated with a Licor secondary antibody (1:15,000, IRDye® 800CW LI-COR Biosciences, Lincoln, NE, USA) or a horseradish peroxidase–conjugated antibody (1:2000; Jackson ImmunoResearch Laboratories, West Grove, PA, USA) for 1 h at room temperature. After two washes with TBST and two washes with TBS, bands were visualized using Odissey scanner (LI-COR Biosciences, Lincoln, NE, USA) or a chemiluminescence detection kit (ECL; Amersham Life Science, Buckinghamshire, England) according to the primary antibody used.
Statistical Analyses
All data were analyzed using Statistica 7.0 (Statsoft, Inc., Tulsa, OK, USA). Statistical analyses were performed using ANOVAs. Significant main effects were further analyzed through Fisher’s LSD post hoc test (Fisher’s least significance difference test with a Type I error set at 0.05).
Results
In Vivo Administration of Amphetamine Increases Dendritic Spine Density in CA1 Hippocampal Pyramidal Neurons
We have previously shown that adolescent rats develop and express locomotor sensitization after an amphetamine two-injection protocol (4 mg/kg) when they were trained and evaluated in a novel environment (Mlewski et al., 2016). To test if this exposure to amphetamine affects dendritic spines in hippocampal neurons, we used the same protocol in male P35–40 thy1-GFP transgenic mice (line M), which express GFP in pyramidal neurons in a Golgi-type fashion (Feng et al., 2000; Figures 1A,B).
Amphetamine induced a significant increase of the density of spine in apical dendrites of CA1 pyramidal neurons in all groups (sal/amph, amph/sal and amph/amph) compared to sal/sal controls. ANOVA revealed a significant main effect of amphetamine (one-way ANOVA F(3,30) = 10.304, *p < 0.0001), indicating that mice treated with amphetamine have higher dendritic spine density than sal/sal controls (Figure 1C). Amphetamine also modulated the morphology of individual spines, increasing the density of stubby spines (one-way ANOVA F(3,30) = 7.3287, *p = 0.001; Figure 1D).
These results demonstrate that amphetamine, administrated in a two-injection protocol known to trigger locomotor sensitization, increases dendritic spine density in CA1 pyramidal neurons favoring the mature stubby type.
In Vitro Administration of Amphetamine Increases Dendritic Spine Density in Hippocampal Pyramidal Neurons
To develop an in vitro assay amenable for testing molecular mechanisms, we exposed YFP-expressing cultured hippocampal neurons to amphetamine (50 μM) for 48 h (Figure 2A). Consistent with in vivo observations, amphetamine significantly increased spine density in YFP-expressing neurons maintained in primary culture (Figures 2B,C; one-way ANOVA F(1,22) = 16.576, *p = 0.001). In addition, amphetamine (50 μM) induced a significant increase of spine density in YFP-expressing CA1 pyramidal neurons maintained in hippocampal slice cultures (Figures 2A,D,E; one way ANOVA F(1,26) = 9.9457, *p = 0.005). Similar to its effects in vivo, amphetamine increased the density of mature stubby spines (one way ANOVA, F(1,26) = 5.5401, *p = 0.05), and also of the mushroom type (one way ANOVA, F(1,26) = 4.7140, *p = 0.05; Figure 2F).
Together with the observations in vivo, these results in two different in vitro preparations provide strong evidence of amphetamine-induced structural plasticity in pyramidal neurons of the hippocampus.
Cdk5 Activity Is Necessary for Amphetamine to Increase Dendritic Spine Density in Hippocampal Pyramidal Neurons
Considering that Cdk5 plays a role in cocaine-induced dendritic spine formation in the nucleus accumbens (Norrholm et al., 2003), we tested whether it is also involved in the effects of amphetamine on CA1 hippocampal pyramidal neurons. To this end, we used three strategies: (1) slice cultures were biolistically co-transfected with YFP and dominant negative Cdk5 (dnCdk5), which abolishes endogenous Cdk5 activation by p35/p25 (Tsai et al., 1994); (2) pharmacological inhibition of Cdk5 kinase activity with the specific inhibitor roscovitine; and (3) slice cultures were biolistically co-transfected with YFP and a Cdk5 short harping interfering RNA (shCdk5) to suppress Cdk5 expression.
Expression of dnCdk5, shRNA-mediated Cdk5 knockdown, and pharmacological Cdk5 inhibition with roscovitine all prevented the increase in spine density induced by amphetamine (Figures 3A,B). In addition, all three Cdk5 manipulations blocked the increase in the density of mushroom spines induced by amphetamine (two-way ANOVA F(3,68) = 7.47556 *p = 0.0001; Figure 3C). None of these Cdk5 manipulations by themselves affected dendritic spine density or morphology (two-way ANOVA F(3,68) = 3.6015, *p = 0.01; Figures 3A,B). These results demonstrate that amphetamine’s effects on dendritic spines require intact Cdk5 activity.
Cdk5 activity is induced by the interaction with a regulatory subunit, either p35 or p25 (proteolytic fragment of p35; Tsai et al., 1994; Tang et al., 1995). In addition, the level of p35 is the rate-limiting factor for the activity of Cdk5 (Takahashi et al., 2005). To examine the role of p35 and p25 in the amphetamine effect on spine density, CA1 pyramidal neurons in slice cultures were biolistically co-transfected with YFP and either p35 or p25. Overexpression of p35 prevented the increase in spine density induced by amphetamine. In contrast, p25-expressing neurons exposed to amphetamine showed the typical increase in spine density (Figures 3D,E; two-way ANOVA F(2,60) = 3.8051 *p = 0.005). In addition, p25-expressing neurons exposed to amphetamine showed higher densities of mature mushroom spine as well as immature thin spines (Figure 3F; one-way ANOVA, F(1,18) = 8.1234 *p = 0.01 and F(1,18) = 5.7472 *p = 0.05 respectively).
Together, these results demonstrate that amphetamine’s effect on spine density requires proper levels of p35 to regulate Cdk5 activity, and that overexpression of p25 does not affect the increase of dendritic spine density induced by amphetamine.
Calpain Activity Is Necessary for Amphetamine-Induced Increase Dendritic Spine Density in Hippocampal Pyramidal Neurons
Proteolytic cleavage of p35 by the Ca2+ activated cysteine protease calpain (Liu X. et al., 2008) results in the formation of p25, which causes hyperactivation of Cdk5 (Kusakawa et al., 2000; Lee et al., 2000). Considering that the only way to generate p25 is by proteolytic cleavage of p35 protein by calpain (Kusakawa et al., 2000), and that amphetamine increases intracellular Ca2+ levels (Licata and Pierce, 2003; Gnegy et al., 2004; Mills et al., 2007), we tested if calpain activity is involved in amphetamine’s effects on dendritic spine density in hippocampal neurons.
We first tested if exposure to amphetamine activates calpain activity. Cell lysates from hippocampal neurons were prepared 60 min after application of amphetamine (50 μM) because that is when its effect on dendritic spine density becomes statistically significant (Figures 4A,B; one way ANOVA, F(3,24) = 3.5324 *p = 0.05). Calpain activity was examined by Western immunoblots using an antibody that recognizes well-characterized calpain-specific α-spectrin breakdown products (α-spectrin-BDP; Zhang et al., 2009; Ma et al., 2012). A 60 min exposure to amphetamine induced robust calpain activity (Figure 4C, line 2), an effect blocked by preincubation with the calpain inhibitor ALLN (Figure 4C, lines 3, 4).
Finally, we tested if inhibition of calpain activity prevents amphetamine’s effect on spine density. Indeed, the calpain inhibitor ALLN abolished the increase in spine density observed 60 min after exposure to amphetamine (Figures 4D,E; two way ANOVA, F(3,57) = 12.18 *p = 0.000001). Calpain inhibition also prevented the amphetamine-induced increase in the densities of mushroom and thin spines types (Figures 4F,G; one way ANOVA, F(3,57) = 11.021 *p = 0.0001 and F(3,57) = 7.9454 *p = 0.01, respectively).
Taken altogether, our findings demonstrate that amphetamine increases dendritic spine density in hippocampal neurons by triggering the proteolytic cleavage of p35 by calpain, generating p25 to activate Cdk5.
Discussion
This study provides strong findings and novel insight into the molecular bases by which amphetamine increases dendritic spine density in hippocampal neurons. Using thy1-GFP transgenic mice, we show that amphetamine administrated in a two-injection protocol increased dendritic spine density in CA1 pyramidal neurons and shifted the density of morphological types in favor of mature stubby spines. Using two different in vitro systems, we also show this effect of amphetamine requires proteolytic cleavage of p35 by calpain to generate p25 that in turn activates Cdk5.
A great number of studies into the pathophysiology of drug abuse have concentrated on the dopaminergic reward system, valuing the neural changes induced during addiction, relapse, and abstinence in this natural reward regions (Chen et al., 2010; Deadwyler, 2010; Collo et al., 2014). Even though the dopaminergic system is central of intense research of motivational and reward-related learning, the neurobiological basis of reward learning and memory are uncompleted without the contribution of the hippocampus, which sends a main output to the reward system. Considering that this brain structure lies upstream of the striatal dopaminergic circuit, synaptic plasticity within the hippocampus may alter the transmission of information throughout the brain’s reward system (Grace et al., 2007). Several studies exhibit that psychostimulant drugs produce persistent alterations in other forebrain regions as well, in particular, in the orbital frontal cortex (Fein et al., 2002; Franklin et al., 2002; Paulus et al., 2002; Adinoff et al., 2003; Bolla et al., 2003; Matochik et al., 2003). Moreover, human psychostimulants users, as well as cocaine-exposed monkeys, show deficits in the execution of cognitive tasks (Rogers et al., 1999; Ornstein et al., 2000; Jentsch et al., 2002). Also, studies in rats have shown that chronic cocaine-treatment produce deficits in discrimination learning (Schoenbaum et al., 2004). Together, these studies indicate that psychostimulant drugs also produce lasting alterations in other brain areas related with higher-order associative learning, in addition to changes in reward circuit. However, little is known, about the cellular and molecular mechanisms of the effects of psychostimulant drugs in the hippocampus. In the few studies performed in rodents, psychostimulant-induced structural plasticity in non-reward regions like the hippocampus, has been studied using long chronic administration regimens (Rademacher et al., 2006; Shen et al., 2006; Boikess et al., 2010; Ahn et al., 2013). Here, we show for the first time that amphetamine administrated in a two-injection protocol increases dendritic spine density in CA1 pyramidal neurons of thy1-GFP transgenic mice. Interestingly, the largest effect was observed in mice that had the first experience with the drug (group sal-amph or amph/sal), suggesting that the first exposure to amphetamine is enough to induce structural plasticity in the hippocampus. Amphetamine also modulated the morphology of individual spines by increasing the density of stubby spines. This type of spines is the most abundant during development and is considered very plastic structures, strongly coupled to the dendritic shaft (Noguchi et al., 2005; Schmidt and Eilers, 2009). Interestingly, similar to its effects in vivo, amphetamine increased the density of stubby spines and also mushroom type in vitro condition. Taken together, these data provide strong evidences of amphetamine-induced structural plasticity in hippocampal pyramidal neurons, both in vivo and in vitro conditions. The particular morphology of a dendritic spine may play an important role in determining its function, a concept that has been extensively reviewed (Shepherd, 1996; Yuste and Majewska, 2001; Nimchinsky et al., 2002). Two morphological features of dendritic spines seem to be critical for their role as biochemical compartments: the geometrical dimensions of the head and neck, which affect the diffusion and/or accumulation and retention of ions and molecules generated by synaptic signaling at the postsynaptic density (e.g., free ionized Ca2+, activated CaMKII). In addition, the morphology and size of spines correlate with the strength of the synapses they bear (Pierce and Lewin, 1994). The spine head volume is directly proportional to the number of docked vesicles at the active zone of presynaptic terminals (Harris and Sultan, 1995; Boyer et al., 1998; Schikorski and Stevens, 1999) and to the number of AMPARs (Nusser et al., 1998). Moreover, the expression of AMPARs within individual spines of different morphologies was mapped by 2-photon uncaging of MNI-glutamate, revealing that mushroom (Type-II) spines show larger AMPAR-mediated responses than thin (Type-III) spines (Matsuzaki et al., 2001). These studies provide functional evidence that larger spines represent stronger synapses, as defined by their expression of AMPARs (Kasai et al., 2003). The induction of LTP leads to an increase in spine head size, while long-term depression (LTD) causes spine shrinkage and retraction, which correlates well with the synaptic levels of AMPARs observed after the induction of such bi-directional synaptic plasticity. Despite the large literature, it remains unclear to what extent the plasticity of dendritic spines drives the pathological behavior evident in psychostimulants addiction.
Previously, we demonstrated that both acute and chronic amphetamine treatment in vivo induced a significant increase in Cdk5 activity (Mlewski et al., 2008). Other studies explored the relationship between the activity of Cdk5 and dendritic spine maintenance (Fu et al., 2007; Lai et al., 2012). Here, we show that the expression and activity of Cdk5 is necessary for amphetamine to modulate dendritic spine density and their morphology: shRNA-mediated Cdk5 knockdown, pharmacological inhibition with roscovitine, and suppression of endogenous Cdk5 activity with a dominant negative mutant, all completely abolished the amphetamine-induced increase in spine density in hippocampal CA1 pyramidal neurons maintained in slice culture. These results are consistent with previous reports of reduction in cocaine-induced increase in spine density in the nucleus accumbens by roscovitine (Norrholm et al., 2003). In addition, dendritic spine density is lower in hippocampal neurons of the double inducible-p35cKO and p39KO mice, indicating that Cdk5 activity is necessary for the maintenance of the density of dendritic spine, at least in hippocampal neurons in adult naïve mice (Mita et al., 2016).
Considering that Cdk5 and its activators p35 and p25 have been involved in the effects of chronic exposures to cocaine (Bibb et al., 2001), methamphetamine (Chen and Chen, 2005), and amphetamine (Mlewski et al., 2008), we further investigated the role of p35 and p25 in amphetamine’s effect on dendritic spine density in hippocampal CA1 pyramidal neurons. Overexpression of p25 did not affect the amphetamine-evoked increase in spine density. This result is in line with the increase in dendritic spine density and number of synapses in an inducible transgenic mouse that transiently overexpress p25 in the hippocampus (Fischer et al., 2005). In contrast, overexpression of p35 completely abolished the effect of amphetamine on dendritic spine density. Consistent with this result, it has been demonstrated that high p35 levels binds microtubules through its N- terminal domain and removes it from Cdk5, rendering the kinase inactive (Shah and Lahiri, 2014). In addition, p35 binding to microtubules promotes microtubule bundling (He et al., 2008) and their stabilization (Hou et al., 2007). Moreover, p35 phosphorylation at Ser-8 and Thr-138 sites by Cdk5 suppresses its calpain-catalyzed truncation, but increases its F-actin bundling and binding activities, which in turn would stabilize actin filaments (Kamei et al., 2007; He et al., 2011).
We previously demonstrated a transient increase of p25 expression in the synaptosomal fraction of rat striatum after acute amphetamine treatment (Mlewski et al., 2008). These enhanced p25 levels were associated with a significant rise in Cdk5 activity and an enrichment of p21-activated kinase 1 (PAK1) phosphorylated on Thr-212, the specific site for Cdk5 kinase (Mlewski et al., 2008). PAK1 is involved in actin remodeling, dendritic spine morphogenesis, excitatory synapse formation, and synaptic plasticity (Zhang, 2005). It has been suggested that Cdk5 allows PAK1 to switch from an active to an inactive state and thereby mediate a rapid and dynamic regulation of the actin cytoskeleton (Rashid et al., 2001), thus facilitating dendritic spine formation. Altogether, these observations support the model that p25-regulated Cdk5 activity is specifically involved in cellular events underlying psychostimulant-induced synaptic plasticity.
Considering that calpain-mediated cleavage of p35 is the only way to generate p25 (Kusakawa et al., 2000; Kerokoski et al., 2004; Zhu et al., 2011), and that cocaine and amphetamine increase intracellular Ca2+ levels, we next tested whether the amphetamine effect on dendritic spine density in hippocampal neurons requires calpain activity. Calpains are a family of cysteine, non-lysosomal, calcium-dependent proteases whose substrates are involved in signal transduction, cytoskeleton remodeling, cell differentiation, vesicular trafficking, axonal degeneration and neuronal death (Liu J. et al., 2008). A unique feature of this protease is that its processed substrates often acquire different functions (Ma, 2013). An example of this is p35, which is cleaved to p25 by calpain. The resultant Cdk5/p25 complex exerts several functions associated to corticogenesis, cytoskeleton dynamic, synaptic plasticity, and dendritic spine formation (Kerokoski et al., 2004; Fischer et al., 2005; Mlewski et al., 2008; Barnett and Bibb, 2011; Lai et al., 2012). Considering that Ca2+-dependent processes are fast and transient, we first determined the time course of amphetamine’s effect on dendritic spine density in hippocampal cultures. Sixty minutes of amphetamine exposure were sufficient to significantly increase dendritic spine density, suggesting rapid post-translational modification of specific proteins, such as calpain and its substrates. Consistently, strong calpain proteolytic activity was detected in amphetamine-treated cultures, and pharmacological inhibition prevented the increase in dendritic spine density in hippocampal CA1 pyramidal neurons exposed to amphetamine. In line with these observations, conditional deletion of calpain-1 and calpain-2 results in lower dendritic spine density in the apical and basal dendrites of hippocampal CA1 pyramidal neurons (Amini et al., 2013). Interestingly, calpain inhibition also prevented the amphetamine-induced increase in the densities of mushroom and thin spines types, precisely the same dendritic spines types that are increased by amphetamine in p25-overexpressing neurons. Initially it was thought that spine synapses originate from established synapses first on the dendritic shaft by the enlargement and then by narrowing of the neck and the emergence of the spine head (Miller and Peters, 1981). Otherwise, filopodia have been considered as precursors of mature dendritic spines. Considering that most of spines have a presynaptic partner (Gray, 1959; Harris and Kater, 1994), it has been suggested that dendritic filopodia search for presynaptic axon terminals to form synapses, giving rise to more developed spines after the first contact (Jontes and Smith, 2000; Yuste and Bonhoeffer, 2004). Furthermore, time-lapse experiments in hippocampal slices maintained in culture, subsequently demonstrated that filopodia are highly dynamic structures, extending and retracting in short time periods (Dailey and Smith, 1996). Besides, it has been shown that filopodia from cultured neurons establish contact with axons, and its number decreased in correlation with the increase in spines number (Ziv and Smith, 1996). Thus, the initial contact with axons triggers the formation of spines from filopodia. Finally, in vivo imaging of dendritic spines in the neocortex of developing and adult animals from multiphoton microscopy supports the fact that spines constantly form by seeking out presynaptic partners in the surrounding neuropil and stabilizing into functional spines of varied morphology, a process driven by sensory experience (Lendvai et al., 2000; Trachtenberg et al., 2002; Holtmaat et al., 2005; Knott et al., 2006).
In summary, we demonstrate that amphetamine increases dendritic spine density in hippocampal neurons through the proteolytic cleavage of p35 into p25 by calpain, which in turn results in p25-depedent activation of Cdk5. p25-activated Cdk5 likely endows different properties to Cdk5, resulting in different roles at discrete levels of organization and different regulatory strategies in the recognition of particular substrates. The apparent lack of p25 in naïve conditions, together with its ability to enhance synaptic plasticity, suggests that it is shaped only during particular events, such as during learning (Fischer et al., 2005) or exposure to drugs of abuse (Mlewski et al., 2008). It is possible that in turn, the Cdk5/p25 complex could modulate critical substrates involved in the reorganization and dynamics of the actin cytoskeleton (Dietz et al., 2012), a fundamental mechanism in the formation of dendritic spines and the excitatory synapses they are part of. Revealing the cellular and synaptic mechanisms underlying drug addiction and the identification of the molecules involved will yield novel opportunities of potential therapeutic targets for the treatment of addictive disorders.
Author Contributions
All authors were responsible for the design of the study. SF, GF, VD and MVP performed experimental procedures and collected data. ECM, SF and CA conducted statistical data analysis. LM and CAC performed cell culture and FAK conducted biochemical assays. The findings were interpreted and the manuscript was drafted by SF, GF, LP-M, DHM and MGP. All authors critically reviewed content and approved the final version for publication.
Funding
This work was supported by grants from Argentinean Agencia Nacional de Promoción Científica y Tecnológica (FONCyT PICT 07-2168 and FONCyT PICT 11-0892), from Consejo Nacional de Investigaciones Científicas y Técnicas—CONICET PIP 2014, and traveling fellowships from the International Society for Neurochemistry (ISN), the Journal of Cell Science and the Company of Biologists, Limited (Travel fellowship to SF).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Gonzalo Quasollo and Mariano Bisbal for their help with the design of shCdk5. We also thank Gastón Calfa for helping with experimental design, and Laura Gastaldi and Mónica Remedi for technical support. We would particularly like to thank Damian Revillo and Evelin Cotella for helpful discussions, and the vivarium technicians for their assistance with animal husbandry. Confocal microscopy was performed at the Centro de Microscopía Óptica y Confocal de Avanzada, CIQUIBIC-INIMEC-CIBICI, CONICET, Universidad Nacional de Córdoba, Argentina.
References
Adinoff, B., Devous, M. D., Cooper, D. B., Best, S. E., Chandler, P., Harris, T., et al. (2003). Resting regional cerebral blood flow and gambling task performance in cocaine-dependent subjects and healthy comparison subjects. Am. J. Psychiatry 160, 1892–1894. doi: 10.1176/appi.ajp.160.10.1892
Ahn, S., Linsenbardt, D. N., Lapish, C. C., and Rubchinsky, L. L. (2013). Repeated injections of D-Amphetamine evoke rapid and dynamic changes in phase synchrony between the prefrontal cortex and hippocampus. Front. Behav. Neurosci. 7:92. doi: 10.3389/fnbeh.2013.00092
Alonso, M., Medina, J. H., and Pozzo-Miller, L. (2004). ERK1/2 activation is necessary for BDNF to increase dendritic spine density in hippocampal CA1 pyramidal neurons. Learn. Mem. 11, 172–178. doi: 10.1101/lm.67804
Amini, M., Ma, C., Farazifard, R., Zhu, G., Zhang, Y., Vanderluit, J., et al. (2013). Conditional disruption of calpain in the CNS alters dendrite morphology, impairs LTP, and promotes neuronal survival following injury. J. Neurosci. 33, 5773–5784. doi: 10.1523/jneurosci.4247-12.2013
Angelo, M., Plattner, F., and Giese, K. P. (2006). Cyclin-dependent kinase 5 in synaptic plasticity, learning and memory. J. Neurochem. 99, 353–370. doi: 10.1111/j.1471-4159.2006.04040.x
Barnett, D. G. S., and Bibb, J. A. (2011). The role of Cdk5 in cognition and neuropsychiatric and neurological pathology. Brain Res. Bull. 85, 9–13. doi: 10.1016/j.brainresbull.2010.11.016
Benavides, D. R., and Bibb, J. A. (2004). Role of Cdk5 in drug abuse and plasticity. Ann. N Y Acad. Sci. 1025, 335–344. doi: 10.1196/annals.1316.041
Berke, J. D., and Hyman, S. E. (2000). Addiction, dopamine and the molecular mechanisms of memory. Neuron 25, 515–532. doi: 10.1016/s0896-6273(00)81056-9
Bibb, J. A., Chen, J., Taylor, J. R., Svenningsson, P., Nishi, A., Snyder, G. L., et al. (2001). Effects of chronic exposure to cocaine are regulated by the neuronal protein Cdk5. Nature 410, 376–380. doi: 10.1038/35066591
Bignante, E. A., Rodriguez Manzanares, P. A., Mlewski, E. C., Bertotto, M. E., Bussolino, D. F., Paglini, G., et al. (2008). Involvement of septal Cdk5 in the emergence of excessive anxiety induced by stress. Eur. Neuropsychopharmacol. 18, 578–588. doi: 10.1016/j.euroneuro.2008.02.007
Boda, B., Alberi, S., Nikonenko, I., Node-Langlois, R., Jourdain, P., Moosmayer, M., et al. (2004). The mental retardation protein PAK3 contributes to synapse formation and plasticity in hippocampus. J. Neurosci. 24, 10816–10825. doi: 10.1523/jneurosci.2931-04.2004
Boikess, S. R., O’Dell, S. J., and Marshall, J. F. (2010). A sensitizing D-amphetamine dose regimen induces long-lasting spinophilin and VGLUT1 protein upregulation in the rat diencephalon. Neurosci. Lett. 469, 49–54. doi: 10.1016/j.neulet.2009.11.042
Bolla, K. I., Eldreth, D. A., London, E. D., Kiehl, K. A., Mouratidis, M., Contoreggi, C., et al. (2003). Orbitofrontal cortex dysfunction in abstinent cocaine abusers performing a decision-making task. Neuroimage 19, 1085–1094. doi: 10.1016/s1053-8119(03)00113-7
Boyer, C., Schikorski, T., and Stevens, C. F. (1998). Comparison of hippocampal dendritic spines in culture and in brain. J. Neurosci. 18, 5294–5300.
Brinkkoetter, P. T., Pippin, J. W., and Shankland, S. J. (2010). Cyclin I-Cdk5 governs survival in post-mitotic cells. Cell Cycle 9, 1729–1731. doi: 10.4161/cc.9.9.11471
Chae, T., Kwon, Y. T., Bronson, R., Dikkes, P., Li, E., and Tsai, L.-H. (1997). Mice lacking p35, a neuronal specific activator of Cdk5, display cortical lamination defects, seizures, and adult lethality. Neuron 18, 29–42. doi: 10.1016/s0896-6273(01)80044-1
Chen, P.-C., and Chen, J.-C. (2005). Enhanced Cdk5 activity and p35 translocation in the ventral striatum of acute and chronic methamphetamine-treated rats. Neuropsychopharmacology 30, 538–549. doi: 10.1038/sj.npp.1300604
Chen, B. T., Hopf, F. W., and Bonci, A. (2010). Synaptic plasticity in the mesolimbic system: therapeutic implications for substance abuse. Ann. N Y Acad. Sci. 1187, 129–139. doi: 10.1111/j.1749-6632.2009.05154.x
Chen, J.-R., Wang, Y.-J., and Tseng, G.-F. (2003). The effect of epidural compression on cerebral cortex: a rat model. J. Neurotrauma 20, 767–780. doi: 10.1089/089771503767869999
Cheung, Z. H., and Ip, N. Y. (2012). Cdk5: a multifaceted kinase in neurodegenerative diseases. Trends Cell Biol. 22, 169–175. doi: 10.1016/j.tcb.2011.11.003
Chuang, J. Z., Yeh, T. Y., Bollati, F., Conde, C., Canavosio, F., Caceres, A., et al. (2005). The dynein light chain tctex-1 has a dynein-independent role in actin remodeling during neurite outgrowth. Dev. Cell 9, 75–86. doi: 10.1016/j.devcel.2005.04.003
Cingolani, L. A., and Goda, Y. (2008). Actin in action: the interplay between the actin cytoskeleton and synaptic efficacy. Nat. Rev. Neurosci. 9, 344–356. doi: 10.1038/nrn2373
Collo, G., Cavalleri, L., and Spano, P. F. (2014). Structural plasticity in mesencephalic dopaminergic neurons produced by drugs of abuse: critical role of BDNF and dopamine. Front. Pharmacol. 5:259. doi: 10.3389/fphar.2014.00259
Dailey, M. E., and Smith, S. J. (1996). The dynamics of dendritic structure in developing hippocampal slices. J. Neurosci. 16, 2983–2994.
De Falco, M., Fedele, V., De Luca, L., Penta, R., Cottone, G., Cavallotti, I., et al. (2004). Evaluation of cyclin D1 expression and its subcellular distribution in mouse tissues. J. Anat. 205, 405–412. doi: 10.1111/j.0021-8782.2004.00347.x
Deadwyler, S. A. (2010). Electrophysiological correlates of abused drugs. Ann. N Y Acad. Sci. 1187, 140–147. doi: 10.1111/j.1749-6632.2009.05155.x
Dhavan, R., and Tsai, L. H. (2001). A decade of CDK5. Nat. Rev. Mol. Cell Biol. 2, 749–759. doi: 10.1038/35096019
Dietz, D. M., Sun, H., Lobo, M. K., Cahill, M. E., Chadwick, B., Gao, V., et al. (2012). Rac1 is essential in cocaine-induced structural plasticity of nucleus accumbens neurons. Nat. Neurosci. 15, 891–896. doi: 10.1038/nn.3094
Engmann, O., Giralt, A., Gervasi, N., Marion-Poll, L., Gasmi, L., Filhol, O., et al. (2015). DARPP-32 interaction with adducin may mediate rapid environmental effects on striatal neurons. Nat. Commun. 6:10099. doi: 10.1038/ncomms10099
Fein, G., Di Sclafani, V., and Meyerhoff, D. J. (2002). Prefrontal cortical volume reduction associated with frontal cortex function deficit in 6-week abstinent crack-cocaine dependent men. Drug Alcohol Depend. 68, 87–93. doi: 10.1016/s0376-8716(02)00110-2
Feng, G., Mellor, R. H., Bernstein, M., Keller-Peck, C., Nguyen, Q. T., Wallace, M., et al. (2000). Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron 28, 41–51. doi: 10.1016/s0896-6273(00)00084-2
Fischer, M., Kaech, S., Wagner, U., Brinkhaus, H., and Matus, A. (2000). Glutamate receptors regulate actin-based plasticity in dendritic spines. Nat. Neurosci. 3, 887–894. doi: 10.1038/78791
Fischer, A., Sananbenesi, F., Pang, P. T., Lu, B., and Tsai, L. H. (2005). Opposing roles of transient and prolonged expression of p25 in synaptic plasticity and hippocampus-dependent memory. Neuron 48, 825–838. doi: 10.1016/j.neuron.2005.10.033
Fischer, A., Sananbenesi, F., Schrick, C., Spiess, J., and Radulovic, J. (2002). Cyclin-dependent kinase 5 is required for associative learning. J. Neurosci. 22, 3700–3707.
Franklin, T. R., Acton, P. D., Maldjian, J. A., Gray, J. D., Croft, J. R., Dackis, C. A., et al. (2002). Decreased gray matter concentration in the insular, orbitofrontal, cingulate, and temporal cortices of cocaine patients. Biol. Psychiatry 51, 134–142. doi: 10.1016/s0006-3223(01)01269-0
Fu, W.-Y., Chen, Y., Sahin, M., Zhao, X.-S., Shi, L., Bikoff, J. B., et al. (2007). Cdk5 regulates EphA4-mediated dendritic spine retraction through an ephexin1-dependent mechanism. Nat. Neurosci. 10, 67–76. doi: 10.1038/nn1811
Gnegy, M. E., Khoshbouei, H., Berg, K. A., Javitch, J. A., Clarke, W. P., Zhang, M., et al. (2004). Intracellular Ca2+ regulates amphetamine-induced dopamine efflux and currents mediated by the human dopamine transporter. Mol. Pharmacol. 66, 137–143. doi: 10.1124/mol.66.1.137
Grace, A. A., Floresco, S. B., Goto, Y., and Lodge, D. J. (2007). Regulation of firing of dopaminergic neurons and control of goal-directed behaviors. Trends Neurosci. 30, 220–227. doi: 10.1016/j.tins.2007.03.003
Gray, E. G. (1959). Electron microscopy of synaptic contacts on dendrite spines of the cerebral cortex. Nature 183, 1592–1593. doi: 10.1038/1831592a0
Harris, K. M., and Kater, S. B. (1994). Dendritic spines: cellular specializations imparting both stability and flexibility to synaptic function. Annu. Rev. Neurosci. 17, 341–371. doi: 10.1146/annurev.neuro.17.1.341
Harris, K. M., and Sultan, P. (1995). Variation in the number, location and size of synaptic vesicles provides an anatomical basis for the nonuniform probability of release at hippocampal CA1 synapses. Neuropharmacology 34, 1387–1395. doi: 10.1016/0028-3908(95)00142-s
He, L., Hou, Z., and Qi, R. Z. (2008). Calmodulin binding and Cdk5 phosphorylation of p35 regulate its effect on microtubules. J. Biol. Chem. 283, 13252–13260. doi: 10.1074/jbc.M706937200
He, L., Zhang, Z., Yu, Y., Ahmed, S., Cheung, N. S., and Qi, R. Z. (2011). The neuronal p35 activator of Cdk5 is a novel F-actin binding and bundling protein. Cell. Mol. Life Sci. 68, 1633–1643. doi: 10.1007/s00018-010-0562-9
Holtmaat, A. J. G. D., Trachtenberg, J. T., Wilbrecht, L., Shepherd, G. M., Zhang, X., Knott, G. W., et al. (2005). Transient and persistent dendritic spines in the neocortex in vivo. Neuron 45, 279–291. doi: 10.1016/j.neuron.2005.01.003
Hou, Z., Li, Q., He, L., Lim, H. Y., Fu, X., Nam, S. C., et al. (2007). Microtubule association of the neuronal p35 activator of Cdk5. J. Biol. Chem. 282, 18666–18670. doi: 10.1074/jbc.C700052200
Hyman, S. E., and Malenka, R. C. (2001). Addiction and the brain: the neurobiology of compulsion and its persistence. Nat. Rev. Neurosci. 2, 695–703. doi: 10.1038/35094560
Jentsch, J. D., Olausson, P., De La Garza, R. II, and Taylor, J. R. (2002). Impairments of reversal learning and response perseveration after repeated, intermittent cocaine administrations to monkeys. Neuropsychopharmacology 26, 183–190. doi: 10.1016/s0893-133x(01)00355-4
Jontes, J. D., and Smith, S. J. (2000). Filopodia, spines, and the generation of synaptic diversity. Neuron 27, 11–14. doi: 10.1016/s0896-6273(00)00003-9
Kamei, H., Saito, T., Ozawa, M., Fujita, Y., Asada, A., Bibb, J. A., et al. (2007). Suppression of calpain-dependent cleavage of the CDK5 activator p35 to p25 by site-specific phosphorylation. J. Biol. Chem. 282, 1687–1694. doi: 10.1074/jbc.M610541200
Kasai, H., Matsuzaki, M., Noguchi, J., Yasumatsu, N., and Nakahara, H. (2003). Structure-stability-function relationships of dendritic spines. Trends Neurosci. 26, 360–368. doi: 10.1016/s0166-2236(03)00162-0
Katz, L. C. (1987). Local circuitry of identified projection neurons in cat visual cortex brain slices. J. Neurosci. 7, 1223–1249.
Kerokoski, P., Suuronen, T., Salminen, A., Soininen, H., and Pirttilä, T. (2004). Both N-methyl-D-aspartate (NMDA) and non-NMDA receptors mediate glutamate-induced cleavage of the cyclin-dependent kinase 5 (cdk5) activator p35 in cultured rat hippocampal neurons. Neurosci. Lett. 368, 181–185. doi: 10.1016/j.neulet.2004.07.007
Knott, G. W., Holtmaat, A., Wilbrecht, L., Welker, E., and Svoboda, K. (2006). Spine growth precedes synapse formation in the adult neocortex in vivo. Nat. Neurosci. 9, 1117–1124. doi: 10.1038/nn1747
Kolb, B., Gorny, G., Li, Y., Samaha, A.-N., and Robinson, T. E. (2003). Amphetamine or cocaine limits the ability of later experience to promote structural plasticity in the neocortex and nucleus accumbens. Proc. Natl. Acad. Sci. U S A 100, 10523–10528. doi: 10.1073/pnas.1834271100
Krapacher, F. A., Mlewski, E. C., Ferreras, S., Pisano, V., Paolorossi, M., Hansen, C., et al. (2010). Mice lacking p35 display hyperactivity and paradoxical response to psychostimulants. J. Neurochem. 114, 203–214. doi: 10.1111/j.1471-4159.2010.06748.x
Kunda, P., Paglini, G., Quiroga, S., Kosik, K., and Cáceres, A. (2001). Evidence for the involvement of tiam1 in axon formation. J. Neurosci. 21, 2361–2372.
Kusakawa, G. I., Saito, T., Onuki, R., Ishiguro, K., Kishimoto, T., and Hisanaga, S. I. (2000). Calpain-dependent proteolytic cleavage of the p35 cyclin-dependent kinase 5 activator to p25. J. Biol. Chem. 275, 17166–17172. doi: 10.1074/jbc.M907757199
Kwon, Y. T., Tsai, L. H., and Crandall, J. E. (1999). Callosal axon guidance defectes in p35−/− mice. J. Comp. Neurol. 415, 218–229. doi: 10.1002/(SICI)1096-9861(19991213)415:23.0.CO;2-F
Lai, K. O., and Ip, N. Y. (2013). Structural plasticity of dendritic spines: the underlying mechanisms and its dysregulation in brain disorders. Biochim. Biophys. Acta 1832, 2257–2263. doi: 10.1016/j.bbadis.2013.08.012
Lai, K.-O., Wong, A. S. L., Cheung, M.-C., Xu, P., Liang, Z., Lok, K.-C., et al. (2012). TrkB phosphorylation by Cdk5 is required for activity-dependent structural plasticity and spatial memory. Nat. Neurosci. 15, 1506–1515. doi: 10.1038/nn.3237
Lee, M. S., Kwon, Y. T., Li, M., Peng, J., Friedlander, R. M., and Tsai, L. H. (2000). Neurotoxicity induces cleavage of p35 to p25 by calpain. Nature 405, 360–364. doi: 10.1038/35012636
Lendvai, B., Stern, E. A., Chen, B., and Svoboda, K. (2000). Experience-dependent plasticity of dendritic spines in the developing rat barrel cortex in vivo. Nature 404, 876–881. doi: 10.1038/35009107
Li, Y., Kolb, B., and Robinson, T. E. (2003). The location of persistent amphetamine-induced changes in the density of dendritic spines on medium spiny neurons in the nucleus accumbens and caudate-putamen. Neuropsychopharmacology 28, 1082–1085. doi: 10.1038/sj.npp.1300115
Licata, S. C., and Pierce, R. C. (2003). The roles of calcium/calmodulin-dependent and Ras/mitogen-activated protein kinases in the development of psychostimulant-induced behavioral sensitization. J. Neurochem. 85, 14–22. doi: 10.1046/j.1471-4159.2003.01662.x
Liu, X., Gong, H., Li, X., and Zhou, W. (2008). Monitoring calcium concentration in neurons with cameleon. J. Biosci. Bioeng. 105, 106–109. doi: 10.1263/jbb.105.106
Liu, J., Liu, M. C., and Wang, K. K. W. (2008). Calpain in the CNS: from synaptic function to neurotoxicity. Sci. Signal. 1:re1. doi: 10.1126/stke.114re1
Ma, M. (2013). Role of calpains in the injury-induced dysfunction and degeneration of the mammalian axon. Neurobiol. Dis. 60, 61–79. doi: 10.1016/j.nbd.2013.08.010
Ma, S., Liu, S., Huang, Q., Xie, B., Lai, B., Wang, C., et al. (2012). Site-specific phosphorylation protects glycogen synthase kinase-3β from calpain-mediated truncation of its N and C termini. J. Biol. Chem. 287, 22521–22532. doi: 10.1074/jbc.M111.321349
Matochik, J. A., London, E. D., Eldreth, D. A., Cadet, J.-L., and Bolla, K. I. (2003). Frontal cortical tissue composition in abstinent cocaine abusers: a magnetic resonance imaging study. Neuroimage 19, 1095–1102. doi: 10.1016/s1053-8119(03)00244-1
Matsuzaki, M., Ellis-Davies, G. C., Nemoto, T., Miyashita, Y., Iino, M., and Kasai, H. (2001). Dendritic spine geometry is critical for AMPA receptor expression in hippocampal CA1 pyramidal neurons. Nat. Neurosci. 4, 1086–1092. doi: 10.1038/nn736
Miller, M., and Peters, A. (1981). Maturation of rat visual cortex. II. A combined Golgi-electron microscope study of pyramidal neurons. J. Comp. Neurol. 203, 555–573. doi: 10.1002/cne.902030402
Mills, K., Ansah, T. A., Ali, S. F., Mukherjee, S., and Shockley, D. C. (2007). Augmented behavioral response and enhanced synaptosomal calcium transport induced by repeated cocaine administration are decreased by calcium channel blockers. Life Sci. 81, 600–608. doi: 10.1016/j.lfs.2007.06.028
Mita, N., He, X., Sasamoto, K., Mishiba, T., and Ohshima, T. (2016). Cyclin-dependent kinase 5 regulates dendritic spine formation and maintenance of cortical neuron in the mouse brain. Cereb. Cortex 26, 967–976. doi: 10.1093/cercor/bhu264
Mlewski, E. C., Arias, C., and Paglini, G. (2016). Association between the expression of amphetamine-induced behavioral sensitization and Cdk5/p35 activity in dorsal striatum. Behav. Neurosci. 130, 114–122. doi: 10.1037/bne0000118
Mlewski, E. C., Krapacher, F. A., Ferreras, S., and Paglini, G. (2008). Transient enhanced expression of Cdk5 activator p25 after Acute and chronic d -amphetamine administration. Ann. N Y Acad. Sci. 1139, 89–102. doi: 10.1196/annals.1432.039
Moore, C. D., Thacker, E. E., Larimore, J., Gaston, D., Underwood, A., Kearns, B., et al. (2007). The neuronal Arf GAP centaurin α1 modulates dendritic differentiation. J. Cell Sci. 120, 2683–2693. doi: 10.1242/jcs.006346
Nath, R., Davis, M., Probert, A. W., Kupina, N. C., Ren, X., Schielke, G. P., et al. (2000). Processing of cdk5 activator p35 to its truncated form (p25) by calpain in acutely injured neuronal cells. Biochem. Biophys. Res. Commun. 274, 16–21. doi: 10.1006/bbrc.2000.3070
Nikolic, M., Dudek, H., Kwon, Y. T., Ramos, Y. F. M., and Tsai, L. H. (1996). The cdk5/p35 kinase is essential for neurite outgrowth during neuronal differentiation. Genes Dev. 10, 816–825. doi: 10.1101/gad.10.7.816
Nimchinsky, E. A., Sabatini, B. L., and Svoboda, K. (2002). Structure and function of dendritic spines. Annu. Rev. Physiol. 64, 313–353. doi: 10.1146/annurev.physiol.64.081501.160008
Noguchi, J., Matsuzaki, M., Ellis-Davies, G. C. R., and Kasai, H. (2005). Spine-neck geometry determines NMDA receptor-dependent Ca2+ signaling in dendrites. Neuron 46, 609–622. doi: 10.1016/j.neuron.2005.03.015
Norrholm, S. D., Bibb, J. A., Nestler, E. J., Ouimet, C. C., Taylor, J. R., and Greengard, P. (2003). Cocaine-induced proliferation of dendritic spines in nucleus accumbens is dependent on the activity of cyclin-dependent kinase-5. Neuroscience 116, 19–22. doi: 10.1016/s0306-4522(02)00560-2
Nusser, Z., Lujan, R., Laube, G., Roberts, J. D. B., Molnar, E., and Somogyi, P. (1998). Cell type and pathway dependence of synaptic AMPA receptor number and variability in the hippocampus. Neuron 21, 545–559. doi: 10.1016/s0896-6273(00)80565-6
Odajima, J., Wills, Z. P., Ndassa, Y. M., Terunuma, M., Kretschmannova, K., Deeb, T. Z., et al. (2011). Cyclin E constrains Cdk5 activity to regulate synaptic plasticity and memory formation. Dev. Cell 21, 655–668. doi: 10.1016/j.devcel.2011.08.009
Ornstein, T. J., Iddon, J. L., Baldacchino, A. M., Sahakian, B. J., London, M., Everitt, B. J., et al. (2000). Profiles of cognitive dysfunction in chronic amphetamine and heroin abusers. Neuropsychopharmacology 23, 113–126. doi: 10.1016/s0893-133x(00)00097-x
Paglini, G., Peris, L., Diez-Guerra, J., Quiroga, S., and Cáceres, A. (2001). The Cdk5–p35 kinase associates with the Golgi apparatus and regulates membrane traffic. EMBO Rep. 2, 1139–1144. doi: 10.1093/embo-reports/kve250
Paglini, G., Pigino, G., Kunda, P., Morfini, G., Maccioni, R., Quiroga, S., et al. (1998). Evidence for the participation of the neuron-specific CDK5 activator P35 during laminin-enhanced axonal growth. J. Neurosci. 18, 9858–9869.
Patrick, G. N., Zukerberg, L., Nikolic, M., de la Monte, S., Dikkes, P., and Tsai, L.-H. (1999). Conversion of p35 to p25 deregulates Cdk5 activity and promotes neurodegeneration. Nature 402, 615–622. doi: 10.1038/45159
Patzke, H., Maddineni, U., Ayala, R., Morabito, M., Volker, J., Dikkes, P., et al. (2003). Partial rescue of the p35−/− brain phenotype by low expression of a neuronal-specific enolase p25 transgene. J. Neurosci. 23, 2769–2778.
Paulus, M. P., Hozack, N. E., Zauscher, B. E., Frank, L., Brown, G. G., Braff, D. L., et al. (2002). Behavioral and functional neuroimaging evidence for prefrontal dysfunction in methamphetamine-dependent subjects. Neuropsychopharmacology 26, 53–63. doi: 10.1016/s0893-133x(01)00334-7
Pierce, J. P., and Lewin, G. R. (1994). An ultrastructural size principle. Neuroscience 58, 441–446. doi: 10.1016/0306-4522(94)90071-x
Rademacher, D. J., Kovacs, B., Shen, F., Napier, T. C., and Meredith, G. E. (2006). The neural substrates of amphetamine conditioned place preference: implications for the formation of conditioned stimulus-reward associations. Eur. J. Neurosci. 24, 2089–2097. doi: 10.1111/j.1460-9568.2006.05066.x
Rasakham, K., Schmidt, H. D., Kay, K., Huizenga, M. N., Calcagno, N., Christopher Pierce, R., et al. (2014). Synapse density and dendritic complexity are reduced in the prefrontal cortex following seven days of forced abstinence from cocaine self-administration. PLoS One 9:e102524. doi: 10.1371/journal.pone.0102524
Rashid, T., Banerjee, M., and Nikolic, M. (2001). Phosphorylation of Pak1 by the p35/Cdk5 kinase affects neuronal morphology. J. Biol. Chem. 276, 49043–49052. doi: 10.1074/jbc.m105599200
Robinson, T. E., and Kolb, B. (1997). Persistent structural modifications in nucleus accumbens and prefrontal cortex neurons produced by previous experience with amphetamine. J. Neurosci. 17, 8491–8497.
Robinson, T. E., and Kolb, B. (2004). Structural plasticity associated with exposure to drugs of abuse. Neuropharmacology 47, 33–46. doi: 10.1016/j.neuropharm.2004.06.025
Rodriguez, A., Ehlenberger, D. B., Dickstein, D. L., Hof, P. R., and Wearne, S. L. (2008). Automated three-dimensional detection and shape classification of dendritic spines from fluorescence microscopy images. PLoS One 3:e1997. doi: 10.1371/journal.pone.0001997
Rogers, R. D., Everitt, B. J., Baldacchino, A., Blackshaw, A. J., Swainson, R., Wynne, K., et al. (1999). Dissociable deficits in the decision-making cognition of chronic amphetamine abusers, opiate abusers, patients with focal damage to prefrontal cortex, and tryptophan-depleted normal volunteers evidence for monoaminergic mechanisms. Neuropsychopharmacology 20, 322–339. doi: 10.1016/s0893-133x(98)00091-8
Scheibel, M. E., Lindsay, R. D., Tomiyasu, U., and Scheibel, A. B. (1975). Progressive dendritic changes in aging human cortex. Exp. Neurol. 47, 392–403. doi: 10.1016/0014-4886(75)90072-2
Schikorski, T., and Stevens, C. F. (1999). Quantitative fine-structural analysis of olfactory cortical synapses. Proc. Natl. Acad. Sci. U S A 96, 4107–4112. doi: 10.1073/pnas.96.7.4107
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., et al. (2012). Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682. doi: 10.1038/nmeth.2019
Schmidt, H., and Eilers, J. (2009). Spine neck geometry determines spino-dendritic cross-talk in the presence of mobile endogenous calcium binding proteins. J. Comput. Neurosci. 27, 229–243. doi: 10.1007/s10827-009-0139-5
Schoenbaum, G., Saddoris, M. R., Ramus, S. J., Shaham, Y., and Setlow, B. (2004). Cocaine-experienced rats exhibit learning deficits in a task sensitive to orbitofrontal cortex lesions. Eur. J. Neurosci. 19, 1997–2002. doi: 10.1111/j.1460-9568.2004.03274.x
Schubert, V., and Dotti, C. G. (2007). Transmitting on actin: synaptic control of dendritic architecture. J. Cell Sci. 120, 205–212. doi: 10.1242/jcs.03337
Shah, K., and Lahiri, D. K. (2014). Cdk5 activity in the brain-multiple paths of regulation. J. Cell Sci. 127, 2391–2400. doi: 10.1242/jcs.147553
Shen, F., Meredith, G. E., and Napier, T. C. (2006). Amphetamine-induced place preference and conditioned motor sensitization requires activation of tyrosine kinase receptors in the hippocampus. J. Neurosci. 26, 11041–11051. doi: 10.1523/jneurosci.2898-06.2006
Shepherd, G. M. (1996). The dendritic spine: a multifunctional integrative unit. J. Neurophysiol. 75, 2197–2210.
Smith, D. S., and Tsai, L.-H. (2002). Cdk5 behind the wheel: a role in trafficking and transport? Trends Cell Biol. 12, 28–36. doi: 10.1016/s0962-8924(01)02181-x
Stoppini, L., Buchs, P.-A., and Muller, D. (1991). A simple method for organotypic cultures of nervous tissue. J. Neurosci. Methods 37, 173–182. doi: 10.1016/0165-0270(91)90128-m
Sun, K. H., Chang, K. H., Clawson, S., Ghosh, S., Mirzaei, H., Regnier, F., et al. (2011). Glutathione-S-transferase P1 is a critical regulator of Cdk5 kinase activity. J. Neurochem. 118, 902–914. doi: 10.1111/j.1471-4159.2011.07343.x
Tada, T., and Sheng, M. (2006). Molecular mechanisms of dendritic spine morphogenesis. Curr. Opin. Neurobiol. 16, 95–101. doi: 10.1016/j.conb.2005.12.001
Takahashi, S., Ohshima, T., Cho, A., Sreenath, T., Iadarola, M. J., Pant, H. C., et al. (2005). Increased activity of cyclin-dependent kinase 5 leads to attenuation of cocaine-mediated dopamine signaling. Proc. Natl. Acad. Sci. U S A 102, 1737–1742. doi: 10.1073/pnas.0409456102
Tang, D., Yeung, J., Lee, K. Y., Matsushita, M., Matsui, H., Tomizawa, K., et al. (1995). An isoform of the neuronal cyclin-dependent kinase 5 (Cdk5) activator. J. Biol. Chem. 270, 26897–26903. doi: 10.1074/jbc.270.45.26897
Trachtenberg, J. T., Chen, B. E., Knott, G. W., Feng, G., Sanes, J. R., Welker, E., et al. (2002). Long-term in vivo imaging of experience-dependent synaptic plasticity in adult cortex. Nature 420, 788–794. doi: 10.1038/nature01273
Tsai, L.-H., Delalle, I., Caviness, V. S., Chae, T., and Harlow, E. (1994). p35 is a neural-specific regulatory subunit of cyclin-dependent kinase 5. Nature 371, 419–423. doi: 10.1038/371419a0
Tseng, G. F., and Hu, M. E. (1996). Axotomy induces retraction of the dendritic arbor of adult rat rubrospinal neurons. Acta Anat. 155, 184–193. doi: 10.1159/000147803
Tyler, W. J., and Pozzo-Miller, L. D. (2001). BDNF enhances quantal neurotransmitter release and increases the number of docked vesicles at the active zones of hippocampal excitatory synapses. J. Neurosci. 21, 4249–4258.
Tyler, W. J., and Pozzo-Miller, L. (2003). Miniature synaptic transmission and BDNF modulate dendritic spine growth and form in rat CA1 neurones. J. Physiol. 553, 497–509. doi: 10.1113/jphysiol.2003.052639
Wang, T.-J., Chen, J.-R., Wang, Y.-J., and Tseng, G.-F. (2009). The cytoarchitecture and soma-dendritic arbors of the pyramidal neurons of aged rat sensorimotor cortex: an intracellular dye injection study. Neuroscience 158, 776–785. doi: 10.1016/j.neuroscience.2008.10.025
Yuste, R., and Bonhoeffer, T. (2001). Morphological changes in dendritic spines associated with long-term synaptic plasticity. Annu. Rev. Neurosci. 24, 1071–1089. doi: 10.1146/annurev.neuro.24.1.1071
Yuste, R., and Bonhoeffer, T. (2004). Genesis of dendritic spines: insights from ultrastructural and imaging studies. Nat. Rev. Neurosci. 5, 24–34. doi: 10.1038/nrn1300
Yuste, R., and Majewska, A. (2001). On the function of dendritic spines. Neuroscientist 7, 387–395. doi: 10.1177/107385840100700508
Zhang, H. (2005). A GIT1/PIX/Rac/PAK signaling module regulates spine morphogenesis and synapse formation through MLC. J. Neurosci. 25, 3379–3388. doi: 10.1523/jneurosci.3553-04.2005
Zhang, Z., Larner, S. F., Liu, M. C., Zheng, W., Hayes, R. L., and Wang, K. K. W. (2009). Multiple αII-spectrin breakdown products distinguish calpain and caspase dominated necrotic and apoptotic cell death pathways. Apoptosis 14, 1289–1298. doi: 10.1007/s10495-009-0405-z
Zhu, J., Li, W., and Mao, Z. (2011). Cdk5: mediator of neuronal development, death and the response to DNA damage. Mech. Ageing Dev. 132, 389–394. doi: 10.1016/j.mad.2011.04.011
Keywords: amphetamine, hippocampus, dendritic spines, Cdk5/p25, calpain, organotypic slice cultures
Citation: Ferreras S, Fernández G, Danelon V, Pisano MV, Masseroni L, Chapleau CA, Krapacher FA, Mlewski EC, Mascó DH, Arias C, Pozzo-Miller L and Paglini MG (2017) Cdk5 Is Essential for Amphetamine to Increase Dendritic Spine Density in Hippocampal Pyramidal Neurons. Front. Cell. Neurosci. 11:372. doi: 10.3389/fncel.2017.00372
Received: 08 September 2017; Accepted: 08 November 2017;
Published: 24 November 2017.
Edited by:
Claudio Rivera, Aix-Marseille University, FranceReviewed by:
Pirta Elina Hotulainen, Minerva Institute for Medical Research, FinlandGonzalo Flores, Benemérita Universidad Autónoma de Puebla, Mexico
Copyright © 2017 Ferreras, Fernández, Danelon, Pisano, Masseroni, Chapleau, Krapacher, Mlewski, Mascó, Arias, Pozzo-Miller and Paglini. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María G. Paglini, gpaglini@immf.uncor.edu
† These authors have contributed equally to this work.