 Lízbeth Ayala-Domínguez1,2Leslie Olmedo-Nieva2,3
Lízbeth Ayala-Domínguez1,2Leslie Olmedo-Nieva2,3 J. Omar Muñoz-Bello2Adriana Contreras-Paredes2
J. Omar Muñoz-Bello2Adriana Contreras-Paredes2 Joaquín Manzo-Merino4Imelda Martínez-Ramírez2
Joaquín Manzo-Merino4Imelda Martínez-Ramírez2 Marcela Lizano2,5*
Marcela Lizano2,5*- 1Programa de Doctorado en Ciencias Biomédicas, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 2Unidad de Investigación Biomédica en Cáncer, Instituto Nacional de Cancerología-Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 3Programa de Doctorado en Ciencias Bioquímicas, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 4Cátedras CONACyT-Instituto Nacional de Cancerología, Mexico City, Mexico
- 5Departamento de Medicina Genómica y Toxicología Ambiental, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, Mexico City, Mexico
Solid tumors carry out the formation of new vessels providing blood supply for growth, tumor maintenance, and metastasis. Several processes take place during tumor vascularization. In angiogenesis, new vessels are derived from endothelial cells of pre-existing vessels; while in vasculogenesis, new vessels are formed de novo from endothelial progenitor cells, creating an abnormal, immature, and disorganized vascular network. Moreover, highly aggressive tumor cells form structures similar to vessels, providing a pathway for perfusion; this process is named vasculogenic mimicry (VM), where vessel-like channels mimic the function of vessels and transport plasma and blood cells. VM is developed by numerous types of aggressive tumors, including ovarian carcinoma which is the second most common cause of death among gynecological cancers. VM has been associated with poor patient outcome and survival in ovarian cancer, although the involved mechanisms are still under investigation. Several signaling molecules have an important role in VM in ovarian cancer, by regulating the expression of genes related to vascular, embryogenic, and hypoxic signaling pathways. In this review, we provide an overview of the current knowledge of the signaling molecules involved in the promotion and regulation of VM in ovarian cancer. The clinical implications and the potential benefit of identification and targeting of VM related molecules for ovarian cancer treatment are also discussed.
Introduction
Ovarian cancer is the second most common and lethal gynecological cancer (1). Among ovarian cancer types, the epithelial ovarian cancer accounts for almost 90% of such malignancy (2), which is usually diagnosed in advanced aggressive stages due to its asymptomatic nature. Extensive tumor invasion, peritoneal metastases, and treatment failure are frequent in advanced epithelial ovarian cancer (3).
The normal physiology of the ovary is characterized by increased permeability of blood vessels during follicular development, ovulation, and subsequent formation of the corpus luteum, with cyclic changes in the formation, differentiation, and regression of ovarian vasculature (4). These vascular processes are deregulated in ovarian cancer, which is characterized by intense neovascularization (5, 6). Neovasculature in ovarian cancer is formed not only from endothelial cells but also from endothelial progenitor cells and/or cells from the tumor itself, allowing the supply of blood and nutrients to the tumor with great efficiency (7).
The versatility of the vascularization processes in ovarian cancer could partially explain its aggressive nature and the limited efficacy of anti-angiogenic therapies (8). An alternative vascularization process, vasculogenic mimicry (VM), has been shown to increase after anti-angiogenic treatment with bevacizumab, in preclinical models of ovarian cancer (9). This finding suggests that VM could be a strategy for escaping anti-angiogenic treatment, highlighting the importance to study the mechanisms involved in vascular remodeling.
In this review, we provide an overview of the current knowledge of the mechanisms and signaling molecules involved in the promotion and regulation of VM in ovarian cancer, its clinical implications and the potential benefit of therapeutic approaches based on the identification and targeting of VM related molecules.
Tumor Vascularization Processes in Ovarian Cancer
The study of the vascularization processes in solid tumors has gained importance due to its implication in growth and metastasis, as well as its possible implication for anti-angiogenic treatment resistance (10). The most studied tumor vascularization process is angiogenesis, although tumor tissue has the capacity to generate its own vasculature from alternative mechanisms such as vasculogenesis, vessel co-option, and VM (11–13).
Angiogenesis
Angiogenesis is a highly regulated process aimed to produce new blood vessels with a key role in development and postnatal life; it is also involved in invasion, growth, and metastasis of solid tumors (14, 15). The onset of angiogenesis occurs in response to hypoxia or ischemia where pro-angiogenic signals overcome anti-angiogenic signals. The vascular endothelial growth factor A (VEGF-A) is the master regulator of angiogenesis, both in physiological and pathological conditions (16). During angiogenesis activation, a complex signaling cascade begins, leading to the proliferation of endothelial cells (ECs) that assemble new vascular networks from the pre-existing vessels, increasing permeability and leakage, and restoring the supply of oxygen and nutrients toward the tumor mass (15, 17).
Angiogenesis is essential for the growth of ovarian cancer cells and their spreading to the peritoneum. VEGF-A has been associated with peritoneal ECs proliferation, migration, and formation of tube-like structures (18). The inhibition of VEGF-A does not revert these processes, suggesting that another pro-angiogenic factors secreted by surrounding ovarian cancer cells or their microenvironment could be involved in the angiogenic activation of peritoneal ECs during metastasis (18, 19). A high level of pro-angiogenic signals has been associated with the formation of ascites, a frequent feature of advanced ovarian cancer (20, 21).
Vasculogenesis
Vasculogenesis, a de novo vessel formation process, is distinguished by the in situ differentiation of ECs from myeloid cells or endothelial progenitor cells (EPCs). This process takes place at the beginning of vascular development and during post-natal life (11, 22). Myeloid cells and EPCs are recruited by pro-angiogenic or pro-inflammatory factors to the tumor vascular bed, where they differentiate into ECs and give place to neovasculature (23–25). Vasculogenesis has a modest impact on tumor vascularization when the angiogenesis pathway is active, however, it is recognized as an important rescue process when this pathway is blocked (10, 26). For instance, when angiogenesis is inhibited after anti-angiogenic treatment or radiotherapy, myeloid cells, and EPCs are recruited by the stroma-derived factor 1 (SDF-1) in response to an increased level of hypoxia-inducible factor 1α (HIF-1α) (10, 26).
Vasculogenesis has an important role in ovarian cancer. It has been related to treatment resistance as a consequence of the overexpression of matrix metalloproteinase 2 and 9 (MMP-2 and MMP-9) after radiotherapy (27). Furthermore, CD34+ EPCs from peripheral blood incorporate into vasculogenic active sites (25) as well as CD11b+ and CD11c+ myeloid cells, recruited by SDF-1 and β-defensins, that contribute to vasculogenesis (28). β-defensins chemoattract CD11c+ dendritic cell precursors and then VEGF-A induces endothelial-like specialization mediated by VEGF receptor 2 (VEGFR-2); interestingly, recruitment of CD11c+ cells has also been found in ascites (28).
Vessel Co-option
Vessel co-option is a process that differs from angiogenesis; instead of inducing the proliferation of ECs, tumor cells grow by adhering to nearby blood vessels (15). Different patterns of vessel co-option have been described in brain, lung, and liver cancers (12). In glioblastoma, CDC42+ CD44+ tumor cells migrate toward a blood vessel in response to a bradykinin gradient created by ECs; when these cells reach the vessel, they fuse with the pericytes or adhere to the basement membrane (12). Vessel co-option has been observed in a mouse model of ovarian cancer (29), where endostatin inhibited vessel co-option by blocking the attachment of ovarian cancer cells to peritoneal vessels through integrins α5β1.
It has been proposed that after the tumor grows by vessel co-option, co-opted vessels regress, and the tumor enters into an avascular phase followed by the induction of peritumoral angiogenesis (30). Vessel co-option facilitates the metastasis of tumor cells since it increases their motility and migration. There is evidence that tumors can switch between angiogenic and non-angiogenic growth during progression and that they can contain angiogenic and non-angiogenic areas (12). The association between vessel co-option and resistance to anti-angiogenic treatment is not clear, since vessel co-option could be one cause of the resistance to anti-angiogenic treatment or it could be a consequence of the aggressive nature of cancer cells in response to anti-angiogenic treatment (10, 31).
Vasculogenic Mimicry
VM is a process by which tumor cells form capillary-like structures, mimicking the embryonic vascular network pattern, without inducing the proliferation of ECs (15). This process increases blood perfusion, allows tumor cells to obtain oxygen and nutrients, and promotes cancer progression (13, 32). It has been proposed that VM is carried out through cancer stem cell (CSC) trans-differentiation into endothelial-like cells (13, 33). Moreover, tumor cells involved in VM resemble mesenchymal cells derived from epithelial to mesenchymal transition (EMT), which is characterized by a down-regulation of epithelial markers (cytokeratin, for example), a loss of cell polarity (E-cadherin, occludin), and the upregulation of mesenchymal markers (vimentin, N-cadherin, fibronectin) (34, 35). Furthermore, these VM cells have an endothelial phenotype. VM has been associated with unfavorable outcome in patients with malignant tumors (36) and has an important participation in tumor invasion and metastasis (37).
Cell-lined vasculature compatible with VM has been observed in ~30–37% of ovarian cancers (38, 39). The presence of such cell-lined vasculature was associated with a higher histological grade and more aggressive tumors. An increased number of VM channels were found in poorly differentiated ovarian cancer cells (40). The presence of VM, combined with the expression of CD133, was positively associated with poor prognosis in patients with ovarian cancer (41). In a preclinical model of ovarian cancer, an accelerated metastasis was observed together with hypoxia and VM after anti-angiogenic treatment with bevacizumab (an anti-VEGF-A monoclonal antibody) (9). All these findings highlight the importance of identifying the underlying mechanisms and the signaling molecules involved in VM to evaluate their prognostic or predictive value, as well as their use as potential targets for developing more effective therapies (42).
Structural and Functional Description of VM
In 1999, Maniotis et al. performed in vitro and in vivo assays in melanoma and found two VM types: a tubular type, and a patterned matrix type (13). The tubular type consists of hollow cords formed by tumor cells that give place to a tubular network. These tubular structures are also connected to other channels that contain red blood cells. Further studies showed that in some cases, a mixture of tumor cells and ECs form those tubular structures (43). The patterned matrix type consists of a network of loops formed by matrix layers that surround clusters of tumor cells. These layers are not uniformly spaced; therefore, the transport of fluid is not uniform around the cell cluster. However, this patterned matrix could provide a greater surface area for diffusion than that provided by a tubular structure (44). The reorganization of tumor cells into cords or clusters, as well as the formation of matrix layers involve mechanisms such as cell-cell adhesion, migration, and extracellular matrix remodeling, where several signaling molecules have been associated with VM.
VM structures have been identified in tissue samples as positive for periodic acid-Schiff (PAS) and negative for EC markers such as CD31 or CD34 (42). PAS+ regions are rich in components of the extracellular matrix, like laminin. Recently, it has been shown that PAS+ regions could also represent non-functional structures unrelated to VM in in vitro studies (45). Moreover, the different vascular structures aimed to conduct fluids within the tumor share several features. Thus, in order to identify the structures that truly correspond to VM as well as to distinguish them from similar structures from the other vascularization processes, it is necessary to assess their architectural and functional features, in addition to their composition. Recently, Valdivia et al. (46) described the architectural and functional features required for differentiating VM from other vascular structures in tumors (46). Whilst blood and lymph vessels are formed by a single line of ECs surrounded by a continuous and non-continuous basement membrane, respectively, VM structures are formed by cancer cells resting on an inner glycoprotein rich matrix (46). The authors propose that, in addition to the traditional architectural features to identify a VM structure (PAS+ and without EC markers), the presence of red blood cells within the lumen of the structure is an indicator of VM functionality (46).
Early studies in breast cancer and melanoma have shown that tubular and patterned matrix VM structures are capable of conducting plasma and red blood cells in vitro and in vivo (44). Maniotis et al. (47) showed that VM structures formed by aggressive melanoma cells in vitro conducted a tracer by direct microinjection and passive diffusion (47); moreover, the matrix pattern also contained red blood cells. Shirakawa et al. (48) used two breast cancer mice models to evaluate tumor blood flow with micro-magnetic resonance angiography imaging (48). They found that aggressive tumor cells formed VM structures in the center of the tumor, while non-aggressive cells showed necrotic cores. Angiogenic vessels were present in tumor periphery in both types of tumors and blood flow was higher in VM structures than in necrotic cores. Clarijs et al. (49) used a tracer to study perfusion in a melanoma mouse model (49). Tracer distribution suggested that blood vessels could be in contact with VM structures, allowing the perfusion of the latter, mediated by at least three mechanisms: the anastomosis of VM structures to blood vessels (50), an increased leakage from blood vessels (47, 51), and through anticoagulant control exerted by aggressive tumor cells (50).
Mechanisms and Signaling Molecules Involved in VM in Ovarian Cancer
Several mechanisms are involved in VM, including those related to the capacity of aggressive tumor cells to resemble features of the ECs such as cell adhesion (52), migration (53), extracellular matrix remodeling (54), perfusion (50), and maturation of blood vessels (55). Moreover, CSCs promote VM by deregulating pathways involved in embryonic development, such as the transforming growth factor β (TGF-β) (56–58), Wnt (59), Notch (60, 61), Nodal (62–64), and the Hippo pathways (65–67), among others. EMT also plays an important role in VM, and encompasses the pathways previously mentioned as well as transcription factors such as Twist1/2 (68), Snail/Slug (69), and ZEB1/2 (34). Moreover, signaling molecules related to hypoxia (70), inflammation (71–75), and metabolism (76–79) also have an impact on VM. The novel findings regarding these mechanisms and their signaling molecules in the regulation of VM in ovarian cancer are presented in this section and are summarized in Figure 1. Additional proteins and signaling pathways identified in other cancer types are shown in Table 1.
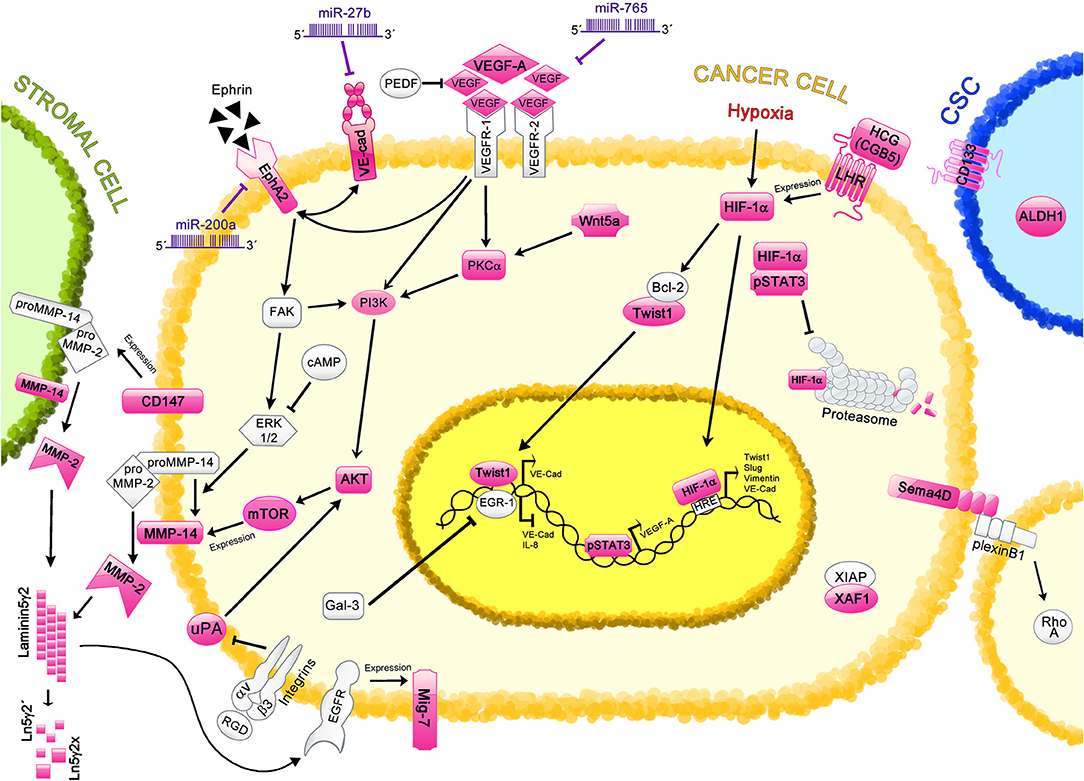
Figure 1. Regulation of Vasculogenic Mimicry by cell signaling molecules in ovarian cancer. Different proteins and signaling pathways involved in VM are shown; those characterized in ovarian cancer VM are highlighted in pink. Cancer cells are depicted in yellow; stromal cell, in green; and cancer stem cell (CSC), in blue. The VE-cadherin/EphA2/MMP-14/MMP-2/Ln5γ2 axis is the main mechanism involved in the induction of VM. This axis is regulated by miR-27b and miR-200a. Other proteins such as VEGF-A (regulated by miR-765 and pSTAT3), CD147, uPA, and Twist1 also regulate this axis through different pathways. Hypoxic-related protein HIF-1α induces the expression of Twist1, VE-Cadherin (VE-Cad), Slug and Vimentin, which are involved in VM induction; moreover, proteins such as pSTAT3, HCG, and LHR regulate the levels of HIF-1α. CSC markers, including ALDH and CD133 are found in ovarian tissues with VM structures. Different cell signaling pathways are also involved in VM, such as Wnt5a, and RTKs pathways, which strongly correlate with VM formation. Additional molecules that have been proposed in VM regulation in ovarian cancer are Sema4, XAF1, and Mig-7, however the precise mechanisms remain unclear.
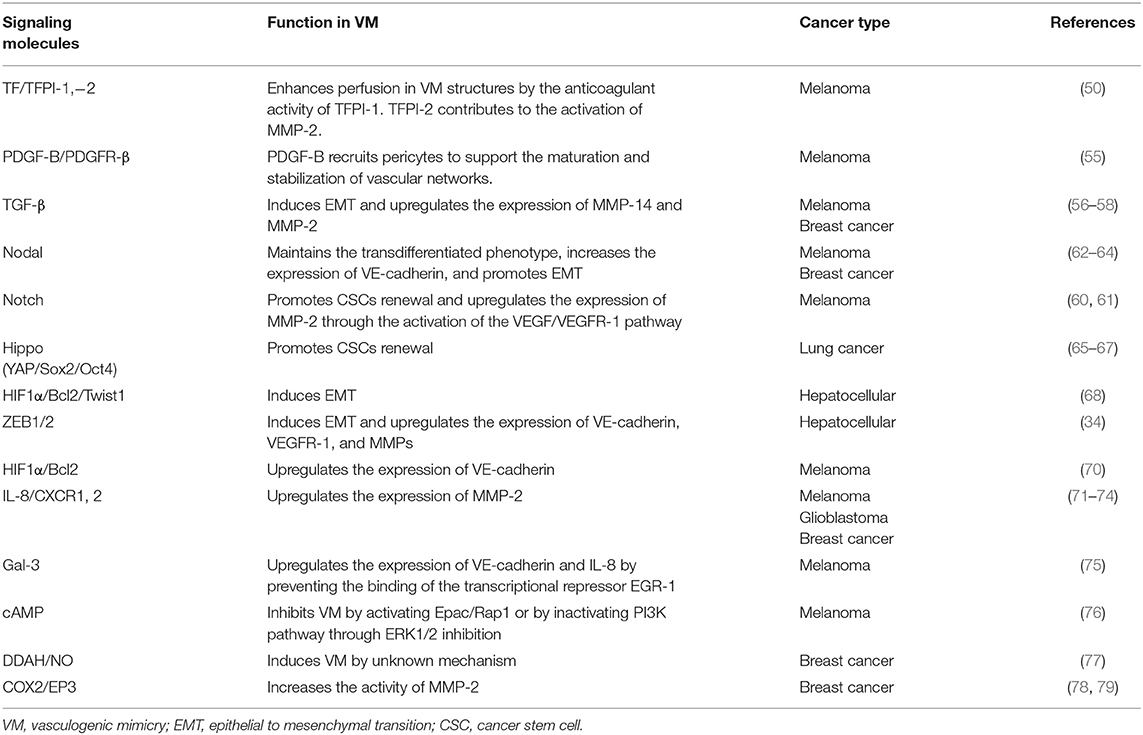
Table 1. Signaling molecules and mechanisms that regulate VM in several types of cancer.
Vascular endothelial (VE)-cadherin, one of the main participants in cell-cell adhesion in endothelial cells, is strongly associated with VM formation (80). This protein recruits the EC-related kinase Ephrin-A2 receptor (EphA2) to the cell membrane (52), increasing the phosphorylation of the focal adhesion kinase (FAK). Consequently, the activation of extracellular regulatory kinases 1 and 2 (ERK1/2) signaling pathway is promoted, allowing the activation of MMP-14 (81). Then, MMP-14 converts proMMP-2 into active MMP-2. These MMPs degrade extracellular matrix components and facilitate invasion, metastasis, and VM (82). Particularly, MMP-2 and MMP-14 induce the Laminin5γ2 (Ln5γ2) cleavage (53, 83). Although the precise mechanism has not been clearly described, it is known that MMP-2 cleavages Ln5γ2 into two pro-metastatic fragments (Ln5γ2′ and Ln5γ2x) (53). Together, these results indicate that the VE-cadherin/EphA2/MMP-2/Ln5γ2 axis is the main regulator of VM.
Interestingly, high expression of VE-cadherin and EphA2 has been found in clinical samples from ovarian cancer patients that exhibit a highly invasive phenotype (84, 85). Additionally, other studies demonstrated that MMP-2 and MMP-14 are also overexpressed in ovarian cancer samples, which is associated with poor clinical outcome (38, 86). It is worth to mention that those findings strongly correlated with the presence of VM structures, suggesting that these molecules are important players in this process.
The Phosphatidylinositol 3-kinase (PI3K) cell signaling pathway regulates MMPs expression in VM (87). This pathway is activated through FAK phosphorylation (88), impacting in the expression of MMP-14. Moreover, the PI3K pathway is frequently activated in ovarian cancer, probably impacting VM (89).
Another regulator of VM is the urokinase plasminogen activator (uPA), which is required to induce the degradation of the extracellular matrix, impacting in tumor angiogenesis. The overexpression of uPA positively correlates with VM formation in ovarian cancer tissues (54). In addition, it was demonstrated that in SKOV-3 and OVCAR-3 ovarian cancer cells, the ablation of uPA expression results in a decrement of complete VM structures formation and such mechanism involves the participation of AKT/mTOR/MMP-2/Laminin5γ2 signal pathways (54).
VEGF-A also upregulates the expression MMPs. It has been shown that in melanoma, VEGF-A induces VM formation by activating the PI3K/protein kinase C α (PKCα) pathway via VEGFR-1 signaling (90). However, in glioblastoma, VM is induced by the VEGFR-2 signaling (91). In an in vitro model of ovarian cancer using SKOV-3 and OVCAR-3 cells, VEGF-A promoted migration, invasion, and VM by up-regulating MMPs via EphA2 (92). This suggests that VEGF-A interacts with the VE-cadherin/EphA2/MMP-2/Ln5γ2 axis in the regulation of VM in ovarian cancer.
The plasma membrane glycoprotein CD147 plays an important role during tumor progression, invasion and metastasis, regulating metalloproteinases expression in peritumoral stromal cells. Invasion capability was evaluated in two different cell lines derived from ovarian cancer with different invasion activity: CABA I and SKOV3 (93). A correlation of CD147 expression with tumor invasiveness, protease activity (MMP-2 and MMP-9), and vascular channels formation was observed. Interestingly, when high invasive cell line was treated with small interfering RNA against CD147, a suppression of non-EC-lined channels was observed. In addition, when CD147 was overexpressed in a low invasive cell line, those cells exhibited an increase of tumor invasion and vascular channel formation. These data suggest that CD147 plays an important role in VM induction in ovarian tumors and CD147 could be an attractive target for therapeutic intervention (93).
Furthermore, Ln5γ2 activates the endothelial growth factor receptor (EGFR) which promotes the expression of the migration-inducing protein 7 (Mig-7), stimulating invasion and VM (94). A study carried out in ovarian cancer samples revealed an association of VM with VE-cadherin and Mig-7 expression (84). It was observed that ovarian tumors without VM frequently expressed low levels of VE-cadherin compared to those with VM. Meanwhile, Mig-7 expression was increased in tumor samples compared to normal tissues, positively correlating with VM and VE-cadherin expression (84).
Some elements involved in apoptosis have been associated with the formation of VM structures, such as the pro-apoptotic XIAP-associated factor 1 (XAF1). Recently, in vivo xenograft models of ovarian cancer have shown that the overexpression of XAF1 decreases the number of VM structures (39). Moreover, in vitro assays with SKOV3 cells revealed that proliferation, migration and invasion were inhibited, and the levels of VEGF were reduced when XAF1 was exogenously overexpressed (39). Therefore, XAF1 is a potent negative regulator of VM in ovarian cancer.
It has been shown that VEGF-A regulates the expression of the axon guidance factor semaphorin 4D (Sema4D) (95), which has been identified as a promotor of VM in non-small cell lung cancer (96), where the recognition of Sema4D by the plexin B1 receptor activates the small GTPase RhoA, which is implicated cell motility. However, when plexin B1 was inhibited, a disruption of the RhoA/ROCK signaling occurred, suppressing VM formation. Additionally, the presence of VM in clinical specimens correlated with increased levels of Sema4D (96). In an ovarian cancer cell line (A2780), soluble Sema4D promoted angiogenesis and VM via plexin B1 (95); moreover, in clinical samples from patients, a high expression of Sema4D had a positive correlation with the malignant degree of epithelial ovarian cancer. Interestingly, it was observed that VEGFR-2, plexin-B1, and Sema4D control the expression of CD31, MMP-2, and VE-cadherin in ovarian cancer cells, which are the markers and initiators of angiogenesis and VM (95).
CSCs are present in ovarian cancer and are positive for CD133, a unique surface marker of CSCs (97). It is known that CD133+ cells promote VM in several cancer types (41, 91, 98–101). The combined expression of CD133 and VM in samples from patients was associated with high-grade ovarian carcinoma, late-stage disease, non-response to chemotherapy and shorter overall survival (41). The trans-differentiation of CD133+ CSCs into ECs may induce VM formation and the expression of EC markers such as VE-cadherin (101) and VEGFR-2 (91). Moreover, it has been shown that in hypoxic environment the subpopulation of CD133+ CSCs is augmented when Twist1 was overexpressed (100). This finding shows that hypoxia may exert an effect on CSCs that probably leads to VM formation.
CSCs can also exhibit a high activity of aldehyde dehydrogenase-1 (ALDH1) (97). The expression of ALDH1 has been evaluated in different types of tumor, including breast cancer, colorectal cancer, and ovarian cancer and strongly correlates with VM, determining an unfavorable clinical outcome (102–104). Although the precise mechanism has not been described, it is known that ALDH1 and VM increase in response to hypoxia (105).
Hypoxia regulates several pathways in cancer, such as angiogenesis, and it has been related to VM in melanoma, glioblastoma, ovarian cancer, and hepatocellular carcinoma (68–70, 106). Hypoxia induces VM formation by up-regulating VE-cadherin expression. The main effectors of this pathways, HIF-1α and HIF-2α, positively regulated VE-cadherin expression; this effect is through the binding of HIF to hypoxia response elements (HRE) located in VE-cadherin promoter in glioblastoma cells (106). Interestingly, it was observed that EMT is promoted in a hypoxic environment and as a result, VM was induced in SKOV3 and OVCAR3 cells (69). In vitro assays showed that hypoxia leads to increased invasion, migration and an enhancement of MMP-2 activity. Therefore, EMT induction as a response to hypoxia is a master regulator of VM in ovarian cancer cells. Moreover, this study demonstrated that in ovarian cancer samples, the levels of HIF-1α were strongly associated with VM formation and the expression of Twist1, Slug, and Vimentin.
Another important regulator of VM under hypoxic conditions are the signal transducer and activator of transcription 3 (STAT3) and the phospho-STAT3 (p-STAT3). It has been suggested that p-STAT3 promote VM, this is due to the binding of pSTAT3 to HIF-1α, which in turn delays its degradation (107, 108). In gastric adenocarcinoma, VM was associated with an increased expression of HIF-1α, STAT3, and p-STAT3 (109). Moreover, STAT3 acts as a transcription factor in VEGF-A transcription (110). Interestingly, in SKOV3 cells p-STAT3 was found in the nucleus, suggesting that was transcriptionally active (111). In addition, when STAT3 was inhibited, the formation of VM structures was completely avoided, suggesting that p-STAT3 is an important regulator of VM in ovarian cancer cells.
The Wnt family members regulate EC differentiation and vascular development (112) and has been associated with VM. In glioma and colon cancer, the canonical Wnt/β-catenin pathway induced VM by increasing the expression of VEGFR-2 and VE-cadherin (59, 113). Interestingly, in ovarian cancer, the non-canonical Wnt signaling is implicated in VM formation. It was found that Wnt5a is overexpressed in tumor samples and is associated with VM (114). Moreover, in vitro analysis revealed that Wnt5a overexpression is linked to PKC pathway activation. Furthermore, it was shown that Wnt5a overexpression induced EMT, increased invasion and motility of SKOV3 cells (114).
An important proangiogenic factor in ovary is the human gonadotropin (HCG). The fifth subunit of β-HCG, CGB5, was shown to promote VM formation in vitro in OVCAR3 cells (115). Additionally, overexpression of CGB5 induced the growth of ovarian cancer cells in a xenograft murine model, as well as VM (116). It was also shown that the activation of luteinizing hormone receptor (LHR), which is the HCG receptor, is required for the promotion of VM formation by CGB5. In another study, it was found that ovarian cancer cells exogenously expressing HCG induced an overexpression of HIF-1α. Importantly, vascular markers such as CD31 and VEGF were also upregulated in those cells (117). Therefore, the HCG/LHR axis induces VM by HIF-1α regulation in ovarian cancer.
Micro-RNAs as Regulators of VM in Ovarian Cancer
Micro-RNAs (miRNAs) are single stranded and non-coding RNA molecules of 19-25 nucleotides in length that have a post-transcriptional regulatory function (118). Different studies have demonstrated that miRNAs are involved in several physiological processes such as cell proliferation, invasion, migration, differentiation, as well as pathological processes including angiogenesis and VM (119–122). The dysregulation in the expression of these RNA molecules is often observed in numerous types of cancer. Diverse studies demonstrate that miRNAs post-transcriptionally regulate different signaling molecules involved in VM process (123–136); examples of these miRNAs are enlisted in Table 2.
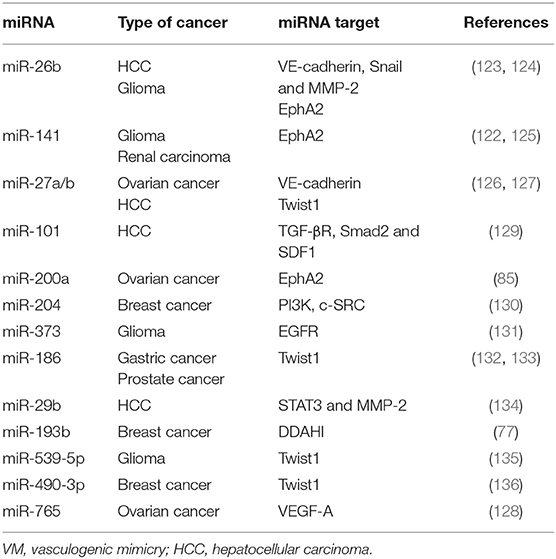
Table 2. VM related miRNAs in different types of cancer.
A well-described miRNA family is miR-26, which includes miR-26a and miR-26b. Those are commonly downregulated in several types of cancer such as glioma, HCC, and gastric cancer (124, 137, 138). For instance, in gastric cancer miR-26a and−26b suppress angiogenesis by targeting hormone growth factor (HGF) mRNA and consequently affecting HGF/VEGF signaling (138). Moreover, in HCC miR-26b has been identified as tumor suppressor since its down-regulation promotes VM and angiogenesis (123).
Similarly, another cluster of miRNAs belonging to the miR-200 family (miR-141, miR-200a, miR-200b, miR-200c, and miR-429) has been widely studied in several types of cancers (125, 139–142). It has been shown that miR-141 overexpression inhibits VM formation through directly targeting EphA2 transcript, decreasing EphA2 protein levels in glioma and renal carcinomas (122, 125).
Hitherto, three miRNAs (miR-200a, miR-27b, and miR-765) have been described as VM regulators in ovarian cancer through directly targeting 3′UTRs of VM-related transcripts (85, 126, 128) (Table 3). The miR-200a was the first microRNA found in ovarian cancer capable of regulating VM (85). Tumors with low miR-200a expression correlate with the presence of VM structures and poor overall survival. An inverse correlation between mRNA and protein EphA2 levels and miR-200a expression was observed in ovarian cancer samples, suggesting a direct regulation among them. In silico assays revealed a miR-200a binding site at EphA2 3′UTR; this observation was confirmed in SKOV3 ovarian cancer cells, where a direct binding of miR-200a to EphA2 3′UTR was observed through luciferase assays. Consequently, the levels of EphA2 protein and mRNA decreased in this model. In agreement, it was shown that the EphA2 overexpression restores VM in miR-200a expressing cells, indicating that miR-200a inhibits VM by mainly targeting EphA2 (85).
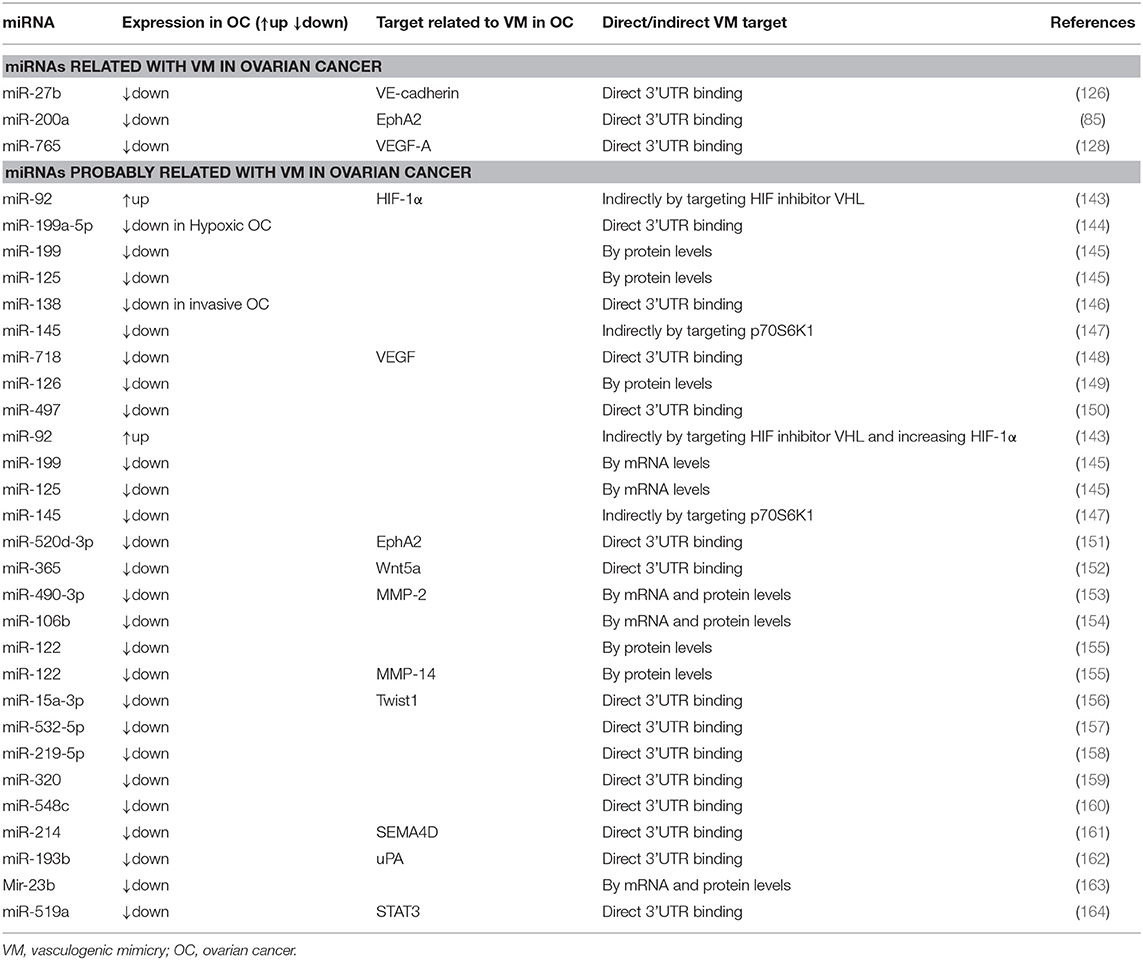
Table 3. VM related miRNAs in ovarian cancer.
Previously, it has been described that VE-cadherin expression is related to VM formation in different types of cancer. A bioinformatic study identified miR-27b as putative regulator of VE-cadherin by the detection of a binding site at VE-cadherin 3′UTR. Concordantly with this result, luciferase assays demonstrated that miR-27b binds to VE-cadherin mRNA 3′UTR in ovarian cancer cells. Furthermore, expression levels of VE-cadherin mRNA and protein in different ovarian cancer cell lines negatively correlate with miR-27b expression. Low metastatic cell lines OVCAR3 and SKOV3 express high amounts of miR-27b and low VE-cadherin mRNA, compared to metastatic cells ES2 and Hey1B that exhibit low amounts of miR-27b and high VE-cadherin mRNA. Overexpression of miR-27b on high VE-cadherin expressing cells decreases VE-cadherin mRNA and protein levels. When miR-27b is overexpressed in metastatic ovarian cancer cell lines (Hey1B and ES2), the migration, invasion, and VM are decreased in in vivo models (126).
A recent study aimed to determine the set of miRNAs regulated in an early stage before complete VM establishment under hypoxia conditions. It was shown that SKOV3 ovarian cancer cells grown under hypoxia conditions form a higher number of 3D capillary-like structures than those cells grown under normoxia conditions (128). A set of miRNAs involved in the regulation of tumorigenesis-related pathways, as well as several genes involved in VM and angiogenesis was found. Among them, miR-765 was highly downregulated under hypoxia (128). Moreover, its restoration promotes a dramatic inhibition of 3D capillary-like structures and down-regulates VEGF-A expression. Importantly, it was demonstrated that VEGF-A mRNA is a direct target of miR-765, since it binds to VEGF-A 3'UTR. Additionally, low levels of miR-765 and high levels of VEGF-A were associated with low overall survival from a cohort of 1,485 ovarian cancer patients (128).
Although only three miRNAs have been directly associated with VM in ovarian cancer, several signaling pathways, and proteins controlling this mechanism are regulated by miRNAs (143–164); therefore, these non-coding transcripts could have a potential role on VM regulation. Table 3 shows the common VM targets in ovarian cancer that are regulated by miRNAs.
Clinical Implications of the Signaling Molecules of VM in Ovarian Cancer
Anti-angiogenic therapies have shown limited effects against cancer progression, due to alternative vascularization processes, such as VM, triggered by aggressive tumor cells (10). The knowledge of the mechanisms and signaling molecules involved in VM may lead to the development of novel anti-vascularization therapies that overcome the limitations found in conventional therapies. Therefore, it is necessary to explore the possible therapeutical strategies that could improve the clinical outcome of ovarian cancer patients.
Therapies targeting VM have not been developed in ovarian cancer so far. However, some inhibitory molecules of VM elements have been studied and have shown promising anti-VM effects (165–174). These inhibitor molecules are summarized in Table 4.
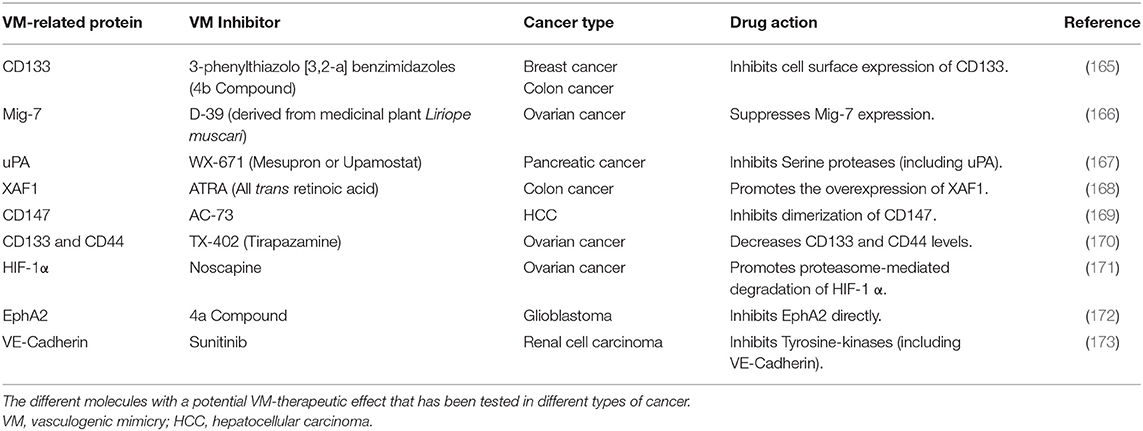
Table 4. Inhibitor molecules that target VM-related proteins.
Studies using pancreatic cancer cells showed that Ginsenoside Rg3, a tetracyclic triterpenoid saponin, reduces VM in xenograft mice models. Moreover, the expression of VE-cadherin, EphA2, MMP-2, and MMP-9 was also down-regulated after the treatment (174). Ginsenoside Rg3 has been proved in ovarian cancer derived cells restraining HIF-1α expression by activating the ubiquitin-proteasome pathway. This effect efficiently blocked migration and EMT in in vitro and in vivo ovarian cancer models, promising a novel anti-VM therapeutic agent (175, 176).
It has been shown that PARP inhibition sensitizes for chemo and radiotherapy in different types of tumors. In melanoma cells that were treated with PARP inhibitors (PJ-34, Isoquinolinone, or Olaparib) a reduction of pro-metastatic and VM markers was observed (177). PARP I inhibitors, such as Olaparib and Rupaparib, have been approved for the treatment of recurrent BRCA-associated ovarian cancer by the Food and Drug Administration (FDA); while Niraparib is used as maintenance therapy following chemotherapy for recurrent ovarian cancer (178). Nevertheless, to date there is no information about their effect on VM in ovarian cancer.
Thalidomide is an immunomodulatory agent with strong anti-angiogenic properties and has been proved in ovarian cancer, glioblastoma, hepatocellular carcinoma, and multiple myeloma in diverse clinical trials. Induction therapy with thalidomide significantly improved the overall response rate, progression free survival and overall survival (179). Previously, it has been shown that thalidomide suppresses tumor growth and angiogenesis in murine models (180). Interestingly, in a xenograft mouse model of melanoma, it was observed that mice treated with Thalidomide induced necrosis in melanoma cells. In addition, VM and tumor growth were significantly reduced compared to non-treated specimens. This effect could be related to the down-regulation of NF-kappaB signaling pathway (181). However, further studies are required to elucidate this statement.
A monoclonal antibody has been developed to target VM, unfortunately it has not been introduced for ovarian cancer treatment. This antibody targets the outer-membrane immunoglobulin-like domains of VE-cadherin, blocking receptor function. In lung cancer cells, it was observed that this antibody functions as an anti-VM agent for cancer treatment, since it inhibited the activation of the VE-cadherin-related pathway in VM (182). Due to the advantages that monoclonal antibody therapies imply, its application in ovarian cancer as an anti-VM agent is promising.
Other molecules implicated in VM in ovarian cancer, such as miR-200a, miR-27b, and miR-765 represent potential candidates for anti-tumoral therapies (85, 126, 128). Importantly, the current strategies are focused in the reduction of cancer through restoring the expression of down-regulated miRNAs, also known as miRNA replacement therapy. There are several ways to harness miRNAs in cancer cells for therapeutic purposes, including introduction of synthetic miRNA mimics, miRNA expressing plasmids, and small molecules that epigenetically alter endogenous expression of miRNAs (183). Such anti-VM strategies could represent an opportunity to venture into the study of new molecules for therapeutic purposes in ovarian cancer. Further studies will be required to prove the effectiveness of such molecules for treatment purposes.
Concluding Remarks
Ovarian cancer is a common gynecological cancer and it is usually diagnosed in advanced stages where therapeutic success is limited. This type of tumors exhibit an aggressive phenotype characterized by a high rate of metastasis, invasion, and poor treatment response. These features are highly associated with the development of neovasculature formed by both endothelial and tumor cells. Particularly, MV is a process that may be influencing ovarian cancer poor prognosis and limited efficacy of anti-angiogenic strategies. Nevertheless, the mechanisms underlying VM formation in ovarian cancer remains unclear and deserves further studies. Recently, molecules that regulate cellular adhesion, hypoxia and EMT have been identified as key regulators of VM. Additionally, it has been shown an important post-transcriptional regulation mediated by microRNAs, that impact on the expression of VM-related proteins such as VE-cadherin, EphA2, and VEGF. Furthermore, this information has allowed the development of strategies with therapeutic potential directed against VM formation. However, subsequent studies will be necessary to elucidate the mechanisms that allow the development of conventional anti-angiogenic therapies combined with the novel anti-VM targets that improve the clinical outcomes of ovarian cancer patients.
Author Contributions
LA-D, LO-N, JM-B, AC-P, JM-M, IM-R, and ML performed the bibliographic review, wrote, and critically revised the manuscript. ML conceived and directed the manuscript.
Funding
This work was partially supported by CONACyT grant CB-251497, PAPIIT-UNAM IN103219, and Instituto Nacional de Cancerología Ref. 018/051/1B1/CE1/1294/18.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
LA-D and LO-N are students from the following programs: Doctorado en Ciencias Biomédicas and Doctorado en Ciencias Bioquímicas, respectively, at the Universidad Nacional Autónoma de México and are recipients of scholarships from CONACyT, México (221487 and 289892, respectively).
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. (2018) 68:394–424. doi: 10.3322/caac.21492
2. Oswald AJ, Gourley C. Low-grade epithelial ovarian cancer: a number of distinct clinical entities? Curr Opin Oncol. (2015) 27:412–9. doi: 10.1097/CCO.0000000000000216
3. Ricci F, Guffanti F, Damia G, Broggini M. Combination of paclitaxel, bevacizumab and MEK162 in second line treatment in platinum-relapsing patient derived ovarian cancer xenografts. Mol Cancer. (2017) 16:97. doi: 10.1186/s12943-017-0662-3
4. Fraser HM, Duncan WC. Vascular morphogenesis in the primate ovary. Angiogenesis. (2005) 8:101–16. doi: 10.1007/s10456-005-9004-y
5. Kristensen GB, Tropé C. Epithelial ovarian carcinoma. Lancet. (1997) 349:113–7. doi: 10.1016/S0140-6736(96)06071-0
7. Tang H-S, Feng Y-J, Yao L-Q. Angiogenesis, vasculogenesis, and vasculogenic mimicry in ovarian cancer. Int J Gynecol Cancer. (2009) 19:605–10. doi: 10.1111/IGC.0b013e3181a389e6
8. Choi H-J, Armaiz Pena GN, Pradeep S, Cho MS, Coleman RL, Sood AK. Anti-vascular therapies in ovarian cancer: moving beyond anti-VEGF approaches. Cancer Metastasis Rev. (2015) 34:19–40. doi: 10.1007/s10555-014-9538-9
9. Xu Y, Li Q, Li X-Y, Yang Q-Y, Xu W-W, Liu G-L. Short-term anti-vascular endothelial growth factor treatment elicits vasculogenic mimicry formation of tumors to accelerate metastasis. J Exp Clin Cancer Res. (2012) 31:16. doi: 10.1186/1756-9966-31-16
10. van Beijnum JR, Nowak-Sliwinska P, Huijbers EJM, Thijssen VL, Griffioen AW. The great escape; the hallmarks of resistance to antiangiogenic therapy. Pharmacol Rev. (2015) 67:441–61. doi: 10.1124/pr.114.010215
11. Dome B, Timar J, Dobos J, Meszaros L, Raso E, Paku S, et al. Identification and clinical significance of circulating endothelial progenitor cells in human non-small cell lung cancer. Cancer Res. (2006) 66:7341–7. doi: 10.1158/0008-5472.CAN-05-4654
12. Donnem T, Reynolds AR, Kuczynski EA, Gatter K, Vermeulen PB, Kerbel RS, et al. Non-angiogenic tumours and their influence on cancer biology. Nat Rev Cancer. (2018) 18:323–36. doi: 10.1038/nrc.2018.14
13. Maniotis AJ, Folberg R, Hess A, Seftor EA, Gardner LM, Pe'er J, et al. Vascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicry. Am J Pathol. (1999) 155:739–52. doi: 10.1016/S0002-9440(10)65173-5
14. Tung JJ, Tattersall IW, Kitajewski J. Tips, stalks, tubes: notch-mediated cell fate determination and mechanisms of tubulogenesis during angiogenesis. Cold Spring Harb Perspect Med. (2012) 2:a006601. doi: 10.1101/cshperspect.a006601
15. Carmeliet P, Jain RK. Molecular mechanisms and clinical applications of angiogenesis. Nature. (2011) 473:298–307. doi: 10.1038/nature10144
16. Leung DW, Cachianes G, Kuang WJ, Goeddel DV, Ferrara N. Vascular endothelial growth factor is a secreted angiogenic mitogen. Science. (1989) 246:1306–9. doi: 10.1126/science.2479986
17. Potente M, Gerhardt H, Carmeliet P. Basic and therapeutic aspects of angiogenesis. Cell. (2011) 146:873–87. doi: 10.1016/j.cell.2011.08.039
18. Winiarski BK, Wolanska KI, Rai S, Ahmed T, Acheson N, Gutowski NJ, et al. Epithelial ovarian cancer-induced angiogenic phenotype of human omental microvascular endothelial cells may occur independently of VEGF signaling. Transl Oncol. (2013) 6:703–14. doi: 10.1593/tlo.13529
19. Ghoneum A, Afify H, Salih Z, Kelly M, Said N. Role of tumor microenvironment in the pathobiology of ovarian cancer: insights and therapeutic opportunities. Cancer Med. (2018) 7:5047–56. doi: 10.1002/cam4.1741
20. Sallinen H, Heikura T, Koponen J, Kosma V-M, Heinonen S, Ylä-Herttuala S, et al. Serum angiopoietin-2 and soluble VEGFR-2 levels predict malignancy of ovarian neoplasm and poor prognosis in epithelial ovarian cancer. BMC Cancer. (2014) 14:696. doi: 10.1186/1471-2407-14-696
21. Sönmezer M, Güngör M, Ensari A, Ortaç F. Prognostic significance of tumor angiogenesis in epithelial ovarian cancer: in association with transforming growth factor beta and vascular endothelial growth factor. Int J Gynecol Cancer. (2004) 14:82–8. doi: 10.1111/j.1048-891x.2004.14202.x
22. Shmilovich H, Ben-Shoshan J, Tal R, Afek A, Barshack I, Maysel-Auslander S, et al. B-type natriuretic peptide enhances vasculogenesis by promoting number and functional properties of early endothelial progenitor cells. Tissue Eng Part A. (2009) 15:2741–9. doi: 10.1089/ten.tea.2008.0414
23. Carbone C, Moccia T, Zhu C, Paradiso G, Budillon A, Chiao PJ, et al. Anti-VEGF treatment-resistant pancreatic cancers secrete proinflammatory factors that contribute to malignant progression by inducing an EMT cell phenotype. Clin Cancer Res. (2011) 17:5822–32. doi: 10.1158/1078-0432.CCR-11-1185
24. Nolan DJ, Ciarrocchi A, Mellick AS, Jaggi JS, Bambino K, Gupta S, et al. Bone marrow-derived endothelial progenitor cells are a major determinant of nascent tumor neovascularization. Genes Dev. (2007) 21:1546–58. doi: 10.1101/gad.436307
25. Xu F, Feng Y. Peripheral blood CD34+-positive mononuclear cells participate in neovasculogenesis of human ovarian epithelial carcinoma. Zhonghua Chan Ke Za Zhi. (2004) 39:620–3. doi: 10.3760/j.issn:0529-567x.2004.09.013
26. Brown JM. Vasculogenesis: a crucial player in the resistance of solid tumours to radiotherapy. Br J Radiol. (2014) 87:20130686. doi: 10.1259/bjr.20130686
27. Liu T, Du X, Sheng X. Genetic alterations following ionizing radiation in human ovarian cancer-derived endothelial cells. Mol Med Rep. (2014) 9:2257–64. doi: 10.3892/mmr.2014.2096
28. Coukos G, Benencia F, Buckanovich RJ, Conejo-Garcia JR. The role of dendritic cell precursors in tumour vasculogenesis. Br J Cancer. (2005) 92:1182–7. doi: 10.1038/sj.bjc.6602476
29. Yokoyama Y, Sedgewick G, Ramakrishnan S. Endostatin binding to ovarian cancer cells inhibits peritoneal attachment and dissemination. Cancer Res. (2007) 67:10813–22. doi: 10.1158/0008-5472.CAN-07-0172
30. Holash J, Maisonpierre PC, Compton D, Boland P, Alexander CR, Zagzag D, et al. Vessel cooption, regression, and growth in tumors mediated by angiopoietins and VEGF. Science. (1999) 284:1994–8. doi: 10.1126/science.284.5422.1994
31. Naumov GN, Akslen LA, Folkman J. Role of angiogenesis in human tumor dormancy: animal models of the angiogenic switch. Cell Cycle. (2006) 5:1779–87. doi: 10.4161/cc.5.16.3018
32. Sood AK, Seftor EA, Fletcher MS, Gardner LM, Heidger PM, Buller RE, et al. Molecular determinants of ovarian cancer plasticity. Am J Pathol. (2001) 158:1279–88. doi: 10.1016/S0002-9440(10)64079-5
33. Ricci-Vitiani L, Pallini R, Biffoni M, Todaro M, Invernici G, Cenci T, et al. Tumour vascularization via endothelial differentiation of glioblastoma stem-like cells. Nature. (2010) 468:824–8. doi: 10.1038/nature09557
34. Yang Z, Sun B, Li Y, Zhao XX, Zhao XX, Gu Q, et al. ZEB2 promotes vasculogenic mimicry by TGF-β1 induced epithelial-to-mesenchymal transition in hepatocellular carcinoma. Exp Mol Pathol. (2015) 98:352–9. doi: 10.1016/j.yexmp.2015.03.030
35. Fan Y-L, Zheng M, Tang Y-L, Liang X-H. A new perspective of vasculogenic mimicry: EMT and cancer stem cells (Review). Oncol Lett. (2013) 6:1174–80. doi: 10.3892/ol.2013.1555
36. Cao Z, Bao M, Miele L, Sarkar FH, Wang Z, Zhou Q. Tumour vasculogenic mimicry is associated with poor prognosis of human cancer patients: a systemic review and meta-analysis. Eur J Cancer. (2013) 49:3914–23. doi: 10.1016/j.ejca.2013.07.148
37. Seftor REB, Hess AR, Seftor EA, Kirschmann DA, Hardy KM, Margaryan NV, et al. Tumor cell vasculogenic mimicry: from controversy to therapeutic promise. Am J Pathol. (2012) 181:1115–25. doi: 10.1016/j.ajpath.2012.07.013
38. Sood AK, Fletcher MS, Coffin JE, Yang M, Seftor EA, Gruman LM, et al. Functional role of matrix metalloproteinases in ovarian tumor cell plasticity. Am J Obstet Gynecol. (2004) 190:899–909. doi: 10.1016/j.ajog.2004.02.011
39. Wang Y, Liu P, Wang X, Mao H. Role of X-linked inhibitor of apoptosis-associated factor-1 in vasculogenic mimicry in ovarian cancer. Mol Med Rep. (2017) 16:325–30. doi: 10.3892/mmr.2017.6597
40. Gao Y, Zhao X, Gu Q, Wang J, Zhang S, Zhang D, et al. Correlation of vasculogenic mimicry with clinicopathologic features and prognosis of ovarian carcinoma. Zhonghua Bing Li Xue Za Zhi. (2009) 38:585–9. doi: 10.3760/cma.j.issn.0529-5807.2009.09.003
41. Liang J, Yang B, Cao Q, Wu X. Association of vasculogenic mimicry formation and CD133 expression with poor prognosis in ovarian cancer. Gynecol Obstet Invest. (2016) 81:529–36. doi: 10.1159/000445747
42. Ge H, Luo H. Overview of advances in vasculogenic mimicry - a potential target for tumor therapy. Cancer Manag Res. (2018) 10:2429–37. doi: 10.2147/CMAR.S164675
43. El Hallani S, Boisselier B, Peglion F, Rousseau A, Colin C, Idbaih A, et al. A new alternative mechanism in glioblastoma vascularization: tubular vasculogenic mimicry. Brain. (2010) 133:973–82. doi: 10.1093/brain/awq044
44. Folberg R, Maniotis AJ. Vasculogenic mimicry. APMIS. (2004) 112:508–25. doi: 10.1111/j.1600-0463.2004.apm11207-0810.x
45. Racordon D, Valdivia A, Mingo G, Erices R, Aravena R, Santoro F, et al. Structural and functional identification of vasculogenic mimicry in vitro. Sci Rep. (2017) 7:6985. doi: 10.1038/s41598-017-07622-w
46. Valdivia A, Mingo G, Aldana V, Pinto MP, Ramirez M, Retamal C, et al. Fact or fiction, it is time for a verdict on vasculogenic mimicry? Front Oncol. (2019) 9:680. doi: 10.3389/fonc.2019.00680
47. Maniotis AJ, Chen X, Garcia C, DeChristopher PJ, Wu D, Pe'er J, et al. Control of melanoma morphogenesis, endothelial survival, and perfusion by extracellular matrix. Lab Invest. (2002) 82:1031–43. doi: 10.1097/01.LAB.0000024362.12721.67
48. Shirakawa K, Kobayashi H, Heike Y, Kawamoto S, Brechbiel MW, Kasumi F, et al. Hemodynamics in vasculogenic mimicry and angiogenesis of inflammatory breast cancer xenograft. Cancer Res. (2002) 62:560–6.
49. Clarijs R, Otte-Höller I, Ruiter DJ, de Waal RMW. Presence of a fluid-conducting meshwork in xenografted cutaneous and primary human uveal melanoma. Invest Ophthalmol Vis Sci. (2002) 43:912–8.
50. Ruf W, Seftor EA, Petrovan RJ, Weiss RM, Gruman LM, Margaryan NV, et al. Differential role of tissue factor pathway inhibitors 1 and 2 in melanoma vasculogenic mimicry. Cancer Res. (2003) 63:5381–9.
51. Frenkel S, Barzel I, Levy J, Lin AY, Bartsch D-U, Majumdar D, et al. Demonstrating circulation in vasculogenic mimicry patterns of uveal melanoma by confocal indocyanine green angiography. Eye. (2008) 22:948–52. doi: 10.1038/sj.eye.6702783
52. Hess AR, Seftor EA, Gruman LM, Kinch MS, Seftor REB, Hendrix MJC. VE-cadherin regulates EphA2 in aggressive melanoma cells through a novel signaling pathway: implications for vasculogenic mimicry. Cancer Biol Ther. (2006) 5:228–33. doi: 10.4161/cbt.5.2.2510
53. Giannelli G, Falk-Marzillier J, Schiraldi O, Stetler-Stevenson WG, Quaranta V. Induction of cell migration by matrix metalloprotease-2 cleavage of laminin-5. Science. (1997) 277:225–8. doi: 10.1126/science.277.5323.225
54. Tang J, Wang JJ, Fan L, Li X, Liu N, Luo W, et al. cRGD inhibits vasculogenic mimicry formation by down-regulating uPA expression and reducing EMT in ovarian cancer. Oncotarget. (2016) 7:24050–62. doi: 10.18632/oncotarget.8079
55. Thijssen VL, Paulis YW, Nowak-Sliwinska P, Deumelandt KL, Hosaka K, Soetekouw PM, et al. Targeting PDGF-mediated recruitment of pericytes blocks vascular mimicry and tumor growth. J Pathol. (2018) 246:447–58. doi: 10.1002/path.5152
56. Ling G, Wang S, Song Z, Sun X, Liu Y, Jiang X, et al. Transforming growth factor-β is required for vasculogenic mimicry formation in glioma cell line U251MG. Cancer Biol Ther. (2011) 12:978–88. doi: 10.4161/cbt.12.11.18155
57. Kuo Y-C, Su C-H, Liu C-Y, Chen T-H, Chen C-P, Wang H-S. Transforming growth factor-beta induces CD44 cleavage that promotes migration of MDA-MB-435s cells through the up-regulation of membrane type 1-matrix metalloproteinase. Int J Cancer. (2009) 124:2568–76. doi: 10.1002/ijc.24263
58. Kim E-S, Sohn Y-W, Moon A. TGF-beta-induced transcriptional activation of MMP-2 is mediated by activating transcription factor (ATF)2 in human breast epithelial cells. Cancer Lett. (2007) 252:147–56. doi: 10.1016/j.canlet.2006.12.016
59. Wang H, Fu J, Xu D, Xu W, Wang S, Zhang L, et al. Downregulation of Pygopus 2 inhibits vascular mimicry in glioma U251 cells by suppressing the canonical Wnt signaling pathway. Oncol Lett. (2016) 11:678–84. doi: 10.3892/ol.2015.3917
60. Vartanian A, Gatsina G, Grigorieva I, Solomko E, Dombrovsky V, Baryshnikov A, et al. The involvement of Notch signaling in melanoma vasculogenic mimicry. Clin Exp Med. (2013) 13:201–9. doi: 10.1007/s10238-012-0190-9
61. Wang Y, Yang R, Wang X, Ci H, Zhou L, Zhu B, et al. Evaluation of the correlation of vasculogenic mimicry, Notch4, DLL4, and KAI1/CD82 in the prediction of metastasis and prognosis in non-small cell lung cancer. Medicine. (2018) 97:e13817. doi: 10.1097/MD.0000000000013817
62. McAllister JC, Zhan Q, Weishaupt C, Hsu M-Y, Murphy GF. The embryonic morphogen, Nodal, is associated with channel-like structures in human malignant melanoma xenografts. J Cutan Pathol. (2010) 37(Suppl. 1):19–25. doi: 10.1111/j.1600-0560.2010.01503.x
63. Topczewska JM, Postovit L-M, Margaryan NV, Sam A, Hess AR, Wheaton WW, et al. Embryonic and tumorigenic pathways converge via Nodal signaling: role in melanoma aggressiveness. Nat Med. (2006) 12:925–32. doi: 10.1038/nm1448
64. Gong W, Sun B, Zhao X, Zhang D, Sun J, Liu T, et al. Nodal signaling promotes vasculogenic mimicry formation in breast cancer via the Smad2/3 pathway. Oncotarget. (2016) 7:70152–67. doi: 10.18632/oncotarget.12161
65. Azad T, Ghahremani M, Yang X. The role of YAP and TAZ in angiogenesis and vascular mimicry. Cells. (2019) 8:407. doi: 10.3390/cells8050407
66. Bora-Singhal N, Nguyen J, Schaal C, Perumal D, Singh S, Coppola D, et al. YAP1 regulates OCT4 activity and SOX2 expression to facilitate self-renewal and vascular mimicry of stem-like cells. Stem Cells. (2015) 33:1705–18. doi: 10.1002/stem.1993
67. Wei H, Wang F, Wang Y, Li T, Xiu P, Zhong J, et al. Verteporfin suppresses cell survival, angiogenesis and vasculogenic mimicry of pancreatic ductal adenocarcinoma via disrupting the YAP-TEAD complex. Cancer Sci. (2017) 108:478–87. doi: 10.1111/cas.13138
68. Liu K, Sun B, Zhao X, Wang X, Li Y, Qiu Z, et al. Hypoxia induced epithelial-mesenchymal transition and vasculogenic mimicry formation by promoting Bcl-2/Twist1 cooperation. Exp Mol Pathol. (2015) 99:383–91. doi: 10.1016/j.yexmp.2015.08.009
69. Du J, Sun B, Zhao X, Gu Q, Dong X, Mo J, et al. Hypoxia promotes vasculogenic mimicry formation by inducing epithelial-mesenchymal transition in ovarian carcinoma. Gynecol Oncol. (2014) 133:575–83. doi: 10.1016/j.ygyno.2014.02.034
70. Zhao N, Sun B, Sun T, Ma Y, Zhao X, Liu Z, et al. Hypoxia-induced vasculogenic mimicry formation via VE-cadherin regulation by Bcl-2. Med Oncol. (2012) 29:3599–607. doi: 10.1007/s12032-012-0245-5
71. Luca M, Huang S, Gershenwald JE, Singh RK, Reich R, Bar-Eli M. Expression of interleukin-8 by human melanoma cells up-regulates MMP-2 activity and increases tumor growth and metastasis. Am J Pathol. (1997) 151:1105–13.
72. Angara K, Borin TF, Rashid MH, Lebedyeva I, Ara R, Lin P-C, et al. CXCR2-expressing tumor cells drive vascular mimicry in antiangiogenic therapy-resistant glioblastoma. Neoplasia. (2018) 20:1070–82. doi: 10.1016/j.neo.2018.08.011
73. Sharma I, Singh A, Siraj F, Saxena S. IL-8/CXCR1/2 signalling promotes tumor cell proliferation, invasion and vascular mimicry in glioblastoma. J Biomed Sci. (2018) 25:62. doi: 10.1186/s12929-018-0464-y
74. Aikins AR, Kim M, Raymundo B, Kim C-W. Downregulation of transgelin blocks interleukin-8 utilization and suppresses vasculogenic mimicry in breast cancer cells. Exp Biol Med. (2017) 242:573–83. doi: 10.1177/1535370216685435
75. Mourad-Zeidan AA, Melnikova VO, Wang H, Raz A, Bar-Eli M. Expression profiling of Galectin-3-depleted melanoma cells reveals its major role in melanoma cell plasticity and vasculogenic mimicry. Am J Pathol. (2008) 173:1839–52. doi: 10.2353/ajpath.2008.080380
76. Lissitzky J-C, Parriaux D, Ristorcelli E, Vérine A, Lombardo D, Verrando P. Cyclic AMP signaling as a mediator of vasculogenic mimicry in aggressive human melanoma cells in vitro. Cancer Res. (2009) 69:802–9. doi: 10.1158/0008-5472.CAN-08-2391
77. Hulin J-A, Tommasi S, Elliot D, Hu DG, Lewis BC, Mangoni AA. MiR-193b regulates breast cancer cell migration and vasculogenic mimicry by targeting dimethylarginine dimethylaminohydrolase 1. Sci Rep. (2017) 7:13996. doi: 10.1038/s41598-017-14454-1
78. Basu GD, Liang WS, Stephan DA, Wegener LT, Conley CR, Pockaj BA, et al. A novel role for cyclooxygenase-2 in regulating vascular channel formation by human breast cancer cells. Breast Cancer Res. (2006) 8:R69. doi: 10.1186/bcr1626
79. Robertson FM, Simeone A-M, Lucci A, McMurray JS, Ghosh S, Cristofanilli M. Differential regulation of the aggressive phenotype of inflammatory breast cancer cells by prostanoid receptors EP3 and EP4. Cancer. (2010) 116:2806–14. doi: 10.1002/cncr.25167
80. Hendrix MJ, Seftor EA, Meltzer PS, Gardner LM, Hess AR, Kirschmann DA, et al. Expression and functional significance of VE-cadherin in aggressive human melanoma cells: role in vasculogenic mimicry. Proc Natl Acad Sci USA. (2001) 98:8018–23. doi: 10.1073/pnas.131209798
81. Hess AR, Postovit L-M, Margaryan NV, Seftor EA, Schneider GB, Seftor REB, et al. Focal adhesion kinase promotes the aggressive melanoma phenotype. Cancer Res. (2005) 65:9851–60. doi: 10.1158/0008-5472.CAN-05-2172
82. Liu W, Xu G, Jia W, Li J, Ma J, Chen K, et al. Prognostic significance and mechanisms of patterned matrix vasculogenic mimicry in hepatocellular carcinoma. Med Oncol. (2011) 28(Suppl. 1):S228–38. doi: 10.1007/s12032-010-9706-x
83. Koshikawa N, Giannelli G, Cirulli V, Miyazaki K, Quaranta V. Role of cell surface metalloprotease MT1-MMP in epithelial cell migration over laminin-5. J Cell Biol. (2000) 148:615–24. doi: 10.1083/jcb.148.3.615
84. Czekierdowski A, Czekierdowska S, Stachowicz N, Łozinski T, Gurynowicz G. Mig-7 expression and vasculogenic mimicry in malignant ovarian tumors. Ginekol Pol. (2017) 88:552–561. doi: 10.5603/GP.a2017.0100
85. Sun Q, Zou X, Zhang T, Shen J, Yin Y, Xiang J. The role of miR-200a in vasculogenic mimicry and its clinical significance in ovarian cancer. Gynecol Oncol. (2014) 132:730–8. doi: 10.1016/j.ygyno.2014.01.047
86. Davidson B, Goldberg I, Gotlieb WH, Kopolovic J, Ben-Baruch G, Nesland JM, et al. High levels of MMP-2, MMP-9, MT1-MMP and TIMP-2 mRNA correlate with poor survival in ovarian carcinoma. Clin Exp Metastasis. (1999) 17:799–808. doi: 10.1023/A:1006723011835
87. Hess AR, Seftor EA, Seftor REB, Hendrix MJC. Phosphoinositide 3-kinase regulates membrane Type 1-matrix metalloproteinase (MMP) and MMP-2 activity during melanoma cell vasculogenic mimicry. Cancer Res. (2003) 63:4757–62.
88. Zhao X, Guan J-L. Focal adhesion kinase and its signaling pathways in cell migration and angiogenesis. Adv Drug Deliv Rev. (2011) 63:610–5. doi: 10.1016/j.addr.2010.11.001
89. Mabuchi S, Kuroda H, Takahashi R, Sasano T. The PI3K/AKT/mTOR pathway as a therapeutic target in ovarian cancer. Gynecol Oncol. (2015) 137:173–9. doi: 10.1016/j.ygyno.2015.02.003
90. Vartanian A, Stepanova E, Grigorieva I, Solomko E, Baryshnikov A, Lichinitser M. VEGFR1 and PKCα signaling control melanoma vasculogenic mimicry in a VEGFR2 kinase-independent manner. Melanoma Res. (2011) 21:91–8. doi: 10.1097/CMR.0b013e328343a237
91. Yao X, Ping Y, Liu Y, Chen K, Yoshimura T, Liu M, et al. Vascular endothelial growth factor receptor 2 (VEGFR-2) plays a key role in vasculogenic mimicry formation, neovascularization and tumor initiation by Glioma stem-like cells. PLoS ONE. (2013) 8:e57188. doi: 10.1371/journal.pone.0057188
92. Wang J-Y, Sun T, Zhao X-L, Zhang S-W, Zhang D-F, Gu Q, et al. Functional significance of VEGF-a in human ovarian carcinoma: role in vasculogenic mimicry. Cancer Biol Ther. (2008) 7:758–66. doi: 10.4161/cbt.7.5.5765
93. Millimaggi D, Mari M, D'Ascenzo S, Giusti I, Pavan A, Dolo V. Vasculogenic mimicry of human ovarian cancer cells: role of CD147. Int J Oncol. (2009) 35:1423–8. doi: 10.3892/ijo_00000460
94. Petty AP, Garman KL, Winn VD, Spidel CM, Lindsey JS. Overexpression of carcinoma and embryonic cytotrophoblast cell-specific Mig-7 induces invasion and vessel-like structure formation. Am J Pathol. (2007) 170:1763–80. doi: 10.2353/ajpath.2007.060969
95. Chen Y, Zhang L, Liu W-X, Wang K. VEGF and SEMA4D have synergistic effects on the promotion of angiogenesis in epithelial ovarian cancer. Cell Mol Biol Lett. (2018) 23:2. doi: 10.1186/s11658-017-0058-9
96. Xia Y, Cai X-Y, Fan J-Q, Zhang L-L, Ren J-H, Li Z-Y, et al. The role of sema4D in vasculogenic mimicry formation in non-small cell lung cancer and the underlying mechanisms. Int J Cancer. (2019) 144:2227–38. doi: 10.1002/ijc.31958
97. Taniguchi H, Suzuki Y, Natori Y. The evolving landscape of cancer stem cells and ways to overcome cancer heterogeneity. Cancers. (2019) 11:532. doi: 10.3390/cancers11040532
98. Chiao M-T, Yang Y-C, Cheng W-Y, Shen C-C, Ko J-L. CD133+ glioblastoma stem-like cells induce vascular mimicry in vivo. Curr Neurovasc Res. (2011) 8:210–9. doi: 10.2174/156720211796558023
99. Lai C-Y, Schwartz BE, Hsu M-Y. CD133+ melanoma subpopulations contribute to perivascular niche morphogenesis and tumorigenicity through vasculogenic mimicry. Cancer Res. (2012) 72:5111–8. doi: 10.1158/0008-5472.CAN-12-0624
100. Zhang D, Sun B, Zhao X, Ma Y, Ji R, Gu Q, et al. Twist1 expression induced by sunitinib accelerates tumor cell vasculogenic mimicry by increasing the population of CD133+ cells in triple-negative breast cancer. Mol Cancer. (2014) 13:207. doi: 10.1186/1476-4598-13-207
101. Wang S-S, Gao X-L, Liu X, Gao S-Y, Fan Y-L, Jiang Y-P, et al. CD133+ cancer stem-like cells promote migration and invasion of salivary adenoid cystic carcinoma by inducing vasculogenic mimicry formation. Oncotarget. (2016) 7:29051–62. doi: 10.18632/oncotarget.8665
102. Xing P, Dong H, Liu Q, Zhao T, Yao F, Xu Y, et al. ALDH1 expression and vasculogenic mimicry are positively associated with poor prognosis in patients with breast cancer. Cell Physiol Biochem. (2018) 49:961–70. doi: 10.1159/000493227
103. Zhu B, Zhou L, Yu L, Wu S, Song W, Gong X, et al. Evaluation of the correlation of vasculogenic mimicry, ALDH1, KAI1 and microvessel density in the prediction of metastasis and prognosis in colorectal carcinoma. BMC Surg. (2017) 17:47. doi: 10.1186/s12893-017-0246-6
104. Yu L, Zhu B, Wu S, Zhou L, Song W, Gong X, et al. Evaluation of the correlation of vasculogenic mimicry, ALDH1, KiSS-1, and MACC1 in the prediction of metastasis and prognosis in ovarian carcinoma. Diagn Pathol. (2017) 12:23. doi: 10.1186/s13000-017-0612-9
105. Wang H-F, Wang S-S, Zheng M, Dai L-L, Wang K, Gao X-L, et al. Hypoxia promotes vasculogenic mimicry formation by vascular endothelial growth factor A mediating epithelial-mesenchymal transition in salivary adenoid cystic carcinoma. Cell Prolif. (2019) 52:e12600. doi: 10.1111/cpr.12600
106. Mao X-G, Xue X-Y, Wang L, Zhang X, Yan M, Tu Y-Y, et al. CDH5 is specifically activated in glioblastoma stemlike cells and contributes to vasculogenic mimicry induced by hypoxia. Neuro Oncol. (2013) b:865–79. doi: 10.1093/neuonc/not029
107. Jung JE, Lee HG, Cho IH, Chung DH, Yoon S-H, Yang YM, et al. STAT3 is a potential modulator of HIF-1-mediated VEGF expression in human renal carcinoma cells. FASEB J. (2005) 19:1296–8. doi: 10.1096/fj.04-3099fje
108. Li S, Meng W, Guan Z, Guo Y, Han X. The hypoxia-related signaling pathways of vasculogenic mimicry in tumor treatment. Biomed Pharmacother. (2016) 80:127–35. doi: 10.1016/j.biopha.2016.03.010
109. Song Y-Y, Sun L-D, Liu M-L, Liu Z-L, Chen F, Zhang Y-Z, et al. STAT3, p-STAT3 and HIF-1α are associated with vasculogenic mimicry and impact on survival in gastric adenocarcinoma. Oncol Lett. (2014) 8:431–7. doi: 10.3892/ol.2014.2059
110. Xu Q, Briggs J, Park S, Niu G, Kortylewski M, Zhang S, et al. Targeting Stat3 blocks both HIF-1 and VEGF expression induced by multiple oncogenic growth signaling pathways. Oncogene. (2005) 24:5552–60. doi: 10.1038/sj.onc.1208719
111. Gest C, Mirshahi P, Li H, Pritchard L-L, Joimel U, Blot E, et al. Ovarian cancer: Stat3, RhoA and IGF-IR as therapeutic targets. Cancer Lett. (2012) 317:207–17. doi: 10.1016/j.canlet.2011.11.026
112. Dejana E. The role of wnt signaling in physiological and pathological angiogenesis. Circ Res. (2010) 107:943–52. doi: 10.1161/CIRCRESAHA.110.223750
113. Qi L, Song W, Liu Z, Zhao X, Cao W, Sun B. Wnt3a promotes the vasculogenic mimicry formation of colon cancer via Wnt/β-catenin signaling. Int J Mol Sci. (2015) 16:18564–79. doi: 10.3390/ijms160818564
114. Qi H, Sun B, Zhao X, Du J, Gu Q, Liu Y, et al. Wnt5a promotes vasculogenic mimicry and epithelial-mesenchymal transition via protein kinase Cα in epithelial ovarian cancer. Oncol Rep. (2014) 32:771–9. doi: 10.3892/or.2014.3229
115. Su M, Wei W, Xu X, Wang X, Chen C, Su L, et al. Role of hCG in vasculogenic mimicry in OVCAR-3 ovarian cancer cell line. Int J Gynecol Cancer. (2011) 21:1366–74. doi: 10.1097/IGC.0b013e31822c7529
116. Gao S, Fan C, Huang H, Zhu C, Su M, Zhang Y. Effects of HCG on human epithelial ovarian cancer vasculogenic mimicry formation in vivo. Oncol Lett. (2016) 12:459–66. doi: 10.3892/ol.2016.4630
117. Su M, Xu X, Wei W, Gao S, Wang X, Chen C, et al. Involvement of human chorionic gonadotropin in regulating vasculogenic mimicry and hypoxia-inducible factor-1α expression in ovarian cancer cells. Cancer Cell Int. (2016) 16:50. doi: 10.1186/s12935-016-0327-0
118. Di Leva G, Garofalo M, Croce CM. MicroRNAs in cancer. Annu Rev Pathol. (2014) 9:287–314. doi: 10.1146/annurev-pathol-012513-104715
119. Wang Z, Zheng C, Jiang K, He J, Cao X, Wu S. MicroRNA-503 suppresses cell proliferation and invasion in osteosarcoma via targeting insulin-like growth factor 1 receptor. Exp Ther Med. (2017) 14:1547–53. doi: 10.3892/etm.2017.4648
120. Zeng Q, Wang Y, Gao J, Yan Z, Li Z, Zou X, et al. miR-29b-3p regulated osteoblast differentiation via regulating IGF-1 secretion of mechanically stimulated osteocytes. Cell Mol Biol Lett. (2019) 24:11. doi: 10.1186/s11658-019-0136-2
121. Li Q, Li Y, Zhang D, Gao H, Gao X. Downregulation of microRNA-451 improves cell migration, invasion and tube formation in hypoxia-treated HUVECs by targeting MIF. Mol Med Rep. (2019) 20:1167–77. doi: 10.3892/mmr.2019.10357
122. Li G, Huang M, Cai Y, Ke Y, Yang Y, Sun X. miR-141 inhibits glioma vasculogenic mimicry by controlling EphA2 expression. Mol Med Rep. (2018) 18:1395–1404. doi: 10.3892/mmr.2018.9108
123. Wang Y, Sun B, Sun H, Zhao X, Wang X, Zhao N, et al. Regulation of proliferation, angiogenesis and apoptosis in hepatocellular carcinoma by miR-26b-5p. Tumour Biol. (2016) 37:10965–79. doi: 10.1007/s13277-016-4964-7
124. Wu N, Zhao X, Liu M, Liu H, Yao W, Zhang Y, et al. Role of microRNA-26b in glioma development and its mediated regulation on EphA2. PLoS ONE. (2011) 6:e16264. doi: 10.1371/journal.pone.0016264
125. Chen X, Wang X, Ruan A, Han W, Zhao Y, Lu X, et al. miR-141 is a key regulator of renal cell carcinoma proliferation and metastasis by controlling EphA2 expression. Clin Cancer Res. (2014) 20:2617–30. doi: 10.1158/1078-0432.CCR-13-3224
126. Liu W, Lv C, Zhang B, Zhou Q, Cao Z. MicroRNA-27b functions as a new inhibitor of ovarian cancer-mediated vasculogenic mimicry through suppression of VE-cadherin expression. RNA. (2017) 23:1019–27. doi: 10.1261/rna.059592.116
127. Zhao N, Sun H, Sun B, Zhu D, Zhao X, Wang Y, et al. miR-27a-3p suppresses tumor metastasis and VM by down-regulating VE-cadherin expression and inhibiting EMT: an essential role for Twist-1 in HCC. Sci Rep. (2016) 6:23091. doi: 10.1038/srep23091
128. Salinas-Vera YM, Gallardo-Rincón D, García-Vázquez R, Hernández-de la Cruz ON, Marchat LA, González-Barrios JA, et al. HypoxamiRs profiling identify miR-745 as a regulator of the early stages of vasculogenic mimicry in SKOV3 ovarian cancer cells. Front Oncol. (2019) 9:381. doi: 10.3389/fonc.2019.00381
129. Yang J, Lu Y, Lin Y-Y, Zheng Z-Y, Fang J-H, He S, et al. Vascular mimicry formation is promoted by paracrine TGF-β and SDF1 of cancer-associated fibroblasts and inhibited by miR-101 in hepatocellular carcinoma. Cancer Lett. (2016) 383:18–27. doi: 10.1016/j.canlet.2016.09.012
130. Salinas-Vera YM, Marchat LA, García-Vázquez R, González de la Rosa CH, Castañeda-Saucedo E, Tito NN, et al. Cooperative multi-targeting of signaling networks by angiomiR-204 inhibits vasculogenic mimicry in breast cancer cells. Cancer Lett. (2018) 432:17–27. doi: 10.1016/j.canlet.2018.06.003
131. Gao Y, Yu H, Liu Y, Liu X, Zheng J, Ma J, et al. Long Non-Coding RNA HOXA-AS2 regulates malignant glioma behaviors and vasculogenic mimicry formation via the MiR-373/EGFR Axis. Cell Physiol Biochem. (2018) 45:131–47. doi: 10.1159/000486253
132. Cao C, Sun D, Zhang L, Song L. miR-186 affects the proliferation, invasion and migration of human gastric cancer by inhibition of Twist1. Oncotarget. (2016) 7:79956–63. doi: 10.18632/oncotarget.13182
133. Chang Z, Cui J, Song Y. Long noncoding RNA PVT1 promotes EMT via mediating microRNA-186 targeting of Twist1 in prostate cancer. Gene. (2018) 654:36–42. doi: 10.1016/j.gene.2018.02.036
134. Fang J-H, Zheng Z-Y, Liu J-Y, Xie C, Zhang Z-J, Zhuang S-M. Regulatory role of the microRNA-29b-IL-6 signaling in the formation of vascular mimicry. Mol Ther Nucleic Acids. (2017) 8:90–100. doi: 10.1016/j.omtn.2017.06.009
135. Guo J, Cai H, Liu X, Zheng J, Liu Y, Gong W, et al. Long non-coding RNA LINC00339 stimulates glioma vasculogenic mimicry formation by regulating the miR-539-5p/TWIST1/MMPs Axis. Mol Ther Nucleic Acids. (2018) 10:170–86. doi: 10.1016/j.omtn.2017.11.011
136. Tao W, Sun W, Zhu H, Zhang J. Knockdown of long non-coding RNA TP73-AS1 suppresses triple negative breast cancer cell vasculogenic mimicry by targeting miR-490-3p/TWIST1 axis. Biochem Biophys Res Commun. (2018) 504:629–34. doi: 10.1016/j.bbrc.2018.08.122
137. Kota J, Chivukula RR, O'Donnell KA, Wentzel EA, Montgomery CL, Hwang H-W, et al. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell. (2009) 137:1005–17. doi: 10.1016/j.cell.2009.04.021
138. Si Y, Zhang H, Ning T, Bai M, Wang Y, Yang H, et al. miR-26a/b inhibit tumor growth and angiogenesis by targeting the HGF-VEGF axis in gastric carcinoma. Cell Physiol Biochem. (2017) 42:1670–83. doi: 10.1159/000479412
139. Shimono Y, Zabala M, Cho RW, Lobo N, Dalerba P, Qian D, et al. Downregulation of miRNA-200c links breast cancer stem cells with normal stem cells. Cell. (2009) 138:592–603. doi: 10.1016/j.cell.2009.07.011
140. Karihtala P, Porvari K, Soini Y, Eskelinen M, Juvonen P, Haapasaari K-M. Expression levels of microRNAs miR-93 and miR-200a in pancreatic adenocarcinoma with special reference to differentiation and relapse-free survival. Oncology. (2019) 96:164–70. doi: 10.1159/000494274
141. Liu D, Song L, Dai Z, Guan H, Kang H, Zhang Y, et al. MiR-429 suppresses neurotrophin-3 to alleviate perineural invasion of pancreatic cancer. Biochem Biophys Res Commun. (2018) 505:1077–83. doi: 10.1016/j.bbrc.2018.09.147
142. Li Y, Guan B, Liu J, Zhang Z, He S, Zhan Y, et al. MicroRNA-200b is downregulated and suppresses metastasis by targeting LAMA4 in renal cell carcinoma. EBioMedicine. (2019) 44:439–51. doi: 10.1016/j.ebiom.2019.05.041
143. Guo FJ, Shao YP, Wang YP, Jin YM, Liu SS, Wang QY. MIR-92 stimulates VEGF by inhibiting von Hippel-Lindau gene product in epithelial ovarian cancer. J Biol Regul Homeost Agents. (2017) 31:615–24.
144. Joshi HP, Subramanian IV, Schnettler EK, Ghosh G, Rupaimoole R, Evans C, et al. Dynamin 2 along with microRNA-199a reciprocally regulate hypoxia-inducible factors and ovarian cancer metastasis. Proc Natl Acad Sci USA. (2014) 111:5331–6. doi: 10.1073/pnas.1317242111
145. He J, Jing Y, Li W, Qian X, Xu Q, Li F-S, et al. Roles and mechanism of miR-199a and miR-125b in tumor angiogenesis. PLoS ONE. (2013) 8:e56647. doi: 10.1371/journal.pone.0056647
146. Yeh Y-M, Chuang C-M, Chao K-C, Wang L-H. MicroRNA-138 suppresses ovarian cancer cell invasion and metastasis by targeting SOX4 and HIF-1α. Int J Cancer. (2013) 133:867–78. doi: 10.1002/ijc.28086
147. Xu Q, Liu L-Z, Qian X, Chen Q, Jiang Y, Li D, et al. MiR-145 directly targets p70S6K1 in cancer cells to inhibit tumor growth and angiogenesis. Nucleic Acids Res. (2012) 40:761–74. doi: 10.1093/nar/gkr730
148. Leng R, Zha L, Tang L. MiR-718 represses VEGF and inhibits ovarian cancer cell progression. FEBS Lett. (2014) 588:2078–86. doi: 10.1016/j.febslet.2014.04.040
149. Luo J, Zhu C, Wang H, Yu L, Zhou J. MicroRNA-126 affects ovarian cancer cell differentiation and invasion by modulating expression of vascular endothelial growth factor. Oncol Lett. (2018) 15:5803–08. doi: 10.3892/ol.2018.8025
150. Wang W, Ren F, Wu Q, Jiang D, Li H, Shi H. MicroRNA-497 suppresses angiogenesis by targeting vascular endothelial growth factor A through the PI3K/AKT and MAPK/ERK pathways in ovarian cancer. Oncol Rep. (2014) 32:2127–33. doi: 10.3892/or.2014.3439
151. Nishimura M, Jung E-J, Shah MY, Lu C, Spizzo R, Shimizu M, et al. Therapeutic synergy between microRNA and siRNA in ovarian cancer treatment. Cancer Discov. (2013) 3:1302–15. doi: 10.1158/2159-8290.CD-13-0159
152. Wang Y, Xu C, Wang Y, Zhang X. MicroRNA-365 inhibits ovarian cancer progression by targeting Wnt5a. Am J Cancer Res. (2017) 7:1096–1106.
153. Chen S, Chen X, Xiu Y-L, Sun K-X, Zhao Y. MicroRNA-490-3P targets CDK1 and inhibits ovarian epithelial carcinoma tumorigenesis and progression. Cancer Lett. (2015) 362:122–30. doi: 10.1016/j.canlet.2015.03.029
154. Chen S, Chen X, Xiu Y-L, Sun K-X, Zhao Y. Inhibition of ovarian epithelial carcinoma tumorigenesis and progression by microRNA 106b mediated through the RhoC pathway. PLoS ONE. (2015) 10:e0125714. doi: 10.1371/journal.pone.0125714
155. Duan Y, Dong Y, Dang R, Hu Z, Yang Y, Hu Y, et al. MiR-122 inhibits epithelial mesenchymal transition by regulating P4HA1 in ovarian cancer cells. Cell Biol Int. (2018) 42:1564–74. doi: 10.1002/cbin.11052
156. Fan B, Chen L-P, Yuan Y-H, Xiao H-N, Lv X-S, Xia Z-Y. MiR-15a-3p suppresses the growth and metastasis of ovarian cancer cell by targeting Twist1. Eur Rev Med Pharmacol Sci. (2019) 23:1934–46. doi: 10.26355/eurrev_201903_17232
157. Wei H, Tang Q-L, Zhang K, Sun J-J, Ding R-F. miR-532-5p is a prognostic marker and suppresses cells proliferation and invasion by targeting TWIST1 in epithelial ovarian cancer. Eur Rev Med Pharmacol Sci. (2018) 22:5842–50. doi: 10.26355/eurrev_201809_15911
158. Wei C, Zhang X, He S, Liu B, Han H, Sun X. MicroRNA-219-5p inhibits the proliferation, migration, and invasion of epithelial ovarian cancer cells by targeting the Twist/Wnt/β-catenin signaling pathway. Gene. (2017) 637:25–32. doi: 10.1016/j.gene.2017.09.012
159. Li C, Duan P, Wang J, Lu X, Cheng J. miR-320 inhibited ovarian cancer oncogenicity via targeting TWIST1 expression. Am J Transl Res. (2017) 9:3705–13.
160. Sun X, Cui M, Zhang A, Tong L, Wang K, Li K, et al. MiR-548c impairs migration and invasion of endometrial and ovarian cancer cells via downregulation of Twist. J Exp Clin Cancer Res. (2016) 35:10. doi: 10.1186/s13046-016-0288-0
161. Liu Y, Zhou H, Ma L, Hou Y, Pan J, Sun C, et al. MiR-214 suppressed ovarian cancer and negatively regulated semaphorin 4D. Tumour Biol. (2016) 37:8239–48. doi: 10.1007/s13277-015-4708-0
162. Mitra AK, Chiang CY, Tiwari P, Tomar S, Watters KM, Peter ME, et al. Microenvironment-induced downregulation of miR-193b drives ovarian cancer metastasis. Oncogene. (2015) 34:5923–32. doi: 10.1038/onc.2015.43
163. Yan J, Jiang J, Meng X-N, Xiu Y-L, Zong Z-H. MiR-23b targets cyclin G1 and suppresses ovarian cancer tumorigenesis and progression. J Exp Clin Cancer Res. (2016) 35:31. doi: 10.1186/s13046-016-0307-1
164. Tian F, Jia L, Chu Z, Han H, Zhang Y, Cai J. MicroRNA-519a inhibits the proliferation and promotes the apoptosis of ovarian cancer cells through targeting signal transducer and activator of transcription 3. Exp Ther Med. (2018) 15:1819–24. doi: 10.3892/etm.2017.5600
165. Al-Ansary GH, Eldehna WM, Ghabbour HA, Al-Rashood STA, Al-Rashood KA, Eladwy RA, et al. Cancer stem cells CD133 inhibition and cytotoxicity of certain 3-phenylthiazolo[3,2-a]benzimi. J Enzyme Inhib Med Chem. (2017) 32:986–91. doi: 10.1080/14756366.2017.1347166
166. Huang B, Yin M, Li X, Cao G, Qi J, Lou G, et al. Migration-inducing gene 7 promotes tumorigenesis and angiogenesis and independently predicts poor prognosis of epithelial ovarian cancer. Oncotarget. (2016) 7:27552–66. doi: 10.18632/oncotarget.8487
167. Heinemann V, Ebert MP, Laubender RP, Bevan P, Mala C, Boeck S. Phase II randomised proof-of-concept study of the urokinase inhibitor upamostat (WX-671) in combination with gemcitabine compared with gemcitabine alone in patients with non-resectable, locally advanced pancreatic cancer. Br J Cancer. (2013) 108:766–70. doi: 10.1038/bjc.2013.62
168. Wang J, Peng Y, Sun YW, He H, Zhu S, An X, et al. All-trans retinoic acid induces XAF1 expression through an interferon regulatory factor-1 element in colon cancer. Gastroenterology. (2006) 130:747–58. doi: 10.1053/j.gastro.2005.12.017
169. Fu Z, Wang L, Cui H, Peng J, Wang S, Geng J, et al. A novel small-molecule compound targeting CD147 inhibits the motility and invasion of hepatocellular carcinoma cells. Oncotarget. (2016) 7:9429–47. doi: 10.18632/oncotarget.6990
170. Nozawa-Suzuki N, Nagasawa H, Ohnishi K, Morishige K-I. The inhibitory effect of hypoxic cytotoxin on the expansion of cancer stem cells in ovarian cancer. Biochem Biophys Res Commun. (2015) 457:706–11. doi: 10.1016/j.bbrc.2015.01.053
171. Su W, Huang L, Ao Q, Zhang Q, Tian X, Fang Y, et al. Noscapine sensitizes chemoresistant ovarian cancer cells to cisplatin through inhibition of HIF-1α. Cancer Lett. (2011) 305:94–9. doi: 10.1016/j.canlet.2011.02.031
172. Heinzlmeir S, Lohse J, Treiber T, Kudlinzki D, Linhard V, Gande SL, et al. Chemoproteomics-aided medicinal chemistry for the discovery of EPHA2 inhibitors. Chem Med Chem. (2017) 12:999–1011. doi: 10.1002/cmdc.201700217
173. Polena H, Creuzet J, Dufies M, Sidibé A, Khalil-Mgharbel A, Salomon A, et al. The tyrosine-kinase inhibitor sunitinib targets vascular endothelial (VE)-cadherin: a marker of response to antitumoural treatment in metastatic renal cell carcinoma. Br J Cancer. (2018) 118:1179–88. doi: 10.1038/s41416-018-0054-5
174. Guo J-Q, Zheng Q-H, Chen H, Chen L, Xu J-B, Chen M-Y, et al. Ginsenoside Rg3 inhibition of vasculogenic mimicry in pancreatic cancer through downregulation of VE-cadherin/EphA2/MMP9/MMP2 expression. Int J Oncol. (2014) 45:1065–72. doi: 10.3892/ijo.2014.2500
175. Liu T, Zhao L, Zhang Y, Chen W, Liu D, Hou H, et al. Ginsenoside 20(S)-Rg3 targets HIF-1α to block hypoxia-induced epithelial-mesenchymal transition in ovarian cancer cells. PLoS ONE. (2014) 9:e103887. doi: 10.1371/journal.pone.0103887
176. Liu T, Zhao L, Hou H, Ding L, Chen W, Li X. Ginsenoside 20(S)-Rg3 suppresses ovarian cancer migration via hypoxia-inducible factor 1 alpha and nuclear factor-kappa B signals. Tumour Biol. (2017) 39:1010428317692225. doi: 10.1177/1010428317692225
177. Rodríguez MI, Peralta-Leal A, O'Valle F, Rodriguez-Vargas JM, Gonzalez-Flores A, Majuelos-Melguizo J, et al. PARP-1 regulates metastatic melanoma through modulation of vimentin-induced malignant transformation. PLoS Genet. (2013) 9:e1003531. doi: 10.1371/journal.pgen.1003531
179. Tempfer CB, Schultheis B, Hilal Z, Dogan A, Rezniczek GA. Thalidomide and lenalidomide for recurrent ovarian cancer: a systematic review of the literature. Oncol Lett. (2017) 14:3327–36. doi: 10.3892/ol.2017.6578
180. Bauer JA, Morrison BH, Grane RW, Jacobs BS, Borden EC, Lindner DJ. IFN-alpha2b and thalidomide synergistically inhibit tumor-induced angiogenesis. J Interferon Cytokine Res. (2003) 23:3–10. doi: 10.1089/10799900360520397
181. Zhang S, Li M, Gu Y, Liu Z, Xu S, Cui Y, et al. Thalidomide influences growth and vasculogenic mimicry channel formation in melanoma. J Exp Clin Cancer Res. (2008) 27:60. doi: 10.1186/1756-9966-27-60
182. Ding J, Jia X, Zuo B, He J, Yang J, He Y. A novel monoclonal antibody targeting a novel epitope of VE-cadherin inhibits vasculogenic mimicry of lung cancer cells. Oncol Rep. (2018) 39:2837–44. doi: 10.3892/or.2018.6374
Keywords: vasculogenic mimicry, ovarian cancer, signaling molecules, angiogenesis, anti-angiogenic therapy
Citation: Ayala-Domínguez L, Olmedo-Nieva L, Muñoz-Bello JO, Contreras-Paredes A, Manzo-Merino J, Martínez-Ramírez I and Lizano M (2019) Mechanisms of Vasculogenic Mimicry in Ovarian Cancer. Front. Oncol. 9:998. doi: 10.3389/fonc.2019.00998
Received: 22 June 2019; Accepted: 17 September 2019;
Published: 27 September 2019.
Edited by:
Laurence A. Marchat, National Polytechnic Institute, MexicoReviewed by:
Kwok-Ming Yao, The University of Hong Kong, Hong KongMaja Cemazar, Institute of Oncology Ljubljana, Slovenia
Copyright © 2019 Ayala-Domínguez, Olmedo-Nieva, Muñoz-Bello, Contreras-Paredes, Manzo-Merino, Martínez-Ramírez and Lizano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marcela Lizano, lizano@unam.mx