 Grace Armijo
Grace Armijo Rudolf Schlechter
Rudolf Schlechter Mario Agurto
Mario Agurto Daniela Muñoz
Daniela Muñoz Patricio Arce-Johnson
Patricio Arce-Johnson- Laboratorio de Biología Molecular y Biotecnología Vegetal, Departamento de Genética Molecular y Microbiología, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Santiago, Chile
Grapevine (Vitis vinifera L.) is one of the most important fruit crop worldwide. Commercial cultivars are greatly affected by a large number of pathogenic microorganisms that cause diseases during pre- and/or post-harvest periods, affecting production, processing and export, along with fruit quality. Among the potential threats, we can find bacteria, fungi, oomycete, or viruses with different life cycles, infection mechanisms and evasion strategies. While plant–pathogen interactions are cycles of resistance and susceptibility, resistance traits from natural resources are selected and may be used for breeding purposes and for a sustainable agriculture. In this context, here we summarize some of the most important diseases affecting V. vinifera together with their causal agents. The aim of this work is to bring a comprehensive review of the infection strategies deployed by significant types of pathogens while understanding the host response in both resistance and susceptibility scenarios. New approaches being used to uncover grapevine status during biotic stresses and scientific-based procedures needed to control plant diseases and crop protection are also addressed.
Introduction
During their lifetime, plants are exposed to a wide variety of pathogens, such as bacteria, viruses, fungi, and nematodes. According to their lifecycle and infection strategies, pathogenic microorganisms can be classified as necrotrophics, biotrophics and hemibiotrophics: necrotrophic pathogens feed on dead tissue, secreting lytic enzymes and phytotoxins to promote cell death into the host plant. Biotrophic pathogens on the other hand, feed on living tissue, developing structures in order to invade the cell and obtain metabolism products. Finally, hemibiotrophic pathogens start with a biotrophic infection phase and then turn to a final necrotrophic phase, killing its host at the end of the infection cycle (Glazebrook, 2005).
Plant defense mechanisms are tightly regulated by hormone-mediated signaling pathways, mainly jasmonic acid (JA) and salicylic acid (SA). It is generally considered that JA and ethylene (JA/Et) mediate necrotrophic pathogens defense, while SA is involved against the biotrophic and hemibiotrophic ones (Pieterse et al., 2009).
Plants respond to necrotrophic pathogens through the induction of JA/Et biosynthesis, which is increased locally and systemically when microorganisms secrete cell wall-degrading lytic enzymes (Robert-Seilaniantz et al., 2011). In addition to wall components, some phospholipids released by the degradation of the plasma membrane directly activate JA biosynthesis (Turner et al., 2002). This increase in JA levels induces the expression of defense related genes coding for glucanases, chitinases, protease inhibitors, and enzymes involved in the biosynthesis of secondary metabolites such as phytoalexins (Glazebrook, 2005).
On the other hand, SA mediates the response to biotrophic and hemibiotrophic pathogens, inducing an increase of reactive oxygen species (ROS) and, therefore, a localized programmed cell death (PCD) in the infected tissue (Pieterse et al., 2009). This defense mechanism, called hypersensitive response (HR), restricts pathogen growth by limiting their access to nutrients and water (Glazebrook, 2005).
For these events to occur, plants must be able to recognize these pathogens. Three types of plant–pathogen interactions have been described to date. The first, PTI or PAMPs-Triggered Immunity (recently called MTI or MAMPs-Triggered Immunity), is a basal immune response activated after recognition of non-adapted pathogens. This corresponds to a first line of defense, common in plants of the same species facing a potentially pathogenic microorganism (Jones and Dangl, 2006; Pieterse et al., 2009). PTI is mediated by plasma membrane localized pattern recognition receptors (PRRs), which are composed of an extracellular domain able to detect PAMPs or MAMPs (i.e., generally structural components of the pathogen) and a intracellular domain that amplifies the signal inside the cell (Monaghan and Zipfel, 2012). A second type of interaction is the effector-triggered susceptibility (ETS), named after the ability of certain microorganisms to overcome the basal plant response through the secretion of virulence factors (effectors) which inhibit PTI, thus promoting the disease (Pieterse et al., 2009). Lastly, there is a third type of interaction known as effector-triggered immunity (ETI). In this case, plants of a particular genotype can identify pathogen effectors through a second class of receptors, called resistance proteins (R). If an effector is recognized by an R protein, either directly or indirectly, it is considered an avirulence factor (AVR) and then the pathogen is avirulent to that plant, since this interaction leads to the activation of HR (Jones and Dangl, 2006; Hayward et al., 2009).
The grapevine (Vitis vinifera L.) is one of the most important fruit crops worldwide. This specie is greatly affected by a large number of pathogens that cause diseases in pre- and post-harvest periods, affecting production, processing and export, along with fruit quality. Some of the most important diseases in V. vinifera are the gray mold, powdery mildew, and downy mildew (DM), caused by Botrytis cinerea, Erysiphe necator and Plasmopara viticola, respectively, among others (Figure 1). The overall purpose of this review is to synthetize the most important grapevine pathogens and their infection strategies along with the host responses in resistance and susceptibility scenarios, taking into consideration the understanding of Vitis-pathogens interactions due to its agronomic importance.
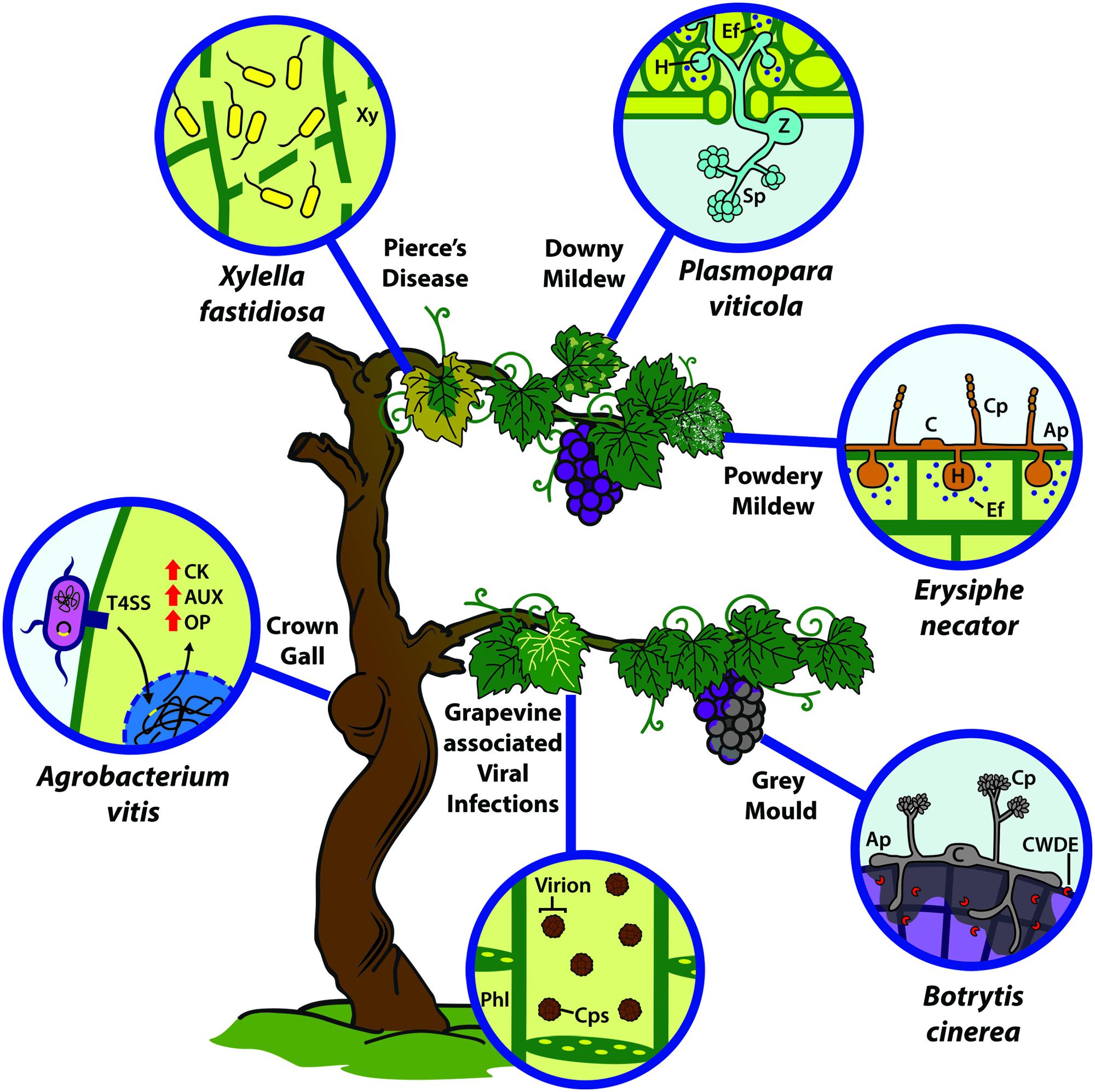
FIGURE 1. Infection strategy of main grapevine pathogenic microorganisms and disease-associated symptoms. Grapevine is affected by different types of microorganisms, i.e., bacteria, fungi, oomycetes and viruses. Agrobacterium vitis causes the grapevine crown gall through the injection of T-DNA sequences via a type-IV secretion system (T4SS), incorporating them in the host genome to induce the synthesis of cytokinins (CK), auxins (AUX) and opines (OP). Xylella fastidiosa is transmitted by insect vectors; it grows and accumulates within the xylem (Xy) vessels causing Pierce’s Disease. Plasmopara viticola zoospores (Z) infect through the stoma in order to accommodate on the host, generating the sporangium (Sp) and generating the grapevine downy mildew (DM) disease. On the other hand, conidia (C) of E. necator, the causal agent of the powdery mildew (PM), infects epidermal layers. However, both P. viticola and E. necator establish their biotrophy by developing haustoria (H) and secreting virulence factors or effectors (Ef) into the host, to manipulate metabolism and defense responses. Botritys cinerea conidia (C) germinate and penetrate the plant tissue by necrotizing the host tissue (mainly in grape berries) through the secretion of cell wall degrading enzymes (CWDE), causing the gray mold disease. Viruses are phloem-limited (Phl) microorganisms, whose infections can cause different symptoms in the host. Ap, appresorium; C, conidium; Cp, conidiophore. Cps, capside.
Necrotrophic Pathogens
Botrytis cinerea
The fungus B. cinerea has the ability to live as a parasite in green tissue and as a saprophyte in dead or decaying ones. This is the reason for its wide distribution in nature and host unspecificity. Particularly in V. vinifera, B. cinerea infection causes a disease known as “gray mold” (Figure 1) in post-harvest periods, affecting complete berry clusters during packing, transport and commercialization and thus becoming one of the most important pathogens affecting export wine and table grapes (Dean et al., 2012).
Infection Strategy
Botrytis cinerea infects grapevine by two main mechanisms: (1) direct mycelium penetration through skin pores or injury, and (2) early invasion, where conidia infect mainly the flower receptacle, and to a lesser extent the stigma and styles, remaining latent within the berry until maturity (Viret et al., 2004). This fungus presents high variability in several biological traits, which can be explained by the great diversity among isolates. However, attachment to a solid surface is a common requisite for conidial germination, though nutritional requirements may differ (Cotoras et al., 2009). When the conidium perceives characteristics and nutrients from the host surface, it develops an infective structure called appressorium that breaches the cuticle by means of a penetration peg (Rolke et al., 2004). It has been described that appressoria-mediated penetration of B. cinerea requires a membrane-associated tetraspanin-encoding gene termed BcPLS1, since Bcpls1-deficient mutants fail to infect intact host cells (Gourgues et al., 2004). The newly formed appressorium secretes lytic enzymes to cross cuticle and outer epithelial wall, like cutinases and lipases, to breach these layers and penetrate the plant cell.
Primary lesion formation triggers an oxidative burst that helps the fungus to kill and degrade the host tissue (Rolke et al., 2004), along with secretion of cell wall degrading enzymes (CWDE) such as endopolygalacturonases (Kars et al., 2005a), pectin methylesterases (Kars et al., 2005b), cellulases and hemicellulases, causing decomposition and consumption of plant biomass. In fact, it has been described that pectin-degrading enzymes are the most abundant plant cell wall modifying proteins expressed during B. cinerea infection in grapevines, such as xyloglucan transglucosylase/hydrolases and glucanases (Blanco-Ulate et al., 2014). Pectin breakdown during infection increases the plant cell wall porosity and may facilitate further polysaccharides degradation. Along with this class of enzymes, B. cinerea secretes toxins and oxalic acid during infection; the latter acidifies locally the infected region, allowing the activation of pectinases and laccases secreted by the fungus, favoring hyphal growth and inducing morphogenic signaling of infectious structures (van Kan, 2006). Furthermore, oxalic acid contributes to the virulence of the pathogen, promoting both host tissue cell death and sporulation in adjacent cells.
The initial step of infection appears to be similar in berries at different developmental stages, since a recent study of Kelloniemi et al. (2015) revealed that germinated conidia developed appressoria on both veraison and berry maturity, and in the two developmental stages expression of genes related to fungal virulence as endopolygalacturonase (BcPG2), pectin methyl esterase (BcPME2), superoxide dismutase (BcSOD1), glutatione-S-transferase (BcGST1), among others, were induced during the first 16–24 h. Nevertheless, B. cinerea failed to colonize berries at veraison and the infection was only successful in mature berries. In the latter, up-regulated genes were enriched in functional categories related to degradation of plant cell wall (CWDE), proteolysis, membrane transport, ROS generation and detoxification, phytotoxic proteins and secondary metabolites such as botrydial and botcinic acid, which suggest that the initial process of infection and penetration by the fungus may be common in these stages of development, but degradation of plant tissue is a critical step for successful infection.
Physiologically, infection of B. cinerea causes significant metabolic changes in berries. High levels of proline, glutamate, arginine, and alanine accumulated in these tissues, accompanied with the production of glycerol, gluconic acid, succinate and largely degraded phenylpropanoids, flavonoid compounds and sucrose (Hong et al., 2012). Moreover, it has been shown that hexokinase activity is required for the development of B. cinerea in the presence of hexoses (Rui and Hahn, 2007), suggesting that the fungus is able to manipulate plant metabolism to use carbon sources required for its growth, in addition to products of tissue degradation.
Host Response and Resistance against B. cinerea
Studies conducted in the Vitis-B. cinerea pathosystem showed that grapevines use natural and pre-existing skin qualities as defense structures, such as number of cell layers, cuticle, and wax content of the fruit (Gabler et al., 2003), along with the quantity and density of stomata and leaf trichomes. Also, activation of inducible defense mechanisms during the infection has been reported, such as the expression of polygalacturonase-inhibiting proteins (PGIP) to reduce the extensive pectin degradation caused by fungal attack (De Lorenzo et al., 2001) or an oxidative burst and phytoalexin accumulation in grape cells and primed leaves (i.e., trans-resveratrol and α-viniferin) necessary for defense activation (Aziz et al., 2003; Verhagen et al., 2011). Structural defenses are related to fungal primary infection processes (i.e., appressoria formation and plant tissue penetration), while inducible responses are associated with subsequent infection processes.
Grapevines are also able to secrete a set of chitinases, e.g., Vvchit1a (acidic class I chitinase), Vvchit1b (basic class I chitinase), and other hydrolytic enzymes to degrade the fungus cell wall (Robert et al., 2002; Chong et al., 2008). However, B. cinerea has the capacity to catalyze the conversion of chitin to chitosan, bypassing the degradation of its cell wall (El Gueddari et al., 2002). Thus, a fine-tuning regulation of different layers of plant defense can be deployed to cope at some extend with the pathogen infection.
As described in other plants, necrotrophic-type infection of V. vinifera activates the JA/ET pathway along with the induction of genes related to phytoalexin biosynthesis, such as phenylalanine ammonia lyase (PAL) and stilbene synthase (STS; Chong et al., 2008; Girault et al., 2008; Thaler et al., 2012; Wang et al., 2015). It has been described that methyl jasmonate (MeJA) treatments in harvested grape berries could induce resistance against B. cinerea and reduce disease incidence. This is associated with an increase of H2O2, enhanced expression of the defense-related gene VvNPR1.1 and accumulation of stilbene phytoalexins, such as tran-resveratrol and its oligomer form, trans-ε-viniferin (Wang et al., 2015). MeJA treatments also induce the biosynthesis of defensive compounds, such as alkaloids, phenolics, flavonoids, and terpenoids along with accumulation of peroxidase, chitinase, β-1,3-glucanase and PAL (Jia et al., 2016).
Wan et al. (2015) demonstrated that a minimal production of ROS and a timely elevation of antioxidative capacity were correlated with a high level of resistance in Chinese wild Vitis, while highly susceptible cultivar ‘Red Globe’ suffered massive infection and sustained ROS production due to relatively unchanged antioxidative activities, suggesting the importance of ROS response for the timely recognition and defense to B. cinerea.
Synthesis and secretion of pathogenesis-related proteins (PR) are also part of the inducible defense mechanism deployed by V. vinifera. For instance, up-regulation of PR-2 in less susceptible plants and accumulation of beta-1,3-glucanases have been described during infection (Derckel et al., 1999). A recent proteomic analysis on V. vinifera cell suspension inoculated with B. cinerea identified proteins involved in defense (e.g., PR10), response to oxidative stress, cell wall modification and protein folding (Dadakova et al., 2015). Besides, the transcripts of five defensin-like genes (DEFL) were identified as significantly up-regulated in grape tissues infected with B. cinerea and three novel defensins were proven to inhibit conidia germination (Giacomelli et al., 2012), suggesting a role of these genes in the defense against this pathogen.
A number of transcription factors have also been identified in V. vinifera with a significant role in the defense response against B. cinerea, whose functional orthologs were previously described in the plant model Arabidopsis thaliana. For example, induction of VvWRKY33 (Group I from WRKY protein family) correlates with the expression of VvPR10.1. Complementation of resistance-compromised Arabidopsis wrky33-1 mutant lines by the constitutive expression of VvWRKY33 restores the resistance against B. cinerea (Merz et al., 2015). Also, transcription factors from the NAC family are known to be involved in the response to environmental stresses, as VvNAC1 expression is induced in B. cinerea infected berries and leaves. Moreover, VvNAC1-overexpressing Arabidopsis plants exhibit enhanced resistance to this fungus. These plants present a modified expression pattern of defense-associated gene markers (AtPR-1, AtPDF1.2), suggesting that VvNAC1 could be a regulatory component of the plant signaling defense cascade (Le Henanff et al., 2013).
The defenses induced in grapevine berries against B. cinerea differ during berry ripening. Agudelo-Romero et al. (2015) described the putative involvement of JA/Et, polyamines and auxins, and evidence of a reprogramming of carbohydrate and lipid metabolisms toward increased synthesis of secondary metabolites involved in plant defense, such as trans-resveratrol and gallic acid, and that, contrary to healthy berries, infected green berries did not activate the SA pathway, suggesting that the pathogen is able to shut down some defenses.
On the other hand, inoculated veraison berries accumulate ROS, activate the salicylate-dependent defense pathway, synthesis of the resveratrol phytoalexin, cell wall reinforcement and formation of papillae underneath the appressoria, thus stopping further infection. As was mentioned above, mature grapevine berries are more susceptible than veraison berries. Infected mature berries activate the jasmonate-dependent pathway, which is unable to stop the fungal necrotrophic infection (Kelloniemi et al., 2015).
Grape genotypes vary in their resistance to infection, degree of fungal colonization and severity of disease. Low or no resistant phenotypes in most common table grape V. vinifera cultivars have been described, whereas high level of resistance has only been found in the species Muscadinia rotundifolia (V. rotundifolia), V. labrusca, and other grape hybrids. This resistance appears to be related to the number and thickness of epidermal and hypodermal cell layers and cuticle and wax contents (Gabler et al., 2003). However, no genetic studies have been conducted to date in these resistant plants.
To date, pre-existing or basal defenses seem to be an important part of the defense against B. cinerea, along with activation of inducible defense mechanisms mediated by SA or JA/Et pathways, depending on the development state, together with an appropriate kinetics between ROS production and generation of antioxidant compounds.
Biotrophic and Hemibiotrophic Pathogens
Erysiphe necator
The biotrophic fungus E. necator is the etiologic agent of the grapevine powdery mildew (PM; Figure 1), affecting species of the genus Vitis. This disease appears as a white-grayish powder on the surface of the infected tissue, mostly leaves and stems. This generates large production losses, reduced yields and fruit quality, mainly by a declining in the sugar content and acidity of the berries (Gadoury et al., 2001; Calonnec et al., 2004).
Infection Strategy
Erysiphe necator depends on its host for growth and development. For this, the conidium attaches to the tissue cells of the plant, allowing the formation of a primary germ tube that differentiates into a specialized infectious structure (i.e., appressorium), which generates a mechanical pressure in order to penetrate and invade the host cell. The successful invasion results in the haustorium formation, by which the fungus absorbs nutrients necessary to complete their life cycle. The haustorium facilitates the dynamic exchange of molecules: the fungus retrieves hexoses, amino acids, vitamins, and other nutrients from host cells, while at the same time secretes proteins to suppress host defenses (Qiu et al., 2015). Once this structure is established, secondary hyphae spread along the infected tissue and finally asexual reproductive bodies (i.e., conidiophores and conidia) emerge from them. When environmental or nutritional conditions become unfavorable, E. necator develops cleistothecia, structures of sexual reproduction, that contain four to six asci at maturity, each of which usually contains four ascospores. However, physiological maturity may not be reached for several months, particularly in colder climates. Like conidia, ascospores germinate with a single germ tube, which terminates in appressorium formation (Gadoury et al., 2012).
As in other obligate biotrophic pathogens, E. necator genome shows a reduced number of genes related to secondary metabolism (i.e., polyketide synthase, non-ribosomal peptide synthase, dimethylallyl tryptophan synthase and terpene synthase), nitrate and sulfate metabolism, and other core ascomycete genes, such as those related to amino acid metabolism, fermentation, channels/transporters and stress response, among others (Jones et al., 2014). Due to these gene losses, nutrient acquisition from the host is essential for E. necator development. Nitrate transporter VvNPF3.2 has shown to be up regulated in the susceptible grapevine cultivar ‘Cabernet sauvignon’ during PM infection, probably to increase nitrate or nitrite transport, whereas this was not observed in the resistant cultivar ‘Norton’ (Pike et al., 2014). It has also been described in PM-infected V. vinifera cv. ‘Cabernet sauvignon’ an increase in the abundance of proteins related to amino acid metabolism (i.e., Alanine aminotransferase and alanine glyoxylate aminotransferases), vitamin biosynthesis and lipid/sterol biosynthesis (Marsh et al., 2010).
In addition, PM exploits as well plant components for its successful penetration and establishment in the host cell. The MLO-like class of R-genes (Mildew Locus O, MLO, belonging to a family of seven-transmembrane domain proteins) is known to be required for successful host-cell invasion. Loss-of-function mlo alleles in barley and A. thaliana lead to enhanced resistance to adapted PM species (Freialdenhoven et al., 1996; Consonni et al., 2006). This family is composed by six members in grapevine (VvMLO3, VvMLO4, VvMLO6, VvMLO9, VvMLO13, and VvMLO17), but early induction of VvMLO3, VvMLO4, and VvMLO17 coincided with the establishment of fungal penetration in grape leaves (Feechan et al., 2009). Recently, the locus Sen1 (susceptibility to E. necator 1) mapped in V. vinifera cv. ‘Chardonnay’ was described as a novel source of PM susceptibility (Barba et al., 2014). Although the biological functions of these genetic components remains elusive, host genes products are likely to be modulated by PM for its successful infection.
Host Response and Resistance against E. necator
Wine and table grape V. vinifera cultivars are very susceptible to E. necator. Marsh et al. (2010) showed that susceptible plants (V. vinifera cv. ‘Cabernet sauvignon’) are able to initiate a basal defense response but unable to restrict fungal growth or reduce the progression of the disease. Differentially expressed proteins during the infection of E. necator are involved in photosynthesis, metabolism, protein destination, and protein synthesis; suggesting that the pathogen manipulates plant energy processes, according to its needs. Furthermore, defense proteins, such as PRs, defensin-like proteins (DEFLs) and proteins involved in ROS detoxification are induced, resembling a basal defense response, however, these mechanisms are inadequate in timing and/or intensity in order to control disease progression (Marsh et al., 2010).
Many species of the Vitis genus, mostly from North America, have varying levels of resistance to PM. Feechan et al. (2011) classified this resistance into four types: susceptibility, partial resistance, penetration resistance, and resistance associated with PCD, based on the degree of development of E. necator in host tissues and the triggered response. Among these, and as described above, V. vinifera is classified as a susceptible species, V. riparia as partially resistant and M. rotundifolia is among the most resistant species mainly due to the induction of PCD.
At the first level of resistance, PEN1- and PEN2/PEN3-like pathways [PENETRATION (PEN) genes] are important components of PTI in grapevine, suggesting a role against non-adapted PM. These genes were described in A. thaliana and their grapevines orthologs may have similar functions. AtPEN1 is a member of the SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor) family which includes proteins that mediate membrane fusion events; AtPEN2 is a myrosinase involved in the biosynthesis of antimicrobial molecules that are delivered to the site of PM penetration via AtPEN3, which is an ATP-binding cassette transporter (Qiu et al., 2015). Their role in PTI response remains elusive, however, it has been shown their importance and functionality in grapevines.
In a second level of resistance, different loci have been identified in several species of the Vitaceae family that confer resistance to E. necator: REN1 (Hoffmann et al., 2008); REN2 (Dalbó et al., 2001); REN3 (Welter et al., 2007); REN4 (Ramming et al., 2011); REN5 (Blanc et al., 2012); REN6, REN7 (Qiu et al., 2015); RUN1 (Barker et al., 2005); RUN2.1, and RUN2.2 (Riaz et al., 2011), although molecular defense mechanisms triggered by these resistance loci are yet poorly understood. Among them, the RUN1 locus of M. rotundifolia was mapped in chromosome 12 and the functional characterization of resistance gene analogs (RGA) within this locus identified a single gene (RGA10 or MrRUN1) that is associated to the complete resistance to PM (Feechan et al., 2013). This gene codes for a resistance protein of the Toll/interleukin-1 receptor-like (TIR)-NB-leucine-rich repeat (LRR) class of immune receptors, composed by a Toll/interleukin-1 receptor-like (TIR), a nucleotide-binding site (NBS) and a leucine-rich repeat (LRR) domains, acting on pathogen effector recognition and signal amplification (McHale et al., 2006). The other resistance loci have also been associated with this type of resistance gene functions; though this has not been experimentally verified to date. Amrine et al. (2015) sequenced and analyzed the transcriptomes of Central Asian grape accessions that were previously shown to carry a REN1-like local haplotype. They confirmed the partial resistance phenotype of these accessions, which is associated with a late post-penetration response to E. necator as it was previously described (Hoffmann et al., 2008; Coleman et al., 2009). In all Central Asian accessions, E. necator germinated and established primary hyphae, but it failed to colonize extensively the leaf tissues. Necrotic spots that colocalize with appressoria of secondary hyphae suggest that late HR and PCD may be key in slowing down fungal spread in the Central Asian accessions (Amrine et al., 2015).
Effective resistance responses of grapevine against E. necator include enhancement of both JA-mediated and systemic acquired resistance (SAR) responses and accumulation of phytoalexins. For example, transcriptomic changes in resistant Chinese wild grapes (V. pseudoreticulata) during PM-infection are related to SA and JA responses, SAR, HR, plant–pathogen interaction, flavonoid biosynthesis and plant hormone signal transduction (Weng et al., 2014). Up regulation of defense-related genes such as glycosyl hydrolases, lipases, PR-5 thaumatin-like proteins and proteinases, among others, has been described in successful response to E. necator. For example VpPR-10.1, isolated from V. pseudoreticulata, has nuclease activity (both RNase and DNase activities) and can lead to PCD and DNA degradation. A correlation between VpPR-10.1 levels and its antifungal property suggest that the nuclease activity is the biochemical basis for the resistance. Thus, VpPR-10.1 could potentially play a dual role degrading pathogen RNA and inducing programmed death of host cells (Xu et al., 2014). Also, the transcription factor VpWRKY1 was rapidly induced in V. pseudoreticulata after E. necator inoculation, and the expression level was found to be correlated with the resistance magnitude (Li et al., 2010). A number of other genes have also been implicated in PM resistance in certain wild Vitis species, showing differential expression levels between PM-resistant wild Vitis species and susceptible V. vinifera cultivars and conferring increased levels of resistance to PM when overexpressed transiently in grapevine leaves or stably transformed into wild-type or mutant lines of A. thaliana (Qiu et al., 2015). The key positive regulator of SA pathway, ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1), was differentially expressed in ‘Norton’ (V. aestivalis cv. ‘Norton’) and ‘Cabernet sauvignon,’ where a constitutive expression in the first and an induced in the latter were observed after inoculation with PM (Fung et al., 2008). This also correlates with the high and constitutive SA levels in ‘Norton’ resistant cultivar. Moreover, analysis of the EDS1 family in grapevine identified both EDS1 and EDL2 (EDS1-LIKE 2) in the resistant and susceptible cultivars as necessary components of the regulatory node EDS-PAD4 in a SA-mediated pathway (Gao et al., 2014).
Lignin, flavonoid phytoalexins, and phenolic compounds also play important roles in the defense response of grapes. The rapid production of resveratrol, major compound of the stilbene family and its transformation into viniferins appear to enhance PM resistance in grapevine cultivars. For example, in the resistant cultivar ‘Norton,’ STS are induced 24 h after inoculation with PM, accumulate higher levels of STS transcripts in mature leaves than the susceptible cultivar ‘Cabernet sauvignon’ and has a differential expression pattern during the berry formation compared to the susceptible cultivar, with a markedly increase at veraison (Dai et al., 2012).
As described above, the presence of resistance loci associated to R gene clusters would be crucial to define the susceptibility or resistance of grapevine plants against E. necator. This suggests that SA pathway is the main mechanism to lead a successful defense response in V. vinifera, since R – AVR signaling is mediated by this hormone. However, other hormones such as JA and the production of secondary metabolites in different cultivars may have a significant role in the defense response against this pathogen.
Plasmopara viticola
Plasmopara viticola (Berk. and Curt.) Berl. and De Toni is the causal agent of grapevine DM (Figure 1) and one of the most important pathogens affecting grapevine production worldwide. It was first collected in North East of the USA in 1834. Since then, it has been classified sequentially as the fungus B. cana, B. viticola, Peronospora viticola and finally as the obligate biotrophic oomycete P. viticola (Burruano, 2000; Gessler et al., 2011).
Plasmopara viticola spread to Europa by the year 1878, probably due to the use of cuttings from American grapes. All cultivated European V. vinifera cultivars are susceptible to P. viticola. Consequently, the production of V. vinifera has been affected by DM to the present. However, several North American Vitis species show resistance to this disease at different levels. This characteristic has also been found in Muscadinia species and some Asian Vitis species. Indeed, the use of natural sources of resistance to pathogens is still a promising tool in breeding programs nowadays (Gessler et al., 2011).
Infection Strategy
Plasmopara viticola infection occurs specifically through stomata. It starts early in the season, when oospores in fallen leaves or mycelium in dormant twigs are activated by adequate climate conditions to produce sporangia. In presence of water, mature sporangium releases self-motile biflagellate zoospores that infect plants tissues. Zoospores are able to place on the abaxial surface of leaves close to stomata, then germinate and penetrate through the stomatal cavity, where they form a substomatal vesicle. This vesicle gives rise to the primary hyphae and mycelium, which grows through intercellular spaces, enclosed by the veins of the leaf and enters to the cell of the mesophyll by its cell-wall-penetrating and feeding haustoria, which invaginates the plasma membrane of the parenchyma cells. As a result, the adaxial surface of the leaf exhibits a typical oil-spot lesion visible in plants affected by this pathogen. The mycelium also develops to form sporangiophores emerging from the stoma and releasing sporangia to the surrounding susceptible tissues (leaves, twigs, or grape clusters) or plants (Burruano, 2000; Perfect and Green, 2001; Gessler et al., 2011).
Stomata remain abnormally open and unresponsive to abscisic acid in grapevine leaves infected by DM. Recently, Guillier et al. (2015) identified two V. vinifera glycoproteins: a phototropin and a lysophospholipase that are induced by pathogen infection, accumulated in apoplastic fluids and considered as candidates for stomatal deregulation.
Little is known about genes involved in pathogenesis in P. viticola. During leaf infection, a gene encoding NADH-ubiquinone oxidoreductase has been indicated to be involved in the generation of the proton motive force to allow ATP biosynthesis and active nutrient transport before acquiring them from the host, mainly during the development of the first hyphal structures and haustoria. Analysis of ESTs from germinated zoospores has shown that the most represented unigene encodes a protein similar to fungal laccases, possibly contributing to pathogenesis by playing a role in stilbene detoxification. This could correspond to one of the mechanisms used by the pathogen to counterbalance the production of plant chemical arsenal. Also, an invertase-coding gene would be involved in pathogen proliferation in plant tissues by allowing P. viticola to uptake carbohydrates from the host (Mestre et al., 2012; Luis et al., 2013).
In cDNA-AFLP analysis carried out on infected grapevine leaves at oil spot stage, Polesani et al. (2008) identified nine transcripts derived fragments uniquely induced by infection, absent in sporangia, which represent putative virulence factors. In order to identify candidate genes encoding effectors from P. viticola several predicted secreted proteins have been identified. Genes corresponding to hydrolytic enzymes, protein inhibitors, elicitor-like proteins and members of the RXLR family of effectors are expressed upon infection. Also, the expression of genes coding for an INL11B-like elicitin, a protein with Kazal-like protease inhibitor fold and a RXLR protein, in infected tissues and germinated zoospores, has been described (Mestre et al., 2012).
Host Response and Resistance against P. viticola
Yu et al. (2012) established five immunity levels to DM in grapevine: (i) if accumulation of callose deposits around the stomata and inhibition of zoospores germination early in the infection process are observed; (ii) emergence of callose deposits around and in the stomata but unable to inhibit the formation of hyphae; (iii) with callose near the stomata and around haustoria, incapable to stop infection; (iv) with more development of hyphae than in (iii); and (v) with hyphae in all mesophyll tissue intercellular spaces. According to this classification, M. rotundifolia was classified as level (i), several Chinese species as level (iii), except for V. amurensis as level (iv) and some Chinese species and V. vinifera as level (v).
Liu et al. (2015) classified three Chinese wild Vitis genotypes according to the histological response to infection. V. piasezkii Liuba-8 was classified as ‘highly resistant,’ showing no sporulation, callose deposition and early production of H2O2, which corresponds to level (i), according to Yu et al. (2012) classification. On the other hand, V. pseudoreticulata Baihe-35-1 and V. davidii var. cyanocarpa Langao-5 showed decreased levels of sporulation and callose deposits in infected tissues, which could be related to level (ii) in Yu classification (Yu et al., 2012).
In grapevine, some QTLs with major DM-resistance effects have been identified, named Rpv (Resistance to P. viticola). Examples of these are Rpv2, derived from M. rotundifolia and located on chromosome 18 (Merdinoglu et al., 2003; Peressotti et al., 2010); Rpv3 found in ‘Bianca’ probably from a V. rupestris background and mapped on the reference chromosome 18 (Bellin et al., 2009; Casagrande et al., 2011; Di Gaspero et al., 2012); Rpv8, and Rpv12 located on chromosome 14 and Rpv10 located on chromosome 9, from V. amurensis (Blasi et al., 2011; Schwander et al., 2012; Venuti et al., 2013).
In the Rpv1 locus derived from M. rotundifolia and located on chromosome 12 (Merdinoglu et al., 2003), one of the first grapevine resistance genes to be cloned and functionally characterized was identified, designated MrRPV1 and conferring strong resistance to P. viticola with a broad isolate specificity described by the severe restriction of hyphal growth and sporangiophore development. MrRPV1 produces four alternatively spliced variants, encoding only one full-length functional TIR-NB-LRR protein (Feechan et al., 2013).
In compatible interactions between grapevine and P. viticola, analysis of differentially expressed grapevine genes during infection have demonstrated that most of these genes are down regulated and only 30% up regulated. Genes related to photosynthesis and primary carbon metabolism were the most negatively affected. Other down-regulated genes are related to protein metabolism, transport and signal transduction. Genes involved in secondary metabolism, defense and response to external stimuli were up regulated (Polesani et al., 2008). Among the genes related to defense response, expression of the PR-2, PR-4, OSM-1 (osmotin), GLP-2 (germin-like protein), GLP-7, TLP-4 (thaumatin-like proteins), CHI (chalcone isomerase), and NAC genes are induced in microdissected stomata and surrounding cells at early times of infection. Thus, there would be a site-specific regulation of grapevine response to P. viticola, with short distance signals released from stomata to adjacent cells (Lenzi et al., 2016).
Stress and defense-related genes have shown higher levels of expression in resistant grapevines, so it has been suggested to be linked to constitutive resistance against P. viticola. Also, high levels of inositol, caffeic acid, and other antimicrobial substances have been related to constitutive response in uninfected resistant leaves. Up regulation of genes coding for PR proteins occurs as soon as resistant grapevines are inoculated with P. viticola. Incompatible interactions are also associated to modulation of genes involved in defense response, photosynthesis, primary and secondary metabolism, signal transduction and transport. This is also followed by an accumulation of metabolites such as resveratrol and viniferins (Gessler et al., 2011; Malacarne et al., 2011).
In Vitis progeny analysis carried out by Malacarne et al. (2011), higher stilbenoid accumulation is associated to individuals with least severe DM symptoms. These compounds were trans-resveratrol, trans-piceid, trans-pterostilbene, and several viniferins. Among the genes induced in the incompatible interaction there are two receptor-like protein kinases, one TIR-NBS receptor, a calcium-dependent protein kinase, as well as transcripts concerning ethylene biosynthesis and responsive factors, PR1, PR2, PR5, PR10, two isoforms of STS, genes encoding a caffeoyl-CoA O-methyltransferase, a flavonoid 3′ 5′-hydroxylase and a dihydroflavonol reductase, among others (Malacarne et al., 2011).
The resistance level of grapevines has been correlated to the quantity of the callose deposition. In this context, strong callose deposition in the P. viticola resistant M. rotundifolia was observed, along with an increased expression of callose synthases genes CalS1 and CalS10 at early times of infection (Yu et al., 2012).
VvWRKY33 has been suggested to play an important role by inducing early defense-related pathways in grapevine leaves against fungal pathogens. VvWRKY33, a class I WRKY transcription factor, activates the promoter of the VvPR10.1 gene. Both are strongly induced when incompatible interaction occurs; in the case of VvWRKY33 just 2 h after P. viticola infection. Ectopic expression of VvWRKY33 in grapevine leaves of a susceptible grapevine cultivar has proved to strongly increase resistance to P. viticola, reducing sporulation between 50 and 70% (Merz et al., 2015).
Thus, as described in E. necator-grapevine pathosystem, some QTLs with major DM-resistance have been identified and would be crucial to define the susceptibility or resistance of grapevine plants against this pathogen. SA pathway, expression of related genes and also production of secondary metabolites may have a significant role in the defense response against this pathogen.
Agrobacterium vitis
Agrobacterium vitis is a specific pathogen of V. vinifera and is the causal agent of the grapevine crown gall disease (Figure 1). Virulent strains of this bacterium induce the formation of tumorigenic structures at the site of infection for nutrient uptake, while necrosis and a HR-like response has been reported in grapevine roots and in non-host plants infected with this bacterium, respectively. A. vitis can be defined as a biotrophic pathogen since it maintains a parasitic relationship with living tissues of their host to complete its life cycle.
Infection Strategy
Similar to Rhizobium radiobacter (formerly known as Agrobacterium tumefaciens), A. vitis contains Ti plasmids carrying vir genes and T-DNA sequences that allows it to transfer stable genetic material into the genome of its host, particularly genes involved in the biosynthesis of opines, auxins, cytokinins, and the utilization of tartrate (Burr and Otten, 1999). The infection starts commonly through plant injuries, particularly by freezing and/or mechanical wounds, to favor the release of phenolic compounds that act as chemoattractants to the attached bacteria. These compounds activate the transcription of vir genes, whose products induce the transfer of the T-DNA into the host genome. VirA is a protein that can detect and interact with the chemical signal and phosphorylate the VirG protein, which in turn transcriptionally activate several other vir genes. For instance, VirG activates the expression of virD2, whose product has the necessary endonuclease activity to release the T-DNA from the Ti plasmid, and virE2, involved in the protection and transport of the bacterial genetic material into the plant (Burr et al., 1998; Burr and Otten, 1999).
The genetic diversity of A. vitis is defined by the nature of their Ti plasmids and T-DNA sequences. Ti plasmids are commonly defined according to the type of opine they produce. In grape crown gall, three Ti plasmids have been defined as the octopine/cucumopine (O/C), the nopaline and vitopine plasmids (Szegedi et al., 1988). These three plasmids contain T-DNA sequences that can be categorized into two functional groups, whether are involved in opine synthesis or in gall induction. Opines are low molecular weight compounds derived from aminoacids, keto-acids, and sugar phosphates that only can be catabolized by Agrobacterium strains (Escobar and Dandekar, 2003). T-DNAs contain genes for the synthesis of these molecules, such as acs (agropine synthase), cus (cucumopine synthase), nos (nopaline synthase), and/or vis (vitopine synthase) depending on the nature of the T-DNA. Moreover, T-DNA sequences also contain genes involved in tumor formation, where genes involved in the biosynthesis of auxins (iaaM and iaaH) and citokinins (ipt genes) can be found (Burr et al., 1998). Oncogenes are also present in these regions, such as the gene 6b that encodes a protein that stimulates cell growth in an auxin-independent manner. For example, the overexpression of T-6b in transgenic tobacco plants leads to an increased cell expansion in leaf disks but with no apparent changes in auxin content (Clement et al., 2006).
A hallmark of the genetic structure of A. vitis is the presence of plasmids with genes encoding proteins involved in tartrate utilization (pTr), an abundant compound in grapevines. Three types of these plasmids have been found in A. vitis and were defined as TAR-I and TAR-II from the AB3 strain, and TAR-III from the AB4 strain (Burr and Otten, 1999). Although there are differences in the size of these plasmids, the genes associated with tartrate utilization (ttuA-E genes) are conserved among the strains of A. vitis. It is suggested that TtuA act as a transcription factor that induce the expression of other ttu genes such as ttuB, which is involved in the tartrate entry into the bacteria; ttuC and ttuD, whose product catalyze the degradation of tartrate; and ttuE, which codes a tartrate-inducible pyruvate kinase that seems not essential for the utilization of tartrate (Salomone et al., 1996). It is important to notice that the particular characteristic of A. vitis to utilize a grape-derived compound can partially explain the high host specificity of this pathogen.
Host Response and Resistance against A. vitis
Natural resistance sources against the grape crown gall are important for the study of the interaction between tumor-forming pathogenic bacteria and its host, and for the introduction of these sources into further breeding programs. Some examples of both cultivated and wild Vitis species are V. riparia cv. ‘Glorie de Montpellier’ and V. amurensis, respectively, who are resistant to the infection of A. vitis. In the case of V. riparia, it has been suggested that the T-DNA from A. vitis is not stably integrated into the host genome given by the low frequency of T-DNA integration and the absence of opine content in infected tissues (Süle et al., 1994). In V. amurensis, a dominant locus has been identified in the chromosome 15 of this specie (Rcg1 locus), where no signs of crown gall formation were observed on populations carrying this resistant locus (Kuczmog et al., 2012). Even though this resistance seems to affect a wider range of Agrobacterium species, the molecular basis underlying the Rcg1 locus remains unclear.
The Korean cultivar V. vinifera cv. ‘Tamnara,’ originated from a grape breeding program between ‘Campbell Early’ (V. labruscana) and ‘Himrod Seedless’ (Vitis sp.; Park et al., 2004), also showed resistance to the grapevine crown gall. Interestingly, genes related to defense responses in Tamnara were expressed in both inoculations with A. vitis and SA-treatments, categorized in groups of ESTs whose genes encode proteins involved in defense (i.e., glucanase, chitinase, thaumatin-like protein, PR-10), signal transduction (NBS-LRR type protein), oxidative burst (i.e., GST, CAT, Glutathione peroxidase) and cell wall fortification (i.e., hydroxyproline-rich protein, extensin-like protein, cinnamyl alcohol dehydrogenase), among others (Choi et al., 2010). It might not be unexpected to find a group of defense related genes that correlate A. vitis inoculations and SA treatments in resistant cultivars, since it has been described that SA and SAR have a role in the defense against R. radiobacter in N. benthamiana, where pre-treatments with SA interferes with the growth and virulence of this bacteria on its host and might affect the expression of several vir genes (Anand et al., 2008).
Thus, it seems that the SA signaling pathway may be operating in the interaction between A. vitis and resistant cultivars of V. vinifera, inducing a defense response similar to those against other biotrophic pathogens mentioned above.
Xylella fastidiosa
Xylella fastidiosa is a gram-negative, xylem-limited bacterium and the causal agent of Pierce’s disease (PD) in V. vinifera (Figure 1). This bacterium can be classified as a biotrophic pathogen, because it does not kill the host tissue until later stages of its life cycle. The growth of this bacterium depends mostly on climate. The optimum growth conditions for X. fastidiosa are warmer environments close to 28°C, this is the main reason that this pathogen is not prevalent in areas where winter temperature drops under 0°C (Lieth et al., 2011).
Infection Strategy
Xylella fastidiosa is transmitted to new host plants by insect vectors such as sharpshooters (Homalodisca coagulate), leafhoppers (Cicadellidae family) and spittlebugs (superfamily Cercopoidea) during xylem sap feeding. This is the major cause for this pathogen to spread and infect several agriculturally important plants, such as citrus, almond, coffee, and grapevine (Newman et al., 2003). In V. vinifera, X. fastidiosa causes the Pierce’s disease (PD), an economically important disease that affects wine, table and raisin grape production. The symptoms of this disease can be severe, including leaf scorching, desiccated fruit, cordon die back, and finally vine death (Roper et al., 2007).
Xylella fastidiosa infects the vine creating a biofilm in xylem vessels that disrupt water and nutrients flow throughout it. This occlusion is composed by host gums, bacterial exopolysaccharide or degradation products from the host cell wall, or a combination of the three. The xylem is a water transport network of vessels composed of lignified dead cells. The vessels are interconnected by bordered pits (channels) and the pit membranes are composed of pectin, cellulose, hemicellulose and proteins. Pectin polymers determine the pore size in the pit membrane, being sizes between 5 nm in grapevine, allowing the passage of sap through xylem but blocking the passage of larger objects, like X. fastidiosa, whose diameter range from 0.3 to 0.5 μm (Davis et al., 1978; Newman et al., 2003; Roper et al., 2007).
Pierce’s disease biofilm formation can be divided in three stages: early biofilm formation, growth, and adaptation. The first stage starts with the transmission of X. fastidiosa directly by insect vectors into the xylem vessel. After entry, X. fastidiosa needs to move through the vascular system. PilT, a type IV fimbriae, is responsible for retracting and extending fimbriae IV in a twitching motility, allowing the bacteria to move through the vascular system. These processes depend on cell–cell aggregation, which involves membrane attachment proteins, such as chaperonin GroEL, and two-component system regulatory proteins (PopP, FeuP, or PhoP) implicated in the correct adhesion of bacterium-bacterium and biofilm formation (Silva et al., 2011). Following this, the bacteria start to break down xylem pit membranes. In order to do this, they secrete CWDs enzymes, such as endo-1,4-β-glucanases, endoxylanases, β-xylosidases, cellobiohydrolase, and a polygalacturonase (PG). Bacterium needs PG to successfully infect and is a critical virulence factor to promote pathogenesis. This enzyme digests cell wall polymers that form the xylem pit membrane. The digestion of pectin component exposes the cell-wall polysaccharides that are target by the other CWDs enzymes. Thus, the pectin digestion allows X. fastidiosa both to use as a carbon source and to move from vessels to vessels (Roper et al., 2007).
The second stage involves a group of metabolic processes to favor biofilm growth, such as amino acid and carbon metabolism and cofactors biosynthesis. Furthermore, at this stage the phenomenon called quorum-sensing (QS) occurs; a bacterial mechanism to sense the population and react to their changing environment. This mechanism is regulated by acylated homoserine lactones (AHLs), which are molecules that trigger the expression of QS regulated genes. QS is involved in the production of cis-11-methyl-2-dodecenoic acid, which participates in virulence, motility, toxin production, aerobic respiration, biofilm dispersal and extracellular polysaccharide (EPS). Another important enzyme for this stage is a polynucleotide phosphorylase that degrades plant chloroplasts to provide carbon sources and facilitate the migration through xylem vessels (Roper et al., 2007).
The third and last stage corresponds to adaptation and protection, involving proteins which confer advantages to the bacterial population like resistance to environmental factors. Within this context, toxin production and heat shock protein (sHsp) expression have been observed. sHsp are a small stress induced proteins that prevent aggregation and are important in protein refolding. A predicted membrane localized GTPase was also found, which may be involved in a signal transduction mechanism, but their specific role is to date unclear. Finally, the two-component system regulatory protein (popP, feuP, or phoP) is required for virulence at the last and long stage of infection (Silva et al., 2011).
Host Response and Resistance against Xylella fastidiosa
The immune response and molecular mechanisms that V. vinifera has developed against infection of X. fastidiosa remain unclear. However, there are some species, such as V. labrusca or M. rotundifolia that when infected with X. fastidiosa, their symptoms are not as severe as in V. vinifera (Lieth et al., 2011). It has been shown that vines employ physical or chemical barriers post infection, such as tylose and gums. Tylose is an outgrowth of parenchyma cells through vessel pit pairs into the lumen of tracheary elements (Sun et al., 2006) and gums are originated from the cell wall or middle lamella (Fry and Milholland, 1990). Some authors suggest that the development of tylose and gums around the infection site is a resistance mechanism. Sun et al. (2006) found that tylose initiation occurred too slowly to restrict spread of mobile pathogens such as X. fastidiosa. However, since this bacterium has a slow basipetal movement in systemic infection, tylose could represent a significant obstacle to this type of infection. Fry and Milholland (1990) studied different species of the Vitis: V. vinifera genus, which is very susceptible to X. fastidiosa, and muscadine grapevine (V. rotundfolia), whose cultivars are tolerant to PD. The authors demonstrated that the susceptible V. vinifera cv. ‘French Colombard’ generates predominantly occlusion pectin, whereas the tolerant V. rotundfolia cv. ‘Carlos’ and ‘Noble’ generate occlusion tylose and gums. Thus, the synthesis of tylose and gums could serve as a defense response to contain the infection of X. fastidiosa.
Mapping for resistance traits against PD has been reported in hybrids of Mexican cultivars of V. arizonica. Within these populations a major QTL has been identified, called PdR1 (Pierce’s disease Resistance 1) in the linkage group 14, which explains 72% of the phenotypic variation (Krivanek et al., 2006; Riaz et al., 2006). Plants carrying this locus have lower levels of bacterial titer in the stems. Moreover, a fine-scale mapping identified two allelic forms in this locus, denominated PdR1a and PdR1b, derived from full siblings of a PD resistant population (V. rupestris ‘A. de Serres’ × V. amazonica/candicans ‘b43-17’). Within this region, 14 putative genes have been identified and 5 of them were annotated with a molecular function (Riaz et al., 2008). Thus, it is yet to be proved whether PdR1a and PdR1b have different genes within this locus or two alleles of the same gene. A subtractive suppression hybridization (SSH) analysis was performed in leaf, stem, and shoot tissues between resistant and susceptible sibling genotypes (V. rupestris × V. amazonica) after inoculation with X. fastidiosa (Lin et al., 2007). Among these tissues, ESTs derived from resistance genotypes where associated with primary cell wall modifying and metabolic enzymes and PR proteins, such as endo-xyloglucan transglycosylase (EXT), xyloglucan endotransglycosylase (XET), PR-2 (β-1,3-glucanase) and PR-10, among others.
To date, identification of resistance locus to PD, called PdR1, and physical or chemical barriers such as tylose and gums are the most relevant known defense mechanism of grapevine to X. fastidiosa.
Viruses
Nearly 70 virus species have been identified to date that are able to infect the Vitis genus, accounting for at least 25 different diseases in grapevine (Martelli, 2014). As other viruses, they are classified by the International Committee on Taxonomy of Viruses (ICTV), classification that is based on several parameters such as: size particle, genome structure (ORFs), nucleotide or amino acid sequence identity of different virus proteins, like heat shock protein 70 (HSP70), polymerases, coat proteins (CP), type of transmission vector and serological information. It should be noted that in many cases they are found associated as a multiplex virus infection complex (Prosser et al., 2007; Martelli, 2014).
From an economic point of view, the most important grapevine viruses are those who cause the leafroll diseases, known as Grapevine Leaf Roll associated Viruses (GLRaV -1, -2, -3, -4, and -7). Moreover, several genetic variants of GLRaV-4 and GLRaV-6 have been described, as well as GLRaV-Pr, GLRaV-Car, and GLRaV-De, based on phylogenetic relationships with the HSP70h gene (Maliogka et al., 2009; Ghanem-Sabanadzovic et al., 2010, 2012). All of the above belong to the Ampelovirus genus, which harbor a monopartite genome composed of genomic and subgenomics positive (+) single stranded (ss) RNAs with around 12 open reading frames. Two exceptions to the rule have been described: GLRaV-2, classified as a Closterovirus, and GLRaV-7, which has not been assigned to any genus to date (Al Rwahnih et al., 2012a; Naidu et al., 2015).
Other important grape viruses are the Grapevine Virus A and B (GVA and GVB, respectively), belonging to the Vitivirus genus, a monopartite genome composed of genomic and subgenomics (+)ssRNAs with five open reading frames (Adams et al., 2004). GVA is related to the Kober stem grooving symptom (Goszczynski et al., 2008; Alabi et al., 2014) and can cause reddening of leaf margins and petioles, poor vigor and leafroll; while GVB is associated with the corky bark syndrome consisting of soft, rubbery, and abnormal swelling of the basal internodes of the canes, longitudinal cracks and cork forming, typical of the rugose wood complex (Bonavia et al., 1996). Vitivirus genus has other less ubiquitous species: GVD, GVE, and most the recently discovered GVF, causing similar corky rugose wood-like symptoms (Al Rwahnih et al., 2012b).
On the other hand, the most representative species of grape viruses in the Nepovirus genus are the grapevine fanleaf virus (GFLV) and the arabis mosaic virus (ArMV), both causing typical fan-leaf degeneration and leaf decline, reduced fruit quality, short internodes, and abnormal bifurcations. In addition, GFLV is considered to be the major threat to grape industry due to its ability to reduce crop yield up to 80% depending on the isolate, the susceptibility of the grapevine variety and environmental factors (Martelli, 2014).
Not only ssRNA viruses can infect grapes. Al Rwahnih et al. (2013) described a monopartite circular ssDNA virus which belongs to an evolutionarily distinct lineage of the Geminiviridae family, the grapevine red blotch-associated virus (GRBaV). This virus causes symptoms such as red blotch, marginal reddening, and red vein in leaves and reduced sugar accumulation in fruits of V. vinifera cv. ‘Californian Cabernet Franc,’ ‘Cabernet sauvignon’ and ‘Zinfandel.’ Similarly, a new ssDNA virus from Brazil was described which is able to infect pears, apples and different grapes varieties causing symptoms such as shrinkage, reddening or red blistering of leaves (Basso et al., 2015).
Infection Strategy
Virus infection starts with the entry of the virus into the plant cell through wounds or graftings, since they cannot cross the cell wall by themselves. In several cases, the spread occurs by transmission vectors, like mealybugs (Pseudococcidae family) and dagger nematodes (Xiphinema index, X. italiae, and X. diversicaudatum among others), which feed on sap phloem or roots, allowing movement of these pathogens into the plant. Also, due to grafting techniques, viruses can flow from an infected bottom (rootstock) to top (scion) through the phloem (Martelli, 2014). Once inside the cell, they must disassemble in order to release their own genome and replicate. Near 70% of plant viruses genomes, including grape-associated ones, are composed of a positive single stranded RNA molecule, or ss(+)RNA. Thus, replication process starts with the expression of a viral replicase, generating negative RNA strands and then subsequently, new positive strands through the activity of a viral RNA dependent RNA polymerase (RdRp; Zabel et al., 1976; Dorssers et al., 1984).
Early translated proteins are involved in the replication process, such as polymerases, helicases, and proteinases. Then, structural proteins such as capsid (CP) and movement proteins (MP) are transcribed, with different functions during viral lifecycle. For example, MP and/or CP are necessary for viral translocation, while CP proteins are also required for virus assembly and cell-to-cell movement (Lucas, 2006). MP proteins can target plasmodesmatas and actively use the host actin/endoplasmic reticulum network to increase the size exclusion limit (SEL) of this structure by either destroying actin filaments and allowing virions to move between cells (Wolf et al., 1989; Waigmann et al., 1994; Harries et al., 2010; Niehl and Heinlein, 2011) or removing desmotubules and expanding membrane pores converting them to tubules (Kawakami et al., 2004; Liu et al., 2005; Schoelz et al., 2011). GFLV replication process occurs in viral compartments from endoplasmic reticulum-derived membranes, and it cannot only can be transported inside Golgi-derived vesicles through microtubule- or microfilament-dependent pathways (Laporte et al., 2003), but they can interact with type-I plasmodesmata located proteins (PDLP) too; these proteins are located on the tubule base of modified plasmodesmatas and genetic disruptions of this interaction can reduce tubule formation, delay infection, and attenuate symptoms (Amari et al., 2010). Nevertheless, not only PDLPs are involved in grape virus movement; class XI myosins (XI-K and XI-2) inactivation produced a mislocalization of PDLP and MP, suppressing GFLV movement, suggesting that these proteins could facilitate the necessary tubule formation for virus translocation (Amari et al., 2011). However, virus translocation is not dependent on virion assembly. This type of cell-to-cell movement can be divided into three steps: first, genome transfer from replication sites to intracellular transport systems; then, assisted intracellular transport of the vira1 genome and finally, the intercellular transport of genomes through plasmodesmata-TMV-like mechanisms (Carrington et al., 1996).
Finally, in phloem-restricted viruses (Figure 1), particles must translocate throughout the plant tissues, moving from mesophyll cells to sieve elements and thus achieve a systemic infection. Viruses take advantage of the phloem vasculature and sink-to-source flux in order to passively move through the plant at a very fast rate (Leisner and Turgeon, 1993). Although many viral determinants have been proposed depending on the viral genus, CP and MP seem to play a major role in this long-distance movement (Hipper et al., 2013). In the end, they are released into systemic tissues, thus starting new infection sites.
Host Response and Resistance against Viral Infection
The common plant cell response to viral infection is the activation of HR, including cell wall fortification and induction of PCD to block the systemic propagation of the virus (Stange et al., 2008; Komatsu et al., 2010). This response has been associated to an R gene-mediated immunity (i.e., ETI), through recognition of AVRs by this class of host immune receptors (Jones and Dangl, 2006). Nevertheless, this kind of response has not been reported within Vitis genus and no direct resistance gene has been described (Martelli, 2014).
During compatible grapevine–virus interactions, host transcriptional reprogramming in both leaves and berries has been reported. For example, presence of GRLaV-3 in V. vinifera cv. ‘Carmenere’ was correlated with leaf red coloration and chlorosis. Transcriptomic and developmental changes were studied during fruit ripening in V. vinifera cv. ‘Cabernet sauvignon’ infected with GRLaV-3. Reduction in sugar and total anthocyanins content, followed by differentially expressed genes, were observed during grape–virus compatible interaction. Among repressed genes, those associated with sugar metabolism, transport, and the phenylpropanoid pathway were severely inhibited at the ripening stage (E-L38), while genes associated to cell rescue, defense, death and aging were up-regulated (Vega et al., 2011). Similar results were found by Espinoza et al. (2007a,b) were same cellular responses plus proteasome-ubiquitin pathway and other biological processes as transport, transcription and RNA processing were induced during viral infections. Moreover, in V. vinifera cv. ‘Merlot’ infected with GRLaV-3, there is an induction of genes related to the flavonoid biosynthetic pathway, specifically the anthocyanin branch, which contributes with the reddish-purple leaf coloration phenotype in GRLaV-3-infected grapevines (Gutha et al., 2010). The same author showed that accumulation of anthocyanins, flavonols and proanthocyanidins are correlated with the induction of chalcone synthase (CHS), flavonoid-3′-hydroxylase (F3′H), flavonoid-3′, 5′-hydroxylase (F3′5′H), dihydroflavonol reductase (DFR), leucoanthocyanidin dioxygenase (LDOX), UDP-glucose:flavonoid 3-O-glycosyltransferase (UFGT) and the transcription factor MYBA1.
A physiological, agronomical and transcriptomic analysis of the infection of V. vinifera cv. ‘Bosco’ by GRSPaV (Grapevine rupestris stem pitting associated-virus) has been conducted by Gambino et al. (2012). At veraison (E-L35), infected grape leaves showed a decrease in their chlorophyll content and net photosynthesis, although up-regulation in photosynthesis, hormones, and secondary metabolism-related genes was observed. However, defense, signal transduction and primary metabolism (TCA, glycolysis and pentose phosphate pathway) related genes were down regulated in this infected tissue. In the same study, GRSPaV-infected berries at veraison showed that 233 genes were differentially expressed, showing down regulation of main functional categories (i.e., stress, disease, senescence, ROS detoxification, ethylene and JA signaling). It is believed that GRSPaV and grapevine have co-existed for a long period of time and these changes account as a form of adaptation and co-evolution between these two species, particularly in the inactivation of stress and defense responses and the favoring of the induction of photosynthesis associated genes.
Infected hosts are able to trigger molecular changes during compatible interactions in order to restrain virus infections. Plants deploy two main endogenous mechanisms to regulate gene expression: transcriptional gene silencing (TGS), which involves decreased RNA synthesis because of promoter methylation (Meyer et al., 1993; Sijen et al., 2001) and post-transcriptional gene silencing (PTGS; de Carvalho et al., 1992; Van Blokland et al., 1994) that recognizes dsRNA as a target (Hammond et al., 2001), which are then trimmed by type-III RNAses (i.e., DICER), producing small RNA fragments (i.e., siRNA). These short fragments act as a signal for a complementary sequence RNA that can be degraded at the end of the cycle. TGS and PTGS were considered as separate pathways. TGS was thought to regulate mainly transposons and accidental transgenes that mimic transposons. By contrast, PTGS was thought to regulate viral infection and accidental transgenes that encode some types of aberrant RNA that mimic viral RNA. The major breakthrough in the distinction came with the discovery that viruses and transgenes encoding dsRNA induce either TGS or PTGS (Vaucheret and Fagard, 2001).
As such, host PTGS machinery might be able to recognize viral dsRNA as a surveillance mechanism to cope with viral infections. For this reason, efforts have been made in order to obtain viral induced silencing transgenic lines expressing sense, antisense or both viral sequences in grapevines. For instance, V. rupestris and 110 Richter rootstock were successfully transformed with a short CP sequence using embryogenic cultures (Krastanova et al., 1995). Another transformations involved sense and antisense-oriented CP gene sequences from GFLV and GLRaV-3, a truncated HSP90-related gene of GLRaV-3 in rootstocks Couderc 3309, V. riparia, Teleki 5C, Millardet et De Grasset 101-14, and 110 Richter (Xue et al., 1999), and the GVA MP sense or antisense sequence expressed in V. rupestris plantlets (Martinelli et al., 2002).
However, virus can counterdefense plant response against infections in at least two known ways: RNAi viral suppressor proteins (VSRs) and RNA silencing suppressors (RSSs; Li and Ding, 2006; Ding and Voinnet, 2007; Murray et al., 2013). In either case, viruses can block the RNA silencing pathway at different stages (Burgyan and Havelda, 2011), affecting dsRNA processing and silencing signal amplification, decreasing siRNA stabilization, suppressing RISC activity or acting as suppressors with unspecified functions (Alvarado and Scholthof, 2009). For example, GLRaV-2 p21 protein suppresses PTGS through direct binding to siRNAs (Ye and Patel, 2005). Also, GLRaV-2 p24 protein and GVA p10 protein have been described to possess silencing suppressor activity (Chiba et al., 2006; Zhou et al., 2006). Another way to counterdefense plant response are protein domains encoded by RNA viruses which decrease alkylation damage. Van den Born et al. (2008) demonstrated that an Alk-B domain encoded by five viruses from the Flexiviridae family, is able to demethylate RNA with a robust repair activity due to its N-terminal extensions beyond the Alk-B core. Several other enzymes are also involved in viral RNAi suppression. Proteinase1/Helper component-proteinase (P1/HC-Pro) acts as a suppressor of both transgene induced and virus-induced gene silencing in tobacco plants (Anandalakshmi et al., 1998). Also, a tandem GLRaV-2 papain-like leader proteases, L1 and L2, are essential to establish the infection process of GLRaV-2 in initially inoculated grapevine (Liu et al., 2009).
Because viruses can affect plant post-transcriptional gene silencing mechanisms, some authors worked on a way to attack this viral counterdefense response. Soler et al. (2012) found that when three VSRs: p25, p20, and p23 from citrus tristeza virus (CTV) are silenced, complete resistance to viral infection in some transgenic lines were obtained. Thus, this strategy could be a promising approach to induce virus resistance in grapevine.
To date, identification of resistance genes has not been reported within Vitis genus, however, virus defense on plants is a common mechanism involving different steps on regulatory pathways. Future research for a deeper understanding of these mechanisms in grapes and other species is required.
Disease Management Outlook
Disease diagnosis is a time-consuming laborious task, especially regarding the identification of a particular pathogen and selection of the best control strategy to prevent or reduce economic losses. However, concerning grapevine disease management, some strategies available are very expensive and/or not tolerated by the importing markets, mainly due to their potentially negative environmental impact.
Since the vast majority of V. vinifera cultivars are susceptible to the pathogens abovementioned in this review, wine and fresh-grape production depends greatly on the use of effective control methods. Cultural control techniques, including thinning and pruning, “cold curing” (i.e., exposing plants to low temperatures to prevent or eliminate the pathogen), chemical control including contact and systemic compounds, biological control and natural products have been included in management systems. Also, in some grapevine disease models applied to decision support systems, other variables have been correlated with the pathogen life cycle, as meteorological conditions and phenological stage of the plant to reduce the use of pesticides applications and increase management efficiency (Gessler et al., 2011; Lieth et al., 2011; Kamoun et al., 2015).
Among the most effective methods, chemical control is one of the most used strategies. Based on the type of chemical (active ingredient), “pesticides” can be classified as copper compounds, sulfur compounds, dithiocarbamates, benzimidazoles, antibiotics, among others. Although these chemicals are effective against fungal and bacterial pathogens, they are not effective against viral diseases. Despite this, certain highly volatile chemicals compounds are toxic for organisms including insects, common viral vectors. Nevertheless, this technique is not highly effective and cannot avoid future infections. For this reason, management control programs nowadays use healthy mother plants insuring virus-free next generation plants (Almeida et al., 2013). Another strategy is to use silencing sequences of suppressor viral proteins that have the potential to avoid future viral infections. For example, the first attempts to artificially induce RNA silencing pathways in grapevines has been done in transgenic V. vinifera cv. “Chardonnay” expressing artificial miRNA targeting GFLV CP (Jelly et al., 2012).
Currently, main concerns in disease management include the environmental and health impact of chemical compounds. Biological control of grapevine and plant diseases in general has become more significant, especially against fungi as B. cinerea, by using antagonistic microorganisms before or after infection (Calvo-Garrido et al., 2014; Parafati et al., 2015). However, the use of disease-resistant cultivars of V. vinifera is the most cost-effective, safe, and environmentally desirable option, since resistant cultivars sometimes offer the only practical control option available. The identification and inclusion of natural sources of resistance in grapevine breeding programs by pyramiding two or more resistance genes or loci, looking for a sustainable viticulture in the future is one of the most commonly used practices. Some of these plants are used as rootstocks, since some plants have substantial levels of resistance, but are not of commercial interest. However, a few Vitis species have been found to be resistant to these pathogens, nevertheless, the source of this resistance has not been completely identified.
The use of biotechnology, for example in the generation of genetically modified plants carrying these resistance elements, could be an effective strategy accepted in the future in agriculture. New molecular approaches are being developed, suggesting that the editing of cis-regulatory elements (CRE) in defense related genes could be a new target in order to increase pathogen resistance (Swinnen et al., 2016). All the aforementioned are excellent candidates to use with novel genome editing tools, such as CRISPR/Cas9being introduced for crop plants with promising results. Big data approaches are also being develop for plant-pathogen interactions in order to facilitate information accessibility for crop developers. Examples as the PhytoPath (Pedro et al., 2016), which provides access to all plant pathogen genomes submitted to the International Nucleotide Sequence Database Nucleotide Consortium are of great significance. It would be an advantage to include this type of approaches for the development of resistant grape cultivars with minimal genome modifications.
At present, integrated disease management, using combined methods to eliminate or reduce pathogen threats and negative impact on the environment and health is a partially effective, economical, and sustainable way to cope with these problems.
Concluding Remarks
Understanding plant responses to different kind of pathogens may show some light on the co-evolution of plant and pathogen strategies, and their impact on resistance or susceptibility to infections.
The study of plant–pathogen interactions in crop plants, like grapevine, is essential to understand how pathogens infect the plant and how plant defenses are activated and enhanced. The understanding of these processes allows looking for biotechnological applications to reduce the economic losses associated to these microorganisms. It is crucial to reduce the handling and treatment with chemical agents, but it is necessary to go deeply into the molecular processes that underlie these interactions in order to achieve this.
Author Contributions
GA, RS, MA, DM, CN, and PA-J designed the work; made the work drafting, did data collection and reviewed it critically for the final version. RS designed the figure.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was partially supported by the National Commission for Science and Technology CONICYT (FONDECYT grant number 1150220) and the Millennium Nucleus for Plant Synthetic Biology and Systems Biology NC130030. GA was supported by FONDECYT postdoctoral grant number 3140324. We thank Dr. F. Restovic (Departamento de Genética Molecular y Microbiología, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile) for helping with the final manuscript revision.
References
Adams, M. J., Antoniw, J. F., Bar-Joseph, M., Brunt, A. A., Candresse, T., Foster, G. D., et al. (2004). The new plant virus family Flexiviridae and assessment of molecular criteria for species demarcation. Arch. Virol. 149, 1045–1060. doi: 10.1007/s00705-004-0304-0
Agudelo-Romero, P., Erban, A., Rego, C., Carbonell-Bejerano, P., Nascimento, T., Sousa, L., et al. (2015). Transcriptome and metabolome reprogramming in Vitis vinifera cv. Trincadeira berries upon infection with Botrytis cinerea. J. Exp. Bot. 66, 1769–1785. doi: 10.1093/jxb/eru517
Al Rwahnih, M., Dave, A., Anderson, M. M., Rowhani, A., Uyemoto, J. K., and Sudarshana, M. R. (2013). Association of a DNA virus with grapevines affected by red blotch disease in California. Phytopathology 103, 1069–1076. doi: 10.1094/PHYTO-10-12-0253-R
Al Rwahnih, M., Dolja, V. V., Daubert, S., Koonin, E. V., and Rowhani, A. (2012a). Genomic and biological analysis of Grapevine leafroll-associated virus 7 reveals a possible new genus within the family Closteroviridae. Virus Res. 163, 302–309. doi: 10.1016/j.virusres.2011.10.018
Al Rwahnih, M., Sudarshana, M. R., Uyemoto, J. K., and Rowhani, A. (2012b). Complete genome sequence of a novel vitivirus isolated from grapevine. J. Virol. 86, 9545. doi: 10.1128/JVI.01444-12
Alabi, O. J., Al Rwahnih, M., Mekuria, T. A., and Naidu, R. A. (2014). Genetic diversity of Grapevine virus A in Washington and California vineyards. Phytopathology 104, 548–560. doi: 10.1094/PHYTO-06-13-0179-R
Almeida, R. P., Daane, K. M., Bell, V. A., Blaisdell, G. K., Cooper, M. L., Herrbach, E., et al. (2013). Ecology and management of grapevine leafroll disease. Front. Microbiol. 4:94. doi: 10.3389/fmicb.2013.00094
Alvarado, V., and Scholthof, H. B. (2009). Plant responses against invasive nucleic acids: RNA silencing and its suppression by plant viral pathogens. Semin. Cell Dev. Biol. 20, 1032–1040. doi: 10.1016/j.semcdb.2009.06.001
Amari, K., Boutant, E., Hofmann, C., Schmitt-Keichinger, C., Fernandez-Calvino, L., Didier, P., et al. (2010). A family of plasmodesmal proteins with receptor-like properties for plant viral movement proteins. PLoS Pathog. 6:e1001119. doi: 10.1371/journal.ppat.1001119
Amari, K., Lerich, A., Schmitt-Keichinger, C., Dolja, V. V., and Ritzenthaler, C. (2011). Tubule-guided cell-to-cell movement of a plant virus requires class XI myosin motors. PLoS Pathog. 7:e1002327. doi: 10.1371/journal.ppat.1002327
Amrine, K. C., Blanco-Ulate, B., Riaz, S., Pap, D., Jones, L., Figueroa-Balderas, R., et al. (2015). Comparative transcriptomics of Central Asian Vitis vinifera accessions reveals distinct defense strategies against powdery mildew. Hortic. Res. 2, 15037. doi: 10.1038/hortres.2015.37
Anand, A., Uppalapati, S. R., Ryu, C. M., Allen, S. N., Kang, L., Tang, Y., et al. (2008). Salicylic acid and systemic acquired resistance play a role in attenuating crown gall disease caused by Agrobacterium tumefaciens. Plant Physiol. 146, 703–715. doi: 10.1104/pp.107.111302
Anandalakshmi, R., Pruss, G. J., Ge, X., Marathe, R., Mallory, A. C., Smith, T. H., et al. (1998). A viral suppressor of gene silencing in plants. Proc. Natl. Acad. Sci. U.S.A. 95, 13079–13084. doi: 10.1073/pnas.95.22.13079
Aziz, A., Poinssot, B., Daire, X., Adrian, M., Bezier, A., Lambert, B., et al. (2003). Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant Microbe Interact. 16, 1118–1128. doi: 10.1094/MPMI.2003.16.12.1118
Barba, P., Cadle-Davidson, L., Harriman, J., Glaubitz, J. C., Brooks, S., Hyma, K., et al. (2014). Grapevine powdery mildew resistance and susceptibility loci identified on a high-resolution SNP map. Theor. Appl. Genet. 127, 73–84. doi: 10.1007/s00122-013-2202-x
Barker, C. L., Donald, T., Pauquet, J., Ratnaparkhe, M. B., Bouquet, A., Adam-Blondon, A. F., et al. (2005). Genetic and physical mapping of the grapevine powdery mildew resistance gene, Run1, using a bacterial artificial chromosome library. Theor. Appl. Genet. 111, 370–377. doi: 10.1007/s00122-005-2030-8
Basso, M. F., da Silva, J. C., Fajardo, T. V., Fontes, E. P., and Zerbini, F. M. (2015). A novel, highly divergent ssDNA virus identified in Brazil infecting apple, pear and grapevine. Virus Res. 210, 27–33. doi: 10.1016/j.virusres.2015.07.005
Bellin, D., Peressotti, E., Merdinoglu, D., Wiedemann-Merdinoglu, S., Adam-Blondon, A. F., Cipriani, G., et al. (2009). Resistance to Plasmopara viticola in grapevine ‘Bianca’ is controlled by a major dominant gene causing localised necrosis at the infection site. Theor. Appl. Genet. 120, 163–176. doi: 10.1007/s00122-009-1167-2
Blanc, S., Wiedemann-Merdinoglu, S., Dumas, V., Mestre, P., and Merdinoglu, D. (2012). A reference genetic map of Muscadinia rotundifolia and identification of Ren5, a new major locus for resistance to grapevine powdery mildew. Theor. Appl. Genet. 125, 1663–1675. doi: 10.1007/s00122-012-1942-3
Blanco-Ulate, B., Morales-Cruz, A., Amrine, K. C., Labavitch, J. M., Powell, A. L., and Cantu, D. (2014). Genome-wide transcriptional profiling of Botrytis cinerea genes targeting plant cell walls during infections of different hosts. Front. Plant Sci. 5:435. doi: 10.3389/fpls.2014.00435
Blasi, P., Blanc, S., Wiedemann-Merdinoglu, S., Prado, E., Ruhl, E. H., Mestre, P., et al. (2011). Construction of a reference linkage map of Vitis amurensis and genetic mapping of Rpv8, a locus conferring resistance to grapevine downy mildew. Theor. Appl. Genet. 123, 43–53. doi: 10.1007/s00122-011-1565-0
Bonavia, M., Digiaro, M., Boscia, D., Boari, A., Bottalico, G., Savino, V., et al. (1996). Studies on “corky rugose wood” of grapevine and on the diagnosis of grapevine virus B. Vitis 35, 53–58.
Burgyan, J., and Havelda, Z. (2011). Viral suppressors of RNA silencing. Trends Plant Sci. 16, 265–272. doi: 10.1016/j.tplants.2011.02.010
Burr, T. J., Bazzi, C., Süle, S., and Otten, L. (1998). Crown gall of grape: biology of Agrobacterium vitis and the development of disease control strategies. Plant Dis. 82, 1288–1297. doi: 10.1094/PDIS.1998.82.12.1288
Burr, T. J., and Otten, L. (1999). CROWN GALL OF GRAPE: biology and disease management. Annu. Rev. Phytopathol. 37, 53–80. doi: 10.1146/annurev.phyto.37.1.53
Burruano, S. (2000). The life-cycle of Plasmopara viticola, cause of downy mildew of vine. Micologist 14, 179–182. doi: 10.1016/S0269-915X(00)80040-3
Calonnec, A., Cartolaro, P., Poupot, C., Dubourdieu, D., and Darriet, P. (2004). Effects of Uncinula necator on the yield and quality of grapes (Vitis vinifera) and wine. Plant Pathol. 53, 434–445. doi: 10.1111/j.0032-0862.2004.01016.x
Calvo-Garrido, C., Vinas, I., Elmer, P. A., Usall, J., and Teixido, N. (2014). Suppression of Botrytis cinerea on necrotic grapevine tissues by early-season applications of natural products and biological control agents. Pest Manag. Sci. 70, 595–602. doi: 10.1002/ps.3587
Carrington, J. C., Kasschau, K. D., Mahajan, S. K., and Schaad, M. C. (1996). Cell-to-cell and long-distance transport of viruses in plants. Plant Cell 8, 1669–1681. doi: 10.1105/tpc.8.10.1669
Casagrande, K., Falginella, L., Castellarin, S. D., Testolin, R., and Di Gaspero, G. (2011). Defence responses in Rpv3-dependent resistance to grapevine downy mildew. Planta 234, 1097–1109. doi: 10.1007/s00425-011-1461-5
Chiba, M., Reed, J. C., Prokhnevsky, A. I., Chapman, E. J., Mawassi, M., Koonin, E. V., et al. (2006). Diverse suppressors of RNA silencing enhance agroinfection by a viral replicon. Virology 346, 7–14. doi: 10.1016/j.virol.2005.09.068
Choi, Y. J., Yun, H. K., Park, K. S., Noh, J. H., Heo, Y. Y., Kim, S. H., et al. (2010). Transcriptional profiling of ESTs responsive to Rhizobium vitis from ‘Tamnara’ grapevines (Vitis sp.). J. Plant Physiol. 167, 1084–1092. doi: 10.1016/j.jplph.2010.02.005
Chong, J., Le Henanff, G., Bertsch, C., and Walter, B. (2008). Identification, expression analysis and characterization of defense and signaling genes in Vitis vinifera. Plant Physiol. Biochem. 46, 469–481. doi: 10.1016/j.plaphy.2007.09.010
Clement, B., Pollmann, S., Weiler, E., Urbanczyk-Wochniak, E., and Otten, L. (2006). The Agrobacterium vitis T-6b oncoprotein induces auxin-independent cell expansion in tobacco. Plant J. 45, 1017–1027. doi: 10.1111/j.1365-313X.2006.02663.x
Coleman, C., Copetti, D., Cipriani, G., Hoffmann, S., Kozma, P., Kovacs, L., et al. (2009). The powdery mildew resistance gene REN1 co-segregates with an NBS-LRR gene cluster in two Central Asian grapevines. BMC Genet. 10:89. doi: 10.1186/1471-2156-10-89
Consonni, C., Humphry, M. E., Hartmann, H. A., Livaja, M., Durner, J., Westphal, L., et al. (2006). Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Genet. 38, 716–720. doi: 10.1038/ng1806
Cotoras, M., Garcia, C., and Mendoza, L. (2009). Botrytis cinerea isolates collected from grapes present different requirements for conidia germination. Mycologia 101, 287–295. doi: 10.3852/08-012
Dadakova, K., Havelkova, M., Kurkova, B., Tlolkova, I., Kasparovsky, T., Zdrahal, Z., et al. (2015). Proteome and transcript analysis of Vitis vinifera cell cultures subjected to Botrytis cinerea infection. J. Proteomics 119, 143–153. doi: 10.1016/j.jprot.2015.02.001
Dai, R., Ge, H., Howard, S., and Qiu, W. (2012). Transcriptional expression of Stilbene synthase genes are regulated developmentally and differentially in response to powdery mildew in Norton and Cabernet Sauvignon grapevine. Plant Sci. 197, 70–76. doi: 10.1016/j.plantsci.2012.09.004
Dalbó, M., Ye, G., Weeden, N., Wilcox, W., and Reisch, B. (2001). Marker-assisted selection for powdery mildew resistance in grapes. J. Am. Soc. Hortic. Sci. 126, 183–189.
Davis, M. J., Purcell, A. H., and Thomson, S. V. (1978). Pierce’s disease of grapevines: isolation of the causal bacterium. Science 199, 75–77. doi: 10.1126/science.199.4324.75
de Carvalho, F., Gheysen, G., Kushnir, S., Van Montagu, M., Inze, D., and Castresana, C. (1992). Suppression of beta-1,3-glucanase transgene expression in homozygous plants. EMBO J. 11, 2595–2602.
De Lorenzo, G., D’Ovidio, R., and Cervone, F. (2001). The role of polygalacturonase-inhibiting proteins (PGIPs) in defense against pathogenic fungi. Annu. Rev. Phytopathol. 39, 313–335. doi: 10.1146/annurev.phyto.39.1.313
Dean, R., Van Kan, J. A., Pretorius, Z. A., Hammond-Kosack, K. E., Di Pietro, A., Spanu, P. D., et al. (2012). The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 13, 414–430. doi: 10.1111/j.1364-3703.2011.00783.x
Derckel, J. P., Baillieul, F., Manteau, S., Audran, J. C., Haye, B., Lambert, B., et al. (1999). Differential induction of grapevine defenses by two strains of Botrytis cinerea. Phytopathology 89, 197–203. doi: 10.1094/PHYTO.1999.89.3.197
Di Gaspero, G., Copetti, D., Coleman, C., Castellarin, S. D., Eibach, R., Kozma, P., et al. (2012). Selective sweep at the Rpv3 locus during grapevine breeding for downy mildew resistance. Theor. Appl. Genet. 124, 277–286. doi: 10.1007/s00122-011-1703-8
Ding, S. W., and Voinnet, O. (2007). Antiviral immunity directed by small RNAs. Cell 130, 413–426. doi: 10.1016/j.cell.2007.07.039
Dorssers, L., van der Krol, S., van der Meer, J., van Kammen, A., and Zabel, P. (1984). Purification of cowpea mosaic virus RNA replication complex: identification of a virus-encoded 110,000-dalton polypeptide responsible for RNA chain elongation. Proc. Natl. Acad. Sci. U.S.A. 81, 1951–1955. doi: 10.1073/pnas.81.7.1951
El Gueddari, N., Rauchhaus, U., Moerschbacher, B., and Deising, H. (2002). Developmentally regulated conversion of surface-exposed chitin to chitosan in cell walls of plant pathogenic fungi. New Phytol. 156, 103–112. doi: 10.1046/j.1469-8137.2002.00487.x
Escobar, M. A., and Dandekar, A. M. (2003). Agrobacterium tumefaciens as an agent of disease. Trends Plant Sci. 8, 380–386. doi: 10.1016/S1360-1385(03)00162-6
Espinoza, C., Medina, C., Somerville, S., and Arce-Johnson, P. (2007a). Senescence-associated genes induced during compatible viral interactions with grapevine and Arabidopsis. J. Exp. Bot. 58, 3197–3212. doi: 10.1093/jxb/erm165
Espinoza, C., Vega, A., Medina, C., Schlauch, K., Cramer, G., and Arce-Johnson, P. (2007b). Gene expression associated with compatible viral diseases in grapevine cultivars. Funct. Integr. Genomics 7, 95–110. doi: 10.1007/s10142-006-0031-6
Feechan, A., Anderson, C., Torregrosa, L., Jermakow, A., Mestre, P., Wiedemann-Merdinoglu, S., et al. (2013). Genetic dissection of a TIR-NB-LRR locus from the wild North American grapevine species Muscadinia rotundifolia identifies paralogous genes conferring resistance to major fungal and oomycete pathogens in cultivated grapevine. Plant J. 76, 661–674. doi: 10.1111/tpj.12327
Feechan, A., Jermakow, A. M., and Dry, I. B. (2009). Grapevine MLO candidates required for powdery mildew pathogenicity? Plant Signal. Behav. 4, 522–523. doi: 10.1071/FP08173
Feechan, A., Kabbara, S., and Dry, I. B. (2011). Mechanisms of powdery mildew resistance in the Vitaceae family. Mol. Plant Pathol. 12, 263–274. doi: 10.1111/j.1364-3703.2010.00668.x
Freialdenhoven, A., Peterhansel, C., Kurth, J., Kreuzaler, F., and Schulze-Lefert, P. (1996). Identification of genes required for the function of non-race-specific mlo resistance to powdery mildew in barley. Plant Cell 8, 5–14. doi: 10.1105/tpc.8.1.5
Fry, S. M., and Milholland, D. (1990). Response of resistant, tolerant, and susceptible grapevine tissue to invasion by the Pierce’s disease bacterium Xylella fastidiosa. Phytopathology 80, 66–69. doi: 10.1094/Phyto-80-66
Fung, R. W., Gonzalo, M., Fekete, C., Kovacs, L. G., He, Y., Marsh, E., et al. (2008). Powdery mildew induces defense-oriented reprogramming of the transcriptome in a susceptible but not in a resistant grapevine. Plant Physiol. 146, 236–249. doi: 10.1104/pp.107.108712
Gabler, F. M., Smilanick, J. L., Mansour, M., Ramming, D. W., and Mackey, B. E. (2003). Correlations of morphological, anatomical, and chemical features of grape berries with resistance to Botrytis cinerea. Phytopathology 93, 1263–1273. doi: 10.1094/PHYTO.2003.93.10.1263
Gadoury, D. M., Cadle-Davidson, L., Wilcox, W. F., Dry, I. B., Seem, R. C., and Milgroom, M. G. (2012). Grapevine powdery mildew (Erysiphe necator): a fascinating system for the study of the biology, ecology and epidemiology of an obligate biotroph. Mol. Plant Pathol. 13, 1–16. doi: 10.1111/j.1364-3703.2011.00728.x
Gadoury, D. M., Seem, R. C., Ficke, A., and Wilcox, W. F. (2001). The epidemiology of powdery mildew on concord grapes. Phytopathology 91, 948–955. doi: 10.1094/PHYTO.2001.91.10.948
Gambino, G., Cuozzo, D., Fasoli, M., Pagliarani, C., Vitali, M., Boccacci, P., et al. (2012). Co-evolution between Grapevine rupestris stem pitting-associated virus and Vitis vinifera L. leads to decreased defence responses and increased transcription of genes related to photosynthesis. J. Exp. Bot. 63, 5919–5933. doi: 10.1093/jxb/ers244
Gao, F., Dai, R., Pike, S. M., Qiu, W., and Gassmann, W. (2014). Functions of EDS1-like and PAD4 genes in grapevine defenses against powdery mildew. Plant Mol. Biol. 86, 381–393. doi: 10.1007/s11103-014-0235-4
Gessler, C., Pertot, I., and Perazzolli, M. (2011). Plasmopara viticola: a review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 50, 3–44.
Ghanem-Sabanadzovic, N. A., Sabanadzovic, S., Gugerli, P., and Rowhani, A. (2012). Genome organization, serology and phylogeny of Grapevine leafroll-associated viruses 4 and 6: taxonomic implications. Virus Res. 163, 120–128. doi: 10.1016/j.virusres.2011.09.001
Ghanem-Sabanadzovic, N. A., Sabanadzovic, S., Uyemoto, J. K., Golino, D., and Rowhani, A. (2010). A putative new Ampelovirus associated with grapevine leafroll disease. Arch. Virol. 155, 1871–1876. doi: 10.1007/s00705-010-0773-2
Giacomelli, L., Nanni, V., Lenzi, L., Zhuang, J., Dalla Serra, M., Banfield, M. J., et al. (2012). Identification and characterization of the defensin-like gene family of grapevine. Mol. Plant Microbe Interact. 25, 1118–1131. doi: 10.1094/MPMI-12-11-0323
Girault, T., Francois, J., Rogniaux, H., Pascal, S., Delrot, S., Coutos-Thevenot, P., et al. (2008). Exogenous application of a lipid transfer protein-jasmonic acid complex induces protection of grapevine towards infection by Botrytis cinerea. Plant Physiol. Biochem. 46, 140–149. doi: 10.1016/j.plaphy.2007.10.005
Glazebrook, J. (2005). Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 43, 205–227. doi: 10.1146/annurev.phyto.43.040204.135923
Goszczynski, D. E., du Preez, J., and Burger, J. T. (2008). Molecular divergence of Grapevine virus A (GVA) variants associated with Shiraz disease in South Africa. Virus Res. 138, 105–110. doi: 10.1016/j.virusres.2008.08.014
Gourgues, M., Brunet-Simon, A., Lebrun, M. H., and Levis, C. (2004). The tetraspanin BcPls1 is required for appressorium-mediated penetration of Botrytis cinerea into host plant leaves. Mol. Microbiol. 51, 619–629. doi: 10.1046/j.1365-2958.2003.03866.x
Guillier, C., Gamm, M., Lucchi, G., Truntzer, C., Pecqueur, D., Ducoroy, P., et al. (2015). Toward the identification of two glycoproteins involved in the stomatal deregulation of downy mildew-infected grapevine leaves. Mol. Plant Microbe Interact. 28, 1227–1236. doi: 10.1094/MPMI-05-15-0115-R
Gutha, L. R., Casassa, L. F., Harbertson, J. F., and Naidu, R. A. (2010). Modulation of flavonoid biosynthetic pathway genes and anthocyanins due to virus infection in grapevine (Vitis vinifera L.) leaves. BMC Plant Biol. 10:187. doi: 10.1186/1471-2229-10-187
Hammond, S. M., Caudy, A. A., and Hannon, G. J. (2001). Post-transcriptional gene silencing by double-stranded RNA. Nat. Rev. Genet. 2, 110–119. doi: 10.1038/35052556
Harries, P. A., Schoelz, J. E., and Nelson, R. S. (2010). Intracellular transport of viruses and their components: utilizing the cytoskeleton and membrane highways. Mol. Plant Microbe Interact. 23, 1381–1393. doi: 10.1094/MPMI-05-10-0121
Hayward, A. P., Tsao, J., and Dinesh-Kumar, S. P. (2009). Autophagy and plant innate immunity: defense through degradation. Semin. Cell Dev. Biol. 20, 1041–1047. doi: 10.1016/j.semcdb.2009.04.012
Hipper, C., Brault, V., Ziegler-Graff, V., and Revers, F. (2013). Viral and cellular factors involved in Phloem transport of plant viruses. Front. Plant Sci. 4:154. doi: 10.3389/fpls.2013.00154
Hoffmann, S., Di Gaspero, G., Kovacs, L., Howard, S., Kiss, E., Galbacs, Z., et al. (2008). Resistance to Erysiphe necator in the grapevine ‘Kishmish vatkana’ is controlled by a single locus through restriction of hyphal growth. Theor. Appl. Genet. 116, 427–438. doi: 10.1007/s00122-007-0680-4
Hong, Y. S., Martinez, A., Liger-Belair, G., Jeandet, P., Nuzillard, J. M., and Cilindre, C. (2012). Metabolomics reveals simultaneous influences of plant defence system and fungal growth in Botrytis cinerea-infected Vitis vinifera cv. Chardonnay berries. J. Exp. Bot. 63, 5773–5785. doi: 10.1093/jxb/ers228
Jelly, N. S., Schellenbaum, P., Walter, B., and Maillot, P. (2012). Transient expression of artificial microRNAs targeting Grapevine fanleaf virus and evidence for RNA silencing in grapevine somatic embryos. Transgenic Res. 21, 1319–1327. doi: 10.1007/s11248-012-9611-5
Jia, H., Zhang, C., Pervaiz, T., Zhao, P., Liu, Z., Wang, B., et al. (2016). Jasmonic acid involves in grape fruit ripening and resistant against Botrytis cinerea. Funct. Integr. Genomics 16, 79–94. doi: 10.1007/s10142-015-0468-6
Jones, J. D., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Jones, L., Riaz, S., Morales-Cruz, A., Amrine, K. C., McGuire, B., Gubler, W. D., et al. (2014). Adaptive genomic structural variation in the grape powdery mildew pathogen, Erysiphe necator. BMC Genomics 15:1081. doi: 10.1186/1471-2164-15-1081
Kamoun, S., Furzer, O., Jones, J. D., Judelson, H. S., Ali, G. S., Dalio, R. J., et al. (2015). The Top 10 oomycete pathogens in molecular plant pathology. Mol. Plant Pathol. 16, 413–434. doi: 10.1111/mpp.12190
Kars, I., Krooshof, G. H., Wagemakers, L., Joosten, R., Benen, J. A., and van Kan, J. A. (2005a). Necrotizing activity of five Botrytis cinerea endopolygalacturonases produced in Pichia pastoris. Plant J. 43, 213–225. doi: 10.1111/j.1365-313X.2005.02436.x
Kars, I., McCalman, M., Wagemakers, L., and Ja, V. A. N. K. (2005b). Functional analysis of Botrytis cinerea pectin methylesterase genes by PCR-based targeted mutagenesis: Bcpme1 and Bcpme2 are dispensable for virulence of strain B05.10. Mol. Plant Pathol. 6, 641–652. doi: 10.1111/j.1364-3703.2005.00312.x
Kawakami, S., Watanabe, Y., and Beachy, R. N. (2004). Tobacco mosaic virus infection spreads cell to cell as intact replication complexes. Proc. Natl. Acad. Sci. U.S.A. 101, 6291–6296. doi: 10.1073/pnas.0401221101
Kelloniemi, J., Trouvelot, S., Heloir, M. C., Simon, A., Dalmais, B., Frettinger, P., et al. (2015). Analysis of the molecular dialogue between gray mold (Botrytis cinerea) and grapevine (Vitis vinifera) reveals a clear shift in defense mechanisms during berry ripening. Mol. Plant Microbe Interact. 28, 1167–1180. doi: 10.1094/MPMI-02-15-0039-R
Komatsu, K., Hashimoto, M., Ozeki, J., Yamaji, Y., Maejima, K., Senshu, H., et al. (2010). Viral-induced systemic necrosis in plants involves both programmed cell death and the inhibition of viral multiplication, which are regulated by independent pathways. Mol. Plant Microbe Interact. 23, 283–293. doi: 10.1094/MPMI-23-3-0283
Krastanova, S., Perrin, M., Barbier, P., Demangeat, G., Cornuet, P., Bardonnet, N., et al. (1995). Transformation of grapevine rootstocks with the coat protein gene of grapevine fanleaf nepovirus. Plant Cell Rep. 14, 550–554. doi: 10.1007/BF00231936
Krivanek, A. F., Riaz, S., and Walker, M. A. (2006). Identification and molecular mapping of PdR1, a primary resistance gene to Pierce’s disease in Vitis. Theor. Appl. Genet. 112, 1125–1131. doi: 10.1007/s00122-006-0214-5
Kuczmog, A., Galambos, A., Horvath, S., Matai, A., Kozma, P., Szegedi, E., et al. (2012). Mapping of crown gall resistance locus Rcg1 in grapevine. Theor. Appl. Genet. 125, 1565–1574. doi: 10.1007/s00122-012-1935-2
Laporte, C., Vetter, G., Loudes, A. M., Robinson, D. G., Hillmer, S., Stussi-Garaud, C., et al. (2003). Involvement of the secretory pathway and the cytoskeleton in intracellular targeting and tubule assembly of Grapevine fanleaf virus movement protein in tobacco BY-2 cells. Plant Cell 15, 2058–2075. doi: 10.1105/tpc.013896
Le Henanff, G., Profizi, C., Courteaux, B., Rabenoelina, F., Gerard, C., Clement, C., et al. (2013). Grapevine NAC1 transcription factor as a convergent node in developmental processes, abiotic stresses, and necrotrophic/biotrophic pathogen tolerance. J. Exp. Bot. 64, 4877–4893. doi: 10.1093/jxb/ert277
Leisner, S. M., and Turgeon, R. (1993). Movement of virus and photoassimilate in the phloem: a comparative analysis. Bioessays 15, 741–748. doi: 10.1002/bies.950151107
Lenzi, L., Caruso, C., Bianchedi, P. L., Pertot, I., and Perazzolli, M. (2016). Laser microdissection of grapevine leaves reveals site-specific regulation of transcriptional response to Plasmopara viticola. Plant Cell Physiol. 57, 69–81. doi: 10.1093/pcp/pcv166
Li, F., and Ding, S. W. (2006). Virus counterdefense: diverse strategies for evading the RNA-silencing immunity. Annu. Rev. Microbiol. 60, 503–531. doi: 10.1146/annurev.micro.60.080805.142205
Li, H., Xu, Y., Xiao, Y., Zhu, Z., Xie, X., Zhao, H., et al. (2010). Expression and functional analysis of two genes encoding transcription factors, VpWRKY1 and VpWRKY2, isolated from Chinese wild Vitis pseudoreticulata. Planta 232, 1325–1337. doi: 10.1007/s00425-010-1258-y
Lieth, J. H., Meyer, M. M., Yeo, K. H., and Kirkpatrick, B. C. (2011). Modeling cold curing of Pierce’s disease in Vitis vinifera ‘Pinot Noir’ and ‘Cabernet Sauvignon’ grapevines in California. Phytopathology 101, 1492–1500. doi: 10.1094/PHYTO-08-10-0207
Lin, H., Doddapaneni, H., Takahashi, Y., and Walker, M. A. (2007). Comparative analysis of ESTs involved in grape responses to Xylella fastidiosa infection. BMC Plant Biol. 7:8. doi: 10.1186/1471-2229-7-8
Liu, J. Z., Blancaflor, E. B., and Nelson, R. S. (2005). The Tobacco mosaic virus 126-kilodalton protein, a constituent of the virus replication complex, alone or within the complex aligns with and traffics along microfilaments. Plant Physiol. 138, 1853–1865. doi: 10.1104/pp.105.065722
Liu, R., Wang, L., Zhu, J., Chen, T., Wang, Y., and Xu, Y. (2015). Histological responses to downy mildew in resistant and susceptible grapevines. Protoplasma 252, 259–270. doi: 10.1007/s00709-014-0677-1
Liu, Y. P., Peremyslov, V. V., Medina, V., and Dolja, V. V. (2009). Tandem leader proteases of Grapevine leafroll-associated virus-2: host-specific functions in the infection cycle. Virology 383, 291–299. doi: 10.1016/j.virol.2008.09.035
Lucas, W. J. (2006). Plant viral movement proteins: agents for cell-to-cell trafficking of viral genomes. Virology 344, 169–184. doi: 10.1016/j.virol.2005.09.026
Luis, P., Gauthier, A., Trouvelot, S., Poinssot, B., and Frettinger, P. (2013). Identification of Plasmopara viticola genes potentially involved in pathogenesis on grapevine suggests new similarities between oomycetes and true fungi. Phytopathology 103, 1035–1044. doi: 10.1094/PHYTO-06-12-0121-R
Malacarne, G., Vrhovsek, U., Zulini, L., Cestaro, A., Stefanini, M., Mattivi, F., et al. (2011). Resistance to Plasmopara viticola in a grapevine segregating population is associated with stilbenoid accumulation and with specific host transcriptional responses. BMC Plant Biol. 11:114. doi: 10.1186/1471-2229-11-114
Maliogka, V. I., Dovas, C. I., Lotos, L., Efthimiou, K., and Katis, N. I. (2009). Complete genome analysis and immunodetection of a member of a novel virus species belonging to the genus Ampelovirus. Arch. Virol. 154, 209–218. doi: 10.1007/s00705-008-0290-8
Marsh, E., Alvarez, S., Hicks, L. M., Barbazuk, W. B., Qiu, W., Kovacs, L., et al. (2010). Changes in protein abundance during powdery mildew infection of leaf tissues of Cabernet Sauvignon grapevine (Vitis vinifera L.). Proteomics 10, 2057–2064. doi: 10.1002/pmic.200900712
Martelli, G. P. (2014). “Virus diseases of grapevine,” in eLS (Chichester: John Wiley & Sons, Ltd). doi: 10.1002/9780470015902.a0000766.pub3
Martinelli, L., Candioli, E., Costa, D., and Minafra, A. (2002). Stable insertion and expression of the movement protein gene of Grapevine Virus A (GVA) in grape (Vitis rupestris S.). Vitis 41, 189–193.
McHale, L., Tan, X., Koehl, P., and Michelmore, R. W. (2006). Plant NBS-LRR proteins: adaptable guards. Genome Biol. 7, 212. doi: 10.1186/gb-2006-7-4-212
Merdinoglu, D., Wiedeman-Merdinoglu, S., Coste, P., Dumas, V., Haetty, S., Butterlin, G., et al. (2003). “Genetic analysis of Downy Mildew resistance derived from Muscadinia rotundifolia,” in Proceedings of the Eighth International Conference on Grape Genetics and Breeding, Leuven, 451–456.
Merz, P. R., Moser, T., Holl, J., Kortekamp, A., Buchholz, G., Zyprian, E., et al. (2015). The transcription factor VvWRKY33 is involved in the regulation of grapevine (Vitis vinifera) defense against the oomycete pathogen Plasmopara viticola. Physiol. Plant. 153, 365–380. doi: 10.1111/ppl.12251
Mestre, P., Piron, M. C., and Merdinoglu, D. (2012). Identification of effector genes from the phytopathogenic Oomycete Plasmopara viticola through the analysis of gene expression in germinated zoospores. Fungal Biol. 116, 825–835. doi: 10.1016/j.funbio.2012.04.016
Meyer, P., Heidmann, I., and Niedenhof, I. (1993). Differences in DNA-methylation are associated with a paramutation phenomenon in transgenic petunia. Plant J. 4, 89–100. doi: 10.1046/j.1365-313X.1993.04010089.x
Monaghan, J., and Zipfel, C. (2012). Plant pattern recognition receptor complexes at the plasma membrane. Curr. Opin. Plant Biol. 15, 349–357. doi: 10.1016/j.pbi.2012.05.006
Murray, G. G., Kosakovsky Pond, S. L., and Obbard, D. J. (2013). Suppressors of RNAi from plant viruses are subject to episodic positive selection. Proc. Biol. Sci. 280:20130965. doi: 10.1098/rspb.2013.0965
Naidu, R. A., Maree, H. J., and Burger, J. T. (2015). Grapevine leafroll disease and associated viruses: a unique pathosystem. Annu. Rev. Phytopathol. 53, 613–634. doi: 10.1146/annurev-phyto-102313-045946
Newman, K. L., Almeida, R. P., Purcell, A. H., and Lindow, S. E. (2003). Use of a green fluorescent strain for analysis of Xylella fastidiosa colonization of Vitis vinifera. Appl. Environ. Microbiol. 69, 7319–7327. doi: 10.1128/AEM.69.12.7319-7327.2003
Niehl, A., and Heinlein, M. (2011). Cellular pathways for viral transport through plasmodesmata. Protoplasma 248, 75–99. doi: 10.1007/s00709-010-0246-1
Parafati, L., Vitale, A., Restuccia, C., and Cirvilleri, G. (2015). Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol. 47, 85–92. doi: 10.1016/j.fm.2014.11.013
Park, K. S., Yun, H. K., Suh, H. S., Jeong, S. B., and Cho, H. M. (2004). Breeding of early season grape cultivar ‘Tamnara’ grapevines (Vitis hybrid) with high quality and disease resistance. Korean J. Hortic. Sci. Technol. 22, 458–461.
Pedro, H., Maheswari, U., Urban, M., Irvine, A. G., Cuzick, A., McDowall, M. D., et al. (2016). PhytoPath: an integrative resource for plant pathogen genomics. Nucleic Acids Res. 44, D688–D693. doi: 10.1093/nar/gkv1052
Peressotti, E., Wiedemann-Merdinoglu, S., Delmotte, F., Bellin, D., Di Gaspero, G., Testolin, R., et al. (2010). Breakdown of resistance to grapevine downy mildew upon limited deployment of a resistant variety. BMC Plant Biol. 10:147. doi: 10.1186/1471-2229-10-147
Perfect, S. E., and Green, J. R. (2001). Infection structures of biotrophic and hemibiotrophic fungal plant pathogens. Mol. Plant Pathol. 2, 101–108. doi: 10.1046/j.1364-3703.2001.00055.x
Pieterse, C. M., Leon-Reyes, A., Van der Ent, S., and Van Wees, S. C. (2009). Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 5, 308–316. doi: 10.1038/nchembio.164
Pike, S., Gao, F., Kim, M. J., Kim, S. H., Schachtman, D. P., and Gassmann, W. (2014). Members of the NPF3 transporter subfamily encode pathogen-inducible nitrate/nitrite transporters in grapevine and Arabidopsis. Plant Cell Physiol. 55, 162–170. doi: 10.1093/pcp/pct167
Polesani, M., Desario, F., Ferrarini, A., Zamboni, A., Pezzotti, M., Kortekamp, A., et al. (2008). cDNA-AFLP analysis of plant and pathogen genes expressed in grapevine infected with Plasmopara viticola. BMC Genomics 9:142. doi: 10.1186/1471-2164-9-142
Prosser, S. W., Goszczynski, D. E., and Meng, B. (2007). Molecular analysis of double-stranded RNAs reveals complex infection of grapevines with multiple viruses. Virus Res. 124, 151–159. doi: 10.1016/j.virusres.2006.10.014
Qiu, W., Feechan, A., and Dry, I. (2015). Current understanding of grapevine defense mechanisms against the biotrophic fungus (Erysiphe necator), the causal agent of powdery mildew disease. Hortic. Res. 2:15020. doi: 10.1038/hortres.2015.20
Ramming, D. W., Gabler, F., Smilanick, J., Cadle-Davidson, M., Barba, P., Mahanil, S., et al. (2011). A single dominant locus, ren4, confers rapid non-race-specific resistance to grapevine powdery mildew. Phytopathology 101, 502–508. doi: 10.1094/PHYTO-09-10-0237
Riaz, S., Krivanek, A. F., Xu, K., and Walker, M. A. (2006). Refined mapping of the Pierce’s disease resistance locus, PdR1, and Sex on an extended genetic map of Vitis rupestris x V. arizonica. Theor. Appl. Genet. 113, 1317–1329. doi: 10.1007/s00122-006-0385-0
Riaz, S., Tenscher, A. C., Ramming, D. W., and Walker, M. A. (2011). Using a limited mapping strategy to identify major QTLs for resistance to grapevine powdery mildew (Erysiphe necator) and their use in marker-assisted breeding. Theor. Appl. Genet. 122, 1059–1073. doi: 10.1007/s00122-010-1511-6
Riaz, S., Tenscher, A. C., Rubin, J., Graziani, R., Pao, S. S., and Walker, M. A. (2008). Fine-scale genetic mapping of two Pierce’s disease resistance loci and a major segregation distortion region on chromosome 14 of grape. Theor. Appl. Genet. 117, 671–681. doi: 10.1007/s00122-008-0802-7
Robert, N., Roche, K., Lebeau, Y., Breda, C., Boulay, M., Esnault, R., et al. (2002). Expression of grapevine chitinase genes in berries and leaves infected by fungal or bacterial pathogens. Plant Sci. 162, 389–400. doi: 10.1016/S0168-9452(01)00576-3
Robert-Seilaniantz, A., Grant, M., and Jones, J. D. (2011). Hormone crosstalk in plant disease and defense: more than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 49, 317–343. doi: 10.1146/annurev-phyto-073009-114447
Rolke, Y., Liu, S., Quidde, T., Williamson, B., Schouten, A., Weltring, K. M., et al. (2004). Functional analysis of H(2)O(2)-generating systems in Botrytis cinerea: the major Cu-Zn-superoxide dismutase (BCSOD1) contributes to virulence on French bean, whereas a glucose oxidase (BCGOD1) is dispensable. Mol. Plant Pathol. 5, 17–27. doi: 10.1111/j.1364-3703.2004.00201.x
Roper, M. C., Greve, L. C., Warren, J. G., Labavitch, J. M., and Kirkpatrick, B. C. (2007). Xylella fastidiosa requires polygalacturonase for colonization and pathogenicity in Vitis vinifera grapevines. Mol. Plant Microbe Interact. 20, 411–419. doi: 10.1094/MPMI-20-4-0411
Rui, O., and Hahn, M. (2007). The Botrytis cinerea hexokinase, Hxk1, but not the glucokinase, Glk1, is required for normal growth and sugar metabolism, and for pathogenicity on fruits. Microbiology 153, 2791–2802. doi: 10.1099/mic.0.2007/006338-0
Salomone, J. Y., Crouzet, P., De Ruffray, P., and Otten, L. (1996). Characterization and distribution of tartrate utilization genes in the grapevine pathogen Agrobacterium vitis. Mol. Plant Microbe Interact. 9, 401–408. doi: 10.1094/MPMI-9-0401
Schoelz, J. E., Harries, P. A., and Nelson, R. S. (2011). Intracellular transport of plant viruses: finding the door out of the cell. Mol. Plant 4, 813–831. doi: 10.1093/mp/ssr070
Schwander, F., Eibach, R., Fechter, I., Hausmann, L., Zyprian, E., and Topfer, R. (2012). Rpv10: a new locus from the Asian Vitis gene pool for pyramiding downy mildew resistance loci in grapevine. Theor. Appl. Genet. 124, 163–176. doi: 10.1007/s00122-011-1695-4
Sijen, T., Vijn, I., Rebocho, A., van Blokland, R., Roelofs, D., Mol, J. N., et al. (2001). Transcriptional and posttranscriptional gene silencing are mechanistically related. Curr. Biol. 11, 436–440. doi: 10.1016/S0960-9822(01)00116-6
Silva, M. S., De Souza, A. A., Takita, M. A., Labate, C. A., and Machado, M. A. (2011). Analysis of the biofilm proteome of Xylella fastidiosa. Proteome Sci. 9, 58. doi: 10.1186/1477-5956-9-58
Soler, N., Plomer, M., Fagoaga, C., Moreno, P., Navarro, L., Flores, R., et al. (2012). Transformation of Mexican lime with an intron-hairpin construct expressing untranslatable versions of the genes coding for the three silencing suppressors of Citrus tristeza virus confers complete resistance to the virus. Plant Biotechnol. J. 10, 597–608. doi: 10.1111/j.1467-7652.2012.00691.x
Stange, C., Matus, J. T., Dominguez, C., Perez-Acle, T., and Arce-Johnson, P. (2008). The N-homologue LRR domain adopts a folding which explains the TMV-Cg-induced HR-like response in sensitive tobacco plants. J. Mol. Graph. Model. 26, 850–860. doi: 10.1016/j.jmgm.2007.05.006
Süle, S., Mozsar, J., and Burr, T. J. (1994). Crown gall resistance of Vitis spp. and grapevine rootstocks. Phytopathology 84, 607–611. doi: 10.1094/Phyto-84-607
Sun, Q., Rost, T. L., and Matthews, M. A. (2006). Pruning-induced tylose development in stems of current-year shoots of Vitis vinifera (Vitaceae). Am. J. Bot. 93, 1567–1576. doi: 10.3732/ajb.93.11.1567
Swinnen, G., Goossens, A., and Pauwels, L. (2016). Lessons from domestication: targeting cis-regulatory elements for crop improvement. Trends Plant Sci. doi: 10.1016/j.tplants.2016.01.014 [Epub ahead of print].
Szegedi, E., Czakó, M., Otten, L., and Koncz, C. (1988). Opines in crown gall tumours induced by biotype 3 isolates of Agrobacterium tumefaciens. Physiol. Mol. Plant Pathol. 32, 237–247. doi: 10.1016/S0885-5765(88)80020-1
Thaler, J. S., Humphrey, P. T., and Whiteman, N. K. (2012). Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 17, 260–270. doi: 10.1016/j.tplants.2012.02.010
Turner, J. G., Ellis, C., and Devoto, A. (2002). The jasmonate signal pathway. Plant Cell 14(Suppl.), S153–S164.
Van Blokland, R., Van der Geest, N., Mol, J. N., and Kooter, J. M. (1994). Transgene-mediated suppression of chalcone synthase expression in Petunia hybrida results from an increase in RNA turnover. Plant J. 6, 861–877. doi: 10.1046/j.1365-313X.1994.6060861.x
Van den Born, E., Omelchenko, M. V., Bekkelund, A., Leihne, V., Koonin, E. V., Dolja, V. V., et al. (2008). Viral AlkB proteins repair RNA damage by oxidative demethylation. Nucleic Acids Res. 36, 5451–5461. doi: 10.1093/nar/gkn519
van Kan, J. A. (2006). Licensed to kill: the lifestyle of a necrotrophic plant pathogen. Trends Plant Sci. 11, 247–253. doi: 10.1016/j.tplants.2006.03.005
Vaucheret, H., and Fagard, M. (2001). Transcriptional gene silencing in plants: targets, inducers and regulators. Trends Genet. 17, 29–35. doi: 10.1016/S0168-9525(00)02166-1
Vega, A., Gutierrez, R. A., Pena-Neira, A., Cramer, G. R., and Arce-Johnson, P. (2011). Compatible GLRaV-3 viral infections affect berry ripening decreasing sugar accumulation and anthocyanin biosynthesis in Vitis vinifera. Plant Mol. Biol. 77, 261–274. doi: 10.1007/s11103-011-9807-8
Venuti, S., Copetti, D., Foria, S., Falginella, L., Hoffmann, S., Bellin, D., et al. (2013). Historical introgression of the downy mildew resistance gene Rpv12 from the Asian species Vitis amurensis into grapevine varieties. PLoS ONE 8:e61228. doi: 10.1371/journal.pone.0061228
Verhagen, B., Trotel-Aziz, P., Jeandet, P., Baillieul, F., and Aziz, A. (2011). Improved resistance against Botrytis cinerea by grapevine-associated bacteria that induce a prime oxidative burst and phytoalexin production. Phytopathology 101, 768–777. doi: 10.1094/PHYTO-09-10-0242
Viret, O., Keller, M., Jaudzems, V. G., and Cole, F. M. (2004). Botrytis cinerea infection of grape flowers: light and electron microscopical studies of infection sites. Phytopathology 94, 850–857. doi: 10.1094/PHYTO.2004.94.8.850
Waigmann, E., Lucas, W. J., Citovsky, V., and Zambryski, P. (1994). Direct functional assay for Tobacco mosaic virus cell-to-cell movement protein and identification of a domain involved in increasing plasmodesmal permeability. Proc. Natl. Acad. Sci. U.S.A. 91, 1433–1437. doi: 10.1073/pnas.91.4.1433
Wan, R., Hou, X., Wang, X., Qu, J., Singer, S. D., Wang, Y., et al. (2015). Resistance evaluation of Chinese wild Vitis genotypes against Botrytis cinerea and different responses of resistant and susceptible hosts to the infection. Front. Plant Sci. 6:854. doi: 10.3389/fpls.2015.00854
Wang, K., Liao, Y., Kan, J., Han, L., and Zheng, Y. (2015). Response of direct or priming defense against Botrytis cinerea to methyl jasmonate treatment at different concentrations in grape berries. Int. J. Food Microbiol. 194, 32–39. doi: 10.1016/j.ijfoodmicro.2014.11.006
Welter, L., Göktürk-Baydar, N., Akkurt, M., Maul, E., Eibach, R., Töpfer, R., et al. (2007). Genetic mapping and localization of quantitative trait loci affecting fungal disease resistance and leaf morphology in grapevine (Vitis vinifera L). Mol. Breed. 20, 359–374. doi: 10.1007/s11032-007-9097-7
Weng, K., Li, Z. Q., Liu, R. Q., Wang, L., Wang, Y. J., and Xu, Y. (2014). Transcriptome of Erysiphe necator-infected Vitis pseudoreticulata leaves provides insight into grapevine resistance to powdery mildew. Hortic. Res. 1, 14049. doi: 10.1038/hortres.2014.49
Wolf, S., Deom, C. M., Beachy, R. N., and Lucas, W. J. (1989). Movement protein of Tobacco mosaic virus modifies plasmodesmatal size exclusion limit. Science 246, 377–379. doi: 10.1126/science.246.4928.377
Xu, T. F., Zhao, X. C., Jiao, Y. T., Wei, J. Y., Wang, L., and Xu, Y. (2014). A pathogenesis related protein, VpPR-10.1, from Vitis pseudoreticulata: an insight of its mode of antifungal activity. PLoS ONE 9:e95102. doi: 10.1371/journal.pone.0095102
Xue, B., Ling, K. S., Reid, C. L., Krastanova, S., Sekiya, M., Momol, E. A., et al. (1999). Transformation of five grape rootstocks with plant virus genes and a virE2 gene from Agrobacterium tumefaciens. In Vitro Cell. Dev. Biol. Plant 35, 226–231. doi: 10.1007/s11627-999-0083-0
Ye, K., and Patel, D. J. (2005). RNA silencing suppressor p21 of Beet yellows virus forms an RNA binding octameric ring structure. Structure 13, 1375–1384. doi: 10.1016/j.str.2005.06.017
Yu, Y., Zhang, Y., Yin, L., and Lu, J. (2012). The mode of host resistance to Plasmopara viticola infection of grapevines. Phytopathology 102, 1094–1101. doi: 10.1094/PHYTO-02-12-0028-R
Zabel, P., Jongen-Neven, I., and Van Krammen, A. (1976). In vitro replication of cowpea mosaic virus RNA. II. Solubilization of membrane-bound replicase and the partial purification of the solubilized enzyme. J. Virol. 17, 679–685.
Keywords: grapevine, pathogenic microorganisms, infection strategy, host response, resistance and susceptibility
Citation: Armijo G, Schlechter R, Agurto M, Muñoz D, Nuñez C and Arce-Johnson P (2016) Grapevine Pathogenic Microorganisms: Understanding Infection Strategies and Host Response Scenarios. Front. Plant Sci. 7:382. doi: 10.3389/fpls.2016.00382
Received: 30 December 2015; Accepted: 13 March 2016;
Published: 30 March 2016.
Edited by:
Dilantha Fernando, University of Manitoba, CanadaReviewed by:
Valerian V. Dolja, Oregon State University, USAEric Kemen, Max Planck Institute for Plant Breeding Research, Germany
Copyright © 2016 Armijo, Schlechter, Agurto, Muñoz, Nuñez and Arce-Johnson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patricio Arce-Johnson, parce@bio.puc.cl
†Present address: Rudolf Schlechter, Department of Biology, Institute of Agricultural Sciences, ETH Zurich, 8092 Zurich, Switzerland
‡These authors have contributed equally to this work.