 Cuilin Zhu
Cuilin Zhu Qing Liu
Qing Liu Xin Li2
Xin Li2 Kexiang Liu
Kexiang Liu Ranji Cui
Ranji Cui- 1Department of Cardiovascular Surgery, The Second Hospital of Jilin University, Changchun, China
- 2Jilin Provincial Key Laboratory on Molecular and Chemical Genetic, The Second Hospital of Jilin University, Changchun, China
- 3Department of Cardiovascular Medicine, University of Tokyo, Tokyo, Japan
- 4Department of Surgery, Western University, London, ON, Canada
Hydrogen sulfide (H2S) is one of most important gas transmitters. H2S modulates many physiological and pathological processes such as inflammation, oxidative stress and cell apoptosis that play a critical role in vascular function. Recently, solid evidence show that H2S is closely associated to various vascular diseases. However, specific function of H2S remains unclear. Therefore, in this review we systemically summarized the role of H2S in vascular diseases, including hypertension, atherosclerosis, inflammation and angiogenesis. In addition, this review also outlined a novel therapeutic perspective comprising crosstalk between H2S and smooth muscle cell function. Therefore, this review may provide new insight inH2S application clinically.
Introduction
Hydrogen sulfide (H2S) is recently recognized as the third gas signaling transmitter after nitric oxide (NO) and carbon monoxide (CO) despite it was once considered as toxic gas. Endogenous H2S production is mainly mediated by cystathionine-b-synthase (CBS), cystathionine-c-lyase (CSE), 3-mercaptopyruvate sulfur transferase (3-MST), which are the most pre-dominant enzymes of H2S production (1–3). Exogenous administration of H2S is mainly performed with NaHS salts and H2S related compounds.
Recent studies have proved H2S to be vasculoprotective by participating different cellular pathways and interfering with a variety of vascular diseases (4–7). H2S is endogenously produced by vascular cells or exogenously administered by H2S releasing donors. In the vasculature, H2S regulates the proliferation and migration of endothelial cells and vascular smooth muscle cell, regulates the apoptosis, oxidative stress and inflammation of vascular cells. Furthermore, H2S has been widely proved to regulate many vascular diseases, including hypertension, atherosclerosis (AS) and angiogenesis (8). Although the beneficial effects of H2S have widely been recognized, the mechanisms into the molecular pathways largely remained unknown. Deeper understanding of the working mechanisms will help put their way into further clinical application.
In this review, we review the recent findings about H2S in vascular diseases, including hypertension, AS and angiogenesis, as well as recent working mechanisms. The role of H2S will be separately discussed endogenously and exogenously here. Finally, we will discuss the possible perspectives of H2S in the future.
H2S in hypertension
Hypertension has been a worldwide disease, accounting for 30-40% of the whole population, posing great danger to people’s health (9, 10). It is reported that H2S plays a role in blood pressure regulation. Despite emerging evidence from experimental studies targeting H2S to protect against hypertension, these results need further clinical research.
Endogenous H2S in hypertension
The concentration of H2S in human blood has been reported within a normal range under physical conditions. However, the change of H2S concentration has been reported to be reduced in high blood pressure (HBP) patients, suggesting the potential regulatory role of H2S in HBP (11, 12). Several clinical studies have reported the relationship between H2S and hypertension related disorders (13). Additionally, decreased H2S plasmatic levels were also found in lead-induced HBP patients (14).
Furthermore, the three major generating enzymes, CSE, CBS and 3-MST were reported to be reduced in HBP patients, suggesting the endogenous synthesis of H2S may participate in the pathogenesis of HBP (15). Aging is an important predisposing factor for HBP. Loss of 3-MST using a genetic mouse model recuses the mouse cardiovascular system from aging-dependent disorders, thus regulating progression of HBP (16). Innate immune and adoptive immune cells are essential in the genesis and target-organ damage of hypertension. In a recent study, Cui reported that CSE-derived H2S promotes Treg differentiation and proliferation in an adenosine monophosphate activated protein kinase (AMPK) dependent pathway, which attenuates the vascular immune-inflammation, thereby preventing hypertension (17). Furthermore, DL-propargylgycine (PPG), a CSE inhibitor, was reported to increase BP in Wistar-Kyoto rats and to promote vascular remodeling, indicating the potential regulatory role of CSE in maintaining normal BP (18). Interestingly, in another study, treatment of Sprague-Dawley rats with CSE inhibitor, DL-propargylglycine (PAG) or CBS inhibitor, aminooxyacetic acid alone, did not alter the BP levels, while treatment with both inhibitors would significantly increase mean arterial pressure. This finding could partly explain the interaction of different H2S producing enzymes in regulating BP (19).
As important gas transmitters, H2S and NO share crosstalk in regulating pathological and physical conditions (20, 21). In hypertension, endogenous H2S production regulated by CSE, inhibits endogenous endothelial NO bioavailability, therefore contributing to blood pressure control (22). Sodium nitroprusside (SNP), a NO donor, was reported to increase H2S production via upregulating the CSE or CBS activity, suggesting the crosstalk between the endogenous production of two gases (23). However, the details of two gases interaction regulating blood pressure remain to be elucidated.
Taken together, endogenous H2S production acts as an important physiological mediator that regulates BP homeostasis and H2S deficiency will contribute to the progress of HBP.
Exogenous H2S in hypertension
Apart from endogenous regulation of H2S in HBP, exogenous administration of H2S would regulate the process of HBP. The effects of exogenous H2S donors have been widely studied in animals in different experimental settings.
NaHS was the most widely used H2S donor for examining the effects in treating HBP. It is reported that early treatment with sodium hydrosulfide (NaHS) (14umol/Kg/day daily intraperitoneal injection for 4 weeks) was proved to prevent the transition from pre-hypertension to hypertension in spontaneously hypertensive rats (SHRs) (24). In another study, NaHS was reported to improve endothelial dysfunction by inhibiting the NLRP3 inflammasome and oxidative stress in SHRs. However, the protective effects were abolished by knocking out Nrf2 (25). The protective effects of NaHS was also testified in an Ang-II induced HBP model, suggesting the universal effects of H2S in treating HBP (26). Xiao and colleagues reported that 20-week administration of NaHS lowered the arterial pressure and increased the production of NO, enhancing eNOS phosphorylation through the activation of peroxisome proliferator-activated receptor/protein kinase B/AMP-activated protein kinase (PPAR-d/Akt/AMPK) signaling pathway (27). Administration of NaHS exerted anti-hypertensive effects, promoted non-NO-mediated relaxation, and decreased oxidative stress in rats with plumbum-induced hypertension (14). Injection of NaHS was demonstrated to ameliorate soluble FMS-like tyrosine kinase 1 (sFlt1)-induced hypertension, proteinuria, and glomerular endotheliosis in rats by increasing vascular endothelial growth factor (VEGF) expression (28). NaHS administration in SHRs were proven to reduce hypertensive related inflammation, partly through regulation of T cell subsets balance by Connexin (Cx) 40/Cx43 expressions inhibition (29). These experiment results demonstrate that NaHS dramatically suppressed the progression of HBP in different experimental settings via different mechanisms.
GYY4137 was synthesized in 2018, characterized as a novel, water-soluble and long-releasing hydrogen sulfide-releasing molecule (30). GYY4137 has been reported to have anti-hypertensive effects, due to upregulating the expression of VEGFR2 (31). In another study, GYY4137 reversed blood pressure increase after Ang-II inducement, which was accompanied by upregulation of microRNA-129 (32). Exogenous GYY4137 supplementation in the paraventricular nucleus (PVN) attenuated sympathetic activity and hypertensive response, partly due to decrease of reactive oxygen species (ROS) and pro-inflammatory cytokines within the PVN in high salt-induced hypertension (33).
Apart from GYY4137 and NaHS, various H2S releasing organic compounds have been shown to exhibit protective effects in treating HBP. Allicin, which comprised a variety of sulfur-containing compounds, has been reported to exert anti-hypertensive effects in an endothelium dependent pathway (34). Sodium thiosulfate, a reversible oxidation product of H2S, has vasodilating and anti-oxidative properties in a N-ω-nitro-L-arginine (L-NNA) induced hypertension model (35). N-phenylthiourea (PTU) and N,N’-diphenylthiourea (DPTU) compounds have been investigated as potential H2S-donors, and also demonstrated typical H2S-mediated vascular properties (36). This experimental evidence advocates more extensive discovery of new H2S donors to exert more extensive application in treating HBP.
H2S does share interplay with NO and CO, regulating the pathogenesis of HBP. H2S and NO are both vasodilating mediators. H2S donors were reported to induce vasorelaxation and promote NO-donor induced vasorelaxation in rat thoracic aorta, showing the possible interaction between NO and H2S in vascular regulation (37). In L-NAME induced hypertensive rats a dysfunctional H2S pathway was revealed and exogenous H2S attenuated the elevated blood pressure in this model (38). Reducing CO levels in Brown-Norway rats increases H2S generation and prevents hypoxia-induced pulmonary edema. Increasing CO levels in SHR has been found to enhance carotid H2S generation, prevent hypersensitivity to hypoxia and control hypertension in SHR (39). H2S has also been demonstrated to exert protective effects for acute CO poisoning patients (40).
As for the mechanisms of H2S in regulating HBP, they share similar functions and several similar pathways to regulate hypertension. The possible mechanisms of H2S on vascular tone include: KATP- channel dependent relaxation, other K+ channels, PKG activation, hyperpolarization, eNOS inhibition, inhibition of cytochrome C oxidase and anti-oxidant effects (21, 41–47).
H2S not only exert anti-hypertensive effects in systematic hypertension, it also has a regulatory role in pulmonary hypertension (PHT). PHT is characterized by blood pressure increased in pulmonary artery, associated with high incidence of mortality and morbidity (48). H2S was reported to exert anti-hypertensive effects in pulmonary hypertension via vaso-relaxative actions (49, 50). H2S has been proven to effectively inhibit hypoxia-induced increase in cell proliferation, migration, and oxidative stress in pulmonary artery smooth muscle cells (PASMCs) in an endoplasmic reticulum (ER) -dependent pathway, therefore exerting protective effects in PHT (51). Rashid et al. demonstrated that the relaxation response of to NaHS in porcine lungs was reduced in the presence of a high concentration of K+, indicating that the mechanism of relaxation depends, in part, on K+ channel activity (52). Du group showed that H2S treatment attenuated the oxidative stress accompanied by PHT, by reducing oxidized glutathione content (53). It was also reported that endogenous sulfur dioxide pathway was down-regulated in rats with PHT, indicating the involvement of sulfur dioxide/aspartate aminotransferase 2 pathway (54).
To summarize, due to the complexity of HBP management and lack of adequate therapy, H2S is gaining increasingly attention as a potential therapeutic target. Therefore, we summarized the role of H2S in regulating hypertension in Table 1. However, the effects and mechanisms by which H2S regulates HBP are complicated and still remaining largely unknown.
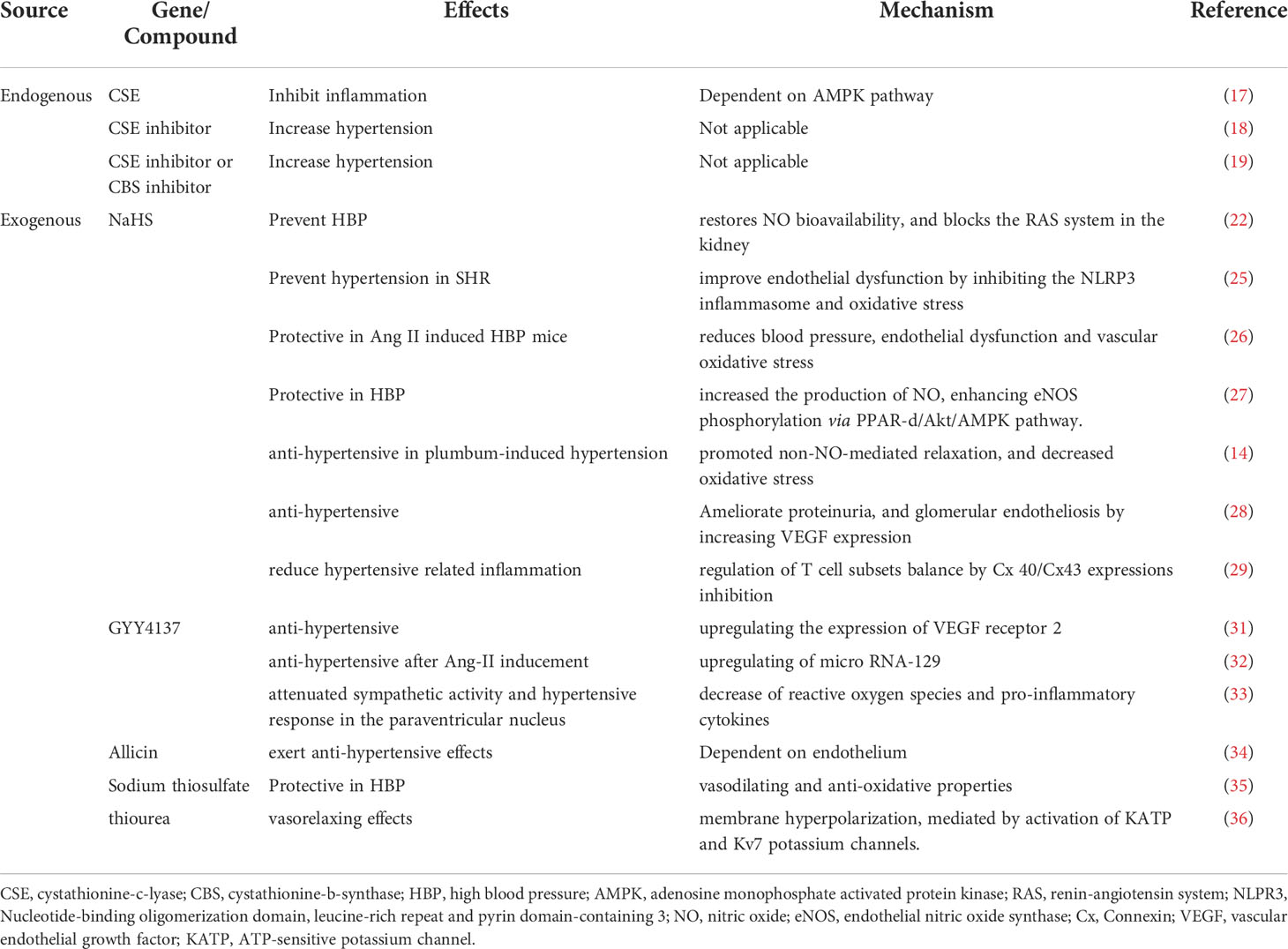
Table 1 Role of H2S in Regulating Hypertension.
H2S and atherosclerosis
Atherosclerosis (AS) is a long-term, chronic inflammatory disease of the vessel wall, which is widely recognized as a high risk for cardiovascular diseases (55). The progression of AS is extremely complex, involving numerous pathophysiological processes, including endothelial dysfunction, oxidative stress, inflammation, vascular smooth muscle cell proliferation and migration (56).
H2S has been reported to be a vaso-relaxant agent, which processes the property of ameliorating vascular dysfunction and mitigating the progression of AS. The potential therapeutic effects in anti-AS include maintaining endothelial cell dysfunction, inhibiting inflammation, suppressing vascular smooth muscle cell (VSMC) proliferation, migration and mitigating oxidative stress (57). However, the mechanisms of H2S to be protective against AS have not been fully elucidated and the therapeutic potential of H2S for AS treatment needs further exploration. Herein, we will review the recent findings of H2S in anti-AS from two main perspectives: endothelial cell dysfunction and inflammation.
Endothelial cell dysfunction
The endothelial cell represents a fundamental barrier for the maintenance of vascular homeostasis. Dysfunction in the endothelium may lead to several cardiovascular diseases (58, 59). Therefore, protecting the vascular endothelium from damage is one of the key factors against AS and AS related disorders.
Endogenous regulation of H2S are observed to play a role in regulating AS. The concentration of H2S was found to be decreased in AS mice, indicating the potential regulatory role of H2S in AS (60). As mentioned previously, the synthesis of H2S are regulated by 3 enzymes: CBS, CSE, and 3-MST. Loss of enzyme functions may lead to endothelial dysfunction in AS. For example, CSE/H2S pathway is reported to involve in AS via the H2S/CSE-TXNIP-NLRP3-IL-18/IL-1β-nitric oxide (NO) signaling pathway (61). Furthermore, Tian and colleagues observed that H2S deficiency derived from CSE depletion contributes to the development of endothelial dysfunction. In their study, MAPK/TXNIP (thioredoxin interacting protein signaling) is positively involved in CSE/H2S deficiency-associated endothelial dysfunction (62). CSE/H2S pathway may be protective against the formation of uremia accelerated atherosclerosis (UAAS) by affecting the expression of downstream molecule endothelial nitric oxide synthase (eNOS), which may be mediated by conventional protein kinase C (PKC) βII/Akt signaling pathway (63). Besides CSE, CBS was also observed to play a role in the process of AS. Mutations in the CBS gene are known to cause endothelial dysfunction responsible for cardiovascular and neurovascular diseases, and CBS/H2S pathway interacts with mitochondrial function and ER-mitochondrial tethering, therefore interfering with endothelial cell dysfunction-related pathologies (64). However, the role of 3-MST in maintain endothelial cell function in AS needs to be investigated. Collectively, the level of H2S and CSE/CSB/3-MST level can be considered as potential biomarkers and therapeutic targets for AS patients.
Numerous studies have demonstrated that exogenous H2S supplementation is another source contributing to the anti-AS effects. For instance, NaHS was proved to be protective against AS by upregulating angiotensin converting enzyme 2 (ACE2) expression in endothelial cells (65). Besides, H2S can reverse the endothelial dysfunction induced by AngII in HUVECs by ER stress pathway (66). Furthermore, H2S can enhance activator protein 1 (AP-1 binding) activity with the sirtuins 3 (SIRT3) promoter, thereby upregulating SIRT3 expression and ultimately reducing oxidant-provoked vascular endothelial dysfunction (67). Also Ford reported that NaHS treatment significantly reduced endothelial dysfunction and inhibited vascular superoxide generation in high-fat diet ApoE(-/-) mice, and therefore impaired atherosclerotic lesion development (68).
Apart from supplementation of traditional donors, administration of organic H2S donors would also be protective in maintaining EC function against AS. GYY4137 can induce autophagy and can protect ECs from Ox-LDL-induced apoptosis by activating Sirt1 (69). AP39 and AP123, the newly synthesized mitochondria-target H2S donors, are reported to protect endothelial cells from highglycemia-induced injury via preserving mitochondria function (70). With the development of pharmacy technology, the synthesis of new H2S releasing compounds are promising against AS.
There is crosstalk between H2S and NO in regulating the pathogenesis of AS. H2S was reported to increase NO production and upregulated the expression of inducible nitric oxide synthase(iNOS) (71). ApoE-/- mice fed with PAG was found with enhanced atherosclerotic lesion area, and with decreased NO levels, suggesting H2S could regulate atherosclerosis progression through NO crosstalk. H2S partially restores aortic endothelium-dependent relaxation in ApoE-/- mice, which may be related to increased phosphorylation of eNOS in the aorta (72).
To summarize, large numbers of studies have demonstrated the protective role of H2S in anti-AS via maintaining normal EC function, however, the mechanisms need deeper understanding. As a result, this will further facilitate the development of drug therapy for treating AS.
Inflammation
AS is a chronic vascular inflammatory disease and inflammation exists at all stages of AS (73). H2S has been reported to have anti-inflammatory effects, further regulating the pathogenesis of AS. Deeper understating of the protective effects of H2S donors via inhibiting inflammation will help provide a new way for future AS treatment.
Endogenous H2S production has been reported to regulate inflammation in AS by its producing enzymes. Alterations of CSE/H2S pathway may thus be involved in atherosclerosis pathogenesis (74). However, the underlying mechanisms are poorly understood. Endogenous CSE/H2S can directly sulfhydrate SIRT1, promote its deacetylation activity, and increase SIRT1 stability, thus reducing atherosclerotic plaque formation, by reducing vascular related inflammation (75). In another study, zofenopril at, the active metabolite of zofenopril has been reported to exert anti-inflammatory activity in vascular cells through its ability to increase H2S availability, therefore providing a potential target for treating AS (76). Moreover, CSE/H2S pathway has been reported to play an anti-inflammatory role in oxidized low-density lipoprotein (ox-LDL)-stimulated macrophage by suppressing c-Jun N-terminal kinase (JNK)/NF-κB signaling pathway (74). Furthermore, high fat diet is a predisposing factor for the progression of AS. It is reported that high fat diet might cause impaired function of CSE/H2S pathway, aggravating inflammation and posing risks to the development of AS (77). Apart from CSE regulation pathway, it is shown that deletion of CBS would impair endogenous H2S production and promote inflammatory reaction in AS-susceptible mice (78). This provided evidence that H2S releasing diet may help protect against AS.
Apart from endogenous H2S in regulating AS related inflammation, exogenous H2S administration also had an important role in AS. NaHS was originally the most widely used H2S donor in studying the effects of H2S in anti-AS. Numerous studies have demonstrated NaHS to be protective against AS by reducing inflammation (74, 75, 79, 80). In addition to traditional H2S releasing salts, new synthesized H2S donors have shown great potential with physiological properties. Whiteman and et al. demonstrated that GYY4137 could significantly inhibit lipopolysaccharide (LPS) -induced release of pro-inflammatory mediators and promoted the release of the anti-inflammatory chemokines. While NaHS exerted a bidirectional effect at high concentrations. This finding can partly explain the complex regulation system of H2S in inflammation (81). GYY4137, has also been proved to be protective against the development of diabetes-accelerated AS by preventing the activation of NLPR3 inflammasome (82). Furthermore, H2S rich compounds are reported to upregulate the expression of glutathione (GSH) and glutamate-cysteine ligase catalytic (GCLC) subunit, inhibiting inflammation, and exerting beneficial effects of mitigating AS (83). The effects of endogenous H2S and exogenous H2S in AS were listed in Table 2.

Table 2 Effects of Endogenous H2S and Exogenous H2S in AS.
H2S and angiogenesis
Angiogenesis is a process of new vessel formation from the existing vasculature (84). It is found that H2S might be a pro-angiogenic factor, promoting angiogenesis in different diseases and increase the expression of angiogenesis related biomarkers, including diabetes mellitus (DM), ischemic diseases and cancer (85).
H2S and DM related angiogenesis
DM is the leading cause of mortality worldwide, causing a variety of vascular complications (86). Impaired angiogenesis is a strong feature of DM and it can commonly induce refractory wound lesions. Therefore, promoting angiogenesis is of crucial importance for DM patients.
DM patients are reported with lower concentration of H2S in serum and in curtenous tissues, indicating the impaired synthesis of H2S production in DM patients (87, 88). Therefore, regulation of endogenous H2S production and production enzymes are a potential treatment for DM related wound healing. CSE down-regulation is reported to play a role in the pathogenesis of diabetic impaired wound healing (89). Danhong, a traditional Chinese herb medicine, has been reported to promote angiogenesis in the diabetic hind limb ischemia model through activation of local CSE-H2S-vascular endothelial growth factor (VEGF) axis (90). Furthermore, DM leads to the dysfunction of 3-MST/H2S and 3-MST might be a therapeutic target for DM patients (91). Besides, 3-MST/H2S axis was also reported to exert pro-angiogeneic effects via modulating mitochondrial respiration and increasing mitochondrial adenosine triphosphate (ATP) production (92).
Numerous studies have proved H2S to be a pro-angiogenetic factor (89, 93, 94). For example, H2S has been reported to increase angiogenesis in injured ischemic adductor muscle and to promote the ischemic diabetic wound healing in type 2 diabetic db/db mice (95). H2S improves wound healing by restoration of endothelial progenitor cell (EPC) functions and activation of Ang-1 in type 2 diabetic mice (94). H2S can also improve diabetic impaired wound healing by attenuating inflammation and increasing angiogenesis (96). These findings together imply that H2S played a role in DM mediated angiogenesis.
Apart from traditional widely-know H2S releasing donors, new and effective donors containing H2S moiety have been synthesized and utilized in DM related diseases. HA-JK1 and SA/JK-1 have been synthesized as examples. For HA-JK1, an in situ forming biomimetic hyaluronic acid (HA) hydrogel was used as a matrix to dope a pH-controllable H2S donor, JK1, to form a novel HA-JK1 hybrid system. This HA-JK1 hydrogel was designed as an ideal delivery scaffold for JK1 with pH-dependent prolonged H2S releasing profile (97). For SA/JK-1, which was capable of releasing H2S consistently under acidic pH conditions by absorbing exudate at the wound interface. The SA/JK-1 sponge exhibited biocompatibility to fibroblasts and promoted cell migration in vitro, and exhibited obviously positive influence on wound healing, therefore providing an effective treatment for non-healing wound (98). Interestingly, microparticles containing NaHS, have been synthesized using the emulsion technique, called NaHS@MPs. It can sustainably release H2S under physiological conditions and promote angiogenesis, further accelerating the healing of full-thickness wounds in diabetic mice (99).
Collectively, the role of H2S in DM related angiogenesis is gaining increasingly attention. However, there remains large space to be explored to clinical practice.
H2S and angiogenesis in ischemic diseases
Ischemic diseases are accompanied by shortage of blood supply. Angiogenesis would potentially increase the blood flow, therefore exerting the treating effects.
Modulation of endogenous H2S generation has a role in angiogenesis. CBS, CSE and 3-MST responded differently to angiogenesis. CSE is reported to promote VEGF-dependent angiogenesis through H2S generation under amino acid restriction (100). However, in another study, Tao and et al. found that CBS could promote vascular endothelial cell migration both under normoxic and minor hypoxia conditions (10% oxygen), while CSE had the opposite effects. 3-MST can accelerate the migration of endothelial cells in hypoxia, while no such effect was observed under normoxic conditions. They further found that 3-MST can modulate the endothelial cell migration, rather than CSE or CBS. Their study highlighted the need to get deeper understanding of the different functions of the H2S producing enzymes under different conditions (101). Furthermore, thiosulfate, one of the products formed during oxidative H2S metabolism, has surprisingly demonstrated inhibitory effects on VEGF-dependent endothelial cell proliferation, combined with reduction of CSE expression level (102). Therefore, the role of endogenous H2S on angiogenesis is controversial and requires more study to elucidate the potential mechanisms.
GYY4137 was reported to promote HHcy-mediated neoangiogenesis impairment in the ischemic hind limbs of post femoral artery ligation model via peroxisome proliferator-activated receptor (PPAR)-γ/VEGF axis (103). DATS, an organic polysulfide releasing H2S, has been demonstrated to promote angiogenesis in hindlimb ischemia via Akt-eNOS signaling pathway (104). Furthermore, NaHS could increase NO bioavailability and promote angiogenesis in ischemia hindlimb (105). NaHS exert proangiogeneic effect mediated by interaction between the upregulated VEGF in the skeletal muscle cells and the VEGF receptor 2 (106). In another report, NaHS exerts pro-angiogeneic effects through dependent on activation of Akt (107).
Recently, with the development of material synthesis technology, various H2S releasing compounds have been synthesized to enhance H2S releasing properties. For instance, A poly (D, L-lactic-co-glycolic acid) microparticle system that contains DATS, called DATS@MPs, possess the property of slow and long-term H2S release. DATS@MPs have been reported to promote therapeutic angiogenesis in an ischemic mouse limb model through activating nuclear respiratory factor 2 (Nrf2) translocation, thus providing therapeutic potential in treating ischemic diseases (108). Moreover, ZYZ-803, a novel synthetic H2S-NO hybrid molecule, which can slowly release H2S and NO, has been reported to exert pro-angiogenetic effects via SIRT1 dependent pathway. The pro-angiogenetic effects of H2S are also dependent on CSE and eNOS expression via cross-talk between signal transducer and activator of transcription 3 (STAT3) and Ca2+/CaM-dependent protein kinase II (CaMKII) activation (109, 110).
Apart from ischemic limb diseases, myocardial infarction (MI) is another serious ischemic disease, which poses great danger to people’s health. GYY4137 was reported to exert pro-angiogenic effects following MI via endogenous natriuretic peptide activation (111). Diallyl trisulfide, a long-lasting H2S donor, can mitigate left ventricular dysfunction via inducing angiogenesis in over-loaded heart failure (112). NaHS was reported to increase angiogenesis and improve left ventricular function after MI (113). Besides, NaHS was also reported to promote angiogenesis, and mitigating the progression of heart failure by inducing matrix metalloproteinase (MMP)-2 activation and inhibiting MMP-9 and tissue inhibitor of matrix metalloproteinase (TIMP)-3 expression (114).
Newly and novel H2S releasing compounds have been synthesized, with the aim to over the limitations of traditional H2S releasing donors. Liang and et al. developed a macromolecular H2S prodrug. The compound comprised of a 2-aminopyridine-5-thiocarboxamide (a small-molecule H2S donor) on partially oxidized alginate (ALG-CHO), to obtain the slow and continuous release of endogenous H2S. They further formed a stem cell-loaded conductive H2S-releasing hydrogel through the Schiff base reaction between ALG-CHO and gelatin. They utilized the hydrogel in treating MI, demonstrating a dramatical improvement of the cardiac functions in rats (115). Moreover, S-Propargyl-Cysteine (SPRC), a novel water-soluble modulator of endogenous H2S production, has been demonstrated to exhibit pro-angiogenetic effects via the activation of STAT3. SPRC therefore provides a novel therapeutic strategy for ischemia heart diseases (116).
H2S and NO shared interactions in regulating angiogenesis. Aortic rings harvested from eNOS−/− mice exhibited no microvessel outgrowth in response to NaHS, compared with wild-type controls, demonstrating that NO was essential for the pro-angiogenic effect of H2S. Besides, chemical inhibition of CSE attenuated NO-mediated cGMP angiogenesis (44). Apart from this, NO donors increased CSE dependent H2S biogenesis in a cGMP-dependent manner. Pre-treating NO donors increased CSE mRNA and protein levels in smooth muscle cells increased H2S production (117). Taken together, NO and H2S contributed mutually in regulating angiogenesis.
In summary, H2S plays an important role in different vascular diseases. The structure of normal artery consisted of 3 layers. The inner layer lined by a monolayer of ECs is closely contacted with blood; the middle layer composed of VSMCs is located at the complex extracellular matrix; and the outer layer of arteries is composed of mast cells, nerve endings, and microvessels. Imbalance and dysfunction of the 3 layers lead to the pathogenesis of vascular diseases, especially dysfunction of EC and SMC (7). This indicates the universal functions of H2S in regulating different vascular diseases. Studied have focused on the effects of H2S from endogenous H2S production and exogenous H2S administration. However, the application of H2S in vascular diseases is still in the basic research stage. Studies and experiments of H2S in treating vascular diseases are required.
Future research should focus on the role and mechanism of H2S and different H2S releasing donors in treating vascular diseases. Synthetic H2S donors have been developed to overcome the disadvantages of traditional H2S donors. They can be categorized by their class of triggering mechanisms, possessing their specific delivery system and H2S releasing properties (118). Continuous improvements in the interaction and crosstalk between different gas transmitters in the control of vascular diseases. Exploring the therapeutic potential in regulating vascular diseases will be promising in the near future.
Author contributions
CZ and QL wrote the first draft. XL, RW, TG, XZ and BL provided the organization and framework of the article. KL and RC provided critical revisions. All authors contributed to the article and approved the submitted version.
Funding
The study was supported by Natural Science Foundation of China (82000390), China Postdoctoral Science Foundation (2020M681048), and Special Fund of Jilin Province (2020SC2T038). Jilin Provincial Finance Department, China (2020SCZT091). The Jilin Science and Technology Agency funds in China, China (20210402003GH).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Li L, Rose P, Moore PK. Hydrogen sulfide and cell signaling. Annu Rev Pharmacol Toxicol (2011) 51:169–87. doi: 10.1146/annurev-pharmtox-010510-100505
2. Mustafa AK, Gadalla MM, Sen N, Kim S, Mu W, Gazi SK, et al. H2S signals through protein s-sulfhydration. Sci Signal (2009) 2:ra72. doi: 10.1126/scisignal.2000464
3. Wallace JL, Wang R. Hydrogen sulfide-based therapeutics: exploiting a unique but ubiquitous gasotransmitter. Nat Rev Drug Discov (2015) 14:329–45. doi: 10.1038/nrd4433
4. Lv B, Chen S, Tang C, Jin H, Du J and Huang Y. Hydrogen sulfide and vascular regulation - an update. J Adv Res (2021) 27:85–97. doi: 10.1016/j.jare.2020.05.007
5. Yuan S, Kevil CG. Nitric oxide and hydrogen sulfide regulation of ischemic vascular remodeling. Microcirculation (2016) 23:134–45. doi: 10.1111/micc.12248
6. Yang G, Wang R. H2S and blood vessels: An overview. Handb Exp Pharmacol (2015) 230:85–110. doi: 10.1007/978-3-319-18144-8_4
7. Kanagy NL. Szabo c and papapetropoulos a. vascular biology of hydrogen sulfide. Am J Physiol Cell Physiol (2017) 312:C537–49. doi: 10.1152/ajpcell.00329.2016
8. Tian X, Zhou D, Zhang Y, Song Y, Zhang Q, Bu D, et al. Persulfidation of transcription factor FOXO1 at cysteine 457: A novel mechanism by which H2S inhibits vascular smooth muscle cell proliferation. J Adv Res (2021) 27:155–64. doi: 10.1016/j.jare.2020.06.023
9. Dong N, Piao H, Li B, Xu J, Wei S, Liu K. Poor management of hypertension is an important precipitating factor for the development of acute aortic dissection. J Clin Hypertens (Greenwich) (2019) 21:804–12. doi: 10.1111/jch.13556
10. Mancia G, Fagard R, Narkiewicz K, Redon J, Zanchetti A, Bohm M, et al. 2013 ESH/ESC guidelines for the management of arterial hypertension: the task force for the management of arterial hypertension of the European society of hypertension (ESH) and of the European society of cardiology (ESC). J Hypertens (2013) 31:1281–357. doi: 10.1097/01.hjh.0000431740.32696.cc
11. Kutz JL, Greaney JL, Santhanam L, Alexander LM. Evidence for a functional vasodilatatory role for hydrogen sulphide in the human cutaneous microvasculature. J Physiol (2015) 593:2121–9. doi: 10.1113/JP270054
12. Sun NL, Xi Y, Yang SN, Ma Z, Tang CS. Plasma hydrogen sulfide and homocysteine levels in hypertensive patients with different blood pressure levels and complications. Zhonghua Xin Xue Guan Bing Za Zhi (2007) 35:1145–8.
13. Shi MM, Wang JL, Zhang LQ, Qin M, Huang YW. Association between hydrogen sulfide and OSA-associated hypertension: a clinical study. Sleep Breath (2020) 24:745–50. doi: 10.1007/s11325-019-01997-y
14. Possomato-Vieira JS, Goncalves-Rizzi VH, do Nascimento RA, Wandekin RR, Caldeira-Dias M, Chimini JS, et al. Clinical and experimental evidences of hydrogen sulfide involvement in lead-induced hypertension. BioMed Res Int (2018) 2018:4627391. doi: 10.1155/2018/4627391
15. Aminzadeh MA, Vaziri ND. Downregulation of the renal and hepatic hydrogen sulfide (H2S)-producing enzymes and capacity in chronic kidney disease. Nephrol Dial Transplant (2012) 27:498–504. doi: 10.1093/ndt/gfr560
16. Peleli M, Bibli SI, Li Z, Chatzianastasiou A, Varela A, Katsouda A, et al. Cardiovascular phenotype of mice lacking 3-mercaptopyruvate sulfurtransferase. Biochem Pharmacol (2020) 176:113833. doi: 10.1016/j.bcp.2020.113833
17. Cui C, Fan J, Zeng Q, Cai J, Chen Y, Chen Z, et al. CD4(+) T-cell endogenous cystathionine gamma lyase-hydrogen sulfide attenuates hypertension by sulfhydrating liver kinase B1 to promote T regulatory cell differentiation and proliferation. Circulation (2020) 142:1752–69. doi: 10.1161/CIRCULATIONAHA.119.045344
18. Yan H, Du J, Tang C. The possible role of hydrogen sulfide on the pathogenesis of spontaneous hypertension in rats. Biochem Biophys Res Commun (2004) 313:22–7. doi: 10.1016/j.bbrc.2003.11.081
19. Roy A, Khan AH, Islam MT, Prieto MC, Majid DS. Interdependency of cystathione gamma-lyase and cystathione beta-synthase in hydrogen sulfide-induced blood pressure regulation in rats. Am J Hypertens (2012) 25:74–81. doi: 10.1038/ajh.2011.149
20. Li L, Hsu A, Moore PK. Actions and interactions of nitric oxide, carbon monoxide and hydrogen sulphide in the cardiovascular system and in inflammation–a tale of three gases! Pharmacol Ther (2009) 123:386–400. doi: 10.1016/j.pharmthera.2009.05.005
21. Ali MY, Ping CY, Mok YY, Ling L, Whiteman M, Bhatia M, et al. Regulation of vascular nitric oxide in vitro and in vivo; a new role for endogenous hydrogen sulphide? Br J Pharmacol (2006) 149:625–34. doi: 10.1038/sj.bjp.0706906
22. Szijarto IA, Marko L, Filipovic MR, Miljkovic JL, Tabeling C, Tsvetkov D, et al. Cystathionine gamma-Lyase-Produced hydrogen sulfide controls endothelial NO bioavailability and blood pressure. Hypertension (2018) 71:1210–7. doi: 10.1161/HYPERTENSIONAHA.117.10562
23. Zhao W, Ndisang JF, Wang R. Modulation of endogenous production of H2S in rat tissues. Can J Physiol Pharmacol (2003) 81:848–53. doi: 10.1139/y03-077
24. Tain YL, Hsu CN, Lu PC. Early short-term treatment with exogenous hydrogen sulfide postpones the transition from prehypertension to hypertension in spontaneously hypertensive rat. Clin Exp Hypertens (2018) 40:58–64. doi: 10.1080/10641963.2017.1313847
25. Li J, Teng X, Jin S, Dong J, Guo Q, Tian D, et al. Hydrogen sulfide improves endothelial dysfunction by inhibiting the vicious cycle of NLRP3 inflammasome and oxidative stress in spontaneously hypertensive rats. J Hypertens (2019) 37:1633–43. doi: 10.1097/HJH.0000000000002101
26. Al-Magableh MR, Kemp-Harper BK, Hart JL. Hydrogen sulfide treatment reduces blood pressure and oxidative stress in angiotensin II-induced hypertensive mice. Hypertens Res (2015) 38:13–20. doi: 10.1038/hr.2014.125
27. Xiao L, Dong JH, Teng X, Jin S, Xue HM, Liu SY, et al. Hydrogen sulfide improves endothelial dysfunction in hypertension by activating peroxisome proliferator-activated receptor delta/endothelial nitric oxide synthase signaling. J Hypertens (2018) 36:651–65. doi: 10.1097/HJH.0000000000001605
28. Holwerda KM, Burke SD, Faas MM, Zsengeller Z, Stillman IE, Kang PM, et al. Hydrogen sulfide attenuates sFlt1-induced hypertension and renal damage by upregulating vascular endothelial growth factor. J Am Soc Nephrol (2014) 25:717–25. doi: 10.1681/ASN.2013030291
29. Ni X, Zhang L, Peng M, Shen TW, Yu XS, Shan LY, et al. Hydrogen sulfide attenuates hypertensive inflammation via regulating connexin expression in spontaneously hypertensive rats. Med Sci Monit (2018) 24:1205–18. doi: 10.12659/MSM.908761
30. Li L, Whiteman M, Guan YY, Neo KL, Cheng Y, Lee SW, et al. Characterization of a novel, water-soluble hydrogen sulfide-releasing molecule (GYY4137): new insights into the biology of hydrogen sulfide. Circulation (2008) 117:2351–60. doi: 10.1161/CIRCULATIONAHA.107.753467
31. Zhu ML, Zhao FR, Zhu TT, Wang QQ, Wu ZQ, Song P, et al. The antihypertension effect of hydrogen sulfide (H2S) is induced by activating VEGFR2 signaling pathway. Life Sci (2021) 267:118831. doi: 10.1016/j.lfs.2020.118831
32. Weber GJ, Pushpakumar SB, Sen U. Hydrogen sulfide alleviates hypertensive kidney dysfunction through an epigenetic mechanism. Am J Physiol Heart Circ Physiol (2017) 312:H874–85. doi: 10.1152/ajpheart.00637.2016
33. Liang YF, Zhang DD, Yu XJ, Gao HL, Liu KL, Qi J, et al. Hydrogen sulfide in paraventricular nucleus attenuates blood pressure by regulating oxidative stress and inflammatory cytokines in high salt-induced hypertension. Toxicol Lett (2017) 270:62–71. doi: 10.1016/j.toxlet.2017.02.004
34. Cui T, Liu W, Chen S, Yu C, Li Y, Zhang JY. Antihypertensive effects of allicin on spontaneously hypertensive rats via vasorelaxation and hydrogen sulfide mechanisms. BioMed Pharmacother (2020) 128:110240. doi: 10.1016/j.biopha.2020.110240
35. Nguyen ITN, Klooster A, Minnion M, Feelisch M, Verhaar MC, van Goor H, et al. Sodium thiosulfate improves renal function and oxygenation in l-NNA-induced hypertension in rats. Kidney Int (2020) 98:366–77. doi: 10.1016/j.kint.2020.02.020
36. Citi V, Martelli A, Bucci M, Piragine E, Testai L, Vellecco V, et al. Searching for novel hydrogen sulfide donors: The vascular effects of two thiourea derivatives. Pharmacol Res (2020) 159:105039. doi: 10.1016/j.phrs.2020.105039
37. Hosoki R, Matsuki N, Kimura H. The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochem Biophys Res Commun (1997) 237:527–31. doi: 10.1006/bbrc.1997.6878
38. Zhong G, Chen F, Cheng Y, Tang C, Du J. The role of hydrogen sulfide generation in the pathogenesis of hypertension in rats induced by inhibition of nitric oxide synthase. J Hypertens (2003) 21:1879–85. doi: 10.1097/00004872-200310000-00015
39. Peng YJ, Makarenko VV, Nanduri J, Vasavda C, Raghuraman G, Yuan G, et al. Inherent variations in CO-H2S-mediated carotid body O2 sensing mediate hypertension and pulmonary edema. Proc Natl Acad Sci U S A (2014) 111:1174–9. doi: 10.1073/pnas.1322172111
40. Yu YP, Li ZG, Wang DZ, Zhan X, Shao JH. Hydrogen sulfide as an effective and specific novel therapy for acute carbon monoxide poisoning. Biochem Biophys Res Commun (2011) 404:6–9. doi: 10.1016/j.bbrc.2010.11.113
41. Mustafa AK, Sikka G, Gazi SK, Steppan J, Jung SM, Bhunia AK, et al. Hydrogen sulfide as endothelium-derived hyperpolarizing factor sulfhydrates potassium channels. Circ Res (2011) 109:1259–68. doi: 10.1161/CIRCRESAHA.111.240242
42. Jiang B, Tang G, Cao K, Wu L, Wang R. Molecular mechanism for H(2)S-induced activation of K(ATP) channels. Antioxid Redox Signal (2010) 12:1167–78. doi: 10.1089/ars.2009.2894
43. Bautista-Nino PK, van der Stel M, Batenburg WW, de Vries R, Roks AJM, Danser AHJ. Endothelium-derived hyperpolarizing factor and protein kinase G ialpha activation: H2O2 versus s-nitrosothiols. Eur J Pharmacol (2018) 827:112–6. doi: 10.1016/j.ejphar.2018.03.019
44. Coletta C, Papapetropoulos A, Erdelyi K, Olah G, Modis K, Panopoulos P, et al. Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endothelium-dependent vasorelaxation. Proc Natl Acad Sci U S A (2012) 109:9161–6. doi: 10.1073/pnas.1202916109
45. Kiss L, Deitch EA, Szabo C. Hydrogen sulfide decreases adenosine triphosphate levels in aortic rings and leads to vasorelaxation via metabolic inhibition. Life Sci (2008) 83:589–94. doi: 10.1016/j.lfs.2008.08.006
46. Hedegaard ER, Gouliaev A, Winther AK, Arcanjo DD, Aalling M, Renaltan NS, et al. Involvement of potassium channels and calcium-independent mechanisms in hydrogen sulfide-induced relaxation of rat mesenteric small arteries. J Pharmacol Exp Ther (2016) 356:53–63. doi: 10.1124/jpet.115.227017
47. Szabo C, Ransy C, Modis K, Andriamihaja M, Murghes B, Coletta C, et al. Regulation of mitochondrial bioenergetic function by hydrogen sulfide. part i. biochemical and physiological mechanisms. Br J Pharmacol (2014) 171:2099–122. doi: 10.1111/bph.12369
48. Nair A. Pharmacologic therapy for pulmonary artery hypertension. Curr Opin Cardiol (2020) 35:643–56. doi: 10.1097/HCO.0000000000000796
49. Ariyaratnam P, Loubani M, Morice AH. Hydrogen sulphide vasodilates human pulmonary arteries: a possible role in pulmonary hypertension? Microvasc Res (2013) 90:135–7. doi: 10.1016/j.mvr.2013.09.002
50. Castro-Piedras I, Perez-Zoghbi JF. Hydrogen sulphide inhibits Ca2+ release through InsP3 receptors and relaxes airway smooth muscle. J Physiol (2013) 591:5999–6015. doi: 10.1113/jphysiol.2013.257790
51. Wu J, Pan W, Wang C, Dong H, Xing L, Hou J, et al. H2S attenuates endoplasmic reticulum stress in hypoxia-induced pulmonary artery hypertension. Biosci Rep (2019) 39:1–13. doi: 10.1042/BSR20190304
52. Rashid S, Heer JK, Garle MJ, Alexander SP, Roberts RE. Hydrogen sulphide-induced relaxation of porcine peripheral bronchioles. Br J Pharmacol (2013) 168:1902–10. doi: 10.1111/bph.12084
53. Wei HL, Zhang CY, Jin HF, Tang CS, Du JB. Hydrogen sulfide regulates lung tissue-oxidized glutathione and total antioxidant capacity in hypoxic pulmonary hypertensive rats. Acta Pharmacol Sin (2008) 29:670–9. doi: 10.1111/j.1745-7254.2008.00796.x
54. Luo L, Liu D, Tang C, Du J, Liu AD, Holmberg L, et al. Sulfur dioxide upregulates the inhibited endogenous hydrogen sulfide pathway in rats with pulmonary hypertension induced by high pulmonary blood flow. Biochem Biophys Res Commun (2013) 433:519–25. doi: 10.1016/j.bbrc.2013.03.014
55. Taleb S. Inflammation in atherosclerosis. Arch Cardiovasc Dis (2016) 109:708–15. doi: 10.1016/j.acvd.2016.04.002
56. Liu Q, Zhang H, Lin J, Zhang R, Chen S, Liu W, et al. C1q/TNF-related protein 9 inhibits the cholesterol-induced vascular smooth muscle cell phenotype switch and cell dysfunction by activating AMP-dependent kinase. J Cell Mol Med (2017) 21:2823–36. doi: 10.1111/jcmm.13196
57. Wang ZJ, Wu J, Guo W, Zhu YZ. Atherosclerosis and the hydrogen sulfide signaling pathway - therapeutic approaches to disease prevention. Cell Physiol Biochem (2017) 42:859–75. doi: 10.1159/000478628
58. Lee DY, Chiu JJ. Atherosclerosis and flow: roles of epigenetic modulation in vascular endothelium. J BioMed Sci (2019) 26:56. doi: 10.1186/s12929-019-0551-8
59. Theodorou K, Boon RA. Endothelial cell metabolism in atherosclerosis. Front Cell Dev Biol (2018) 6:82. doi: 10.3389/fcell.2018.00082
60. Mani S, Li H, Untereiner A, Wu L, Yang G, Austin RC, et al. Decreased endogenous production of hydrogen sulfide accelerates atherosclerosis. Circulation (2013) 127:2523–34. doi: 10.1161/CIRCULATIONAHA.113.002208
61. Yue LM, Gao YM, Han BH. Evaluation on the effect of hydrogen sulfide on the NLRP3 signaling pathway and its involvement in the pathogenesis of atherosclerosis. J Cell Biochem (2019) 120:481–92. doi: 10.1002/jcb.27404
62. Tian D, Dong J, Jin S, Teng X, Wu Y. Endogenous hydrogen sulfide-mediated MAPK inhibition preserves endothelial function through TXNIP signaling. Free Radic Biol Med (2017) 110:291–9. doi: 10.1016/j.freeradbiomed.2017.06.016
63. Xiong R, Lu X, Song J, Li H, Wang S. Molecular mechanisms of hydrogen sulfide against uremic accelerated atherosclerosis through cPKCbetaII/Akt signal pathway. BMC Nephrol (2019) 20:358. doi: 10.1186/s12882-019-1550-4
64. Rao G, Murphy B, Dey A, Dwivedi SKD, Zhang Y, Roy RV, et al. Cystathionine beta synthase regulates mitochondrial dynamics and function in endothelial cells. FASEB J (2020) 34:9372–92. doi: 10.1096/fj.202000173R
65. Lin Y, Zeng H, Gao L, Gu T, Wang C, Zhang H. Hydrogen sulfide attenuates atherosclerosis in a partially ligated carotid artery mouse model via regulating angiotensin converting enzyme 2 expression. Front Physiol (2017) 8:782. doi: 10.3389/fphys.2017.00782
66. Hu HJ, Jiang ZS, Qiu J, Zhou SH, Liu QM. Protective effects of hydrogen sulfide against angiotensin II-induced endoplasmic reticulum stress in HUVECs. Mol Med Rep (2017) 15:2213–22. doi: 10.3892/mmr.2017.6238
67. Xie L, Feng H, Li S, Meng G, Liu S, Tang X, et al. SIRT3 mediates the antioxidant effect of hydrogen sulfide in endothelial cells. Antioxid Redox Signal (2016) 24:329–43. doi: 10.1089/ars.2015.6331
68. Ford A, Al-Magableh M, Gaspari TA, Hart JL. Chronic NaHS treatment is vasoprotective in high-Fat-Fed ApoE(-/-) mice. Int J Vasc Med (2013) 2013:915983. doi: 10.1155/2013/915983
69. Zhu L, Duan W, Wu G, Zhang D, Wang L, Chen D, et al. Protective effect of hydrogen sulfide on endothelial cells through Sirt1-FoxO1-mediated autophagy. Ann Transl Med (2020) 8:1586. doi: 10.21037/atm-20-3647
70. Gero D, Torregrossa R, Perry A, Waters A, Le-Trionnaire S, Whatmore JL, et al. The novel mitochondria-targeted hydrogen sulfide (H2S) donors AP123 and AP39 protect against hyperglycemic injury in microvascular endothelial cells in vitro. Pharmacol Res (2016) 113:186–98. doi: 10.1016/j.phrs.2016.08.019
71. Lin Y, Chen Y, Zhu N, Zhao S, Fan J, Liu E. Hydrogen sulfide inhibits development of atherosclerosis through up-regulating protein s-nitrosylation. BioMed Pharmacother (2016) 83:466–76. doi: 10.1016/j.biopha.2016.07.003
72. Liu Z, Han Y, Li L, Lu H, Meng G, Li X, et al. The hydrogen sulfide donor, GYY4137, exhibits anti-atherosclerotic activity in high fat fed apolipoprotein e(-/-) mice. Br J Pharmacol (2013) 169:1795–809. doi: 10.1111/bph.12246
73. Libby P. Inflammation in atherosclerosis. Arterioscler Thromb Vasc Biol (2012) 32:2045–51. doi: 10.1161/ATVBAHA.108.179705
74. Wang XH, Wang F, You SJ, Cao YJ, Cao LD, Han Q, et al. Dysregulation of cystathionine gamma-lyase (CSE)/hydrogen sulfide pathway contributes to ox-LDL-induced inflammation in macrophage. Cell Signal (2013) 25:2255–62. doi: 10.1016/j.cellsig.2013.07.010
75. Du C, Lin X, Xu W, Zheng F, Cai J, Yang J, et al. Sulfhydrated sirtuin-1 increasing its deacetylation activity is an essential epigenetics mechanism of anti-atherogenesis by hydrogen sulfide. Antioxid Redox Signal (2019) 30:184–97. doi: 10.1089/ars.2017.7195
76. Monti M, Terzuoli E, Ziche M, Morbidelli L. H2S dependent and independent anti-inflammatory activity of zofenoprilat in cells of the vascular wall. Pharmacol Res (2016) 113:426–37. doi: 10.1016/j.phrs.2016.09.017
77. Peh MT, Anwar AB, Ng DS, Atan MS, Kumar SD, Moore PK. Effect of feeding a high fat diet on hydrogen sulfide (H2S) metabolism in the mouse. Nitric Oxide (2014) 41:138–45. doi: 10.1016/j.niox.2014.03.002
78. Zhang D, Jiang X, Fang P, Yan Y, Song J, Gupta S, et al. Hyperhomocysteinemia promotes inflammatory monocyte generation and accelerates atherosclerosis in transgenic cystathionine beta-synthase-deficient mice. Circulation (2009) 120:1893–902. doi: 10.1161/CIRCULATIONAHA.109.866889
79. Xu S, Liu Z, Liu P. Targeting hydrogen sulfide as a promising therapeutic strategy for atherosclerosis. Int J Cardiol (2014) 172:313–7. doi: 10.1016/j.ijcard.2014.01.068
80. Liu YH, Lu M, Hu LF, Wong PT, Webb GD, Bian JS. Hydrogen sulfide in the mammalian cardiovascular system. Antioxid Redox Signal (2012) 17:141–85. doi: 10.1089/ars.2011.4005
81. Whiteman M, Li L, Rose P, Tan CH, Parkinson DB, Moore PK. The effect of hydrogen sulfide donors on lipopolysaccharide-induced formation of inflammatory mediators in macrophages. Antioxid Redox Signal (2010) 12:1147–54. doi: 10.1089/ars.2009.2899
82. Zheng Q, Pan L, Ji Y. H2S protects against diabetes-accelerated atherosclerosis by preventing the activation of NLRP3 inflammasome. J BioMed Res (2019) 34:94–102. doi: 10.7555/JBR.33.20190071
83. Jain SK, Huning L, Micinski D. Hydrogen sulfide upregulates glutamate-cysteine ligase catalytic subunit, glutamate-cysteine ligase modifier subunit, and glutathione and inhibits interleukin-1beta secretion in monocytes exposed to high glucose levels. Metab Syndr Relat Disord (2014) 12:299–302. doi: 10.1089/met.2014.0022
84. Carmeliet P. Mechanisms of angiogenesis and arteriogenesis. Nat Med (2000) 6:389–95. doi: 10.1038/74651
85. Terzuoli E, Monti M, Vellecco V, Bucci M, Cirino G, Ziche M, et al. Characterization of zofenoprilat as an inducer of functional angiogenesis through increased H2 s availability. Br J Pharmacol (2015) 172:2961–73. doi: 10.1111/bph.13101
86. Gregg EW, Williams DE, Geiss L. Changes in diabetes-related complications in the united states. N Engl J Med (2014) 371:286–7. doi: 10.1056/NEJMc1406009
87. Jain SK, Bull R, Rains JL, Bass PF, Levine SN, Reddy S, et al. Low levels of hydrogen sulfide in the blood of diabetes patients and streptozotocin-treated rats causes vascular inflammation? Antioxid Redox Signal (2010) 12:1333–7. doi: 10.1089/ars.2009.2956
88. Brancaleone V, Roviezzo F, Vellecco V, De Gruttola L, Bucci M, Cirino G. Biosynthesis of H2S is impaired in non-obese diabetic (NOD) mice. Br J Pharmacol (2008) 155:673–80. doi: 10.1038/bjp.2008.296
89. Cheng Z, Shen X, Jiang X, Shan H, Cimini M, Fang P, et al. Hyperhomocysteinemia potentiates diabetes-impaired EDHF-induced vascular relaxation: Role of insufficient hydrogen sulfide. Redox Biol (2018) 16:215–25. doi: 10.1016/j.redox.2018.02.006
90. Wu F, He Z, Ding R, Huang Z, Jiang Q, Cui H, et al. Danhong promotes angiogenesis in diabetic mice after critical limb ischemia by activation of CSE-h 2 s-VEGF axis. Evid Based Complement Alternat Med (2015) 2015:276263. doi: 10.1155/2015/276263
91. Coletta C, Modis K, Szczesny B, Brunyanszki A, Olah G, Rios EC, et al. Regulation of vascular tone, angiogenesis and cellular bioenergetics by the 3-mercaptopyruvate Sulfurtransferase/H2S pathway: Functional impairment by hyperglycemia and restoration by DL-alpha-Lipoic acid. Mol Med (2015) 21:1–14. doi: 10.2119/molmed.2015.00035
92. Abdollahi Govar A, Toro G, Szaniszlo P, Pavlidou A, Bibli SI, Thanki K, et al. 3-mercaptopyruvate sulfurtransferase supports endothelial cell angiogenesis and bioenergetics. Br J Pharmacol (2020) 177:866–83. doi: 10.1111/bph.14574
93. Yang HB, Liu HM, Yan JC, Lu ZY. Effect of diallyl trisulfide on ischemic tissue injury and revascularization in a diabetic mouse model. J Cardiovasc Pharmacol (2018) 71:367–74. doi: 10.1097/FJC.0000000000000579
94. Liu F, Chen DD, Sun X, Xie HH, Yuan H, Jia W, et al. Hydrogen sulfide improves wound healing via restoration of endothelial progenitor cell functions and activation of angiopoietin-1 in type 2 diabetes. Diabetes (2014) 63:1763–78. doi: 10.2337/db13-0483
95. Wang GG, Li W. Hydrogen sulfide improves vessel formation of the ischemic adductor muscle and wound healing in diabetic db/db mice. Iran J Basic Med Sci (2019) 22:1192–7. doi: 10.22038/ijbms.2019.36551.8709
96. Zhao H, Lu S, Chai J, Zhang Y, Ma X, Chen J, et al. Hydrogen sulfide improves diabetic wound healing in ob/ob mice via attenuating inflammation. J Diabetes Complications (2017) 31:1363–9. doi: 10.1016/j.jdiacomp.2017.06.011
97. Wu J, Chen A, Zhou Y, Zheng S, Yang Y, An Y, et al. Novel H2S-releasing hydrogel for wound repair via in situ polarization of M2 macrophages. Biomaterials (2019) 222:119398. doi: 10.1016/j.biomaterials.2019.119398
98. Zhao X, Liu L, An T, Xian M, Luckanagul JA, Su Z, et al. A hydrogen sulfide-releasing alginate dressing for effective wound healing. Acta Biomater (2020) 104:85–94. doi: 10.1016/j.actbio.2019.12.032
99. Lin WC, Huang CC, Lin SJ, Li MJ, Chang Y, Lin YJ, et al. In situ depot comprising phase-change materials that can sustainably release a gasotransmitter H2S to treat diabetic wounds. Biomaterials (2017) 145:1–8. doi: 10.1016/j.biomaterials.2017.08.023
100. Longchamp A, Mirabella T, Arduini A, MacArthur MR, Das A, Trevino-Villarreal JH, et al. Amino acid restriction triggers angiogenesis via GCN2/ATF4 regulation of VEGF and H2S production. Cell (2018) 173:117–29.e14. doi: 10.1016/j.cell.2018.03.001
101. Tao B, Wang R, Sun C, Zhu Y. 3-mercaptopyruvate sulfurtransferase, not cystathionine beta-synthase nor cystathionine gamma-lyase, mediates hypoxia-induced migration of vascular endothelial cells. Front Pharmacol (2017) 8:657. doi: 10.3389/fphar.2017.00657
102. Leskova A, Pardue S, Glawe JD, Kevil CG, Shen X. Role of thiosulfate in hydrogen sulfide-dependent redox signaling in endothelial cells. Am J Physiol Heart Circ Physiol (2017) 313:H256–64. doi: 10.1152/ajpheart.00723.2016
103. Majumder A, Singh M, George AK, Behera J, Tyagi N, Tyagi SC. Hydrogen sulfide improves postischemic neoangiogenesis in the hind limb of cystathionine-beta-synthase mutant mice via PPAR-gamma/VEGF axis. Physiol Rep (2018) 6:e13858. doi: 10.14814/phy2.13858
104. Hayashida R, Kondo K, Morita S, Unno K, Shintani S, Shimizu Y, et al. Diallyl trisulfide augments ischemia-induced angiogenesis via an endothelial nitric oxide synthase-dependent mechanism. Circ J (2017) 81:870–8. doi: 10.1253/circj.CJ-16-1097
105. Bir SC, Kolluru GK, McCarthy P, Shen X, Pardue S, Pattillo CB, et al. Hydrogen sulfide stimulates ischemic vascular remodeling through nitric oxide synthase and nitrite reduction activity regulating hypoxia-inducible factor-1alpha and vascular endothelial growth factor-dependent angiogenesis. J Am Heart Assoc (2012) 1:e004093. doi: 10.1161/JAHA.112.004093
106. Wang MJ, Cai WJ, Li N, Ding YJ, Chen Y, Zhu YC. The hydrogen sulfide donor NaHS promotes angiogenesis in a rat model of hind limb ischemia. Antioxid Redox Signal (2010) 12:1065–77. doi: 10.1089/ars.2009.2945
107. Cai WJ, Wang MJ, Moore PK, Jin HM, Yao T, Zhu YC. The novel proangiogenic effect of hydrogen sulfide is dependent on akt phosphorylation. Cardiovasc Res (2007) 76:29–40. doi: 10.1016/j.cardiores.2007.05.026
108. Hsieh MH, Tsai HW, Lin KJ, Wu ZY, Hu HY, Chang Y, et al. An in situ slow-releasing H2S donor depot with long-term therapeutic effects for treating ischemic diseases. Mater Sci Eng C Mater Biol Appl (2019) 104:109954. doi: 10.1016/j.msec.2019.109954
109. Hu Q, Wu D, Ma F, Yang S, Tan B, Xin H, et al. Novel angiogenic activity and molecular mechanisms of ZYZ-803, a slow-releasing hydrogen sulfide-nitric oxide hybrid molecule. Antioxid Redox Signal (2016) 25:498–514. doi: 10.1089/ars.2015.6607
110. Xiong Y, Chang LL, Tran B, Dai T, Zhong R, Mao YC, et al. ZYZ-803, a novel hydrogen sulfide-nitric oxide conjugated donor, promotes angiogenesis via cross-talk between STAT3 and CaMKII. Acta Pharmacol Sin (2020) 41:218–28. doi: 10.1038/s41401-019-0255-3
111. Lilyanna S, Peh MT, Liew OW, Wang P, Moore PK, Richards AM, et al. GYY4137 attenuates remodeling, preserves cardiac function and modulates the natriuretic peptide response to ischemia. J Mol Cell Cardiol (2015) 87:27–37. doi: 10.1016/j.yjmcc.2015.07.028
112. Polhemus D, Kondo K, Bhushan S, Bir SC, Kevil CG, Murohara T, et al. Hydrogen sulfide attenuates cardiac dysfunction after heart failure via induction of angiogenesis. Circ Heart Fail (2013) 6:1077–86. doi: 10.1161/CIRCHEARTFAILURE.113.000299
113. Qipshidze N, Metreveli N, Mishra PK, Lominadze D, Tyagi SC. Hydrogen sulfide mitigates cardiac remodeling during myocardial infarction via improvement of angiogenesis. Int J Biol Sci (2012) 8:430–41. doi: 10.7150/ijbs.3632
114. Givvimani S, Munjal C, Gargoum R, Sen U, Tyagi N, Vacek JC, et al. Hydrogen sulfide mitigates transition from compensatory hypertrophy to heart failure. J Appl Physiol (1985) (2011) 110:1093–100. doi: 10.1152/japplphysiol.01064.2010
115. Liang W, Chen J, Li L, Li M, Wei X, Tan B, et al. Conductive hydrogen sulfide-releasing hydrogel encapsulating ADSCs for myocardial infarction treatment. ACS Appl Mater Interfaces (2019) 11:14619–29. doi: 10.1021/acsami.9b01886
116. Kan J, Guo W, Huang C, Bao G, Zhu Y, Zhu YZ. S-propargyl-cysteine, a novel water-soluble modulator of endogenous hydrogen sulfide, promotes angiogenesis through activation of signal transducer and activator of transcription 3. Antioxid Redox Signal (2014) 20:2303–16. doi: 10.1089/ars.2013.5449
117. Cheng Y, Ndisang JF, Tang G, Cao K, Wang R. Hydrogen sulfide-induced relaxation of resistance mesenteric artery beds of rats. Am J Physiol Heart Circ Physiol (2004) 287:H2316–23. doi: 10.1152/ajpheart.00331.2004
Keywords: hydrogen sulfide, vascular, hypertension, angiogenesis, atherosclerosis
Citation: Zhu C, Liu Q, Li X, Wei R, Ge T, Zheng X, Li B, Liu K and Cui R (2022) Hydrogen sulfide: A new therapeutic target in vascular diseases. Front. Endocrinol. 13:934231. doi: 10.3389/fendo.2022.934231
Received: 02 May 2022; Accepted: 11 July 2022;
Published: 10 August 2022.
Edited by:
Chengqi Xu, Huazhong University of Science and Technology, ChinaReviewed by:
Rongfeng Zhang, Dalian Medical University, ChinaHuiying Liu, 307th Hospital of Chinese People’s Liberation Army, China
Copyright © 2022 Zhu, Liu, Li, Wei, Ge, Zheng, Li, Liu and Cui. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ranji Cui, cuiranji@jlu.edu.cn; Kexiang Liu, kxliu64@hotmail.com
†These authors have contributed equally to this work and share first authorship