 Vera Marie Kroesen1,2
Vera Marie Kroesen1,2 Paula Rodríguez-Martínez3
Paula Rodríguez-Martínez3 Eric García1
Eric García1 Yaiza Rosales1
Yaiza Rosales1 Jorge Díaz1
Jorge Díaz1 Montse Martín-Céspedes3
Montse Martín-Céspedes3 Gustavo Tapia3
Gustavo Tapia3 Maria Rosa Sarrias4,5
Maria Rosa Sarrias4,5 Pere-Joan Cardona1,6
Pere-Joan Cardona1,6 Cristina Vilaplana1,6*
Cristina Vilaplana1,6*
- 1Experimental Tuberculosis Unit (UTE), Fundació Institut Germans Trias i Pujol (IGTP), Universitat Autònoma de Barcelona (UAB), Badalona, Spain
- 2Carl-von-Ossietzky University Oldenburg, Oldenburg, Germany
- 3Pathology Department, Hospital Universitari Germans Trias i Pujol (HUGTIP), Universitat Autònoma de Barcelona (UAB), Badalona, Spain
- 4Innate Immunity Group, Fundació Institut Germans Trias i Pujol (IGTP), Badalona, Spain
- 5Centro de Investigación Biomédica en Red de Enfermedades Hepáticas y Digestivas (CIBEREhD), Madrid, Spain
- 6Centro de Investigación Biomédica en Red de Enfermedades Respiratorias (CIBERES), Madrid, Spain
An excessive, non-productive host-immune response is detrimental in active, chronic tuberculosis (TB) disease as it typically leads to tissue damage. Given their anti-inflammatory effect, non-steroidal anti-inflammatory drugs can potentially attenuate excessive inflammation in active TB disease. As such, we investigated the prophylactic and therapeutic effect of low-dose aspirin (LDA) (3 mg/kg/day), either alone or in combination with common anti-TB treatment or BCG vaccination, on disease outcome in an experimental murine model of active TB. Survival rate, bacillary load (BL) in lungs, and lung pathology were measured. The possible mechanism of action of LDA on the host’s immune response was also evaluated by measuring levels of CD5L/AIM, selected cytokines/chemokines and other inflammatory markers in serum and lung tissue. LDA increased survival, had anti-inflammatory effects, reduced lung pathology, and decreased bacillary load in late-stage TB disease. Moreover, in combination with common anti-TB treatment, LDA enhanced survival and reduced lung pathology. Results from the immunological studies suggest the anti-inflammatory action of LDA at both a local and a systemic level. Our results showed a systemic decrease in neutrophilic recruitment, decreased levels of acute-phase reaction cytokines (IL-6, IL-1β, and TNF-α) at late stage and a delay in the decrease in T cell response (in terms of IFN-γ, IL-2, and IL-10 serum levels) that occurs during the course of Mycobacterium tuberculosis infection. An anti-inflammatory milieu was detected in the lung, with less neutrophil recruitment and lower levels of tissue factor. In conclusion, LDA may be beneficial as an adjunct to standard anti-TB treatment in the later stage of active TB by reducing excess, non-productive inflammation, while enhancing Th1-cell responses for elimination of the bacilli.
Introduction
Tuberculosis (TB), which is a chronic infectious disease caused by Mycobacterium tuberculosis, ranks among the top 10 causes of death worldwide (1). In a clinical context, human TB presents with a wide spectrum of disease, ranging from asymptomatic infection to active TB disease (2). Patients can usually be divided into having latent TB infection or suffering from active TB disease, depending on whether they show clinical signs of the disease or not (2). Patients with co-morbidities that induce a pro-inflammatory milieu, such as diabetes mellitus and tobacco smoking, an impaired immune response, such as the elderly, children, people relying on immunosuppressive therapy, such as anti-TNFs (3) or with immunodeficiencies, such as those co-infected with HIV (1), are particularly at risk of developing active TB disease once infected with M. tuberculosis.
The current approach to treating TB entails antimicrobial drugs that target the mycobacteria (4). Thus, the standard 6-month treatment for TB patients consists of a four-drug regimen containing rifampicin, isoniazid, pyrazinamide, and ethambutol (4). Multi-drug resistant tuberculosis (MDR-TB), defined as resistance to at least two first-line drugs isoniazid and rifampicin, and extensively-drug resistant tuberculosis, where additional resistance to any fluoroquinolone or any of the injectable second-line aminoglycosides occurs, is an emerging challenge in the treatment of TB (1, 5). In 2015, the WHO reported an incidence of 450,000 new cases of MDR-TB worldwide, and it has been reported in virtually every country (1). One strategy that has been suggested to more effectively fight against TB, MDR-TB, and TB/HIV involves drugs that target host immune functions as adjuvants to classic antimicrobial treatment rather than focusing on the bacteria themselves (6). By not targeting the infecting microorganisms, these drugs have the potential advantage of not selecting TB drug resistance. The majority of these so-called host-directed therapies (HDTs) (7), which include non-steroidal anti-inflammatory drugs (NSAIDS) (8, 9), aim to balance host immune responses in order to decrease damage and are potentially able to achieve clinical improvement and decreased morbidity and mortality. Pathological immune reactions in the host, such as an insufficient or excessive inflammatory response, the latter of which leads to severe tissue damage, are considered to be a major cause of failure of current TB treatments (7).
Upon infection, M. tuberculosis is phagocytosed by alveolar macrophages, where it prevents phagosome–lysosome fusion and elimination by the lysosome (10). Infected antigen-presenting cells secrete various cytokines and chemokines to activate the innate immune response and influx of neutrophils (11, 12). Neutrophils usually represent a protective immune response during early infection by secreting oxidizing and hydrolytic agents that target the bacteria (11). However, although this neutrophil-dominated inflammation is beneficial in acute infections, where a fast and strong immune response can potentially lead to control or even clearance of the bacilli, it can be detrimental when non-productive and excessive, as described above, in the context of HIV co-infection (13), or simply in chronic infection (11). In active, chronic TB disease, an early and strong immune response has been reported to destroy delicate neighboring host tissues, thus leading to necrosis and resulting in cavitation, which facilitates spread of the bacilli, rather than containing M. tuberculosis replication and the continuous, excessively aggressive immune response (14). Attenuating the excessive host inflammatory response in active TB disease might thus be vital for treatment and disease outcome (11). Given their anti-inflammatory effect, NSAIDS can potentially attenuate neutrophil-mediated inflammation in TB (9, 8). Similarly, cyclooxygenase (COX) inhibitors have also been suggested to have potential therapeutic applications in other infections, such as in the control of parasite replication and dissemination in Chagas disease (15–17), in Leishmania major infection (18) and in pneumonia and pneumococcal-influenza co-infection (19).
In this study, we aimed to investigate the effect of low-dose aspirin (AAS) (LDA), administered alone or as an adjunct to common anti-TB treatment with the standard antibiotic treatment-regimen for human patients or with preventative BCG vaccination, in a murine model of active TB. Disease outcome was quantified in terms of survival rate, bacillary load (BL) in lungs, and lung pathology. LDA has previously been shown to prolong survival and enhance control of BL in the late stages of TB (12) and was, therefore, to confirm this finding. AAS is a salicylate that inhibits the two isoforms of COX in an irreversible manner, while leaving LOX activity unaffected (20, 21). The LOX pathway results in the synthesis of lipoxins (LX), which are known to be immunoresolvents with a potent anti-inflammatory effect and potential antimicrobial properties (22). The resulting synthesis of LX promotes the switching from a pro-inflammatory to anti-inflammatory milieu, with a decrease in pro-inflammatory cytokines and less neutrophil recruitment. Inhibition of the COX pathway also has an impact on vascularization and is widely used in the prevention of cardiovascular disease and stroke (21).
In light of the above, we decided to explore whether the described beneficial effects of LDA on survival rate, pathology, and BL in lungs in active TB are mediated by an anti-inflammatory effect, as previously found with ibuprofen (23). To assess the potential anti-inflammatory effect of LDA, we measured immunological profiles in serum and lungs in a murine model of active TB. Our specific objectives were: (1) to provide further evidence for the described beneficial effects of LDA on disease outcome in a murine model of active TB, when given prophylactically or therapeutically, by assessing survival rate, pulmonary BL, and lung pathology; (2) to investigate whether the potential beneficial effects are mediated by an anti-inflammatory mechanism of action; (3) to draw conclusions regarding the general immune responses in TB (as possible target for HDTs) by comparing immunological profiles in infected animals and non-infected negative control mice.
Materials and Methods
Experimental Design
A murine model of active TB based on a single intravenous (i.v.) infection [4 × 104–2 × 105/mL colony forming units (CFU)/mL per mouse] with M. tuberculosis H37Rv Pasteur strain in C3HeB/FeJ mice was used, as described previously (15). Animals (n = 164) were divided into five experiments (see Table 1):
(a) To evaluate the prophylactic effect of LDA: Experiments 1 and 2. AAS was given from 1 week before infection and disease outcome compared in aspirin-treated and non-treated animals.
(b) To evaluate the therapeutic effect of LDA: Experiments 3 to 5. In Experiment 3 (n = 24), disease outcome was compared in LDA-treated and non-treated animals. In Experiment 4, LDA was evaluated as a coadjuvant with BCG vaccination (n = 36). Mice were subcutaneously vaccinated with sham (SF) or BCG 106 CFU/animal [ImmunoCyst® BCG, Sanofi Pasteur (batch E-5-C4042A)] alone or in combination with therapeutical LDA (administered 2 weeks post-challenge). The results for the BCG- and BCG + AAS-treated groups were compared with those obtained for the sham group. The endpoint was established at week 18 after the start of the experiment as these BCG-vaccinated or BCG-vaccinated and AAS-treated mice did not die. Experiment 5 (n = 24) was designed to assess the therapeutic effect of LDA in combination with RIMSTAR, the standard antibiotic combination used in human TB treatment.
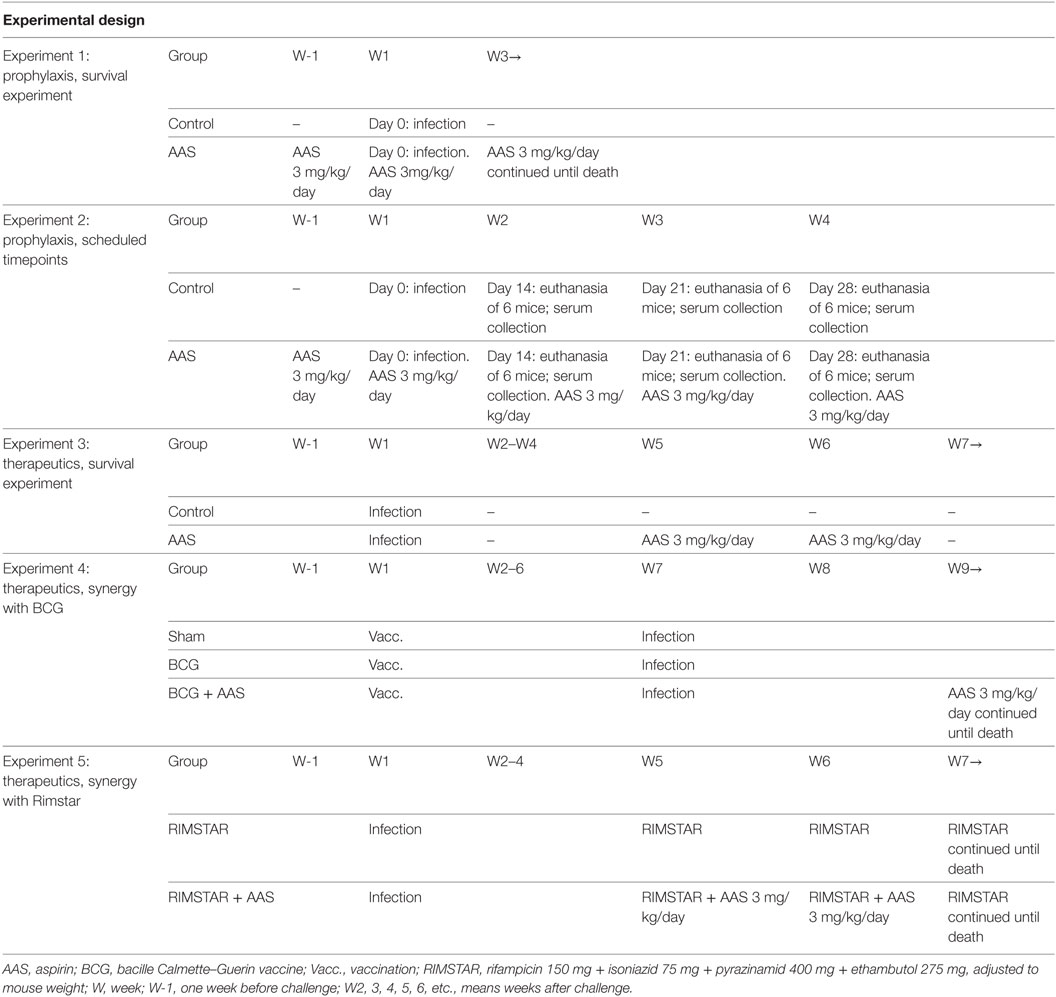
Table 1. Experimental design of the five experiments included in the study.
The Murine Model of Acute Active TB (C3HeB/FeJ Mice)
The C3HeB/FeJ murine model of acute active TB disease in humans was selected due to its suitability for investigating the effect of therapeutical strategies on disease outcome and immune mechanisms during active TB disease with excess inflammation and granuloma formation (12).
Endpoints to Assess the Impact of LDA on Disease Outcome
The effect of LDA on disease outcome was measured in terms of survival rate (all experiments except number 2; n = 108), BL in lungs at different timepoints (days 14, 21, and 28 post-infection; n = 48) and lung pathology analysis (histometry; n = 138) (Table 2). For the BL assessment, samples of lung lobes from animals from experiment 2 were collected, homogenized, and several dilutions plated on nutrient Middlebrook 7H11 agar (BD Diagnostics, Spark, USA). The number of CFU was counted after incubation for 28 days at 37°C and the results expressed as CFU/mL.
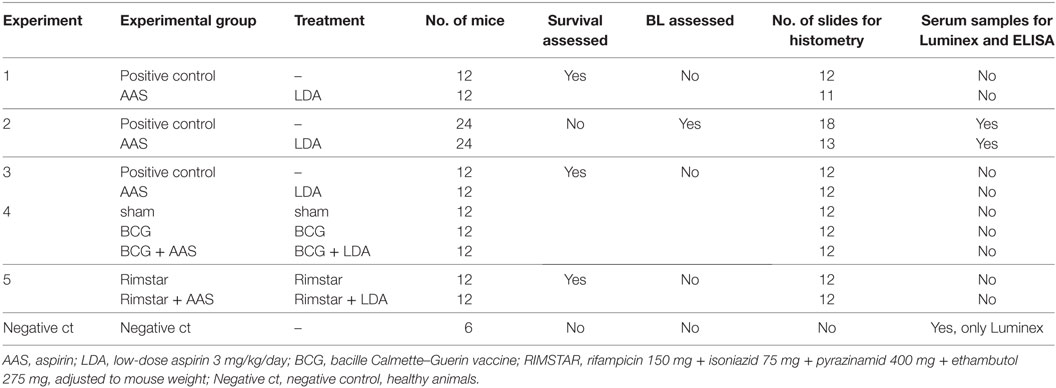
Table 2. Overview of the experiments and methods employed.
For the assessment of lung pathology, histological lung sections from C3HeB/FeJ mice from all experimental groups were analyzed by histometry. Lungs were fixed in 10% buffered formalin, embedded in paraffin and 5-µm sections stained with hematoxylin-eosin (HE) stain for histometric analysis. All slides were scanned at a resolution of 300 dpi (dots per inch), and scanned images analyzed using NIS-Elements Documentation 3.0 Software (by Nikon Instruments; Shinjuku, Tokio, Japan). The percentage of affected area in relation to total lung area was measured.
Methods to Assess the Impact of LDA on Immunological Profiles in Serum and Lung Tissue
The immunological profiles in sera from the experimental groups were assessed in Experiment 2 (Table 2). Ten selected cytokines and chemokines (G-CSF, KC, MIP-2, IL-1α, IL-1β, IL-6, TNFα, IL-2, IFNγ, and IL-17) were measured in serum samples obtained at days 14 (week 2), 21 (week 3), and 28 (week 4) post-infection, using the Luminex MILLIPLEX® MAP Kit, Mouse Cytokine/Chemokine Magnetic Bead Panel, 96-Well Plate Assay (EMD Millipore). Six non-infected mice were used as negative control in the Luminex assay. Circulating CD5L/AIM protein was assessed using the CircuLex Mouse AIM/CD5L/Spα ELISA Kit (MBL International Corporation, Woburn, USA) and the same sera samples. Both kits were used in accordance with their manufacturer’s instructions.
Immunohistochemical stains on paraffined sections were performed using the ultraView DAB (Ventana Medical Systems) in accordance with the manufacturer’s protocol. Monoclonal antibodies against TNF-α (rabbit polyclonal ab6671; Abcam, dilution 1:100), STAT1 (phospho S727) (rabbit monoclonal EPR3146; Abcam, dilution 1:500), myeloperoxidase (MPO) (rabbit polyclonal ab45977, Abcam, dilution 1:500), arginase 1 (rabbit polyclonal ab91279, Abcam, dilution 1:200), iNOS (rabbit polyclonal ab15323, Abcam, dilution 1:100), CD5L (dilution 1:50), tissue factor (TF) (rabbit monoclonal ab151748, Abcam, dilution 1:200) were used. Granulomas were imaged with a camera and NIS-Elements Documentation 3.0 Software connected to a microscope (Nikon Instruments; Shinjuku, Tokyo, Japan). As individual tissue sections from different animals with active TB contain multiple granulomas, we decided to image regions from five different granulomas with representative features, and to use them to analyze the IHC staining (5 per experimental group and timepoint). Images were acquired as TIFF-format images and analyzed with ImageJ (available at https://imagej.nih.gov/ij/). The median stained area for each antibody was graphically represented as square pixels.
Graphics and Statistics
Data are shown as median ± SEM. A non-parametric comparison (Mann–Whitney test) was used for comparisons between LDA and control groups (GraphPad Software v6.0). Statistically significant differences are designated as follows: *p < 0.05; **p < 0.01; ***p < 0.001.
Results
Survival Rate
Low-dose aspirin increased the survival rate when compared to controls in all experiments in which survival was assessed (experiments 1, 3, 4, and 5) (Figure 1). Both prophylactic and therapeutic treatment with LDA (experiments 1 and 3) increased survival compared to non-treated controls in a statistically significant manner (p < 0.001 and p = 0.0032, respectively). Both BCG vaccination alone and in combination with LDA (experiment 4) showed a statistically significant increase in survival compared to the control group (p < 0.001), although no additional effect of LDA was found, at least until termination of the experiment. Two weeks of LDA in addition to antibiotic treatment (experiment 5) increased survival compared to RIMSTAR-only treated animals, although the differences between the survival curves were not statistically significant, at least not with the number of animals included in each treatment group (n = 12).
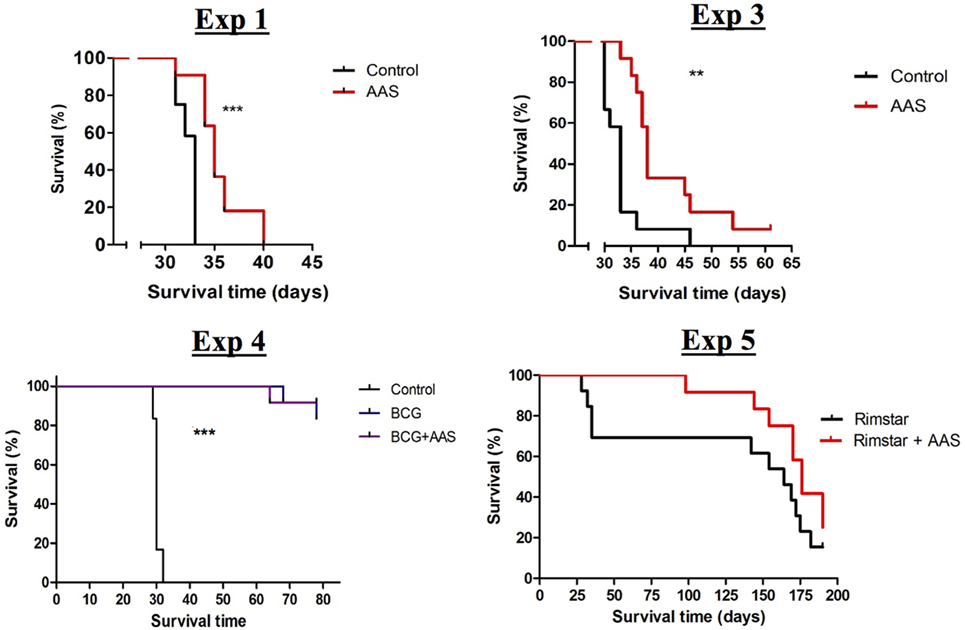
Figure 1. Survival assessment. Curves for the experimental groups achieved in the four experiments in which the impact of treatment on survival was assessed. Abbreviations: AAS, aspirin; BCG, bacille Calmette–Guerin vaccine; Vacc., vaccination; RIMSTAR, rifampicin 150 mg + isoniazid 75 mg + pyrazinamid 400 mg + ethambutol 275 mg, adjusted to mouse weight.
BL in Lungs
Prophylactic treatment with LDA (experiment 2) showed a statistically significant increase in BL on day 14 post-infection (p = 0.0022), but a statistically significant decrease at later stages (day 21, p = 0.0050; day 28, p = 0.0152) when compared to controls (Figure 2A).
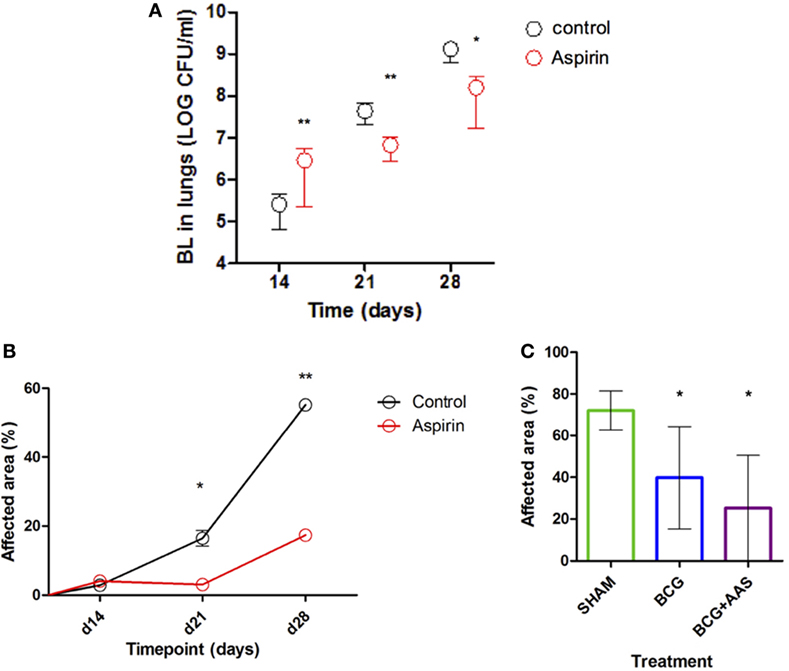
Figure 2. Impact of AAS on the BL and histometry. AAS, aspirin; BCG, bacille Calmette–Guerin vaccine; BL, bacillary load. Panel (A) shows the impact of AAS on the BL with time post-challenge. Panels (B,C) show the impact of AAS on the histometry in terms of affected lung area, both alone [(B): experiment 2] and in combination with BCG vaccine [(C): experiment 4].
Lung Pathology (Histometry)
Histological lung sections obtained from mice from all experiments were stained with HE-stain and analyzed in terms of granulomatous area in lungs (%). A statistically significant increase in lung pathology in both treatment groups over the course of infection was seen in those animals used to evaluate the BL at different timepoints (experiment 2). This increase was statistically significantly delayed and mitigated by preventive treatment with LDA only. As shown in Figure 2B, the % affected area was significantly reduced in LDA-treated mice compared to controls in late-stage infection (day 21, p = 0.0286; day 28, p = 0.0286).
BCG vaccination was found to prevent excess lung pathology compared to sham (p = 0.0169) (Figure 2C), keeping lung pathology very low at least until the end of the experiment at 4 months post-infection [mean of 39.78%; SD 24.5% (BCG group) and 72.07%; SD 9.335% (sham group)]. Adjunctive LDA treatment had an additional beneficial impact on this effect, although this was not statistically significant when compared to BCG alone.
Immunological Studies
Figure 3 shows the results of the Luminex assay in serum for the cytokines. All median serum concentrations (pg/mL, plus 25–75% confidence intervals) for all cytokines/chemokines and CD5L measured by Luminex and ELISA at weeks 2 (day 14), 3 (day 21), and 4 (day 28) post-challenge are shown in Table S1 in Supplementary Material. The IHC values in lungs are shown in Figure 4, and the results of CD5L measurements in serum (measured by ELISA, Figure 5A) and lungs (assessed by IHC, Figure 5B) are shown in Figure 5.
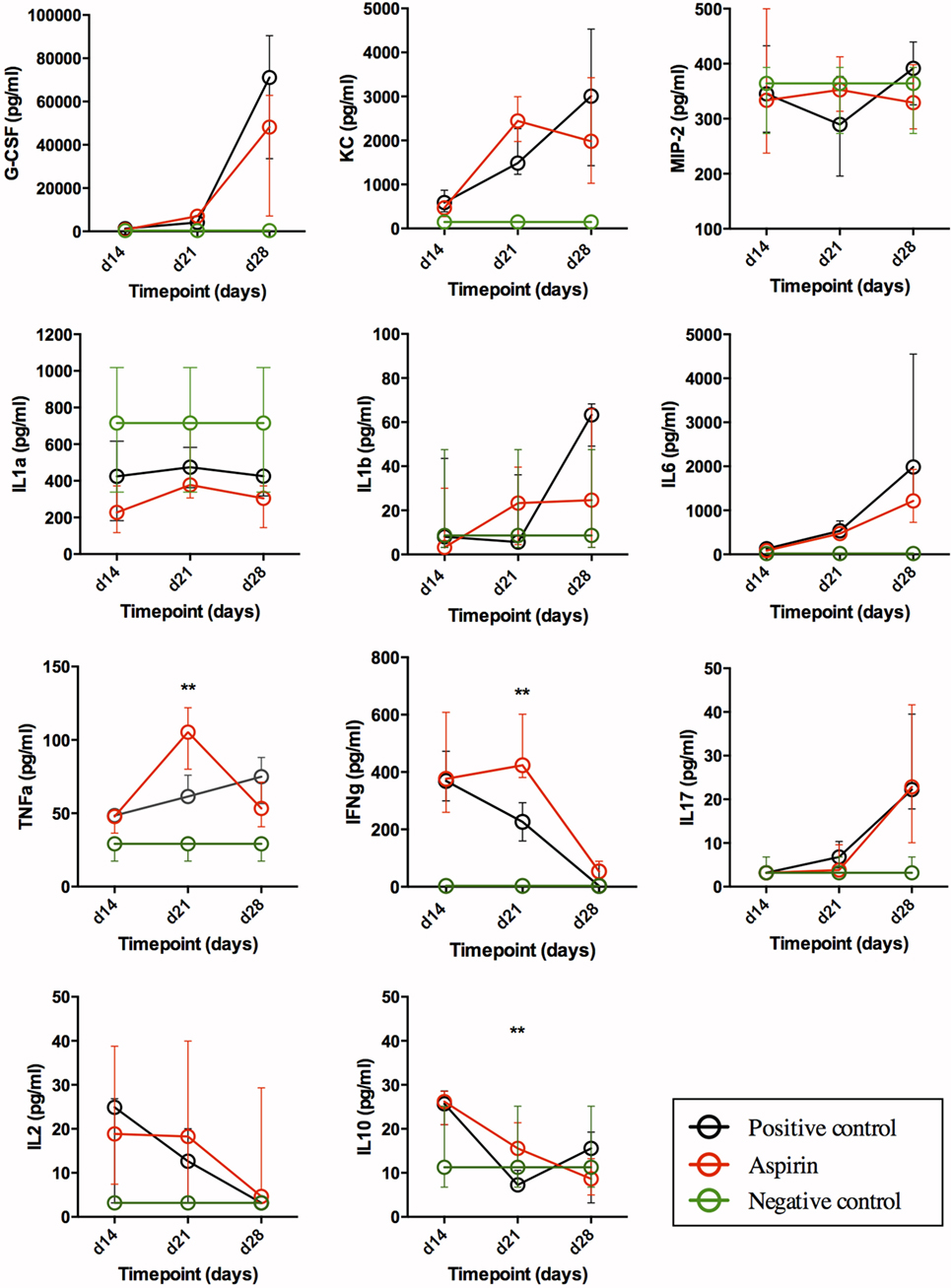
Figure 3. Impact of AAS on serum cytokine/chemokine levels. All data are presented in pg/mL. Abbreviations: AAS, aspirin; G-CSF, granulocyte-colony stimulating factor; KC, chemokine (C-X-C motif) ligand 1 (CXCL-1); MIP-2, macrophage inflammatory protein 2-alpha [also named chemokine (C-X-C motif) ligand 2 (CXCL2)]; IL, interleukin; TNFa, tumor necrosis α; IFNg, interferon γ.
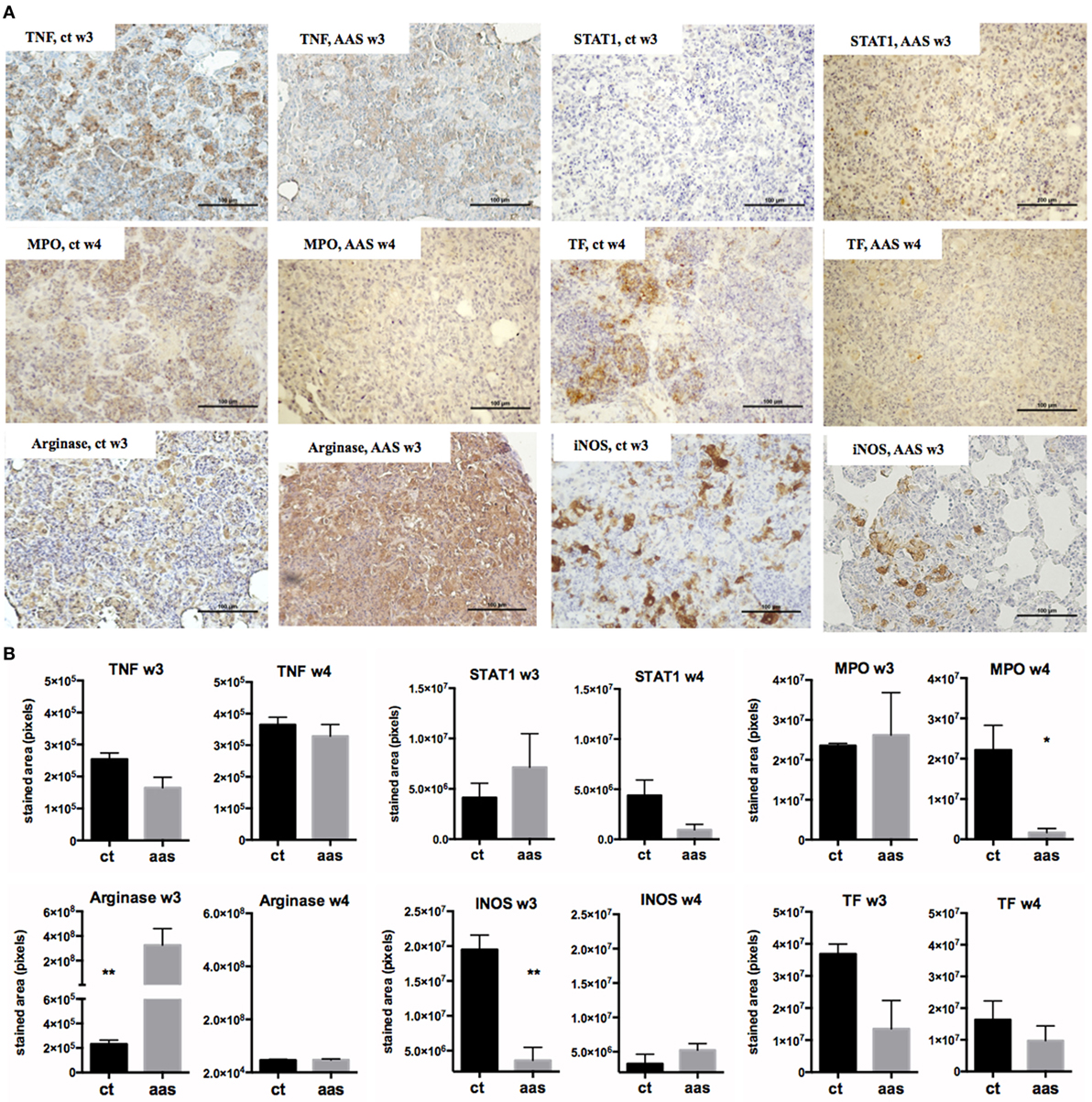
Figure 4. Impact of AAS in lung measured by IHQ. Panel (A) shows some pictures of the staining. The bar represents 100 µm. Panel (B) shows the results of the staining quantification by experimental group and timepoint. Abbreviations: AAS, aspirin; TNF, tumor necrosis α; STAT1, signal transducer and activator of transcription 1; MPO, myeloperoxidase; INOS, inducible nitric oxide synthase; TF, tissue factor.
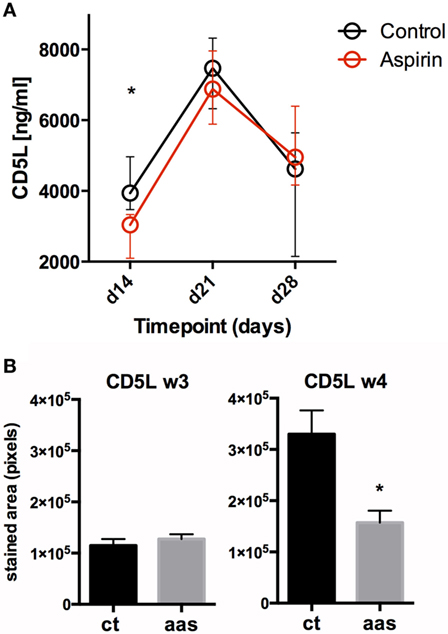
Figure 5. Impact of AAS on CD5L at a systemic and local level. Panel (A) shows serum levels (in pg/mL) for CD5L over time. Panel (B) shows CD5L in lung tissue on days 21 (week 3) and 28 (week 4) after challenge. Abbreviations: Ct, control; AAS, aspirin; d, day; w, week.
Neutrophil Recruitment (G-CSF, KC, and MIP-2)
Our findings in serum (Figure 3) underline a general increase in the recruitment of granulocytes during TB, as the G-CSF and KC (CXCL-1) values for infected groups differed from the negative control in a statistically significant manner. As regards G-CSF serum levels, there were no statistically significant differences between treatment groups, although LDA reduced serum levels in late-stage disease. For KC, serum levels seemed to reach a plateau in week 3 in LDA-treated mice, while in the untreated infected animals (positive control) they appeared to keep rising through week 4, although the difference was not statistically significant. MIP-2 (CXCL2) levels in infected groups remained similar to those in healthy mice, except for week 4 (day 28), when the levels in the positive control animals increased (difference not statistically significant).
Acute-Phase Reaction: IL-1α, IL-1β, IL-6, and TNFα
Our results (Figure 3) suggest an expected systemic (acute-phase) inflammatory reaction in TB. Thus, IL-1α values in infected groups were below the range in healthy animals (not statistically significant), and levels in the LDA group were lower than those obtained from the positive control group on days 14 and 28 post-infection. IL-1β levels in both infected groups increased over time, showing the greatest differences at week 4 (day 28) when compared to healthy animals. This rise seemed to be less pronounced in the LDA group, although no changes were found to be statistically significant. IL-6 and TNFα values in infected groups differed with respect to the negative control at all timepoints in a statistically significant manner. Indeed, there was a statistically significant increase in serum IL-6 levels over the course of infection, with this being mitigated in the AAS group in later stages. TNFα levels in the LDA group peaked at week 3 (day 21) post-infection in a statistically significant manner, but returned to day 14 levels by week 4 (day 28).
T-Cell Response: IL-2, IFNγ, and IL-17
Animals from both infected groups showed increased IFNγ values at weeks 2 and 3 (days 14 and 21) when compared to negative controls, with these levels decreasing to those of healthy animals by day 28 (week 4). AAS significantly delayed this decrease. Thus, on day 28 post-infection, the IL-17 values for infected groups were statistically significantly higher than those for the negative control. No differences were encountered between the control and AAS groups. Although serum IL-2 levels were generally low, they were elevated in the infected groups with respect to levels in healthy mice, dropping back over the course of infection. This effect was less evident in the AAS group (not statistically significant).
The Anti-Inflammatory Cytokine: IL-10
IL-10 levels in infected groups were higher at week 2 (day 14) post-challenge when compared to the negative control, subsequently decreasing to the level found in healthy animals by day 28. The drop in the AAS group in week 3 (day 21) was less pronounced (p = 0.01).
IHC Results
The IHC results (Figure 4) showed differences in terms of stained area in pixels when using anti-TNF and anti-STAT1 antibodies. These differences were observed at a tissue level both over time and when comparing the two experimental groups. The presence of TNF was slightly lower in the AAS group, especially at week 3 post-infection (difference not statistically significant). In contrast, STAT1 was found to be elevated at week 3 and decreased at week 4 in the AAS group with respect to the control group. MPO+ cells were statistically significantly decreased in the AAS group in late-stage disease (week 4). Differences were encountered for both arginase and INOS detection at week 3. Thus, while arginase staining increased, INOS staining decreased in the AAS group, with the differences being statistically significant in both cases. Tissue factor was found to be decreased in the AAS group at both weeks 3 and 4 post-infection, although this decrease was more important at week 3 and was not statistically significant at either timepoint.
CD5L/AIM
Significant changes in serum CD5L/AIM levels were observed in both experimental groups over the course of the infection, dominated by a pronounced peak in week 3 post-infection (day 21), followed by a decrease. Serum CD5L/AIM levels were lower for the LDA group at weeks 2 and 3, although the difference was only statistically significant in week 2 (p = 0.0159) (Figure 5A). The IHC study revealed less CD5L/AIM in lung sections from the AAS-treated group at week 4 (p = 0.031).
Discussion
The present study provides information on different LDA administration regimens (3 mg/kg) and their effect in a murine model of active TB.
The effect of AAS obtained in vivo has been classically considered to depend on the doses used. Thus, while low doses are commonly known to have an antithrombotic effect, only intermediate doses (500 mg to 3 g) were classically considered to be anti-inflammatory (24). However, the literature on preclinical and clinical studies in sepsis (25) shows that LDA triggers lipoxin synthesis, thus mediating anti-inflammatory and inflammation-resolving effects (26). Our results proved that maintained LDA also had an anti-inflammatory effect in the active TB model.
In our hands, when given preventatively or therapeutically in the absence of any other treatment, LDA statistically significantly increased survival in C3HeB/FeJ mice infected with TB, even enhancing the effect of RIMSTAR treatment. This is important as it stands in contrast with other findings, where AAS was described to antagonize the action of isoniazid (27) (which might raise the fear of the drug interfering with the standard treatment), and thus supporting the feasibility of AAS as an adjunct to TB drug therapy. Our findings, therefore, support the feasibility of AAS as an adjunct to TB drug therapy and provide evidence for the beneficial effects of LDA being mediated by an anti-inflammatory and an anti-mycobacterial mechanism, thereby reducing lung pathology over the course of the infection and BL in lungs, at least in late-stage TB.
As regards lung pathology, preventive treatment with LDA alone statistically significantly mitigated and delayed excess pulmonary damage in C3HeB/FeJ mice when compared to controls. Our results suggest that BCG vaccination can indeed prevent or delay terminal pathology due to M. tuberculosis infection in C3HeB/FeJ mice, and that treatment with LDA given therapeutically in addition to BCG vaccination does not provide any additional benefit, at least in the 4 months of this study. To determine whether LDA, administered therapeutically either alone or in combination with RIMSTAR or BCG vaccination, can statistically significantly mitigate and delay pulmonary damage during the course of infection, further experiments should be carried out using the same model, but with an experimental design considering several scheduled timepoints.
Analysis of serum levels for selected cytokines/chemokines showed that, as expected, TB itself generates a systemic inflammatory reaction with elevated levels of major neutrophil recruitment factors. Indeed, our results showed a significant increase in serum G-CSF, KC, IL-6, TNFα, and IL-17 levels when compared to negative controls. In contrast, the infection alone showed an inadequate drop in IFNγ, IL-2, and IL-10 levels over the course of infection. We observed a pronounced increase in serum IL-17 levels in late-stage disease, with this increase correlating with a decrease in IL-10 and CD5L. CD5L usually limits IL-17 production in Th17 and enhances IL-10 production in Th17 (28), and IL-10 has been suggested to play a role in controlling the anti-microbial activity and subsequent pulmonary tissue caseation (29). The observed late-stage rise in IL-17, therefore, probably reflects an increase in pathogenic Th17, thus leading to an increase in PMN infiltration and possibly explaining the excess late-stage lung pathology.
Overall, and although not reaching statistical significance for all measurements, our results suggest a systemic anti-inflammatory effect due to AAS treatment, as also described by Marzo et al. and Vilaplana et al. for ibuprofen in the same mouse model (12, 23). First, our findings point toward a systemic modulating effect of LDA on systemic neutrophil recruitment, thus resulting in a reduction in serum G-CSF and KC levels in late-stage disease. The IHC results confirmed the presence of fewer neutrophils in lungs, with a statistically significant decrease in MPO+ staining being observed at day 28 (week 4). With regards to the effects on neutrophil recruitment, it has been suggested that the anti-inflammatory mechanism of action of LDA is mitigated by both the reduced number (recruitment) of neutrophils and by reduced prostaglandin production in monocytes, lymphocytes, and neutrophils, which is implicated in the origin and development of granulomas (8, 14). These cells possess COX-2 activity and produce the prostaglandins (PG) implicated in the inflammation and lung damage associated with active TB, although in the short-term PGE2 might be beneficial (8). Moreover, the results for the AAS group showed less acute phase reaction cytokines (IL-6, IL-1β, and TNF-α) in the later stages. It has been observed that increased IL-6 levels directly correlate with increased BL (30) and X-ray severity in active TB in humans, therefore, this could be a valuable marker for predicting response to anti-TB treatment, with a pronounced reduction of serum levels being observed following treatment (31). Serum IL-1β levels in infected mice rose above the levels for healthy mice in late-stage disease. It has been described that increased IL-1β levels directly correlate with X-ray severity in active pulmonary TB in humans (31), and increased lung pathology was also observed in our study over the course of infection. Consequently, the lower levels achieved with LDA could be a reflection of the reduced lung pathology found in late-stage TB in the animals in this group.
Eisen et al. previously suggested that the beneficial anti-inflammatory effect of AAS might be mediated by a balancing effect on TNFα and that this could only be achieved at high dose, commonly known as the anti-inflammatory dose (32). Our results suggest that LDA does indeed balance TNF-α in TB in C3HeB/FeJ mice, significantly increasing serum levels in earlier stages (week 3) and reducing serum levels in the later stages of infection compared to non-treated positive controls. However, this increase could also be a consequence of the increase in BL at week 3 observed in the AAS group. Indeed, according to our IHQ results, TNF-α was slightly decreased in lung tissue (especially at week 3). However, the TNF-α increase observed at a serum level was not associated with worsened lung damage at that precise timepoint or subsequently according to the pathology results (33). The increase detected in serum levels at week 3 in the AAS group might enhance the Th1 response, the macrophage response and, consequently, clearance of the mycobacteria, acting synergistically with IL-2 and IFNγ. Increased levels of IFNγ and IL-2 have been found to be associated with moderate cases of human TB rather than more severe cases (34). Indeed, with LDA treatment we see a delay in the decrease in T cell response at a serum level (IFN-γ, IL-2, and IL-10) observed over the course of the infection, which reaches statistical significance at week 3. This delay at this precise timepoint might be responsible for controlling the tissue damage and BL by week 4, when infected animals are usually critically ill because of TB.
Signal transducer and activator of transcription 1 (STAT-1) was found to be elevated in lung tissue from LDA-treated animals compared to controls at week 3 and decreased at week 4, thus mirroring the results obtained in serum. The decrease in serum IL-10 levels over the course of the infection was also delayed in LDA-treated mice, and although the AAS group showed the lowest values in late-stage disease, the anti-inflammatory effect observed, as represented by higher IL-10 levels at week 3, could be considered a problem. It has been found that high serum IL-10 levels in patients with pulmonary TB lead to a slower response to treatment and a lack of bacterial control, thereby underlining the importance of a strong pro-inflammatory response when lung tissues are still intact (30, 35). In addition, reduced IL-10 (and IL-4) levels are associated with more moderate cases of human TB (34). In our study, the impact of the anti-inflammatory effect achieved by LDA seems to be beneficial rather than detrimental as, overall, AAS manages to decrease the inflammation in late-stage disease without impairing the T cell response in the short term. At a tissue level, we were able to demonstrate an increase in arginase-1 and a decrease in INOS at a lung level by week 3 in animals treated with LDA compared to controls. Arginase and INOS have been reported to be markers for characterizing and differentiating different macrophage functions as pro-healing/anti-inflammatory (Arginase) or pro-inflammatory (INOS) in the context of TB (36). As such, LDA treatment may be temporarily switching the phenotype of lung macrophages toward a more pro-healing/anti-inflammatory mode at week 3, which could have an important impact on disease outcome.
As regards CD5L measurements, mice from the AAS group had lower values than those from the control group at early timepoints (weeks 2 and 3) in serum and at later stages (week 4) at a lung level. CD5L/AIM is an innate protein that has been previously described to peak in plasma at week 3 post-infection, which may reflect a strong early innate immune response aimed at controlling M. tuberculosis growth. This enhances autophagy for elimination of the bacilli and acts in an anti-inflammatory (regulatory) manner on macrophages and Th1 (28, 37, 38). According to our results, the observations at a serum level may not be mirrored at the lung level as we see increased CD5L in lungs by IHC in week 4 (less pronounced in AAS-treated animals). This could suggest that this protein may play a role both during early infection and also in late stages of the disease, a point that should be studied in further experiments.
Finally, the measurement of TF by IHQ showed high levels at week 3, which was diminished by AAS treatment. TF is the principal initiator of the coagulation cascade, which plays a role in inflammation (39) and innate immunity and is expressed in response to different stimuli, such as infections and inflammation (40). The role of TF in TB is controversial. Thus, while some authors have stated that although this factor is increased, TF-mediated coagulation does not contribute to the host’s defense (41), others have suggested a protective role for TF during M. tuberculosis infection by demonstrating, in a mouse model of TB, that TF deficiency is associated with increased M. tuberculosis replication in lungs (42). LXA4 was found to be able to promote a proinflammatory and prothrombotic profile in vitro by inducing TF expression (43). However, a tendency toward hypercoagulation is a known problem in TB patients, especially in severe cases, therefore, the modulation of this state has been proposed as a target for AAS treatment in TB meningitis (9, 44). Indeed, our results point in this direction.
Our results are in contrast to some previous in vitro and in vivo studies in the literature, which suggest that inhibition of prostaglandin pathways involving a decrease in PGE2 and production of LXA4 is detrimental in terms of immunity to TB and might impair T cell immunity (45–47). In our opinion, the coexistence of contradictory results in the literature in this sense is probably due to the important matter of timing in HDT administration during TB, as a strong inflammatory response is needed at the beginning of the infection for possible control or even clearance of the mycobacteria. Our study was performed using a murine model based on intravenous infection of C3HeB/FeJ mice, which consistently develop lesions with liquefactive necrosis after infection with M. tuberculosis and die after 30 days in the absence of treatment. The characteristics of these lesions well mimic active TB in humans at diagnosis (when the disease is exacerbated, with an important inflammatory component and subsequent host tissue destruction), but not human latent infection or disease without a predominant inflammatory response. For this reason, we consider that results from studies in C3HeB/FeJ mice are not applicable to immune-competent humans chronically infected with M. tuberculosis or suffering from mild forms of TB. In light of the above, and as a conclusion to our work, we propose that LDA should only be considered as an adjunct to antibiotic treatment and/or vaccination in patients who present with clinical signs of disease (wasting, cough, etc.). On the other hand, the finding that preventative treatment with LDA increases survival, delays excess granuloma formation in lungs, and reduces BL in late-stage TB disease in our model suggests LDA could be given to patients at high risk for TB as a preventive measure, although further studies should be performed to confirm this point.
Ethics Statement
All procedures were performed according to protocol DMAH6119, which was reviewed by the Animal Experimentation Ethics Committee of the Hospital Universitari Germans Trias i Pujol (registered as B9900005) and approved by the Dept d’Agricultura, Ramaderia, Pesca, Alimentació i Medi Natural of the Catalan Government, according to current national and European Union legislation regarding the protection of Experimental animals. Mice were supervised daily following a strict monitoring protocol in order to ensure animal welfare, and euthanized, if required, with isoflurane (inhalation excess).
Author Contributions
CV headed the project. CV, MS, and P-JC conceived and planned the experiments. VK, PR-M, MM-C, GT, EG, and JD carried out the experiments. VK, GT, MS, P-JC, and CV contributed to the interpretation of the results. VK and CV took the lead in writing the manuscript. All authors provided critical feedback and helped to shape the research, analysis, and manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
This work was supported by the Plan Nacional I + D + I co-financed by ISCIII-Subdirección General de Evaluación and Fondo-EU de Desarrollo Regional (FEDER) through CV contract CP13/00174, MRS contract CPII14/00021, and project MS13/00174. VK received a PROMOS grant from the German Academic Exchange Service (DAAD) and a Fernweh grant from the International Student Office of the University of Oldenburg. The study and evaluation of HDTs against tuberculosis is part of the EDCTP2 program supported by the European Union. The Experimental Tuberculosis Unit is accredited by the Catalan Agency for Management of University and Research Grants with code 2017 SGR500 and the IGTP is a member of the CERCA network of institutes. The funders had no role in study design, data collection, and analysis, decision to publish, or preparation of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at https://www.frontiersin.org/articles/10.3389/fimmu.2018.00798/full#supplementary-material.
References
1. World Health Organization (WHO). Global TB Report 2017 (2017). Available from: http://www.who.int/tb/publications/global_report/en/ (Accessed: April 8, 2018).
2. Pai M, Behr MA, Dowdy D, Dheda K, Divangahi M, Boehme CC, et al. Tuberculosis. Nat Rev Dis (2016) 2:16076. doi:10.1038/nrdp.2016.76
3. Bourigault M-L, Segueni N, Rose S, Court N, Vacher R, Vasseur V, et al. Relative contribution of IL-1α, IL-1β and TNF to the host response to Mycobacterium tuberculosis and attenuated M. bovis BCG. Immun Inflamm Dis (2013) 1(1):47–62. doi:10.1002/iid3.9
4. World Health Organization (WHO). Treatment of Tuberculosis: Guidelines. 4th ed. Geneva: World Health Organization (2010). 160 p. Available from: https://www.ncbi.nlm.nih.gov/books/NBK138741/
5. World Health Organization (WHO). Treatment Guidelines for Drug-Resistant Tuberculosis. Geneva: World Health Organization (2016). 61 p. Available from: http://www.who.int/tb/areas-of-work/drug-resistant-tb/MDRTBguidelines2016.pdf%0Ahttp://www.who.int/kms/handbook_2nd_ed.pdf (Accessed: April 8, 2018).
6. Zumla A, Maeurer M, Chakaya J, Hoelscher M, Ntoumi F, Rustomjee R, et al. Host-directed therapies for tackling multi-drug resistant TB – learning from the Pasteur-Bechamp debates, and the Host-Directed Therapies Network (HDT-NET). Clin Infect Dis (2015) 61(9):1432–8. doi:10.1093/cid/civ631
7. Zumla A, Maeurer M, Chakaya J, Hoelscher M, Ntoumi F, Rustomjee R, et al. Towards host-directed therapies for tuberculosis. Nat Rev Drug Discov (2015) 14:511–2. doi:10.1038/nrd4696
8. Ivanyi J, Zumla A. Nonsteroidal antiinflammatory drugs for adjunctive tuberculosis treatment. J Infect Dis (2013) 208(2):185–8. doi:10.1093/infdis/jit153
9. Kroesen VM, Gröschel MI, Martinson N, Zumla A, Maeurer M, van der Werf TS, et al. Non-steroidal anti-inflammatory drugs as host-directed therapy for tuberculosis: a systematic review. Front Immunol (2017) 8:772. doi:10.3389/fimmu.2017.00772
10. Kimmey JM, Huynh JP, Weiss LA, Park S, Kambal A, Debnath J, et al. Unique role for ATG5 in neutrophil-mediated immunopathology during M. tuberculosis infection. Nature (2015) 528(7583):565–9. doi:10.1038/nature16451
11. Hawn TR, Matheson AI, Maley SN, Vandal O. Host-directed therapeutics for tuberculosis: can we harness the host? Microbiol Mol Biol Rev (2013) 77(4): 608–627. doi:10.1128/MMBR.00032-13
12. Marzo E, Vilaplana C, Tapia G, Diaz J, Garcia V, Cardona P-J. Damaging role of neutrophilic infiltration in a mouse model of progressive tuberculosis. Tuberculosis (2014) 94(1):55–64. doi:10.1016/j.tube.2013.09.004
13. Nusbaum RJ, Calderon VE, Huante MB, Sutjita P, Vijayakumar S, Lancaster KL, et al. Pulmonary tuberculosis in humanized mice infected with HIV-1. Sci Rep (2016) 6:21522. doi:10.1038/srep21522
14. Kiran D, Podell BK, Chambers M, Basaraba RJ. Host-directed therapy targeting the Mycobacterium tuberculosis granuloma: a review. Semin Immunopathol (2016) 38(2):167–83. doi:10.1007/s00281-015-0537-x
15. Freire-De-Lima CG, Nascimento DO, Soares MBP, Bozza PT, Castro-Faria-Neto HC, De Mello FG, et al. Uptake of apoptotic cells drives the growth of a pathogenic trypanosome in macrophages. Nature (2000) 403(6766):199–203. doi:10.1038/35003208
16. Lonien SCH, Malvezi AD, Suzukawa HT, Yamauchi LM, Yamada-Ogatta SF, Rizzo LV, et al. Response to Trypanosoma cruzi by human blood cells enriched with dentritic cells is controlled by cyclooxygenase-2 pathway. Front Microbiol (2017) 8:2020. doi:10.3389/fmicb.2017.02020
17. Luna-Gomes T, Filardy AA, Rocha JDB, Decote-Ricardo D, LaRocque-de-Freitas IF, Morrot A, et al. Neutrophils increase or reduce parasite burden in Trypanosoma cruzi-infected macrophages, depending on host strain: role of neutrophil elastase. PLoS One (2014) 9(3):e90582. doi:10.1371/journal.pone.0090582
18. Arcanjo AF, LaRocque-de-Freitas IF, Rocha JDB, Zamith D, Costa-da-Silva AC, Nunes MP, et al. The PGE2/IL-10 axis determines susceptibility of B-1 cell-derived phagocytes (B-1CDP) to Leishmania major infection. PLoS One (2015) 10(5):e0124888. doi:10.1371/journal.pone.0124888
19. Wang H, Anthony D, Yatmaz S, Wijburg O, Satzke C, Levy B, et al. Aspirin-triggered resolvin D1 reduces pneumococcal lung infection and inflammation in a viral and bacterial coinfection pneumonia model. Clin Sci (2017) 131(18):2347–62. doi:10.1042/CS20171006
20. Aspirin Monograph for Professionals. Available from: https://www.Drugs.com
21. Dennis EA, Norris PC. Eicosanoid storm in infection and inflammation. Nat Rev Immunol (2015) 15(8):511–23. doi:10.1038/nri3859
22. Romano M, Cianci E, Simiele F, Recchiuti A. Lipoxins and aspirin-triggered lipoxins in resolution of inflammation. Eur J Pharmacol (2015;) 760:49–63. doi:10.1016/j.ejphar.2015.03.083
23. Vilaplana C, Marzo E, Tapia G, Diaz J, Garcia V, Cardona P-J. Ibuprofen therapy resulted in significantly decreased tissue bacillary loads and increased survival in a new murine experimental model of active tuberculosis. J Infect Dis (2013) 208(2):199–202. doi:10.1093/infdis/jit152
24. Hankey GJ, Eikelboom JW. Antithrombotic drugs for patients with ischaemic stroke and transient ischaemic attack to prevent recurrent major vascular events. Lancet Neurol (2010) 9:273–84. doi:10.1016/S1474-4422(10)70038-7
25. Sossdorf M, Otto GP, Boettel J, Winning J, Lösche W. Benefit of low-dose aspirin and non-steroidal anti-inflammatory drugs in septic patients. Crit Care (2012) 17(1):1–2. doi:10.1186/cc11886
26. Eisen DP. Manifold beneficial effects of acetyl salicylic acid and nonsteroidal anti-inflammatory drugs on sepsis. Intensive Care Med (2012) 38:1249–57. doi:10.1007/s00134-012-2570-8
27. Byrne ST, Denkin SM, Zhang Y. Aspirin antagonism in isoniazid treatment of tuberculosis in mice. Antimicrob Agents Chemother (2007) 51(2):794–5. doi:10.1128/AAC.01145-06
28. Wang C, Yosef N, Gaublomme J, Wu C, Lee Y, Clish CB, et al. CD5L/AIM regulates lipid biosynthesis and restrains Th17 cell pathogenicity. Cell (2015) 163(6):1413–27. doi:10.1016/j.cell.2015.10.068
29. Marakalala MJ, Raju RM, Sharma K, Zhang YJ, Eugenin EA, Prideaux B, et al. Inflammatory signaling in human tuberculosis granulomas is spatially organized. Nat Med (2016) 22(5):531–8. doi:10.1038/nm.4073
30. Mesquita EDD, Gil-Santana L, Ramalho D, Tonomura E, Silva EC, Oliveira MM, et al. Associations between systemic inflammation, mycobacterial loads in sputum and radiological improvement after treatment initiation in pulmonary TB patients from Brazil: a prospective cohort study. BMC Infect Dis (2016) 16:368. doi:10.1186/s12879-016-1736-3
31. Chowdhury IH, Ahmed AM, Choudhuri S, Sen A, Hazra A, Pal NK, et al. Alteration of serum inflammatory cytokines in active pulmonary tuberculosis following anti-tuberculosis drug therapy. Mol Immunol (2014) 62(1):159–68. doi:10.1016/j.molimm.2014.06.002
32. Eisen DP, McBryde ES, Walduck A. Low-dose aspirin and ibuprofen’s sterilizing effects on Mycobacterium tuberculosis suggest safe newadjuvant therapies for tuberculosis. J Infect Dis (2013) 208(11):1925–7. doi:10.1093/infdis/jit476
33. Kaufmann SHE, Dorhoi A, Hotchkiss RS, Bartenschlager R. Host-directed therapies for bacterial and viral infections. Nat Rev Drug Discov (2018) 17:35–56. doi:10.1038/nrd.2017.162
34. Dlugovitzky D, Torres-Morales A, Rateni L, Farroni MA, Largacha C, Molteni O, et al. Circulating profile of Th1 and Th2 cytokines in tuberculosis patients with different degrees of pulmonary involvement. FEMS Immunol Med Microbiol (1997) 18(3):203–7. doi:10.1111/j.1574-695X.1997.tb01046.x
35. Djoba Siawaya JF, Beyers N, Van Helden P, Walzl G. Differential cytokine secretion and early treatment response in patients with pulmonary tuberculosis. Clin Exp Immunol (2009) 156(1):69–77. doi:10.1111/j.1365-2249.2009.03875.x
36. Mattila JT, Ojo OO, Kepka-Lenhart D, Marino S, Kim JH, Eum SY, et al. Microenvironments in tuberculous granulomas are delineated by distinct populations of macrophage subsets and expression of nitric oxide synthase and arginase isoforms. J Immunol (2013) 191(2):773–84. doi:10.4049/jimmunol.1300113
37. Sanjurjo L, Amézaga N, Vilaplana C, Cáceres N, Marzo E, Valeri M, et al. The scavenger protein apoptosis inhibitor of macrophages (AIM) potentiates the antimicrobial response against Mycobacterium tuberculosis by enhancing autophagy. PLoS One (2013) 8(11):e79670. doi:10.1371/journal.pone.0079670
38. Sanjurjo L, Aran G, Roher N, Valledor AF, Sarrias M-R. AIM/CD5L: a key protein in the control of immune homeostasis and inflammatory disease. J Leukoc Biol (2015) 98(2):173–84. doi:10.1189/jlb.3RU0215-074R
39. Schuliga M. The inflammatory actions of coagulant and fibrinolytic proteases in disease. Mediators Inflamm (2015) 2015:437695. doi:10.1155/2015/437695
40. Caccamo N, Dieli F. Inflammation and the coagulation system in tuberculosis: tissue factor leads the dance. Eur J Immunol (2016) 46:303–6. doi:10.1002/eji.201546225
41. Kothari H, Keshava S, Vatsyayan R, Mackman N, Rao LVM, Pendurthi UR. Role of tissue factor in Mycobacterium tuberculosis-induced inflammation and disease pathogenesis. PLoS One (2014) 9(12):e114141. doi:10.1371/journal.pone.0114141
42. Venkatasubramanian S, Tripathi D, Tucker T, Paidipally P, Cheekatla S, Welch E, et al. Tissue factor expression by myeloid cells contributes to protective immune response against Mycobacterium tuberculosis infection. Eur J Immunol (2016) 46(2):464–79. doi:10.1002/eji.201545817
43. Maderna P, Godson C, Hannify G, Murphy M, Brady HR. Influence of lipoxin A4 and other lipoxygenase-derived eicosanoids on tissue factor expression. Am J Physiol Cell Physiol (2000) 279(4):C945–53. doi:10.1152/ajpcell.2000.279.4.C945
44. Schoeman J, Mansvelt E, Springer P, Van Rensburg AJ, Carlini S, Fourie E. Coagulant and fibrinolytic status in tuberculous meningitis. Pediatr Infect Dis J (2007) 26(5):428–31. doi:10.1097/01.inf.0000261126.60283.cf
45. Mayer-Barber KD, Andrade BB, Oland SD, Amaral EP, Barber DL, Gonzales J, et al. Host-directed therapy of tuberculosis based on interleukin-1 and type I interferon crosstalk. Nature (2014) 511(7507):99–103. doi:10.1038/nature13489
46. Divangahi M, Desjardins D, Nunes-Alves C, Remold HG, Behar SM. Eicosanoid pathways regulate adaptive immunity to Mycobacterium tuberculosis. Nat Immunol (2010) 11(8):751–8. doi:10.1038/ni.1904
Keywords: aspirin, host-directed therapies, non-steroidal anti-inflammatory drugs, tuberculosis, Mycobacterium tuberculosis, mouse model
Citation: Kroesen VM, Rodríguez-Martínez P, García E, Rosales Y, Díaz J, Martín-Céspedes M, Tapia G, Sarrias MR, Cardona P-J and Vilaplana C (2018) A Beneficial Effect of Low-Dose Aspirin in a Murine Model of Active Tuberculosis. Front. Immunol. 9:798. doi: 10.3389/fimmu.2018.00798
Received: 07 February 2018; Accepted: 03 April 2018;
Published: 23 April 2018
Edited by:
Jesús Gonzalo-Asensio, University of Zaragoza, SpainReviewed by:
Debora Decote-Ricardo, Universidade Federal Rural do Rio de Janeiro, BrazilNina Ivanovska, Institute of Microbiology (BAS), Bulgaria
Copyright: © 2018 Kroesen, Rodríguez-Martínez, García, Rosales, Díaz, Martín-Céspedes, Tapia, Sarrias, Cardona and Vilaplana. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cristina Vilaplana, cvilaplana@gmail.com, cvilaplana@igtp.cat