 Teun J. de Vries1*
Teun J. de Vries1* Ismail el Bakkali1
Ismail el Bakkali1 Thomas Kamradt2
Thomas Kamradt2 Georg Schett3
Georg Schett3 Ineke D. C. Jansen1
Ineke D. C. Jansen1 Patrizia D'Amelio4
Patrizia D'Amelio4- 1Department of Periodontology, Academic Centre for Dentistry Amsterdam (ACTA), University of Amsterdam and Vrije Universiteit Amsterdam, Amsterdam, Netherlands
- 2Institute of Immunology, Universitätsklinikum Jena, Jena, Germany
- 3Department of Internal Medicine III, Friedrich-Alexander University Erlangen-Nürnberg and Universitatsklinikum Erlangen, Erlangen, Germany
- 4Gerontology and Bone Metabolic Diseases Division, Department of Medical Science, University of Turin, Turin, Italy
Local priming of osteoclast precursors (OCp) has long been considered the main and obvious pathway that takes place in the human body, where local bone lining cells and RANKL-expressing osteocytes may facilitate the differentiation of OCp. However, priming of OCp away from bone, such as in inflammatory tissues, as revealed in peripheral blood, may represent a second pathway, particularly relevant in individuals who suffer from systemic bone loss such as prevalent in inflammatory diseases. In this review, we used a systematic approach to review the literature on osteoclast formation in peripheral blood in patients with inflammatory diseases associated with bone loss. Only studies that compared inflammatory (bone) disease with healthy controls in the same study were included. Using this core collection, it becomes clear that experimental osteoclastogenesis using peripheral blood from patients with bone loss diseases in prevalent diseases such as rheumatoid arthritis, osteoporosis, periodontitis, and cancer-related osteopenia unequivocally point toward an intrinsically increased osteoclast formation and activation. In particular, such increased osteoclastogenesis already takes place without the addition of the classical osteoclastogenesis cytokines M-CSF and RANKL in vitro. We show that T-cells and monocytes as OCp are the minimal demands for such unstimulated osteoclast formation. In search for common and disease-specific denominators of the diseases with inflammation-driven bone loss, we demonstrate that altered T-cell activity and a different composition—such as the CD14+CD16+ vs. CD14+CD16– monocytes—and priming of OCp with increased M-CSF, RANKL, and TNF- α levels in peripheral blood play a role in increased osteoclast formation and activity. Future research will likely uncover the barcodes of the OCp in the various inflammatory diseases associated with bone loss.
Introduction
Close inspection of skeletons as seen in anatomy museums may show signs of inflammatory bone loss, present as joint erosions and bone degradation of the tooth sockets that surround teeth. This betrays an evoked activity of bone degradation by inflammation steered osteoclasts, so different from turn-over osteoclasts that in fact leave the rest of the skeleton seemingly normal. The different bone cells, osteoclasts, bone-lining cells, osteoblasts and osteocytes are responsible for a lifelong balanced remodeling process. When this process becomes unbalanced, such as during inflammatory diseases with bone-loss, it may result in severe bone loss, locally, or systemically. The key cell-type in this disturbed bone balance is the osteoclast, the multinucleated cell responsible for breaking down bone tissue (1).
Osteoclasts derive from cells of the monocyte/macrophage lineage (2–4) present in bone marrow (5), but also present in peripheral blood (6). Osteoclasts play a key role in diseases that are associated with increased bone loss (7). Such diseases include common diseases such as the rheumatic diseases, osteoporosis, periodontitis, cancers that metastasize to bone and Crohn's disease. Less common diseases that also give rise to bone loss are chronic liver disease, Gaucher's disease, Turner's syndrome, and phenylketonuria. Bone loss is also frequently observed in patients with chronic kidney disease. In nearly all of these diseases, excessive osteoclast generation and activation, with a key contribution to the altered immune system plays a dominant role. All these diseases are discussed in detail below.
Growth factors and cytokines that can be produced by a wide range of cells in the human body regulate the activity and formation of osteoclasts. The principle differentiation factors for osteoclast differentiation are macrophage colony-stimulating factor (M-CSF), receptor activator of nuclear factor kappa-B ligand (RANKL) (8) and the inhibitor of osteoclast differentiation, the “protector of bone,” osteoprotegerin (OPG) (9). These cytokines are produced by osteocytes and bone lining cells/osteoblasts (10). However, RANKL can also be produced by T-cells (11, 12), by synovial fibroblasts from inflamed joints (13) and by tooth-associated fibroblasts (14). Apart from this classical pathway, it was demonstrated by Kim et al. using RANK-/- mouse models, that osteoclasts may also form through stimulation with inflammatory cytokines such as tumor necrosis factor-α (TNF-α) (15).
Apart from the common culture methods of osteoclasts using M-CSF and RANKL, there are strong indications that osteoclast precursors may differentiate into multinucleated osteoclasts in the absence of added osteoclastogenesis stimulating factors M-CSF and RANKL. This is referred to as “spontaneous” or unstimulated osteoclast formation (16, 17), recently excellently reviewed by Salamanna et al. (7). A better term for this could be self-stimulatory osteoclastogenesis, where the combination of T-cells that may provide the osteoclastogenesis signals with primed OCp may give rise to osteoclast formation. Here, and experimental evidence is provided below, cells from peripheral blood, such as T-cells, may provide the necessary differentiation factors for the monocytic, CD14+ osteoclast precursor cells in blood. These studies are based on experimental in vitro studies which suggest an activation of the OCp by inflammatory mediators present in the plasma of patients with inflammatory bone disease or an intrinsic change of cells toward more osteoclastic differentiation. Examples include periodontitis, osteoporosis, Crohn's disease, rheumatoid arthritis, and bone metastatic cancer and will be further described later on in this review.
The aim of this systematic literature review is to provide an overview and an interpretation of experimental in vitro studies involving osteoclast formation from peripheral blood of patients with inflammatory diseases that lead to bone loss compared to the osteoclast formation from peripheral blood of healthy controls. This will gain insight into the various mechanisms that play a role in the activation of osteoclasts from peripheral blood in these inflammatory bone diseases. The similarities and differences in peripheral blood-mediated osteoclast formation between the various inflammatory diseases associated with bone loss will be examined and discussed.
Literature Search
The methodological approach of systematic review was used. The PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) was used to reduce the bias in the selection of publications for this review.
Search strategy
Search Query
#1 Peripheral blood
#2 Osteoclast OR osteoclasts
#3 Osteoclast formation OR osteoclast differentiation OR osteoclastogenesis
#4 (Periodontitis OR periodontal disease) OR rheumatoid arthritis OR psoriatic arthritis OR inflammatory bowel disease OR Crohn's disease OR osteoporosis OR bone metastatic cancer
#5 (#1 AND #2 AND #3 AND #4) AND (“2002/01/01”[PDat]: “2018/07/31”[PDat])
The electronic search for relevant studies was carried out in the three databases Pubmed, Embase, and Web of science. The keywords used for the search were “peripheral blood,” “osteoclast,” “osteoclasts,” “osteoclast formation,” “osteoclast differentiation,” “osteoclastogenesis,” “periodontitis,” “periodontal disease,” “rheumatoid arthritis,” “psoriatic arthritis,” “inflammatory bowel disease,” “Crohn's disease,” “osteoporosis,” “bone metastatic cancer.” In terms of time and language, articles published from 2002 until 2018 were assessed and only publications in English were included in this study.
Screening and Selection
A set of inclusion and exclusion criteria were used to retrieve relevant in vitro studies. All in vitro studies involving exogenously added cytokine driven and spontaneous osteoclast formation from peripheral blood from patients with periodontitis or rheumatoid arthritis or psoriatic arthritis or osteoporosis or Crohn's disease or bone metastatic cancer were included for further examination.
The titles and abstracts of all publications identified by the electronic search were manually screened and discussed by two reviewers (IB and TdV). When the suitability of an article for this review could not be determined based on the title and abstract, the full text was read and examined by one reviewer (IB) and reported and discussed (IB and TdV). Outcomes of in vitro studies included comparison between osteoclast formation from peripheral blood of patients with one of the diseases listed above and healthy controls. All articles that did not meet with the primary outcome of interest, (spontaneous) osteoclast formation from PBMCs or monocytes of diseased patients compared to healthy controls were excluded, leaving 29 papers in the core-collection (Figure 1).
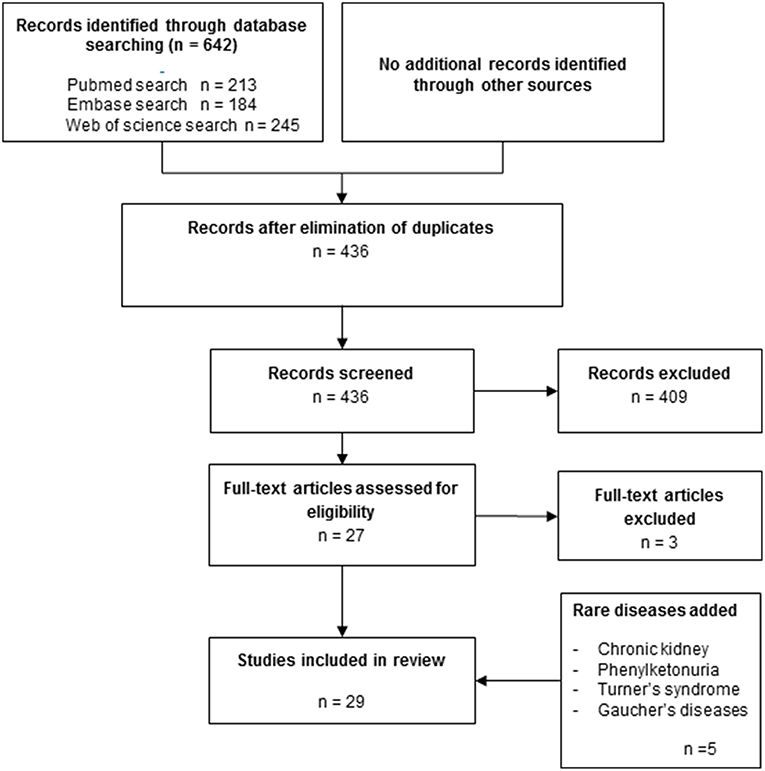
Figure 1. Flow chart of the literature search strategy.
Apart from the systematic approach on the common diseases osteoporosis, periodontitis and rheumatoid arthritis, literature on less frequent diseases (Turner, Gaucher, chronic liver disease, Crohn's disease, and phenylketonuria) that was found with this search strategy was incorporated.
Peripheral Blood Osteoclast Formation Is Increased in a Wide Spectrum of Bone Diseases
Overall it can be concluded that osteoclast formation from peripheral blood cells is increased in a wide range of diseases (Table 1). Results per disease or group of diseases are discussed in detail below.
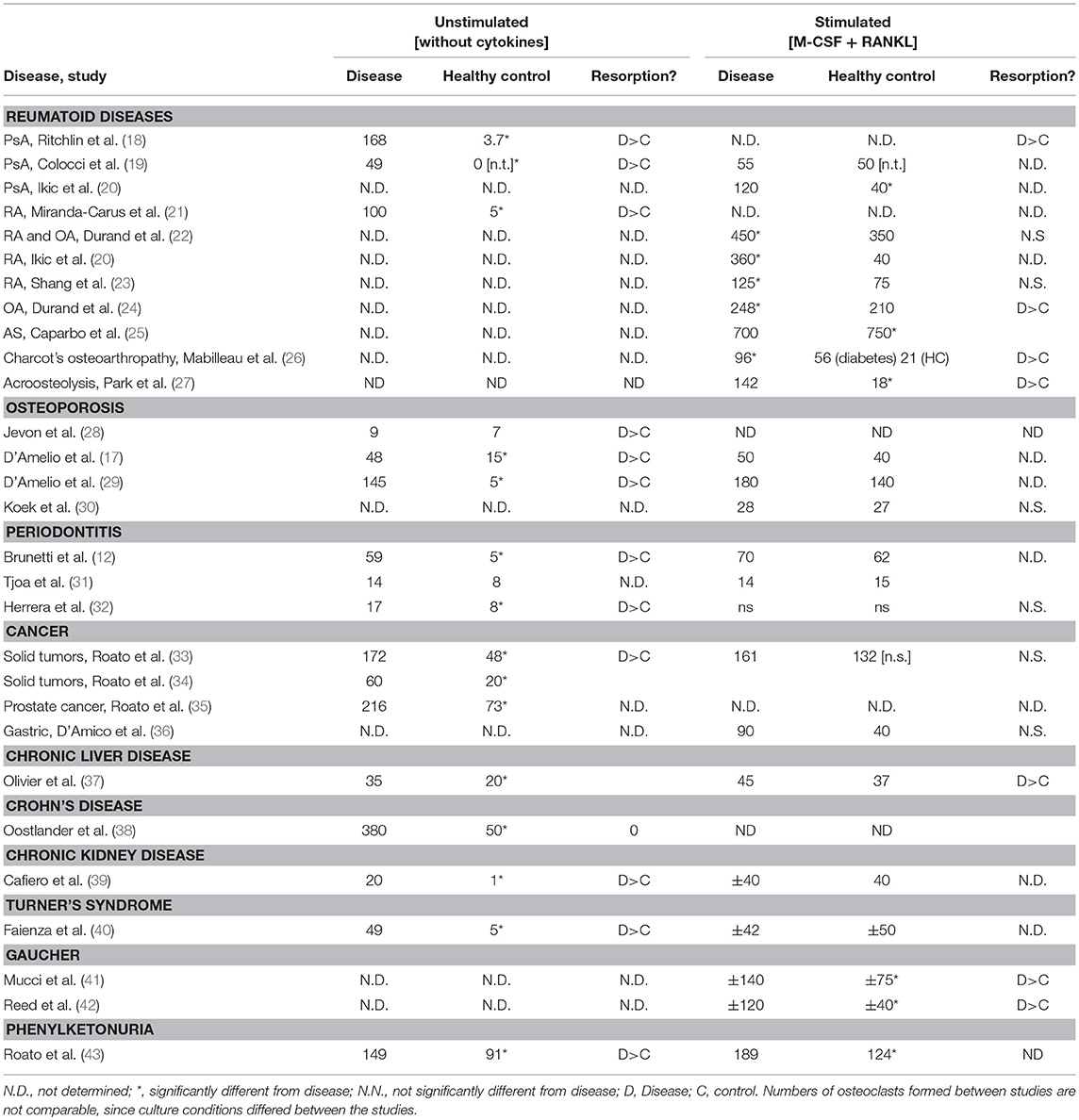
Table 1. Unstimulated and stimulated osteoclast formation is increased in peripheral blood from patients with bone loss.
Rheumatic Diseases
Rheumatoid Arthritis
Rheumatoid arthritis is a chronic inflammatory joint disease that is characterized by chronic synovitis and exaggerated local and systemic bone loss. It was established by Gravallese et al., that osteoclasts accumulate in the joints of RA patients (44) and that RANKL, the key osteoclastogenic mediator, is expressed locally in the joints of RA patients (45). Systemically, increased circulating osteoclast precursors have been reported in RA, which express key osteoclastogenic molecules on their surface (46, 47). Notably, when exposing similar numbers of peripheral CD14+ cells to osteoclastogenic conditions, more osteoclasts form in RA patients than in healthy controls (20), which is a strong argument that these peripheral osteoclast precursors are primed in the circulation. Moreover, elevated osteoclast formation in RA was shown to be correlated to local and systemic bone loss in RA (23). In addition, data showed that RA affects the longevity of osteoclasts, with a significantly lower number of osteoclasts undergoing apoptosis in RA compared to the healthy controls (22).
Enhanced osteoclastogenesis in RA essentially depends on T cells (48). Thus, in T-cell (CD3+ cells) depleted cultures, osteoclast precursors from RA patients show significantly lower spontaneous differentiation. Addition of exogenous RANKL to these cultures resulted in partial recovery of osteoclast formation. Together, these data indicate a crucial role for T-cells in osteoclast formation in RA, in line with earlier studies that demonstrated the activating role of T-cells in osteoclast formation (8, 49) which initiated osteoimmunology research. Miranda-Carus et al. (21) showed that T-cells present in peripheral blood play a major role in the formation of osteoclasts in RA patients. They interact with the osteoclast precursors of the monocyte/ macrophage lineage in vitro. An increased level of TNF-α, IL-1, IL-17, and RANKL was also found in the autologous T-cell/monocyte co-cultures derived from patients with early RA and established RA compared with controls (21) suggesting that these cytokines drive osteoclast differentiation in RA. Especially CD4+ T-cells play a role in osteoclastogenesis. OPG, anti-TNF-α and anti-IL-1 in this study, were shown to inhibit osteoclast formation (21).
Psoriatic Arthritis
Psoriatic arthritis (PsA) is a chronic inflammatory joint disease, which develops in about 30% of patients with psoriasis and is characterized by local bone erosions and systemic bone loss. TNFa and the interleukins 17 and −23 are key mediators of bone loss in PsA (50). In 2003, Ritchlin et al. (18) showed that the number of osteoclast precursors is increased in patients with PsA. In part, the development of osteoclasts in PsA could be independent from M-CSF or RANKL. This could be explained by a higher number of circulating OC precursors or by the ability of maturation of osteoclast precursors without supplementation with exogenous levels of M-CSF or RANKL. Ritchlin et al., also showed strong RANKL expression in synovial tissue of PsA patients as well as strong RANK expression in osteoclasts at areas with bone erosion. Fewer osteoclasts formed from blood of PsA patients after anti-tumor necrosis factor-α (TNF-α) treatment, indicating that TNF-α primes OC precursors in PsA (16). The original findings of Ritchlin et al., were also corroborated by Colucci et al., who showed that more TRAP+ multinucleated cells formed in cultures from PsA patients. Interestingly, supplementation of inflammatory cytokines abrogated these differences between PsA patients and controls (Table 1). This finding indicates an intrinsically higher osteoclastogenesis potential of blood from PsA patients. Increased osteoclast formation in PBMCs appears to be T-cell dependent, since the formation of osteoclasts was absent in T-cell-depleted PBMC cultures, which implies that T-cells are responsible for osteoclast formation. These results also suggest that blockade of RANKL and TNF-α might be used as an effective strategy for inhibiting enhanced osteoclastogenesis in PsA patients (19). Indeed such concept is supported by data on TNF inhibitors, which effectively inhibit osteoclast formation (51). Notably, aside from inflammation, PsA is strongly linked to obesity and the metabolic syndrome. In this context it is interesting that Xue et al., found an elevated value of certain adipokines, cytokines derived from adipose tissues, in the circulation of patients with PsA (52). Adipokines like leptin, adiponectin, chemerin, and omentin may not only play a role in inflammation but also in osteoclastogenesis in patients with PsA. Higher levels of leptin and omentin, for instance, positively correlated with the number of osteoclast precursors found in PsA patients, while adiponectin was negatively correlated with osteoclast precursors.
Osteoarthritis
In osteoarthritis (OA) (24), the key degenerative joint disease, it was found that the number of TRACP+ multinucleated osteoclasts is higher than in healthy controls providing an explanation for the sometimes bone erosive phenotype of OA. No significant difference was found between OA and healthy controls regarding the number of circulating CD14+ cells. However, osteoclasts from the OA patients were found capable of resorbing a significantly larger (4 times) area than the osteoclasts from the healthy controls. The higher number of osteoclast-like cells formed by the PBMCs from the osteoarthritis group compared to the PBMCs from the control group could be responsible for enhanced local bone resorption in OA.
Though strictly speaking not a rheumatoid disease, Charcot's arthropathy that is associated with diabetes co-incides with joint erosions, in particular of the foot (53). Charcot's arthropathy is associated with increased peripheral blood osteoclast formation, with more osteoclasts than in matched diabetes patients without Charcot's arthropathy is or healthy controls (26). A later study from the same group showed that higher numbers of CD14+ cells prevail in blood of patients with Charcot foot, concomitant with higher peripheral blood TNF-α levels (53).
Acroosteolysis
Also patients suffering from acroosteolysis, which is part of the systemic sclerosis disease spectrum, show increased osteoclast formation compared to healthy controls. In this context, the increased osteoclast formation is associated with higher VEGF levels in the peripheral blood (27). VEGF can substitute for M-CSF in driving osteoclast differentiation (54).
Ankylosing Spondylitis
Ankylosing spondylitis is an inflammatory rheumatic disease of the spine, which is characterized by loss of trabecular bone but periosteal apposition of cortical bone leading to bony spur formation sometimes leading to fusion of vertebra (55). Interestingly, ankylosing spondylitis is the only rheumatic disease where data showed that less osteoclasts form in vitro and where data on serum CTX levels show that lower overall bone resorption happens. Lower osteoclast numbers correlated with lower RANKL/OPG ratios. Furthermore, osteoclasts from ankylosing spondylitits patients were less prone to apoptosis (25) which has also been observed in other rheumatic diseases (24).
Osteoporosis
Osteoporosis is a skeletal disease characterized by lower bone mass and micro-architectural deterioration of bone leading to an increased risk of fractures (56). Several indications show that this phenomenon is caused by a higher activity of osteoclasts due to an imbalance of the osteoclasts and osteoblasts, postmenopausal bone loss is triggered by estrogen deficiency that increased osteoclastogenesis through several pathways, and in particular by the activation of T cells that produce higher level of pro-inflammatory and pro-osteoclastogenic cytokines as TNFα and RANKL (57). Bone fractures are the severe consequence of osteoporosis and represent a major health problem in the increasingly older (58). Old patients experiencing a femoral fracture have a decreased life expectancy and may become care-dependent in half the survivors. The presence of a fragility fracture increases the risk of new fractures creating a “domino effect”: one vertebral fracture doubles the risk of subsequent femoral fracture within a year, the presence of vertebral fractures as well as of femoral fracture impair patients' quality of life and increase mortality.
In a first study comparing the peripheral blood osteoclast formation from patients and controls, similar numbers of osteoclasts formed, but the resorptive capacity was higher in osteoclasts from osteoporosis patients (28), suggesting that the peripheral OCp had the same osteoclastogenic potential, but were somehow primed to form more active osteoclasts (28). D'Amelio et al. (17, 57) investigated the osteoclast formation in osteoporosis. The first study compared the osteoclast formation in osteoporotic women compared to healthy controls, without adding M-CSF, TNF-α, or RANKL to the cultures (17). A higher number of osteoclasts was formed in the cultures from osteoporosis blood compared to healthy controls. After supplementation of M-CSF and RANKL the numbers of osteoclasts reached a same level in controls and patients. A significantly higher level of TNF-α and RANKL was found in the PBMC cultures of the osteoporotic group. Adding 1,25-OH vitamin D3 to the PBMCs cultures resulted in both groups in lower numbers of osteoclasts, but higher resorption. The lacunar resorption area was significantly higher in the osteoporotic patients group compared to the healthy subjects with and without the addition of 1,25-OH vitamin D3. Comparable results were reported in the second paper by the same group (29): higher levels of TNF-α and RANKL in the PBMC cultures and higher osteoclast formation in the osteoporotic group compared to the healthy subjects. Antibodies against TNF-α and RANKL decreased spontaneous osteoclast formation strongest in the osteoporotic group. Additionally this study demonstrated that the T-cells of osteoporosis patients play a key role in the osteoclastogenesis by increasing the TNF-α and RANKL production. It was shown that osteoclast formation was severely suppressed when depleting the T-cells from the PBMCs cultures. This indicates that T-cells play a crucial role in osteoclast formation and that they secrete cytokines necessary for osteoclast formation in osteoporosis. In a fourth study, no differences in osteoclast formation were found between controls and osteoporotic patients (30), but in this case only M-CSF and RANKL stimulated cultures were studied. This was in line with the previous studies (17, 28, 57) regarding the stimulated osteoclastogenesis, where addition of M-CSF and RANKL may shield possible differences.
Periodontitis
Periodontitis is the inflammatory bone-destructive disease affecting the alveolar bone between teeth. It's individual susceptibility is driven by an oral bacterial dysbiosis, genetic factors (59) and life style (60). Currently, it is estimated that no <46% of Americans adults have moderate to severe periodontitis (61). When peripheral blood mononuclear cells (PBMC) from chronic periodontitis patients were cultured in the absence of M-CSF and RANKL, more osteoclast-like cells form from chronic periodontitis patients (12). Osteoclast-like cells in the control group were fewer and smaller. The addition of stimulating factors M-CSF and RANKL resulted in comparable numbers of osteoclast-like cells in control and periodontitis group. M-CSF and RANKL only triggered osteoclast formation in the control group, osteoclast numbers of the periodontitis group did not increase after cytokine treatment. The osteoclasts formed spontaneously from the PBMC cultures from the periodontitis patients and showed a significantly higher resorbtive activity compared to the controls. A T-cell dependent osteoclastogenesis was shown in this study, since the number of osteoclasts was low in the T-cell depleted unstimulated PBMCs cultures from the periodontitis patients. Addition of stimulating factors M-CSF and RANKL to these cultures led to a significantly higher formation of numerous large osteoclasts. An explanation for this finding is the overexpression of RANKL and TNF-α by T-cells, which was shown to be higher by the T-cells from the patient group. Addition of anti-RANKL and anti-TNF-α antibodies induced a dose dependent inhibition of the osteoclast formation in the periodontitis group (12). Also Tjoa et al., aimed to determine if there was a difference in osteoclast formation between the PBMC from patients suffering from chronic periodontitis and matched healthy controls (31). In this study, no differences were observed in unstimulated osteoclast formation between controls and periodontitis patients. A significant difference was shown however after the stimulation with M-CSF between control and diseased group. Larger and more multinucleated cells were found in the control group, whereas the patient group was insensitive to stimulation with M-CSF (31), suggesting that the OCp in this group were already primed in the circulation. In another study, it was shown that peripheral blood monocytes from periodontitis patients are more prone to differentiate into mature multinucleated osteoclasts (32). In other words, this study shows that these monocytes were primed in peripheral blood and prepared here for enhanced osteoclast formation. Only the stimulation with RANKL and not with M-CSF and RANKL gave significant differences between the periodontitis group and healthy controls regarding the osteoclast-like cells formed. A significantly higher level of M-CSF was found in the periodontitis group (32). This could be an explanation for the insensitive response of PBMC from periodontitis patients to stimulation with M-CSF in the study of Tjoa et al. (31), since the osteoclast precursors could be already pre-activated by higher M-CSF levels present in serum.
Cancer That Metastasizes to Bone
Hematological Cancers
B cell multiple myeloma was studied by Colucci et al. who cultured 10 times more osteoclasts in the absence of added cytokines from myeloma blood than from control blood. As described repetitively above, this difference was no longer seen when the M-CSF and RANKL was added to these cultures. Colluci et al., found that peripheral blood contained upregulated OPG and RANKL levels. OPG co-precipitated with TRAIL, by which more RANKL became available. Autologous T-cells added to these osteoclasts, prolonged survival of osteoclasts (11).
Solid Tumor
Using blood from patients with bone metastasis from diverse primary tumors such as melanoma, lung, prostate, kidney, breast, and colon, it was found that these cultures gave rise to more osteoclast compared to controls when no cytokines were added (33). Addition of M-CSF and RANKL nullified this effect. OCp from tumor patients were further characterized and apart from CD14 and CD11, these precursors, and not in the control group, also expressed the osteoclast marker the vitronectin receptor αvβ3, which could be typical for precursors that are a bit further differentiated, indicating that the OCp in peripheral blood of cancer patients are already a step further in differentiation. Addition of OPG did not inhibit unstimulated osteoclast formation (33). It was shown that anti-TNF blocked osteoclast formation (33), which was recently confirmed in a separate study (51) Only T-cells from osteolysis patients expressed TNF-α, and also osteoclasts derived from cancer patients expressed TNF-α (33). The same group showed in subsequent years with a relatively large cohort of heterogeneous tumors (34) and in a group of prostate cancer (35) that most osteoclasts are formed without stimulation in patients with bone metastasis, compared to cancer patients without metastasis, and least osteoclasts formed from blood from controls. In both studies, a role of IL-7 was described. IL-7 serum levels were high in patients with metastasis, lower in sera from patients without and lower in control sera. T-lymphocytes could be identified as the source for IL-7, antibodies against IL-7 significantly inhibited osteoclast formation (34, 35). In a group of gastric cancer patients, no differences were observed in numbers of osteoclasts that differentiation from blood from patients with metastases, from patients without metastases and controls (36). An important difference with the other solid tumor studies was that only cytokine stimulated conditions were considered.
Rarer Bone Diseases Associated With Increased Bone Loss
Diseases Associated With Osteopenia: Chronic Liver Disease, Crohn's Disease and Chronic Kidney Disease
Chronic liver disease can lead to osteoporosis, but not in all individuals with chronic liver disease. In attempt to explain this phenomenon, osteoclasts were cultured from blood of chronic liver disease patients with osteopenia, without osteopenia and matched healthy controls. More osteoclasts formed without adding osteoclastic cytokines M-CSF and RANKL from blood of osteopenic patients compared to non-osteopenic and healthy controls (37). Interestingly, and in line with what was found in the periodontitis study from the same group (31), only controls responded with higher osteoclast numbers when stimulated with M-CSF, whereas both osteopenic and non-osteopenic patients did not respond to addition of M-CSF. Serum levels M-CSF of both patient groups were significantly higher than controls, suggesting M-CSF priming of OCp in peripheral blood. Furthermore, number of osteoclasts cultured in vitro, correlated negatively with the lumbar spine density score (37).
Though Crohn's disease is an intestinal disease, it is associated with inflammatory flair-ups periods. Some of the patients develop osteoporosis, including those who are not on high levels of corticosteroids. In a study with patients in a quiescent disease stage, Oostlander et al. (38) were the first to describe the pre-stages of unstimulated osteoclast formation. It was shown that the formation of osteoclasts is preceded with a stage of cell clusters, the number of which correlate with the numbers of osteoclasts that form (38). In a similar approach as in a previous study (57), OCp were either cultured with purified autologous B-cells, T-cells or the combination or without. Osteoclasts only formed in combinations where T-cells were present, which also made part of the cell clusters that preceded osteoclast formation. More osteoclasts formed from OCp:T-cell cultures from Crohn's disease than from controls and correlated to IL-17 levels in vitro. TNF-α levels were highest in OCp+ T-cells, compared to T-cells alone (no secretion) or monocytes only (lower levels), indicating that the T-cell:OCp interaction induces TNF-expression (36), later confirmed by Moonen et al. (3). Interestingly, the diverse combinations of T-cells, B-cells, or T-Cells + B-cells did not affect control monocytes, indicating that the osteoclastogenesis driving T-cell activity is increased in Crohn's disease.
Bone disease in patients with chronic kidney disease (CKD) is a major clinical concern due to its prevalence and consequences that greatly impact patients quality of life (62). CKD patients may be affected by both higher and lower bone turnover disease, in patients with high bone turnover disease increased osteoclastogenesis is sustained by both increased in inflammatory and pro-osteoclastogenic cytokines and by increased PTH due to the decreased ability of kidney to hydroxylate vitamin D into its active form 1,25OHvitamin D. Patients with terminal kidney failure who are on dialysis, experience a chronically inflammatory state with often skeletal complications. Osteoclast formation was studied both without and cytokine stimulation (39). Osteoclast formation was lowest in controls, and higher in early chronic patients and higher in late chronic patients and highest in hemodialysis patients, similar correlations were seen with resorptive capacity. RANKL was expressed on T-lymphocytes in renal patients, not in controls. RANK-Fc can inactive RANKL, and when added it dose dependently decreased osteoclastogenesis, demonstrating that osteoclast formation was RANKL dependent (39).
Rare Diseases Associated With Bone Loss: Turner's Syndrome, Gaucher's Disease and Phenylketonuria
Patients with Turner's syndrome may present with decreased bone due to hypergonadism associated with the disease. Estrogen deficiency could be the course for such bone loss (40). Unstimulated osteoclast formation resulted in more osteoclasts that were more active, both in the group with high levels of follicle stimulating hormone (FSH) and with low levels. These osteoclasts resorbed more calcium phosphate. Osteoclast numbers from monocytes with addition of M-CSF and RANKL were similar between controls and patients. The increased unstimulated osteoclast formation in Turner's syndrome correlated with a lower percentage of osteoclastogenesis inhibitory CD4+CD25+ cells and a higher percentage of CD3+ NKT cells. The CD8+TNF-α+ cells were higher in Turner's syndrome, as well as the CD14+TNF-α+ monocytes. This skewness could contribute and be responsible for the increased osteoclast formation (40).
Gaucher's disease is a heritable disease with deficiency of the lysomal enzyme glucocerebrosidase, resulting in excess of glycosylceramide, which is then stored in high quantities in macrophages, concomitant with a disturbed immunological balance and cytokine secretion profiles (42). Approximately 3-fold more osteoclasts formed from Gaucher patient's PBMC cultured with M-CSF and RANKL. These osteoclasts were larger in size and number of nuclei and resorbed larger areas of bone. When distinguishing between active and non- active bone disease, the Gaucher patients with active bone disease formed more osteoclasts. Especially the Gaucher cultures were relatively independent of M-CSF (42), in line with data from periodontitis (31) and chronic liver disease patients (37). When control PBMC were cultured with inhibitors of glucocerebrosidase, increased numbers of osteoclast formed (42), indicating that this enzyme plays a key role in tempering osteoclast numbers. The increased osteoclast formation in Gaucher's disease was confirmed by Mucci et al. (41). Here, it was shown that Gaucher's disease blood contained a higher percentage of non-classical/inflammatory CD14+CD16+ cells compared to the classical CD14+CD16– monocytes. When culturing with an enzyme that replenishes the missing glucocerebrosidase, osteoclast numbers decreased only in Gaucher patients cultures, again indicating that glucocerebrosidase tempers osteoclast formation. T-cells from Gaucher's disease express more RANKL (41). Osteoclast cultures from controls were insensitive to OPG or anti-TNF-α treatment, whereas osteoclast numbers went significantly down in Gaucher's PBMC cultures that were treated with OPG or anti-TNF-α (41).
Phenylketonuria (PKU) is a rare, inherited disease with a defect in the synthesis of the amino acid phenylalanine. These patients have a hitherto not understood progressive bone impairment. Also with this disease, increased osteoclasts were cultured both without osteoclastogenesis stimulating cytokines and with these cytokines (43). As shown for Gaucher (41), the blood of PKU also contains a higher proportion of CD14+CD16+ monocytes, which co-express CD51/CD61, or αvβ3 integrin, that is typical for osteoclasts (63). The unstimulated osteoclastogenesis cultures contained increased levels of TNF-α and RANKL in PKU patients. Only RANK-Fc, that blocks RANKL activity, decreased osteoclast numbers in PKU cultures. PKU patients contained activated T- cells of the CD4+CD25+CD69+ signature, a cell type that was absent in controls.
General Osteoclastogenesis Features of Inflammatory Bone Diseases
When summarizing the osteoclastogenesis capacity of the various inflammatory bone disease, several features become apparent (Table 1). Firstly, when taking together all 29 summarized studies, more osteoclasts formed, be it unstimulated or stimulated with M-CSF and RANKL. This was the case for 24 of the 29 studies. Secondly, in all cases where resorptive activity was determined, the osteoclasts from bone loss diseases were more active. Thirdly, osteoclasts can be cultured without exogenous addition of M-CSF and RANKL, only when cultured from PBMC or at least the addition of T-cells to OCp. These osteoclasts often displayed lytic activity of calcium phosphate coatings, but also bone resorption activity has been reported. Fourthly, those studies that have compared unstimulated and M-CSF and RANKL stimulated osteoclast formation, often found increased osteoclast formation in unstimulated cultures, whereas these differences were often not found any more when cultured with M-CSF and RANKL. This accounted for a variety of diseases, such as psoriatic arthritis (19), osteoporosis (17, 57), periodontitis (12, 31), multiple myeloma (11), solid tumors (33), chronic liver disease (37), kidney disease (39), and Turner's syndrome (40). This suggests that stimulation with unphysiological levels of M-CSF and RANKL may hide the intrinsically increased osteoclastogenic activities present in peripheral blood. Especially these studies with self-stimulatory osteoclastogenesis cocktails of T-cells and OCp can give us clues of common and disease specific determinants of increased osteoclast formation. Below, three common denominators which stood out when comparing the various inflammatory bone diseases, being (1) inflammatory mediators in serum, (2) the role of T-cells and (3) differential priming or skewness in monocyte distribution are worked out for the inflammatory bone diseases.
Common Inflammatory Mediators in Serum Prepares OCp in the Circulation
A different priming whilst in the circulation by increased levels of pro-osteoclastogenic mediators could make monocytes more equipped to differentiate into osteoclasts. Most commonly described is the increased presence of TNF-α, see the above paragraph where it's presence is discussed in the context of T-cells, but also in serum from patients with Charcot's disease, where it co-incides with increased numbers of CD14+ cells (53). Another such pro-osteoclastogenic cytokine is M-CSF. Higher levels of M-CSF in the circulation have been described for periodontitis (32) and chronic liver disease (37). This could make PBMC from these patients less responsive to exogenous M-CSF (31, 37).
A Common Role for T-Cells in Osteoclast Formation in the Various Inflammatory Bone Diseases
The general role of T- cells in osteoclast regulation (64), the role of osteoclast activating T-cells such as Th17 (65) and the regulatory role of Treggs (66) have been reviewed elsewhere. Here, we describe the T-cell findings in the context of inflammatory diseases with bone loss in the context of the core collection used for this review. When reviewing the above literature it is clear that T-cells are indispensable for spontaneous osteoclastogenesis: without T-cells, no osteoclasts form. This has been investigated for psoriatic arthritis (19, 48), osteoporosis (57), periodontitis (12) and Crohn's disease (38). The mechanism by which they adhere and stimulate OCp is likely by LFA-1: ICAM-1 interaction, since antibodies against LFA-1 interfere both with cell cluster formation and with osteoclast formation (38). TNF-α is most commonly reported as the cytokine secreted by T-cells in the various diseases. T-cells from patients with inflammatory bone loss diseases secrete more TNF-α, as shown for psoriatic arthritis (19), osteoporosis (57), periodontitis (12), and Turner's syndrome (40). Exclusively the T-cells from peripheral blood from patients with osteolytic solid tumors and not those without osteolytic tumors were reported to express detectable levels of TNF-α (34). Treating unstimulated osteoclastogenesis cultures with anti TNF-α agent infliximab reduces both cell cluster formation and osteoclast formation (51). Anti-TNF-α treatment also decreased osteoclast formation in patients with RA (18), periodontitis (12), and Gaucher's disease (41). T-cells from inflammatory bone diseases express more RANKL, as reported for RA (21), osteoporosis (57), and chronic kidney disease (39). Anti RANKL reduced osteoclast formation in Gaucher's disease more than in controls (41), and also in phenylketonuria patients (43), chronic kidney disease (39). Studies with RANKL-independent osteoclast formation were also reported (33, 37). In these studies, OPG or RANK-Fc did not affect osteoclast formation. Two osteoclastogenesis studies from solid tumors report spontaneous osteoclastogenesis stimulated by T-cell secreted IL-7 (34, 35). This has not been studied in the other inflammatory bone loss diseases. Finally, it has been reported for myeloma derived osteoclasts that autologous T-cells added to osteoclast cultures may prolong osteoclast survival (11).
A Different Distribution of Monocytes Subtypes and Different Monocyte Priming in Inflammatory Diseases
A third commonality between the different diseases is that due to the inflammatory disease, a skewed distribution of monocytes that is better equipped to differentiate into osteoclasts populate the peripheral blood. The existence of different monocyte populations (2, 67) which distributions are then different between disease and controls, is indeed a very attractive explanation for the outcome of more osteoclasts in inflammatory disease. One way to achieve a relatively crude and likely heterogeneous populations of OCp is with CD14+ positive isolation. Studies that show more osteoclasts as outcome, where equal numbers of purified CD14+ monocytes were uses in disease vs. healthy controls, provide a first indication for a better equipment of these cells for osteoclast differentiation. This has been shown for RA and psoriatic arthritis (20) and for periodontitis (32). When comparing monocyte heterogeneity, two different criteria have been used in the articles assessed in this review: an approach based on CD14, CD11b, and the vitronectin receptor (VNR) expression and the more commonly used CD14/CD16 expression of monocytes. One study that has used CD14+ in conjunction with CD11b and the vitronectin receptor or αvβ3 showed that peripheral blood of osteolytic cancers contained more VNR+ monocytes, which correlated to higher osteoclast formation (33).
The much more commonly used classification of monocytes is based on their CD14 and CD16 expression. The CD14+CD16– are the classical monocytes that become the phagocytic cells in tissues and comprise the majority of blood monocytes, intermediate monocytes are CD14+CD16+ and are pro-inflammatory and play a role in wound healing, and the CD14+CD16++ cells are the non-classical monocytes that play a role in patrolling and fibrosis (2, 67). Initial description on either CD16– and CD16+, before the subsequent refinement of CD16+ monocytes into intermediate and non-classical monocytes (68) and also recent literature on the transcriptome of the classical and non-classical monocytes (69) suggest that the physiological osteoclasts derive from classical, CD14+CD16– monocytes. This was refined recently by Sprangers et al., who found that all three monocyte subtypes differentiate equally well on plastic, but that only the classical and intermediate ones form resorbing osteoclasts on bone (6). On bone slices, non-classical monocytes hardly differentiated and no resorption was observed. For some of the inflammatory bone loss diseases, however, several studies indicate that likely the CD16+ monocytes are important for osteoclast formation. A skew distribution compared to healthy controls was demonstrated for several inflammatory bone loss diseases. It was demonstrated for multiple myeloma, that CD14+CD16+ monocytes are predominant in active disease compared to smoldering disease. These cells gave rise to larger osteoclasts (70). Also in kidney disease, more CD14+CD16+ monocytes were found in peripheral blood, and exclusively more “inflammatory” monocytes expressed RANKL, providing the possibility for auto-stimulation (39). Patients with Gaucher's disease (41) and patients with phenylketonuria (43) have more CD14+CD16+ monocytes in peripheral blood than matched controls. All the above studies are association studies, where a higher percentage of CD14+CD16+ in blood of patients is associated with increased osteoclast formation. To prove that these cells indeed give rise to more osteoclasts, Chiu et al., have sorted the three subtypes of OCp from psoriatic arthritis patients and healthy controls (71). Interestingly, the CD16+ monocytes from controls gave rise to low levels of osteoclasts, whereas the CD16+ monocytes from patients gave rise to high numbers of osteoclasts. This suggests that apart from the common distinction of OCp with the CD14 and CD16 markers, other features have been acquired by psoriatic arthritis patients, making them more equipped to differentiate into osteoclasts. An experiment with healthy control OCps confirmed the relative inertness of CD16+ monocytes in healthy controls. When adding increasing numbers of CD16+ monocytes to a constant number of CD14+CD16– monocytes in an osteoclastogenesis experiment, there was no increase in osteoclast numbers, suggesting that under these conditions, only the CD14+CD16– monocytes contributed to the formation of a syncytium (72).
Concluding Remarks on the Common Denominators for Increased Osteoclast Formation in Patients With Inflammation Related Bone Loss
In summary, this core collection of studies with well-matched bone-loss patient—healthy controls, unequivocally shows an increased osteoclast formation and activity in patients with inflammatory bone loss. In search for the conditions that give rise to this, several factors that are shared between diseases can be identified. First of all, the serum that surrounds OCp in the circulation is beneficiary for the differentiation of OCp. Second, T-cells that secrete TNF-α and RANKL are present in the circulation of patients. In light of the fact that inflammatory bone loss diseases may occur simultaneously, anti-TNF-α treatment could benefit more than one disease, as was shown for instance in rheumatoid arthritis patients receiving anti-TNF-α medication infliximab, who had lower periodontal indices. Finally, the patient blood may contain more OCp and differently primed OCp, probably containing the CD14+CD16+ phenotype rather than CD14+CD16–. On top of that, either disease specific or a general immunological stimulus has given the OCp from bone loss patients a profile to facilitate enhance osteoclast formation (Figure 2). Future research will likely uncover the barcodes of the OCps in the various inflammatory diseases associated with bone loss. This knowledge of the biological mechanisms underlying the alterations of monocytes and osteoclasts will likely reveal future therapeutic targets that will specifically target the immune system-steered osteoclast formation.
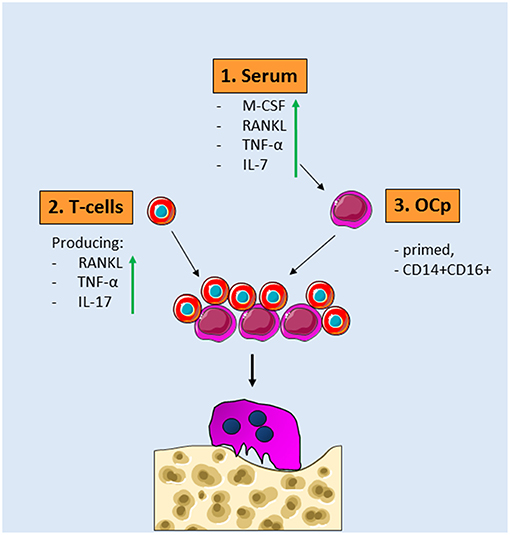
Figure 2. Common denominators for osteoclast formation in diseases with inflammatory bone loss. 1. Serum of patients with inflammatory bone loss diseases contains more osteoclastogenesis stimulating factors such as RANKL, TNF-α, IL-7, and M-CSF. 2. These serum factors prime OCp present in peripheral blood. These OCp are skewed toward more CD14+CD16+ cells. 3. T-cells from patients with inflammatory bone diseases express more IL-17, RANKL, and TNF-α. When these different OCp and T-cells—both of them different from controls—are added together, more osteoclasts are formed that are more active in bone resorption.
Author Contributions
TV wrote most of the manuscript and coordinated feedback. IB performed literature search and drafted the initial parts of the manuscript. TK commented on earlier versions of the manuscript. GS critically evaluated the rheumatology part. IJ critically evaluated all stages of the manuscript. GS and PD put the manuscript in a clinical context. PD wrote the sections on osteoporosis.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Boyce BF, Yao Z, Xing L. Osteoclasts have multiple roles in bone in addition to bone resorption. Crit Rev Eukaryot Gene Expr. (2009) 19:171–80. doi: 10.1615/CritRevEukarGeneExpr.v19.i3.10
2. Sprangers S, de Vries TJ, Everts V. Monocyte heterogeneity: consequences for monocyte-derived immune cells. J Immunol Res. (2016) 2016:1475435. doi: 10.1155/2016/1475435
3. Moonen CGJ, Alders ST, Bontkes HJ, Schoenmaker T, Nicu EA, Loos BG, et al. Survival, retention, and selective proliferation of lymphocytes is mediated by gingival fibroblasts. Front Immunol. (2018) 9:1725. doi: 10.3389/fimmu.2018.01725
4. Mbalaviele G, Novack DV, Schett G, Teitelbaum SL. Inflammatory osteolysis: a conspiracy against bone. J Clin Invest. (2017) 127:2030–9. doi: 10.1172/jci93356
5. De Vries TJ, Schoenmaker T, Hooibrink B, Leenen PJ, Everts V. Myeloid blasts are the mouse bone marrow cells prone to differentiate into osteoclasts. J Leukoc Biol. (2009) 85:919–27. doi: 10.1189/jlb.0708402
6. Sprangers S, Schoenmaker T, Cao Y, Everts V, de Vries TJ. Different blood-borne human osteoclast precursors respond in distinct ways to IL-17A. J Cell Physiol. (2016) 231:1249–60. doi: 10.1002/jcp.25220
7. Salamanna F, Maglio M, Borsari V, Giavaresi G, Aldini NN, Fini M. Peripheral blood mononuclear cells spontaneous osteoclastogenesis: mechanisms driving the process and clinical relevance in skeletal disease. J Cell Physiol. (2016) 231:521–30. doi: 10.1002/jcp.25134
8. Kong YY, Feige U, Sarosi I, Bolon B, Tafuri A, Morony S, et al. Activated T cells regulate bone loss and joint destruction in adjuvant arthritis through osteoprotegerin ligand. Nature. (1999) 402:304–9. doi: 10.1038/46303
9. Yasuda H, Shima N, Nakagawa N, Mochizuki SI, Yano K, Fujise N, et al. Identity of osteoclastogenesis inhibitory factor (OCIF) and osteoprotegerin (OPG): a mechanism by which OPG/OCIF inhibits osteoclastogenesis in vitro. Endocrinology. (1998) 139:1329–37. doi: 10.1210/endo.139.3.5837
10. Xiong J, Onal M, Jilka RL, Weinstein RS, Manolagas SC, O'Brien CA. Matrix-embedded cells control osteoclast formation. Nat Med. (2011) 17:1235–41. doi: 10.1038/nm.2448
11. Colucci S, Brunetti G, Rizzi R, Zonno A, Mori G, Colaianni G, et al. T cells support osteoclastogenesis in an in vitro model derived from human multiple myeloma bone disease: the role of the OPG/TRAIL interaction. Blood. (2004) 104:3722–30. doi: 10.1182/blood-2004-02-0474
12. Brunetti G, Colucci S, Pignataro P, Coricciati M, Mori G, Cirulli N, et al. Grano. T cells support osteoclastogenesis in an in vitro model derived from human periodontitis patients. J Periodontol. (2005) 76:1675–80. doi: 10.1902/jop.2005.76.10.1675
13. Takayanagi H, Iizuka H, Juji T, Nakagawa T, Yamamoto A, Miyazaki T, et al. Involvement of receptor activator of nuclear factor kappaB ligand/osteoclast differentiation factor in osteoclastogenesis from synoviocytes in rheumatoid arthritis. Arthritis Rheum. (2000) 43:259–69. doi: 10.1002/1529-0131(200002)43:2<259::Aid-anr4>3.0.Co;2-w
14. Sokos D, Everts V, De Vries TJ. Role of periodontal ligament fibroblasts in osteoclastogenesis: a review. J Periodontal Res. (2015) 50:152–9. doi: 10.1111/jre.12197
15. Kim N, Kadono Y, Takami M, Lee J, Lee SH, Okada F, et al. Osteoclast differentiation independent of the TRANCE-RANK-TRAF6 axis. J Exp Med. (2005) 202:589–95. doi: 10.1084/jem.20050978
16. Anandarajah AP, Schwarz EM, Totterman S, Monu J, Feng CY, Shao T, et al. The effect of etanercept on osteoclast precursor frequency and enhancing bone marrow oedema in patients with psoriatic arthritis. Ann Rheum Dis. (2008) 67:296–301. doi: 10.1136/ard.2007.076091
17. D'Amelio P, Grimaldi A, Pescarmona GP, Tamone C, Roato I, Isaia G. Spontaneous osteoclast formation from peripheral blood mononuclear cells in postmenopausal osteoporosis. FASEB J. (2005) 19, 410–2. doi: 10.1096/fj.04-2214fje
18. Ritchlin CT, Haas-Smith SA, Li P, Hicks DG, Schwarz EM. Mechanisms of TNF-alpha- and RANKL-mediated osteoclastogenesis and bone resorption in psoriatic arthritis. J Clin Invest. (2003) 111:821–31. doi: 10.1172/JCI200316069
19. Colucci S, Brunetti G, Cantatore FP, Oranger A, Mori G, Quarta L, et al. Lymphocytes and synovial fluid fibroblasts support osteoclastogenesis through RANKL, TNFalpha, and IL-7 in an in vitro model derived from human psoriatic arthritis. J Pathol. (2007) 212:47–55. doi: 10.1002/path.2153
20. Ikic M, Jajic Z, Lazic E, Ivcevic S, Grubisic F, Marusic A, et al. Association of systemic and intra-articular osteoclastogenic potential, pro-inflammatory mediators and disease activity with the form of inflammatory arthritis. Int Orthop. (2014) 38:183–92. doi: 10.1007/s00264-013-2121-0
21. Miranda-Carus ME, Benito-Miguel M, Balsa A, Cobo-Ibanez T, Perez DA, Pascual-Salcedo D, et al. Peripheral blood T lymphocytes from patients with early rheumatoid arthritis express RANKL and interleukin-15 on the cell surface and promote osteoclastogenesis in autologous monocytes. Arthritis Rheum. (2006) 54:1151–64. doi: 10.1002/art.21731
22. Durand M, Boire G, Komarova SV, Dixon SJ, Sims SM, Harrison RE, et al. The increased in vitro osteoclastogenesis in patients with rheumatoid arthritis is due to increased percentage of precursors and decreased apoptosis—the In Vitro Osteoclast Differentiation in Arthritis (IODA) study. Bone. (2011) 48:588–96. doi: 10.1016/j.bone.2010.10.167
23. Shang W, Zhao LJ, Dong XL, Zhao ZM, Li J, Zhang BB, et al. Curcumin inhibits osteoclastogenic potential in PBMCs from rheumatoid arthritis patients via the suppression of MAPK/RANK/c-Fos/NFATc1 signaling pathways. Mol Med Rep. (2016) 14:3620–6. doi: 10.3892/mmr.2016.5674
24. Durand M, Komarova SV, Bhargava A, Trebec-Reynolds DP, Li K, Fiorino C, et al. Monocytes from patients with osteoarthritis display increased osteoclastogenesis and bone resorption: the In Vitro Osteoclast Differentiation in Arthritis study. Arthritis Rheum. (2013) 65:148–58. doi: 10.1002/art.37722
25. Caparbo VF, Saad CGS, Moraes JC, de Brum-Fernandes AJ, Pereira RMR. Monocytes from male patients with ankylosing spondylitis display decreased osteoclastogenesis and decreased RANKL/OPG ratio. Osteoporos Int. (2018) 29:2565–73. doi: 10.1007/s00198-018-4629-z
26. Mabilleau G, Petrova NL, Edmonds ME, Sabokbar A. Increased osteoclastic activity in acute Charcot's osteoarthropathy: the role of receptor activator of nuclear factor-kappaB ligand. Diabetologia. (2008) 51:1035–40. doi: 10.1007/s00125-008-0992-1
27. Park JK, Fava A, Carrino J, Del Grande F, Rosen A, Boin F. Association of acroosteolysis with enhanced osteoclastogenesis and higher blood levels of vascular endothelial growth factor in systemic sclerosis. Arthritis Rheumatol. (2016) 68:201–9. doi: 10.1002/art.39424
28. Jevon M, Hirayama T, Brown MA, Wass JA, Sabokbar A, Ostelere S, et al. Osteoclast formation from circulating precursors in osteoporosis. Scand J Rheumatol. (2003) 32:95–100. doi: 10.1080/03009740310000102
29. D'Amelio P, Grimaldi A, Di Bella S, Tamone C, Brianza SZ, Ravazzoli MG, et al. Risedronate reduces osteoclast precursors and cytokine production in postmenopausal osteoporotic women. J Bone Miner Res. (2008) 23:373–9. doi: 10.1359/jbmr.071031
30. Koek WNH, van der Eerden BCJ, Alves R, van Driel M, Schreuders-Koedam M, Zillikens MC, et al. Osteoclastogenic capacity of peripheral blood mononuclear cells is not different between women with and without osteoporosis. Bone. (2017) 95:108–14. doi: 10.1016/j.bone.2016.11.010
31. Tjoa STS, De Vries TJ, Schoenmaker T, Kelder A, Loos BG, Everts V. Formation of osteoclast-like cells from peripheral blood of periodontitis patients occurs without supplementation of M-CSF. J Clin Periodontol. (2008) 35:568575. doi: 10.1111/j.1600-051X.2008.01241.x
32. Herrera BS, Bastos AS, Coimbra LS, Teixeira SA, Rossa C Jr, Van Dyke TE, et al. Peripheral blood mononuclear phagocytes from patients with chronic periodontitis are primed for osteoclast formation. J Periodontol. (2014) 85:e72–81. doi: 10.1902/jop.2013.130280
33. Roato I, Grano M, Brunetti G, Colucci S, Mussa A, Bertetto O, et al. Mechanisms of spontaneous osteoclastogenesis in cancer with bone involvement. FASEB J. (2005) 19:228–30. doi: 10.1096/fj.04-1823fje
34. Roato I, Gorassini E, Brunetti G, Grano M, Ciuffreda L, Mussa A, et al. IL-7 modulates osteoclastogenesis in patients affected by solid tumors. Ann N Y Acad Sci. (2007) 1117:377–84. doi: 10.1196/annals.1402.002
35. Roato I, D'Amelio P, Gorassini E, Grimaldi A, Bonello L, Fiori C, et al. Osteoclasts are active in bone forming metastases of prostate cancer patients. PLoS ONE. (2008) 3:e3627. doi: 10.1371/journal.pone.0003627
36. D'Amico L, Satolli MA, Mecca C, Castiglione A, Ceccarelli M, D'Amelio P, et al. Bone metastases in gastric cancer follow a RANKL-independent mechanism. Oncol Rep. (2013) 29:1453–8. doi: 10.3892/or.2013.2280
37. Olivier BJ, Schoenmaker T, Mebius RE, Everts V, Mulder CJ, van Nieuwkerk KM, et al. Increased osteoclast formation and activity by peripheral blood mononuclear cells in chronic liver disease patients with osteopenia. Hepatology. (2008) 47, 259–67. doi: 10.1002/hep.21971
38. Oostlander A, Everts V, Schoenmaker T, Bravenboer N, van Vliet SJ, van Bodegraven AA, et al. T cell-mediated increased osteoclast formation from peripheral blood as a mechanism for Crohn's disease-associated bone loss. J Cell Biochem. (2012) 113:260–8. doi: 10.1002/jcb.23352
39. Cafiero C, Gigante M, Brunetti G, Simone S, Chaoul N, Oranger A, et al. Inflammation induces osteoclast differentiation from peripheral mononuclear cells in chronic kidney disease patients: crosstalk between the immune and bone systems. Nephrol Dial Transplant. (2018) 33:65–75. doi: 10.1093/ndt/gfx222
40. Faienza MF, Brunetti G, Ventura A, Piacente L, Messina MF, De Luca F, et al. Mechanisms of enhanced osteoclastogenesis in girls and young women with Turner's Syndrome. Bone. (2015) 81:228–36. doi: 10.1016/j.bone.2015.07.021
41. Mucci JM, Cuello MF, Kisinovsky I, Larroude M, Delpino MV, Rozenfeld PA. Proinflammatory and proosteoclastogenic potential of peripheral blood mononuclear cells from Gaucher patients: implication for bone pathology. Blood Cells Mol Dis. (2015) 55:134–43. doi: 10.1016/j.bcmd.2015.05.009
42. Reed M, Baker RJ, Mehta AB, Hughes DA. Enhanced differentiation of osteoclasts from mononuclear precursors in patients with Gaucher disease. Blood Cells Mol Dis. (2013) 51:185–94. doi: 10.1016/j.bcmd.2013.04.006
43. Roato I, Porta F, Mussa A, D'Amico L, Fiore L, Garelli D, et al. Bone impairment in phenylketonuria is characterized by circulating osteoclast precursors and activated T cell increase. PLoS ONE. (2010) 5:e14167. doi: 10.1371/journal.pone.0014167
44. Gravallese EM, Goldring SR. Cellular mechanisms and the role of cytokines in bone erosions in rheumatoid arthritis. Arthritis Rheum. (2000) 43:2143–51. doi: 10.1002/1529-0131(200010)43:10<2143::Aid-anr1>3.0.Co;2-s
45. Gravallese EM, Manning C, Tsay A, Naito A, Pan C, Amento E, et al. Synovial tissue in rheumatoid arthritis is a source of osteoclast differentiation factor. Arthritis Rheum. (2000) 43:250–8. doi: 10.1002/1529-0131(200002)43:2<250::AID-ANR3>3.0.CO;2-P
46. Herman S, Muller RB, Kronke G, Zwerina J, Redlich K, Hueber AJ, et al. Induction of osteoclast-associated receptor, a key osteoclast costimulation molecule, in rheumatoid arthritis. Arthritis Rheum. (2008) 58:3041–50. doi: 10.1002/art.23943
47. Gengenbacher M, Sebald HJ, Villiger PM, Hofstetter W, Seitz M. Infliximab inhibits bone resorption by circulating osteoclast precursor cells in patients with rheumatoid arthritis and ankylosing spondylitis. Ann Rheum Dis. (2008) 67:620–4. doi: 10.1136/ard.2007.076711
48. Vandooren B, Melis L, Veys EM, Tak PP, Baeten D. In vitro spontaneous osteoclastogenesis of human peripheral blood mononuclear cells is not crucially dependent on T lymphocytes. Arthritis Rheum. (2009) 60:1020–5. doi: 10.1002/art.24413
49. Horwood NJ, Kartsogiannis V, Quinn JM, Romas E, Martin TJ, Gillespie MT. Activated T lymphocytes support osteoclast formation in vitro. Biochem Biophys Res Commun. (1999) 265:144–50. doi: 10.1006/bbrc.1999.1623
50. Gravallese EM, Schett G. Effects of the IL-23-IL-17 pathway on bone in spondyloarthritis. Nat Rev Rheumatol. (2018) 14:631–40. doi: 10.1038/s41584-018-0091-8
51. de Vries TJ, Yousovich J, Schoenmaker T, Scheres N, Everts V. Tumor necrosis factor-alpha antagonist infliximab inhibits osteoclast formation of peripheral blood mononuclear cells but does not affect periodontal ligament fibroblast-mediated osteoclast formation. J Periodontal Res. (2016) 51:186–95. doi: 10.1111/jre.12297
52. Xue Y, Jiang L, Cheng Q, Chen H, Yu Y, Lin Y, et al. Adipokines in psoriatic arthritis patients: the correlations with osteoclast precursors and bone erosions. PLoS ONE. (2012) 7:e46740. doi: 10.1371/journal.pone.0046740
53. Mabilleau G, Petrova N, Edmonds ME, Sabokbar A. Number of circulating CD14-positive cells and the serum levels of TNF-α are raised in acute charcot foot. Diabetes Care. (2011) 34:e33. doi: 10.2337/dc10-1695
54. Niida S, Kaku M, Amano H, Yoshida H, Kataoka H, Nishikawa S, et al. Vascular endothelial growth factor can substitute for macrophage colony-stimulating factor in the support of osteoclastic bone resorption. J Exp Med. (1999) 190:293–8. doi: 10.1084/jem.190.2.293
55. Watad A, Bridgewood C, Russell T, Marzo-Ortega H, Cuthbert R, McGonagle D. The early phases of ankylosing spondylitis: emerging insights from clinical and basic science. Front Immunol. (2018) 9:2668. doi: 10.3389/fimmu.2018.02668
56. Tella SH, Gallagher JC. Prevention and treatment of postmenopausal osteoporosis. J Steroid Biochem Mol Biol. (2014) 142:155–70. doi: 10.1016/j.jsbmb.2013.09.008
57. D'Amelio P, Grimaldi A, Di Bella S, Brianza SZ, Cristofaro MA, Tamone C, et al. Estrogen deficiency increases osteoclastogenesis up-regulating T cells activity: a key mechanism in osteoporosis. Bone. (2008) 43:92–100. doi: 10.1016/j.bone.2008.02.017
58. Drake MT, Clarke BL, Lewiecki EM. The pathophysiology and treatment of osteoporosis. Clin Ther. (2015) 37:1837–50. doi: 10.1016/j.clinthera.2015.06.006
59. de Vries TJ, Andreotta S, Loos BG, Nicu EA. Genes critical for developing periodontitis: lessons from mouse models. Front Immunol. (2017) 8:1395. doi: 10.3389/fimmu.2017.01395
60. Loos BG, Papantonopoulos G, Jepsen S, Laine ML. What is the contribution of genetics to periodontal risk? Dent Clin N Am. (2015) 59:761–80. doi: 10.1016/j.cden.2015.06.005
61. Bakri Y, Sarrazin S, Mayer UP, Tillmanns S, Nerlov C, Boned A, et al. Balance of MafB and PU.1 specifies alternative macrophage or dendritic cell fate. Blood. (2005) 105:2707–16. doi: 10.1182/blood-2004-04-1448
62. Nitta K, Yajima A, Tsuchiya K. Management of osteoporosis in chronic kidney disease. Intern Med. (2017) 56:3271–6. doi: 10.2169/internalmedicine.8618-16
63. Horton MA, Taylor ML, Arnett TR, Helfrich MH. Arg-Gly-Asp (RGD) peptides and the anti-vitronectin receptor antibody 23C6 inhibit dentine resorption and cell spreading by osteoclasts. Exp Cell Res. (1991) 195:368–75. doi: 10.1016/0014-4827(91)90386-9
64. D'Amico L, Roato I. Cross-talk between T cells and osteoclasts in bone resorption. Bonekey Rep. (2012) 1:82. doi: 10.1038/bonekey.2012.82
65. Wakkach A, Rouleau M, Blin-Wakkach C. Osteoimmune interactions in inflammatory bowel disease: central role of bone marrow Th17 TNFalpha cells in osteoclastogenesis. Front Immunol. (2015) 6:640. doi: 10.3389/fimmu.2015.00640
66. Bozec A, Zaiss MM. T regulatory cells in bone remodelling. Curr Osteoporos Rep. (2017) 15:121–5. doi: 10.1007/s11914-017-0356-1
67. Ziegler-Heitbrock L, Ancuta P, Crowe S, Dalod M, Grau V, Hart DN, et al. Nomenclature of monocytes and dendritic cells in blood. Blood. (2010) 116:74–80. doi: 10.1182/blood-2010-02-258558
68. Geissmann F, Jung S, Littman DR. Blood monocytes consist of two principal subsets with distinct migratory properties. Immunity. (2003) 19:71–82. doi: 10.1016/S1074-7613(03)00174-2
69. Anbazhagan K, Duroux-Richard I, Jorgensen C, Apparailly F. Transcriptomic network support distinct roles of classical and non-classical monocytes in human. Int Rev Immunol. (2014) 33:470–89. doi: 10.3109/08830185.2014.902453
70. Bolzoni M, Ronchetti D, Storti P, Donofrio G, Marchica V, Costa F, et al. IL21R expressing CD14+CD16+ monocytes expand in multiple myeloma patients leading to increased osteoclasts. Haematologica. (2017) 102:773–84. doi: 10.3324/haematol.2016.153841
71. Chiu YG, Shao T, Feng C, Mensah KA, Thullen M, Schwarz EM, et al. CD16 (FcRgammaIII) as a potential marker of osteoclast precursors in psoriatic arthritis. Arthritis Res Ther. (2010) 12:R14. doi: 10.1186/ar2915
Keywords: peripheral blood, osteoclast formation, T-cells, CD14+CD16+, osteoclast precursor priming, inflammatory bone diseases
Citation: de Vries TJ, el Bakkali I, Kamradt T, Schett G, Jansen IDC and D'Amelio P (2019) What Are the Peripheral Blood Determinants for Increased Osteoclast Formation in the Various Inflammatory Diseases Associated With Bone Loss? Front. Immunol. 10:505. doi: 10.3389/fimmu.2019.00505
Received: 29 November 2018; Accepted: 25 February 2019;
Published: 19 March 2019.
Edited by:
Takayuki Yoshimoto, Tokyo Medical University, JapanReviewed by:
Guillaume Mabilleau, Université d'Angers, FranceFrancesca Salamanna, Rizzoli Orthopedic Institute, Italy
Copyright © 2019 de Vries, el Bakkali, Kamradt, Schett, Jansen and D'Amelio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Teun J. de Vries, teun.devries@acta.nl