 María-José Ferrándiz
María-José Ferrándiz David Carreño1†
David Carreño1† Silvia Ayora
Silvia Ayora Adela G. de la Campa
Adela G. de la Campa- 1Unidad de Genética Bacteriana, Centro Nacional de Microbiología, Instituto de Salud Carlos III, Madrid, Spain
- 2Departamento de Biotecnología Microbiana, Centro Nacional de Biotecnología, Consejo Superior de Investigaciones Científicas, Madrid, Spain
- 3Presidencia, Consejo Superior de Investigaciones Científicas, Madrid, Spain
The histone-like protein HU is a conserved nucleoid-associated protein that is involved in the maintenance of the bacterial chromosome architecture. It is the only known nucleoid-associated protein in Streptococcus pneumoniae, but it has not been studied. The pneumococcal gene encoding this protein, hlp, is shown herein to be essential for cell viability. Its disruption was only possible either when it was duplicated in the chromosome and its expression induced from the PZn promoter, or when hlp was cloned into a plasmid under the control of the inducible Pmal promoter. In vitro assays indicated that pneumococcal HU shows a preference for binding to supercoiled DNA rather than to linear or nicked DNA. In vivo experiments in which the amount of HU was manipulated showed a relationship between the amount of HU and the level of DNA supercoiling. A twofold reduction in the amount of HU triggered a 21% increase in DNA relaxation in untreated cells. However, in cells treated with novobiocin, a drug that relaxes DNA by inhibiting DNA gyrase, a 35% increase in DNA relaxation was observed, instead of the expected 20% in cells with a constitutive HU amount. Conversely, a fourfold HU increase caused only 14% of DNA relaxation in the presence of novobiocin. Taken together, these results support an essential role for HU in the maintenance of DNA supercoiling in S. pneumoniae.
Introduction
Streptococcus pneumoniae (the pneumococcus) is the causative agent of respiratory tract infections and invasive serious illness including meningitis and bacteremia (Lynch and Zhanel, 2009). More than 1.6 million people die of pneumococcal infection every year and half of these deaths are in children aged less than 5 years according to World Health Organization estimations (World Health Organization, 2007).
As in other bacteria, the chromosome of S. pneumoniae is confined within the nucleoid. Chromosome compaction is achieved by the action of several factors, including DNA supercoiling, nucleoid-associated proteins (NAPs) (Pato and Banerjee, 1996) and macromolecular crowding (Wang et al., 2013). In bacteria, DNA is usually negatively supercoiled and it is mainly found in a plectonemic form (Travers and Muskhelishvili, 2007). This plectoneme formation leads to a 10% reduction in the radius of gyration of DNA, which is a measure of the volume (Luijsterburg et al., 2008). In addition, negative supercoiling eases the association of architectural proteins (de los Rios and Perona, 2007; Luijsterburg et al., 2008), which also affects the volume that a DNA molecule occupies in the cell. The DNA supercoiling balance results from the joint action of DNA topoisomerases with opposing activities (Zechiedrich et al., 2000). Topoisomerase I and topoisomerase IV relax negatively supercoiled DNA, whereas DNA gyrase introduces negative supercoiling. The alteration of this equilibrium leads to a cellular response to restore DNA superhelicity (Menzel and Gellert, 1983; Tse-Dinh, 1985; Ferrándiz et al., 2010). The large concentration of macromolecules found in the bacterial cytoplasm (>0.3 g of RNA and protein/ml in Escherichia coli) (Zimmerman and Trach, 1991) promotes the compaction of the DNA directly (Zimmerman, 1993) or indirectly by increasing the binding of proteins (Murphy and Zimmerman, 1995). In addition to these factors, various polycationic species neutralize DNA charges that attach DNA (de Vries, 2010).
The architectural properties of NAPs are derived from their capacity to bind DNA, wrapping it, bending the double helix or forming bridges among separate DNA segments (Luijsterburg et al., 2006; de los Rios and Perona, 2007; Luijsterburg et al., 2008). In eukaryotes the main mechanism of packaging DNA is wrapping it via the action of histones. Several NAPs have been described in bacteria and play an important role in the organization of chromatin (Luijsterburg et al., 2006). Members of the Lrp/AsnC family form octameric structures that wrap DNA around themselves in a right-handed superhelix to promote DNA compaction (Thaw et al., 2006; de los Rios and Perona, 2007). DNA bridging favored by NAPs leads to formation of complexes between DNA duplexes or between DNA and other architectural proteins resulting in the formation of loops. The protein archetypes of this group are H-NS and the H-NS-like proteins found in Gram-negative proteobacteria (Bertin et al., 1999; Dame et al., 2000). The proteins that bend DNA are widely distributed among prokaryotes, and the most well-known are those in the HU/IHF family (Swinger and Rice, 2004).
The architectural function of NAPs is exerted through their capacity to non-specifically bind DNA. H-NS, HU, and Fis have a preference for A+T-rich DNA, and H-NS has a greater affinity for curved DNA (reviewed by Dorman, 2014). In addition to their architectural role, NAPs have an important function in the regulation of gene expression. The preference for A+T-rich DNA allows these proteins to target horizontally acquired elements (with higher-than-average A+T content) including pathogenicity islands encoding virulence factors (reviewed by Dorman, 2014). NAPs and DNA supercoiling influences the transcription of many bacterial genes in a co-operative way, as the topological state of the DNA target is important for the binding of NAPs (Dorman, 2013). The same phenomenon occurs for H-NS and Fis, which modulate the topological response of pel genes (major virulence factors) in the plant pathogen Dickeya dadantii (Ouafa et al., 2012), and of Fis in the control of virulence genes in Salmonella (Croinin et al., 2006).
By bending or bridging DNA, NAPs facilitate cellular processes. HU, IHF, and Fis are involved in DNA replication (Chodavarapu et al., 2008; Kasho et al., 2014); HU participates in recombination and DNA repair (Kamashev and Rouviere-Yaniv, 2000); and H-NS, IHF, and HU act as transcriptional regulators (Dillon and Dorman, 2010; Berger et al., 2010; Kahramanoglou et al., 2011).
HU (also called HlpA, Hlp or Hup) is a small, basic, and highly conserved protein in the prokaryotic kingdom (Drlica and Rouvière-Yaniv, 1987; Azam and Ishihama, 1999) and can also be found in chloroplasts (Briat et al., 1984) and in yeast mitochondria (Caron et al., 1979). HU forms homo- or heterodimers that bind to double-stranded DNA non-specifically and with low affinity and prefers to bind intrinsically flexible DNA (Tanaka et al., 1993). A role in transcriptional regulation has been shown for this protein in E. coli, modulating genes that respond to anaerobiosis, acid stress, high osmolarity and SOS induction (Oberto et al., 2009). It also regulates the spatial distribution of RNA polymerase in the nucleoid, implicating a role for HU in coordinating the genomic structure and transcription (Berger et al., 2010). A role in virulence has been attributed to this protein when released into tissues during infection by Streptococcus pyogenes and other streptococcal species (Bergey and Stinson, 1988; Choi and Stinson, 1989, 1991; Winters et al., 1993). This virulence may be explained by the release of HU by these streptococci during the stationary phase due to the autolysis process (Stinson et al., 1998) triggering a cascade of events that induce pro-inflammatory responses that contribute to the activation of host innate immunity during bacterial infection (Liu et al., 2008a). In S. pneumoniae, the roles of NAPs in the chromosome architecture are unknown, and as in other streptococcal species, important proteins involved in the organization of the nucleoid, such as H-NS, IHF, or Fis, are absent. These observations suggest that HU is a key protein in the organization and compaction of the chromosome in S. pneumoniae. In this study, we characterized the pneumococcal HU protein (SpnHU), which is essential for the cell viability of S. pneumoniae due to its role in preserving DNA supercoiling.
Materials and Methods
Bacterial Strains, Growth and Transformation
Streptococcus pneumoniae R6 strain, which was used in all experiments, was grown in a casein hydrolysate-based medium (AGCH) supplemented with 0.2% yeast extract and 0.3–0.8% sucrose or 0.8% maltose as a carbon source. Transformation was performed in R6 strain with chromosomal or plasmid DNA as previously reported (Lacks et al., 1986). Transformants were selected in medium containing 1 μg/ml tetracycline for plasmids pMV158 and pMVHU (Lacks et al., 1986), and 2.5 μg/ml chloramphenicol or 250 μg/ml kanamycin for chromosomal insertions. To induce DNA relaxation, 1 μg/ml (1 × MIC) of novobiocin (NOV) was added to cultures. Strains containing plasmids pMV158, pMVHU, and pLS1 were grown in medium containing 1 μg/ml tetracycline. Growth was followed by measuring the optical density at 620 nm (OD620 nm) either in an UV-visible spectrophotometer (Evolution 201, Thermo Scientific) or in a microplate reader (Infinite F200, Tecan). Measurements of the two devices correlated linearly by means of the equation y = 0.2163 x + 0.1151 (y = microplate reader measure, x = spectrophotometer), with an R2 of 0.98.
DNA Manipulation and Constructs
Chromosomal DNA and plasmids from S. pneumoniae were obtained as described previously (Fenoll et al., 1994). Restriction endonucleases and DNA ligase (Fermentas) were used following the supplier’s specifications. PCR was performed using 1 U of PfuI (Fermentas). Conditions for PCR were as follows: an initial cycle of 30 s denaturation at 94°C, and 30 cycles of denaturation at 94°C for 15 s, annealing at 50°C for 30 s and extension at 68°C for 1 min per kb of PCR product.
The hlp gene (spr1020) was inactivated in the chromosome by allelic replacement by homologous recombination. Three DNA fragments, obtained by PCR amplification, were digested with the appropriate restriction enzymes and ligated as follows. Two fragments upstream and downstream of the hlp gene of 1238 and 1151 bp were amplified with primers pairs 101829R/HU15RPAE (with the PaeI restriction site) and HU73FXBA/1021208F (with the XbaI restriction site), respectively (Table 1), using DNA from strain R6 as template. A third DNA fragment of 929 bp bearing the cat gene was amplified from plasmid pJS3 with primers CATUP1PAE and CATDOWN1XBA, which contained restriction sites for PaeI and XbaI, respectively (Table 1). The three fragments were digested and ligated, and the ligation product was used as a template to obtain a 3604 bp PCR product (oligonucleotide pair 101829R/1021208F), which was used to transform R6 competent cells. Transformants were selected by plating in AGCH-agar medium supplemented with 0.2% yeast extract and 0.3% sucrose and containing 2.5 μg/ml chloramphenicol. To confirm the disruption, amplification from the chromosome was performed with primers 1018131R and 1021339F (Table 1) flanking the replaced DNA. Primers UPHU1, CATMED, and CAT191 (Table 1) were used to sequence the construct.

TABLE 1. List of primers used in this work.
Plasmid pMVHU, which contains the hlp gene under the control of the maltose- inducible promoter Pmal, was constructed as follows. First, plasmid pMV158GFP was digested with NdeI, treated with Klenow to fill in recessed 3′ ends and obtained blunted 5′ overhang, and digested again with SphI, rendering a fragment of 6186 bp without the gfp gene. This fragment was ligated to a 314-bp DNA obtained by amplification of R6 DNA with primers HURTF that had been previously phosphorylated and HUSPH (Table 1), which included a SphI restriction site, and digested with this enzyme. This same DNA fragment of 314 bp digested with SphI was also cloned into plasmid TAGZyme pQE-1 from Qiagen and, digested with PvuII and SphI to obtain six histidine (H6) codons fused to the 5′ end of hlp. The new plasmid was named pQEHU.
To clone hlp under the control of the Zn-inducible promoter PZn, this copy was positioned in the chromosome of pneumococcal R6 strain at the dispensable spr1866 locus (Martín-Galiano et al., 2014) as follows. The gene was amplified with HUATGSAL and HUTAASAC primers (Table 1) containing SalI and SacI restriction sites, respectively. This PCR product was digested with SalI and SacI and ligated into plasmid pZ0479 (Martín-Galiano et al., 2014). The obtained plasmid, pZ0479HU, was used as a DNA template to amplify hlp under PZn together with a kanamycin resistance cassette (Figure 2) using the oligonucleotide pairs KmR_B1_R/ pZK_Xb1_R (Table 1), and attached to regions flanking the disposable spr1866 gene (Martín-Galiano et al., 2014). This construct was introduced into the S. pneumoniae R6 chromosome by genetic transformation. Successful integration was checked by PCR using primers 1865EXF and 1865EXR (Table 1) flanking the replaced DNA. Primers HUATGSAL and HUTAASAC (Table 1) were used to sequence the construct.
Southern Blot Analysis
Chromosomal DNA (3 μg) from R6, R6 PZn-hlp, and R6 PZn-hlpΔhlp was digested with HindIII and separated by 0.8% agarose gel electrophoresis, transferred to Nylon membranes and hybridized to a biotinylated 141-bp hlp probe obtained by PCR amplification with 5′ biotinylated oligonucleotides HU20BIOT and HU61RBIOT (Table 1). Blots were developed with the Phototope-Star Detection Kit (New England Biolabs) following the manufacturer’s instructions.
SpnHU Purification
For overexpression of hlp fused to H6 codons at its 5′ end, E. coli XL1 (pQEHU) was grown to an OD600 nm of 0.5–0.6, and expression was induced for 2 h with 1 mM IPTG. Cells from 2 l cultures were harvested, suspended in lysis buffer (50 mM phosphate buffer pH 8.0, 300 mM NaCl, 10 mM imidazole), and chilled on ice for 1 h with 1 mg/ml lysozyme and 0.25% Triton X-100. Lysis was completed by five cycles of 20 s of sonication with an amplitude of 30% in a Sonifier B-12 (Branson Co, Danbury, CT, United States). Cell debris was pelleted by centrifugation at 20000 ×g for 10 min, and the resulting supernatant was filtered through a 0.45 μm Millipore Millex® HA filter. This supernatant was loaded into a Ni-NTA (QIAGEN) column in an ÄKTA FPLC system following the manufacturer’s instructions. (H6)SpnHU was eluted with 20 ml of buffer containing a 200–350 mM gradient of imidazole. Fractions of 1 ml were collected and analyzed by SDS-12% polyacrylamide gel electrophoresis and stained with Coomassie blue. Fractions containing a protein of the expected size were dialyzed against 20 mM NaH2PO4, 150 mM NaCl, pH 7.0. To complete removal of the H6-tag and additional amino acids from the amino terminus, (H6)SpnHU was treated with the TAGZyme System (Qiagen) following the manufacturer’s instructions. This system consists of dipeptidyl aminopeptidase I (DPase I) in combination with glutamine cyclotransferase (Qcyclase) and pyroglutamyl aminopeptidase (pGAPase). The three enzymes contain a C-terminal His tag that allows their removal from the reaction solution by additional Ni-Affinity chromatography steps. The purified SpnHU was dialyzed in 10 mM TrisHCl pH 8.0, 50 mM KCl, 0.5 mM DTT, 0.5 mM EDTA and 50% glycerol and stored at -20°C. The protein concentration was measured using a colorimetric protein assay (Bio-Rad) following the manufacturer’s instructions. The concentration of SpnHU is expressed in monomeric form.
Cross-Linking of SpnHU
SpnHU was incubated at room temperature for 30 min in 0.1% glutaraldehyde in a total volume of 10 μl containing 5 mM Tris-HCl, pH 8.0, 25 mM KCl, 25% glycerol, 0.25 mM EDTA, 0.25 mM DTT. Samples were diluted with an equal volume of Laemmli sample buffer, loaded into an SDS-polyacrylamide gel (15%), electrophoresed, and stained with Coomassie blue.
Electrophoretic Mobility-Shift Assay (EMSA)
Plasmid pBR322 DNA (2 nM) that was supercoiled, PstI-linearized, or nicked with Nt. BstNBI (New England Biolabs) was incubated at room temperature for 20 min with different amounts of purified SpnHU in 15 μl of 10 mM Tris-HCl, pH 8.0, 50 mM KCl, 0.05% Brij 58, 0.01 mg/ml BSA, 5% glycerol, 0.01 mM EDTA and 0.01 mM DTT. Samples were loaded in a 0.5% agarose gel and electrophoresed in Tris-borate-EDTA at 18 V for 14 h at room temperature. After electrophoresis, the gels were stained with a 0.5 μg/ml ethidium bromide solution.
Supercoiling Assays
Supercoiled plasmid pBR322 (0.18 nM) was relaxed with 5.3 nM topoisomerase I of S. pneumoniae obtained in our laboratory (García et al., 2011) in a final volume of 190 μl by incubation at 37°C for 30 min in 20 mM TrisHCl pH 8.0, 100 mM KCl, 10 mM MgCl2, 1 mM DTT, 50 μg/ml BSA. Increasing concentrations of SpnHU were added to the mix to reach a final volume of 200 μl. Reactions were incubated for 1 h at 37°C and stopped by addition of 50 mM EDTA, 1% SDS and 100 μg/ml proteinase K, followed by incubation at 37°C for 1 h. DNA was precipitated and suspended in 1 × loading buffer, electrophoresed on a 1.2% agarose gel at 18 V for 18 h, and stained with 1 μg/ml ethidium bromide.
Two-Dimensional Agarose Gel Electrophoresis
The distribution of topoisomers of plasmid pMVHU was analyzed in neutral/neutral two-dimensional agarose gels. Plasmid DNA was obtained as previously described (Martín-Parras et al., 1998). The first dimension was run in a 0.4% (w/v) agarose (Seakem; FMC Bioproducts) gel in Tris-borate-EDTA containing 1 μg/ml of chloroquine (Sigma) at 1.5 V/cm at room temperature for 19 h. The second dimension was performed in 1% agarose gel in the same buffer containing 2 μg/ml of chloroquine at 7.5 V/cm for 7–9 h at 4°C. Chloroquine was added to both, the agarose and the running buffer. After electrophoresis, gels were subjected to Southern hybridization using as the probe a 240-bp PCR fragment obtained from pMVHU DNA with primers REPUPBIOT (5′ biotinylated) and REPDOWN (Ferrándiz et al., 2010). Chemiluminescent detection of DNA was performed with the Phototope®-Star kit (New England Biolabs). Images were captured in a VersaDoc MP400 system (BioRad) and analyzed with the Quantity One program. The DNA linking number (Lk) was calculated by quantifying the amount of every given topoisomer. The DNA supercoiling density (σ) was calculated with σ = ΔLk/Lk0. Linking number differences (ΔLk) were determined with the equation ΔLk = Lk - Lk0, in which Lk0 = N/10.5, where N is the DNA size in bp and 10.5 is the number of bp per complete turn of B-DNA, the most probable helical repeat of DNA under the used conditions.
RNA Extraction and Real Time qRT-PCR Experiments
RNA was extracted from 2.5 to 3 × 1010 cells with the RNeasy kit (Qiagen). cDNAs were synthesized from 5 μg of RNA with SuperScript™ III Reverse Transcriptase (Invitrogen) for 1 h at 55°C. These cDNAs were subjected to quantitative qRT-PCR (Chromo 4, Bio-Rad) as described previously (Ferrándiz and de la Campa, 2014). The oligonucleotide pairs used are indicated in Table 1. To normalize the three independent qRT-PCR values, they were divided by those obtained by amplification of an internal fragment of 16S rDNA (Table 1).
Western Blot Analysis
Rabbit polyclonal antibodies against SpnHU were obtained after three subcutaneous injections at 3-week intervals of polyacrylamide portions containing (H6)SpnHU. Blood was recovered and the serum was stored at -80°C. Polyclonal antibodies against LytA were kindly provided by Ernesto García (CIB, CSIC, Madrid, Spain). Whole cell lysates were obtained by centrifugation of approximately ≈ 5 × 1010 cells, suspended in 400 μl of phosphate buffered saline and sonicated 3 × 20 s with a Vibra Cell 75043 (Bioblock Scientific). Lysates were separated on 4–20% Mini-Protean™ TGX gels (Bio-Rad) and transferred to PVDF membranes. Membranes with transferred proteins were probed with anti-SpnHU (diluted 1:100) and anti-LytA (diluted 1:20000) antibodies. Super Signal West Pico chemiluminescent substrate (Thermo Scientific) was used to develop the membranes and monitored with a ChemiDoc™ MP system (Bio-Rad). Image analysis was performed with Image Lab™ software (Bio-Rad).
Measurement of the Intracellular Amount of SpnHU
To calculate the number of SpnHU molecules per cell, Western blotting was performed using anti-SpnHU antibodies as previously described (Ali Azam et al., 1999). Briefly, quantities between 1 and 0.062 μg of purified SpnHU, and lysates of R6 cells from 5.9 × 107 ± 0.71 × 107 colonies-forming units (CFUs) were loaded, electrophoresed in 4–20% gels, and transferred to PVDF membranes. Blots were developed and scanned to quantify the band intensities as indicated above. The linear correlation between the amount of SpnHU and the immunostaining intensity served to interpolate the amount of protein from the cell lysate. CFUs were calculated by plating cell extracts on blood agar plates. The molecular mass of SpnHU is 9.6 kDa. Determination was performed with three independent cell extracts.
Statistical Analysis
GraphPad Prism 7.02 was used for the statistical analysis. The SpnHU level among ΔHU (pMVHU) cells grown either with sucrose or maltose and R6 was determined by one-way analysis of variance. Supercoiling density values (σ) were determined using two-way analysis of variance with Tukey’s multiple comparisons test. The 95% confidence interval was used. Statistical significance was defined as P < 0.05.
Results
HU from Streptococcus pneumoniae R6 Is Highly Conserved Among Pneumococcal Strains and Streptococci of the Mitis Group
Analysis of the genomic sequence of S. pneumoniae, revealed a single gene encoding SpnHU, suggesting that the protein is a homodimer, as in other Gram-positive bacteria (Tanaka et al., 1984; Stinson et al., 1998; Liu et al., 2008b). SpnHU from R6 shares 59 and 55% of identity with Escherichia coli HU subunits alpha and beta, respectively. The identity increased to 75.3 and 89% when compared with HU from Gram-positive bacteria such as Staphylococcus aureus (SauHU) and Streptococcus mutans (SmuHU), respectively. When compared with HU of streptococcal species of the mitis group, the identity increased to 95.6%. Its predicted secondary structure included three α-helices and five β-sheets following the α1-α2-β1-β2-β3-β4-β5-α3 organization (Figure 1A). This secondary structure is highly conserved among HU proteins and is divided in three domains: helix-turn-helix (HTH), dimerization signal (DS) and DNA-binding domain (DBD) (Christodoulou and Vorgias, 2002; Christodoulou et al., 2003). The predicted structure of the SpnHU homodimer based on SauHU and SmuHU crystal structures is shown in Figure 1B. This structure in SauHU and SmuHU has a V-like form that is transversally divided in two parts: an α-helical body and two protruding β-ribbon arms, which have a flexible nature and clamp DNA along the minor groove (Kim et al., 2014; O’Neil et al., 2016).
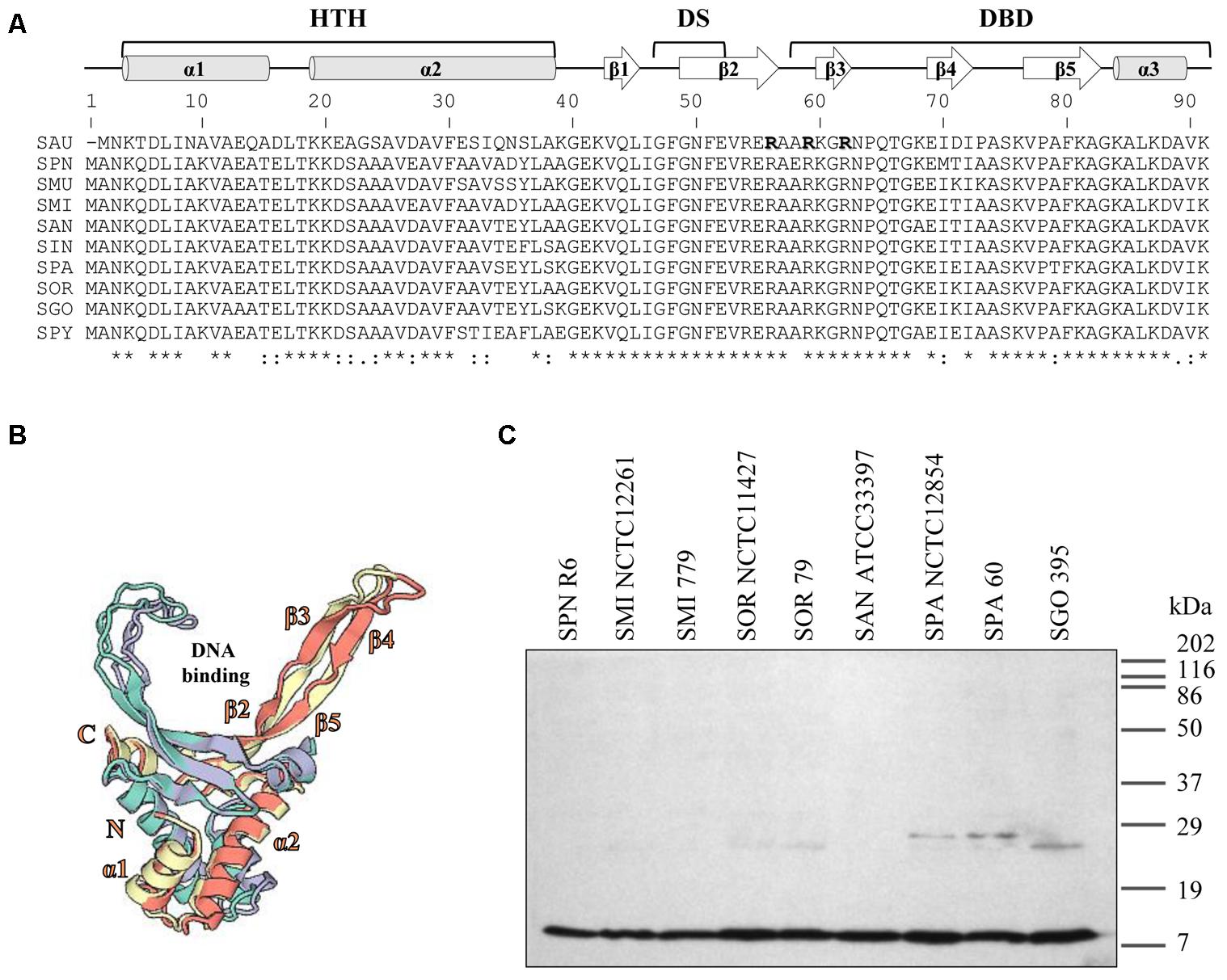
FIGURE 1. Streptococcal HU homologs are highly conserved. (A) Multiple sequence alignment of streptococcal HU homologs and Staphylococcus aureus (SAU) HU protein (SauHU). Above the alignment, secondary structural elements are shown. The secondary structure was predicted based on the position specific-scoring matrices generated by PSI-BLAST with the Pyre2 portal for protein modeling, prediction, and analysis (Kelley et al., 2015). HTH, helix-turn-helix; DS, dimerization signal; and DBD, DNA-binding domain. Residues that are crucial for DNA recognition and binding are denoted in bold face and shadowed. Below the alignment, conserved residues are identified by asterisks. SPN (Streptococcus pneumoniae), SMU (Streptococcus mutans), SMI (Streptococcus mitis), SAN (Streptococcus anginosus), SIN (Streptococcus intermedius), SOR (Streptococcus oralis), SGO (Streptococcus gordoni), SPA (Streptococcus parasanguinis), and SPY (Streptococcus pyogenes). (B) The 3D model of the SpnHU homodimer based on the crystal structures of SauHU (4qjn, homodimer subunits in green and purple) and SMU (5fbm, homodimer subunits in yellow and pink). This structure has been modeled with coverages of 98% (SAU) and 100% (SMU) of the SpnHU sequence using Swiss-Model (Biasini et al., 2014). (C) Western blot analysis of HU protein in different streptococcal species. Crude cell extracts (20 μg) were separated by SDS-PAGE, blotted, and incubated with an antibody directed against SpnHU protein.
Western blot analysis using a polyclonal antibody directed against SpnHU revealed a protein with the same size as SpnHU (9.6 kDa) in the different streptococcal extracts (Figure 1C), highlighting the high identity among streptococcal HU proteins.
The Biochemical Behavior of SpnHU Is Similar to That of Other HU Proteins
To assess the in vitro activity of SpnHU, the protein was purified to approximately 98% homogeneity (Figure 2A) after removal of the H6-tag, as described in Section “Materials and Methods.” In solution, the protein formed dimers at all tested concentrations (Figure 2B), consistent with the results obtained for other HU proteins (Christodoulou and Vorgias, 2002; Chen et al., 2004; Ghosh and Grove, 2004) and with the cocrystal structures of dimeric Anabaena HU bound to DNA (Swinger et al., 2003). At protein concentrations higher than 10 μM, multimeric species, including trimers, were observed, and some of them failed to enter the gel (Figure 2B). The formation of oligomeric structures in vitro has also been observed for other HUs and can be explained as the likely association of free monomers, either with themselves or with assembled dimers, resulting in non-specific aggregation (Ghosh and Grove, 2004; Kamau et al., 2005).
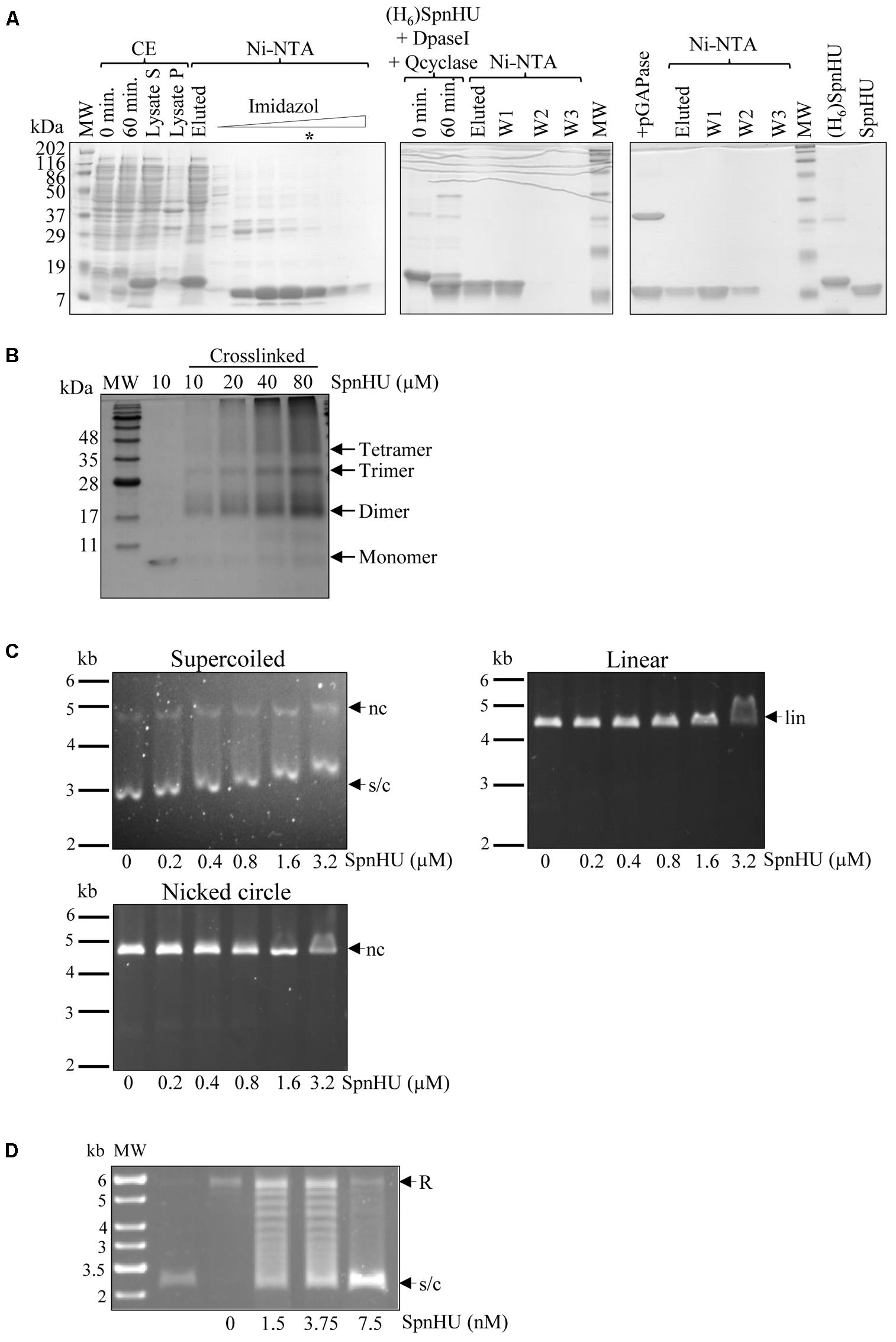
FIGURE 2. In vitro characterization of SpnHU activity. (A) Purification of recombinant [(H6)SpnHU] and native (SpnHU) proteins. A culture of E. coli XL1blue (pQEHU) was grown and induced with IPTG as described in Section “Materials and Methods.” Samples were electrophoresed in SDS-12% polyacrylamide gels and polypeptides stained with Coomassie Blue. Left image: expression of (H6)SpnHU and purification steps. CE, crude extract (20 μg per lane); 0 and 60 min, samples after IPTG induction; lysate S and lysate P, supernatant and pellet after lysis and centrifugation. Fractions (15 μl) of a Ni-NTA column in which an imidazole gradient was applied. The lane marked with an asterisk contains the (H6)SpnHU that was used for further purification. Middle image: DpaseI and Qcyclase treatment followed by Ni-NTA purification. Right image: pGAPase treatment followed by Ni-NTA purification and 3 μg of purified proteins (H6)SpnHU and SpnHU. MW, molecular weight markers. (B) Oligomerization status of SpnHU. The indicated amounts of crosslinked native protein were separated by SDS-PAGE, and the gels were stained with Coomassie. (C) Agarose gel-based EMSAs showing the binding of increasing concentrations of SpnHU to 0.03 pmol supercoiled (s/c), linear (lin) or nicked (nc) plasmid pBR322. (D) Agarose gel electrophoresis showing the supercoiling s/c of relaxed (R) pBR322 plasmid after incubation with pneumococcal topoisomerase I in the presence of the indicated amounts of SpnHU (second and consecutive lanes). The first lane shows the DNA molecular weight marker with sizes indicated in kb, and the second lane shows supercoiled pBR322 before relaxation with topoisomerase I.
The affinity of SpnHU for supercoiled, linear, or nicked DNA was tested by gel-shift assays using 2 nM plasmid pBR322. While shifts in mobility were observed with a concentration of 0.2 μM SpnHU when supercoiled pBR322 was used, eight-fold and sixteen-fold more SpnHU (1.6 μM and 3.2 μM) was required to shift 2 nM linear or nicked pBR322, respectively (Figure 2C). This preference for supercoiled DNA has been previously described for other bacterial HU proteins (Chen et al., 2004; Kamau et al., 2005; Mukherjee et al., 2008). SpnHU is also able to efficiently constrain pBR322 relaxed with S. pneumoniae topoisomerase I (García et al., 2011), at concentrations lower than 5 nM (Figure 2D).
SpnHU Is Essential for Cell Viability
Previous studies of other bacteria have shown that most homodimeric HU proteins are essential (Micka and Marahiel, 1992; Liu et al., 2008b), although there are exceptions such as HU of Mycobacterium smegmatis, which can be deleted (Hołówka et al., 2017). However, heterodimeric proteins are not essential (Wada et al., 1988). In S. pneumoniae, several studies have been conducted to identify essential genes through the use of different high-throughput gene disruption systems and showed discrepancies regarding the essentiality of hlp (Thanassi et al., 2002; Song et al., 2005; van Opijnen et al., 2009; van Opijnen and Camilli, 2012; Mobegi et al., 2014; Verhagen et al., 2014). To discern whether SpnHU is essential for cell viability, we attempted to knockout the hlp gene by replacing it with a chloramphenicol resistance cassette (cat). A fragment of 3.3 kb containing the hlp flanking genes spr1019 and spr1021 ligated to cat was constructed (Figure 3A) and introduced into the R6 strain by genetic transformation. The frequency of appearance of chloramphenicol-resistant (CmR) colonies was approximately 2 × 104-fold lower (Table 2) than that obtained either by a similar construction generated to replace the dispensable spxB gene by cat (Ferrándiz and de la Campa, 2014), or by transforming plasmid pJS3 containing cat (Ballester et al., 1986). To analyze whether the cat gene had been properly integrated, eight CmR colonies were analyzed by PCR using primers bordering spr1019 and spr1021 (Table 1). All colonies amplified a fragment of 3.2 kb, consistent with the size of wild-type hlp (data not shown). The chloramphenicol resistance of these colonies could be explained by integration of the cassette in other chromosomal loci sharing low homology with homologous arms present in the cassette (the spr1019 and/ or spr1020 regions), which would explain the low frequency of transformation. These data suggest that SpnHU is an essential protein. To confirm this finding, the hlp gene was either cloned in a plasmid, rendering strain R6 (pMVHU) (Figure 3B), or duplicated in the chromosome, rendering strain R6 PZn-hlp (Figure 3C). In both constructs, hlp was cloned under the control of an inducible promoter, such as Pmal, in plasmid pMVHU, which is activated in the presence of maltose (Nieto et al., 2001), and PZn in strain R6 PZn-hlp, which is induced by addition of ZnSO4 to the culture (Kloosterman et al., 2007). Strain R6 PZn-hlp contained an additional copy of hlp ectopically integrated into the spr1865 locus (Figure 3C). The introduction of ectopic DNA in this locus did not affect the cell viability (data not shown). Both strains, R6 (pMVHU) and R6 PZn-hlp, were transformed with the 3.3-kb spr1119-cat-spr1021cassette (Figure 3A), and colonies were selected under conditions that allowed expression of their ectopic hlp. In both cases, CmR transformants were obtained with transformation frequencies of 2.50% and 1.37% in R6 (pMVHU) and R6 PZn-hlp receptor strains, respectively (Table 2). These frequencies were in agreement with those obtained with controls: 2.54% for plasmid pJS3 and 1.89% for the 4.5-kb fragment containing spxB-cat (Table 2). Ten CmR colonies from these transformations of R6 (pMVHU) and R6 PZn-hlp were analyzed by PCR using primers bordering spr1019 and spr1021 (Table 1). All analyzed colonies amplified a 3.9 kb fragment, consistent with the hlp replaced with cat. The sequence of chromosomal DNA confirmed the deletion of hlp. Additionally, strain R6 PZn-hlpΔhlp obtained from R6 PZn-hlp by deletion of hlp (Figure 3C) was also checked by Southern blot analysis (Figure 3D). All these data support that SpnHU is essential for growth.
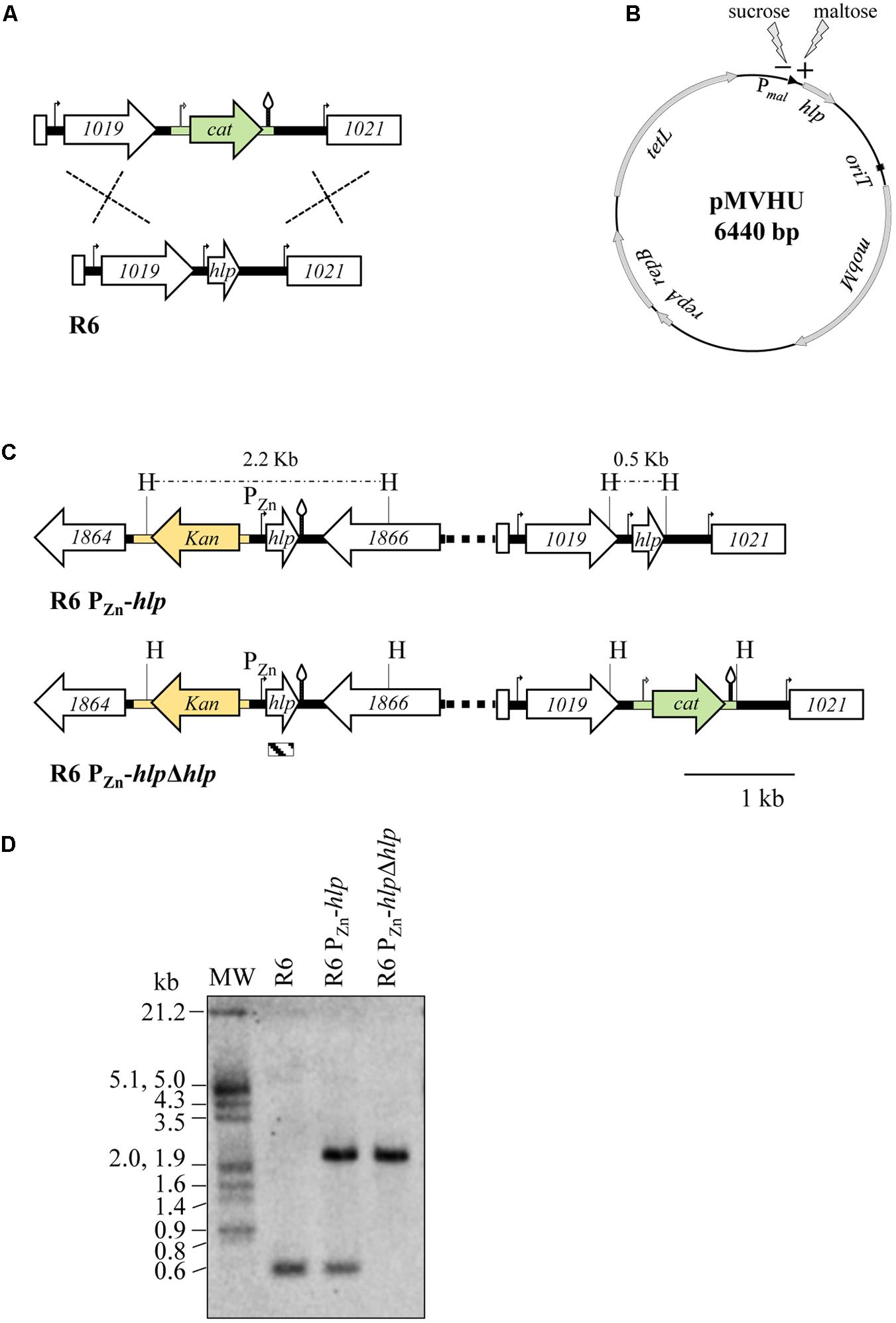
FIGURE 3. Strategies to knockout the hlp gene in S. pneumoniae. (A) Schematic showing the replacement of the chromosomal hlp gene with the chloramphenicol resistance cassette (green drawing). R6 genes (spr numbering) are indicated by white arrows. Promoters are shown as curved arrows and transcription terminators as stem and loop structures. (B) Representation of plasmid pMVHU containing the hlp gene under the control of the maltose promoter. (C) Schematic showing the in cis duplication of the hlp gene in strain R6 PZn-hlp, by placing an additional copy of the gene under the control of a Zn-inducible promoter. The kanamycin resistance cassette is presented in yellow. The organization of strain R6 PZn-hlpΔhlp is also represented. The chloramphenicol resistance cassette, promoters, and transcription terminators are represented as in (A), H, HindIII targets and predicted sizes of fragments. Distances between HindIII targets are shown above the dashed lines. The striped box at the bottom of the figure represents the hlp probe used for Southern analysis and its position inside the hlp gene. (D) Southern blot analysis of the R6, R6 PZn-hlp, and R6 PZn-hlpΔhlp strains. Chromosomal DNA from the strains was cut with HindIII, separated by agarose gel electrophoresis, transferred to a nylon membrane and hybridized with the biotinylated hlp probe shown in (C).

TABLE 2. Attempts to disrupt hlp in the chromosome of R6 or R6 complemented with hlp.
The Levels of SpnHU Affect the in Vivo Level of DNA Supercoiling
To study the influence of SpnHU on DNA supercoiling in vivo, we used strain ΔHU (pMVHU). This strain, constructed from R6, contained the chromosomal copy of hlp replaced with cat and an ectopic copy of the gene cloned under the control of Pmal in plasmid pMVHU (Figures 3A,B). As Pmal is inducible by maltose and repressed in the presence of sucrose (Nieto et al., 2001), strain ΔHU (pMVHU) would produce SpnHU, depending on the presence of sucrose or maltose as the carbon source in the growing medium. We grew ΔHU (pMVHU) and its parental strain R6 as a control, either in medium with maltose or sucrose. No remarkable differences in growth were observed among the strains in any of the growth media used (Figure 4A). We measured the amount of SpnHU under both conditions in the exponential phase of growth by Western blot analysis. As expected, the amount of SpnHU in R6 did not change in any of the media used (Figure 4B). However, for ΔHU (pMVHU), the amount of SpnHU increased significantly by 3.9-fold (P < 0.0001) under growth induction conditions (maltose, M) with respect to the values detected under repression conditions (sucrose, S) (Figure 4B). Although Pmal was not completely silenced under suppression conditions, the amount of SpnHU decreased significantly by twofold (P = 0.0052) compared with R6 grown under the same conditions (Figure 4B).
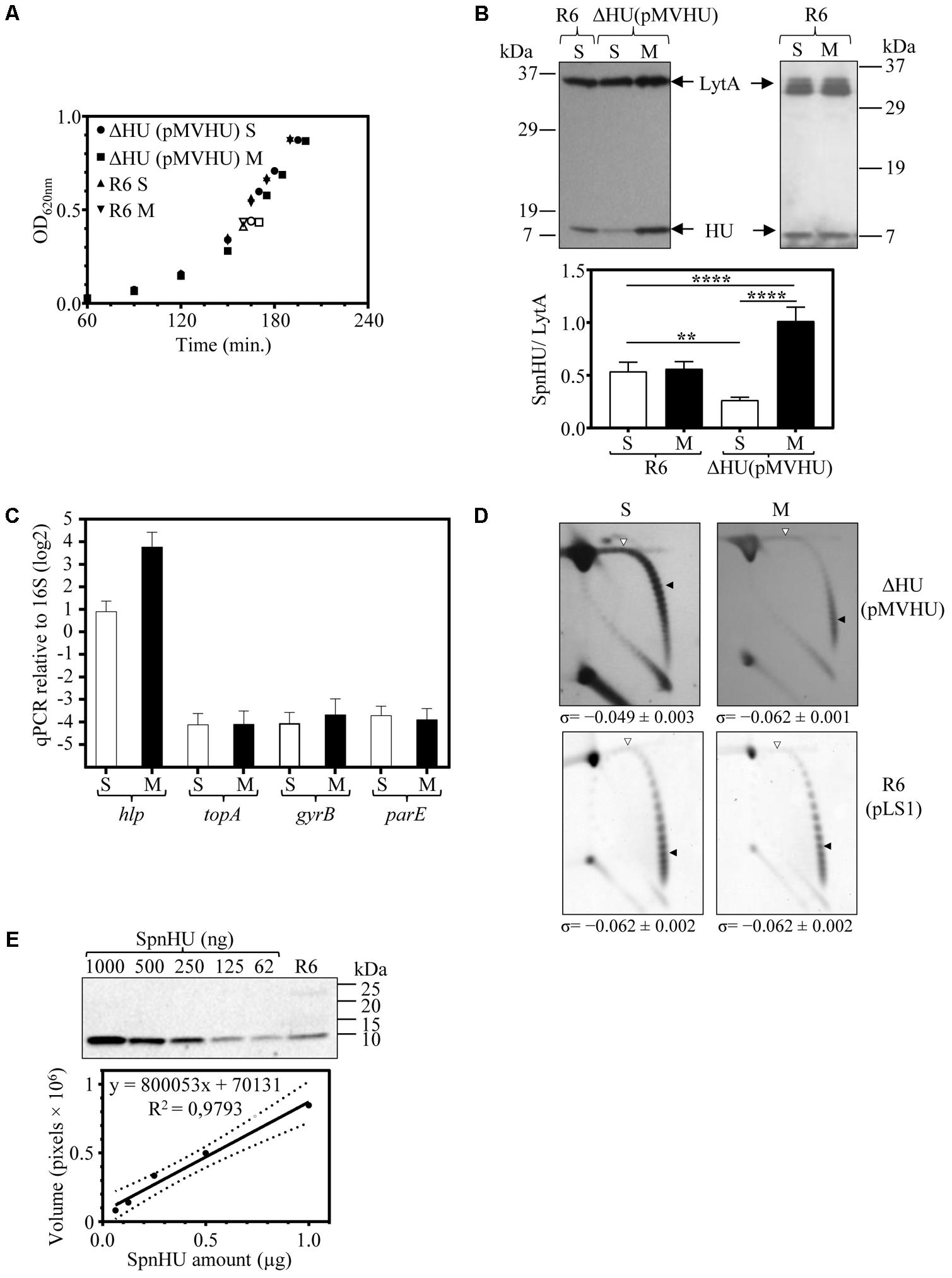
FIGURE 4. Oscillations in the amount of SpnHU affect the level of DNA supercoiling. Cultures of R6, R6 (pLS1), and ΔHU (pMVHU) were grown in medium containing sucrose. When the cultures reached OD620 nm= 0.2, they were diluted 50-fold in medium containing either sucrose (S) or maltose (M). Samples were obtained when the cultures reached OD620 nm= 0.4. (A) Growth kinetics of ΔHU (pMVHU) and R6 strains grown in sucrose and maltose-supplemented media. Open points indicate the time at which samples were collected. (B) Western blot analysis of SpnHU levels in samples grown in medium containing sucrose (S) or maltose (M). Crude cell extracts (20 μg) were separated by SDS-PAGE and blotted. To quantify SpnHU, values were divided by the amount of LytA protein as a loading control. Values are the average ± SD of three independent replicates. (∗P < 0.0332, ∗∗∗P < 0.0002). (C) qRT-PCR values of hlp, topA, gyrB, and parE genes in cells of ΔHU (pMVHU) grown in medium containing maltose or sucrose. Values of represented amplicons were calculated relative to those of 16S rDNA. Represented values are the average ± SD of three independent replicates. (D) Plasmid DNAs were isolated and subjected to 2D agarose gel electrophoresis as described in Section “Materials and Methods.” Supercoiling density (σ) values are averages ± SD from three independent replicates. (E) Western blot analysis of different amounts of purified SpnHU and a crude cell extract of R6 strain to calculate the number of SpnHU molecules per cell. The linearity of the SpnHU amount and volume expressed is presented as the number of pixels.
The expression of hlp in strain ΔHU (pMVHU) grown in maltose or sucrose was evaluated by qRT-PCR as indicated in Section “Materials and Methods”. The induction of Pmal triggered an increase by 7.3-fold in the expression of hlp (Figure 4C). Taken together these data revealed that variations in the amount of SpnHU were possible when the Pmal expression system was used in strain ΔHU (pMVHU).
To assess whether these SpnHU variations affected DNA supercoiling levels in vivo, the level of chromosome supercoiling in strains ΔHU (pMVHU) and R6 (pLS1) was inferred from the distribution of topoisomers of plasmids pMVHU and pLS1 present in the cells. It has been previously shown that the supercoiling density of small plasmids can be subrogated to the chromosome (Pruss et al., 1982; Weitao et al., 2000; Ferrándiz et al., 2010). Plasmids pMVHU and pLS1 replicate by a rolling circle mechanism (de la Campa et al., 1990), and all their genes are transcribed in the same direction, thus avoiding transcription interference during replication. Plasmids were extracted from cells grown in medium either with sucrose or maltose. The distribution of topoisomers was analyzed by two-dimensional agarose gel electrophoresis in the presence of chloroquine, which intercalates in the DNA and thus permits the separation of molecular DNA species by mass and shape. Chloroquine was used in the second dimension at 2 μg/ml, which induced ΔLk of -20 and -14 for plasmids pMVHU and pLS1, respectively.
Quantification of DNA supercoiling was performed by calculating the supercoiling density (σ) as described in Section “Materials and Methods”. Values of σ for pMVHU extracted from ΔHU cells grown in medium supplemented either with sucrose or maltose were -0.049 ± 0.003 and -0.062 ± 0.001, respectively (Figure 4D), which indicated a significant increase of 21% in the value of σ (P = 0.0004) when SpnHU levels dropped 3.9-fold. In the case of pLS1 extracted from strain R6 grown in either sucrose or maltose-supplemented medium, the σ values remained the same at -0.062 ± 0.002, (Figure 4D), demonstrating that the carbon source employed did not affect DNA supercoiling. Taken together these data revealed that the decrease in DNA supercoiling of ΔHU (pMVHU) grown in sucrose occurred independently of the carbon source and therefore correlated with the decline in the amount of SpnHU. This decrease was 50.9 and 72.3% compared with the R6 grown in either sucrose or maltose, and ΔHU (pMVHU) grown in maltose, respectively (Figure 4B). Conversely, we did not observe an increase in the level of negative supercoiling when the levels of protein duplicated, i.e., in the case of ΔHU (pMVHU) grown with maltose compared with R6 grown in either sucrose or maltose (Figure 4D).
In pneumococcus, the decrease in negative DNA supercoiling, i.e., relaxation triggers a homeostatic response that involves changes in the expression of genes encoding DNA topoisomerases (Ferrándiz et al., 2010). We examined whether the DNA relaxation caused by the decrease in SpnHU prompted this effect. We measured the expression of genes coding topoisomerase I, gyrase, and one of the subunits of topoisomerase IV: topA, gyrB, and parE, respectively, by qRT-PCR. No alterations were found in the expression of either topA, gyrB or parE when ΔHU (pMVHU) was grown in the presence of both sucrose and maltose (Figure 4C). This result supports the conclusion that the drop of 50.9% in SpnHU was the responsible for the 21% increase in DNA relaxation, which did not affect the levels of DNA topoisomerases.
To determine the number of SpnHU molecules present in cells during the exponential growth phase, Western blot analysis with different amounts of purified SpnHU and crude extracts of the R6 strain (Figure 4E) were performed, as explained in Section “Materials and Methods.” The estimated number of SpnHU molecules was 67240 ± 14700.
An Increase in SpnHU Attenuated DNA Relaxation Induced by Novobiocin
To further analyze the contribution of SpnHU to DNA supercoiling levels, ΔHU (pMVHU) was grown in medium containing either sucrose or maltose in the absence or presence of novobiocin (NOV), which specifically inhibits the activity of gyrase B subunit, leading to DNA relaxation (Ferrándiz et al., 2010). No differences in growth were found when ΔHU (pMVHU) was cultured with sucrose or maltose in the absence of NOV (duplication times of 84.1 ± 2.1 min and 84.1 ± 4.3 min, n = 3, respectively) (Figure 5A). This finding shows that small changes in SpnHU protein levels do not contribute to a loss in cell viability. However, in the presence of 1 μg/ml NOV (1 × MIC), ΔHU (pMVHU) only grew in the maltose medium (doubling time of 288.5 ± 26.4 min, n = 3) (Figure 5A), which shows that more SpnHU partially counteracts the effect of NOV on DNA supercoiling.
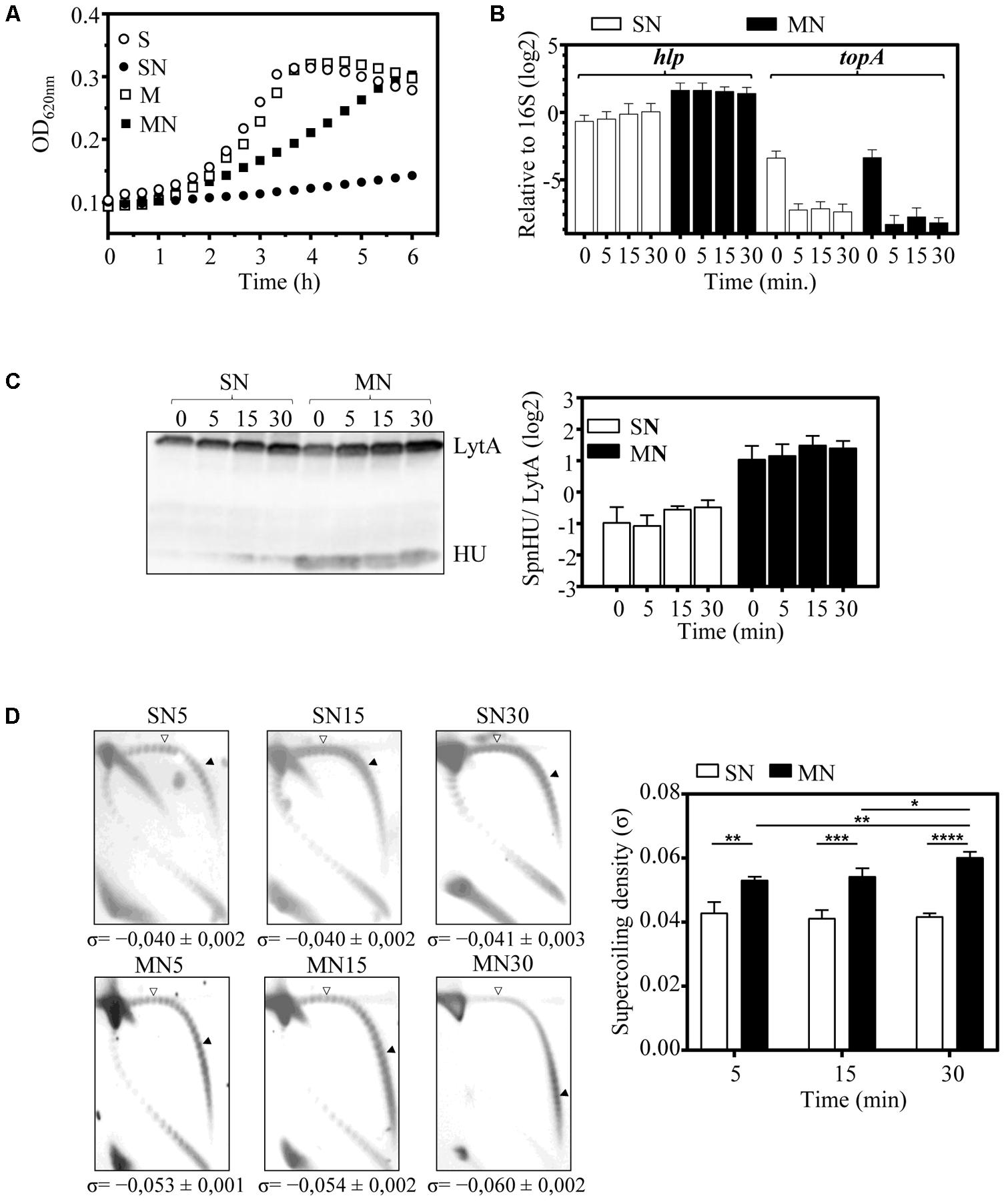
FIGURE 5. The increase in SpnHU protein levels attenuates the relaxation effect of NOV. A culture of ΔHU (pMVHU) was grown in medium containing sucrose until it reached an OD620 nm= 0.2, and then it was diluted 50-fold in four different media: two containing either sucrose (S), or maltose (M) and two containing either sucrose or maltose, with 1 μg/ml of NOV (SN or MN). Cells were collected 5, 15, and 30 min after addition of the drug. (A) Growth kinetics of ΔHU (pMVHU) cultures. (B) qRT-PCR values of hlp and topA genes relative to those of 16S rDNA after treatment with NOV. Represented values are the average ± SD of three independent replicates. (C) Western blot analysis of SpnHU protein levels in cells exposed to NOV. Crude cell extracts (20 μg) were separated by SDS-PAGE, blotted, and incubated with antibodies directed against SpnHU and LytA. To quantify SpnHU, values were divided by the amount of LytA protein as a loading control. Three independent replicates are represented. (D) Plasmid DNA from cells exposed to NOV at different times was isolated and subjected to 2D agarose gel electrophoresis as described in Section “Materials and Methods.” Supercoiling density (σ) values are averages ± SD from three independent replicates. (∗P < 0.0332, ∗∗P < 0.0021, ∗∗∗P < 0.0002, ∗∗∗∗P < 0.0001).
It has been previously shown that the treatment of S. pneumoniae with NOV induces DNA relaxation and triggers a homeostatic response to restore the level of DNA supercoiling when the induced relaxation is less than 25% (Ferrándiz et al., 2010). This response includes transcriptional changes in topoisomerase genes: up-regulation of gyrase genes and down-regulation of topoisomerase I and IV genes (Ferrándiz et al., 2010). Among topoisomerases, topoisomerase I (encoded by topA) is mainly responsible for the maintenance of DNA supercoiling levels in S. pneumoniae, since the amount of topA mRNA correlated with the level of DNA supercoiling during cellular homeostatic responses either to DNA relaxation or to increased negative DNA supercoiling (Ferrándiz et al., 2016). Expression of both topA and hlp genes was measured by qRT-PCR after treatment of ΔHU (pMVHU) with NOV. We observed decreases of 16- and 24-fold (mean of 5, 15, and 30 min) in topA mRNA when the medium contained sucrose and maltose, respectively (Figure 5B). These levels were in the range of those observed previously (Ferrándiz et al., 2010), and indicated that NOV was working as expected. For hlp, 3.6-fold (mean of 5, 15, and 30 min) overexpression was detected in cells grown with maltose compared with cells grown with sucrose (Figure 5B). This overexpression of hlp corresponded to a fourfold (mean of 5, 15, and 30 min) increase in the amount of SpnHU (Figure 5C). The difference in the amount of SpnHU observed between cells grown with maltose and cells grown with sucrose remained similar in the presence and absence of NOV in ΔHU (pMVHU) (Figures 4B, 5C). This result is expected since the hlp gene in this strain is under the control of the inducible promoter of maltose, the activity of which is not affected by NOV. Based on these results, we deduced that more SpnHU would allow the survival of ΔHU (pMVHU) to 1 × MIC NOV treatment probably because the level of DNA supercoiling was closer to the physiological value.
The DNA supercoiling level of ΔHU (pMVHU) grown with sucrose and maltose and in the presence or absence of 1 × MIC NOV was analyzed by two-dimensional agarose gel electrophoresis. Treatment of ΔHU (pMVHU) grown in sucrose with NOV resulted in σ = -0.040 (mean of 5, 15, and 30 min) (Figure 5D). This value was 18.4% (P = 0.03) and 35.5% (P = 0.0001) lower than those of ΔHU (pMVHU) grown in sucrose and without NOV (σ = -0.049) and R6 grown both in sucrose or maltose and without NOV (σ = -0.062), respectively (Figure 4D). The 35.5% drop in the level of DNA supercoiling in ΔHU (pMVHU) grown in sucrose with NOV resulted from the sum of the relaxation induced by both the decrease in the amount of SpnHU (Figures 4B,D) and NOV treatment (Figure 5D). This DNA supercoiling level could not be restored over time, as observed for R6 treated with concentrations of NOV that induced decreases in supercoiling higher than 25% (Ferrándiz et al., 2010). When we compared the DNA supercoiling of ΔHU (pMVHU) grown in maltose with NOV, we observed a significant decrease of 14.5% in DNA supercoiling 5 min after NOV addition with respect to the untreated ΔHU (pMVHU) or R6 (P = 0.0002 and P = 0.0005, respectively) (Figures 4D, 5D). This decrease was 1.7-fold lower than that observed for R6 treated with the same concentration of NOV (Ferrándiz et al., 2010) and correlated with a 2.1-fold higher amount of SpnHU (Figure 4B). The level of DNA supercoiling was fully recovered after 30 min of treatment (Figure 5D).
Discussion
We investigated SpnHU in this study, first by characterizing its biochemical activity and second by analyzing its activity in vivo. In terms of its in vitro activity, we have observed that it behaves like most of their homologous HU proteins. It preferably forms dimers and binds to supercoiled DNA with higher affinity than to linear or nicked DNA. Furthermore, it is able to constrain relaxed DNA in the presence of topoisomerase I in an efficient manner.
Regarding the in vivo activity of SpnHU, we have shown that this protein is involved in the maintenance of DNA supercoiling, and only small variations in the amount of SpnHU are possible to avoid disturbing the proper DNA topology. Therefore, a tight regulation of hlp would be expected to preserve an adequate amount of this protein. Transcriptional regulation of hlp in S. pneumoniae would differ from that of the hupA and hupB genes in E. coli, the expression of which is modulated by Fis and by the catabolite repressor protein CRP (Claret and Rouvière-Yaniv, 1996). Fis is absent in S. pneumoniae, and hlp is most likely regulated by global changes in DNA supercoiling. Moreover, this gene is located in one of the topology-reactive gene clusters, or domains, in which the pneumococcus genome is divided (Ferrándiz et al., 2010). Genes in these domains show a coordinated transcriptional regulation in response to topological changes (Ferrándiz et al., 2010; de la Campa et al., 2017). Specifically, hlp is located in a domain in which transcription is down-regulated when the DNA is relaxed by the inhibition of DNA gyrase by NOV. This kind of regulation by DNA supercoiling also affects all the DNA topoisomerase genes in S. pneumoniae (Ferrándiz et al., 2010). Transcriptional down-regulation of hlp would be involved in the known homeostatic response to DNA relaxation by which gyrase genes gyrA and gyrB are up-regulated and topA is down-regulated (Ferrándiz et al., 2010). All these changes would balance the DNA supercoiling after relaxation induced by NOV.
In addition, we have shown that SpnHU is essential for the viability of pneumococcal R6 cells, supporting some previous studies of essential genes in S. pneumoniae (Thanassi et al., 2002; Song et al., 2005; Mobegi et al., 2014). These studies were performed with strain D39, its derivatives Rx1 and R6, and TIGR4. Other studies in which hlp was not essential, used TIGR4, D39, and the Spain9V-3 SP195 strain (van Opijnen et al., 2009; van Opijnen and Camilli, 2012; Verhagen et al., 2014). Differences with respect to the essentiality of the hlp gene may be related to the length of the gene, with short genes such as hlp (273 bp) being less susceptible to disruption by transposon insertions. In fact, among all the essential genes found in two of the mentioned studies (van Opijnen et al., 2009; Mobegi et al., 2014), only 9% have sizes < 300 bp despite representing 18% of the genome. However, 81% have sizes > 300 bp (representing 91% of the genome).
The survival of S. pneumoniae seems to be possible only when at least a minimum amount of this protein is present in the cell. The cellular dependence of this protein could be due to its importance in preserving DNA topology, which is crucial for DNA metabolic processes. In fact, in E. coli, HU modulates the transcription of genes that respond to stress conditions and SOS induction (Oberto et al., 2009), and a possible role in the coordination of replication with chromosome segregation has also been observed in Mycobacterium tuberculosis as HupB in this bacterium preferably associates with the origin organizing the newly replicated oriC regions (Hołówka et al., 2017). In the phylogenetically related S. intermedius, SiHU downregulation leads to an alteration of the cell transcription program, with changes in nucleoid segregation, cell division and cell surface properties (Liu et al., 2008b).
In contrast to other bacteria, such as E. coli, the landscape of NAPs in S. pneumoniae is drastically reduced such that NAPs such as FIS, H-NS, or IHF are not present. The same occurs in other bacteria of the Firmicutes phylum, such as B. subtilis, which also lacks FIS, H-NS, or IHF. However, this bacterium has some DNA-bridging proteins that belong to the Lrp family, which could be involved in the maintenance of DNA topology (Beloin et al., 1997). These proteins are absent in S. pneumoniae, although we cannot exclude the possibility that a structural homologue of Fis or other NAPs are present. This situation suggests that SpnHU plays an important role in the preservation of DNA supercoiling. Indeed, the observed 21% reduction of the supercoiling level when the cell has a twofold lower amount of SpnHU can be ascribed to the wrapping property of this protein since the level of topoisomerases, which are mainly responsible for supercoiling variations, does not change. In S. intermedius, downregulation of SiHU is possible due to the induced expression of a Si-hlp antisense RNA, which triggers nucleoids expansion (Liu et al., 2008b), supporting the decrease in HU and leading to a lower level of restrained DNA supercoils.
Taken together these results show that in vivo, SpnHU contributes to the maintenance of DNA supercoiling and that this essential protein can only be partially removed. The reduction of SpnHU seems to only be possible up to a level where the negative supercoiling is not reduced beyond 20%, which is the limit of cell survival (Ferrándiz et al., 2010). Although up to a 40% increase in negative supercoiling can be tolerated by the cell (Ferrándiz et al., 2016), hypernegative supercoiling is not observed when SpnHU levels are increased. Similarly, overproduction of HU from E. coli causes no change in DNA supercoiling (Mc Govern et al., 1994). Unlike E. coli, in which the protein is induced approximately 40-fold, a moderate increase (only fourfold) was observed herein. The possible effect of a saturation of SpnHU bound to the DNA does not explain the inability of the increase in HU to result in an increase in supercoiling. In fact, HU and Fis are the most abundant NAPs in exponentially growing E. coli cells. The approximate number of dimeric molecules per cell is 30000 (Ali Azam et al., 1999), which would allow HU dimers to be uniformly distributed along the E. coli chromosome every ≈ 190 bp (Azam et al., 2000). In S. pneumoniae, we estimated that the number of SpnHU molecules per cell is approximately 34000 dimers (67240 ± 14700 monomers), which is in the same range as that observed for E. coli. However, the size of pneumococcus chromosome (2038615 bp) is almost half that of E. coli, resulting in an HU distribution of 1 dimer per ≈ 60 bp. As in E. coli, the amount of SpnHU is not enough to coat the entire chromosome, taking into account that HU dimers need 21 bp DNA to form a helical complex, as demonstrated for Anabaena HU-DNA cocrystals (Swinger et al., 2003). The formation of flexible bends induced by HU dimers has been proposed to explain the DNA compaction. These nucleosome-like structures are rescinded in the case of high HU to DNA ratios due to the formation of a rigid nucleoprotein filament (Sagi et al., 2004; van Noort et al., 2004) potentially explaining why increases in HU not result in hypernegative supercoiling.
We found that a moderate increase in SpnHU levels attenuates the effect of relaxation induced by NOV, in agreement with our biochemical results. SpnHU constrains the supercoiling, counteracting the effect of a less active gyrase. Both a drop in topoisomerase I and an increased amount of SpnHU can attenuate the effect of the drug, restoring an adequate DNA supercoiling balance and thus allowing cells to grow at the inhibitory concentration of NOV. Taken together, these data seem to indicate that the amount of this protein is critical for pneumococcus survival, and ability to cope with moderate changes in DNA supercoiling.
Author Contributions
AC and M-JF conceived, designed, and supervised the study. DC purified (H6)SpnHU and participated with SA in the EMSA experiments. M-JF carried out the remaining experiments and wrote the manuscript. All authors read and actively participated in the correction of the manuscript. The manuscript has been approved by all authors for publication.
Funding
This work was supported by the Ministerio de Economía y Competitividad (BIO2014-55462-R and BIO2017-82951-R).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Ali Azam, T., Iwata, A., Nishimura, A., Ueda, S., and Ishihama, A. (1999). Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid. J. Bacteriol. 181, 6361–6370.
Azam, T. A., Hiraga, S., and Ishihama, A. (2000). Two types of localization of the DNA-binding proteins within the Escherichia coli nucleoid. Genes Cells 5, 613–626. doi: 10.1046/j.1365-2443.2000.00350.x
Azam, T. A., and Ishihama, A. (1999). Twelve species of the nucleoid-associated protein from Escherichia coli. Sequence recognition specificity and DNA binding affinity. J. Biol. Chem. 274, 33105–33113. doi: 10.1074/jbc.274.46.33105
Ballester, S., López, P., Alonso, J. C., Espinosa, M., and Lacks, S. A. (1986). Selective advantage of deletions enhancing chloramphenicol acetyltransferase gene expression in S. pneumoniae plasmids. Gene 41, 153–163. doi: 10.1016/0378-1119(86)90094-6
Beloin, C., Ayora, S., Exley, R., Hirschbein, L., Ogasawara, N., Kasahara, Y., et al. (1997). Characterization of an lrp-like (lrpC) gene from Bacillus subtilis. Mol. Gen. Genet. 256, 63–71. doi: 10.1007/s004380050546
Berger, M., Farcas, A., Geertz, M., Zhelyazkova, P., Brix, K., Travers, A., et al. (2010). Coordination of genomic structure and transcription by the main bacterial nucleoid-associated protein HU. EMBO Rep. 11, 59–64. doi: 10.1038/embor.2009.232
Bergey, E. J., and Stinson, M. W. (1988). Heparin-inhibitable basement membrane-binding protein of Streptococcus pyogenes. Infect. Immun. 56, 1715–1721.
Bertin, P., Benhabiles, N., Krin, E., Laurent-Winter, C., Tendeng, C., Turlin, E., et al. (1999). The structural and functional organization of H-NS-like proteins is evolutionarily conserved in gram-negative bacteria. Mol. Microbiol. 31, 319–329. doi: 10.1046/j.1365-2958.1999.01176.x
Biasini, M., Bienert, S., Waterhouse, A., Arnold, K., Studer, G., Schmidt, T., et al. (2014). SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 42, W252–W258. doi: 10.1093/nar/gku340
Briat, J. F., Letoffee, S., Mache, R., and Rouviere-Yaniv, J. (1984). Similarity between the bacterial histone-like HU and a protein from spinach chloroplasts. FEBS Lett. 172, 75–79. doi: 10.1016/0014-5793(84)80877-7
Caron, F., Jacq, C., and Rouvière-Yaniv, J. (1979). Characterization of a histone-like protein extracted from yeast mitochondria. Proc. Natl. Acad. Sci. U.S.A. 76, 4265–4269. doi: 10.1073/pnas.76.9.4265
Chen, C., Ghosh, S., and Grove, A. (2004). Substrate specificity of Helicobacter pylori histone-like HU protein is determined by insufficient stabilization of DNA flexure points. Biochem. J. 383, 343–351. doi: 10.1042/BJ20040938
Chodavarapu, S., Felczak, M. M., Yaniv, J. R., and Kaguni, J. M. (2008). Escherichia coli DnaA interacts with HU in initiation at the E. coli replication origin. Mol. Microbiol. 67, 781–792. doi: 10.1111/j.1365-2958.2007.06094.x
Choi, S. H., and Stinson, M. W. (1989). Purification of a Streptococcus mutans protein that binds to heart tissue and glycosaminoglycans. Infect. Immun. 57, 3834–3840.
Choi, S. H., and Stinson, M. W. (1991). Binding of a Streptococcus mutans cationic protein to kidney in vitro. Infect. Immun. 59, 537–543.
Christodoulou, E., Rypniewski, W. R., and Vorgias, C. R. (2003). High-resolution X-ray structure of the DNA-binding protein HU from the hyper-thermophilic Thermotoga maritima and the determinants of its thermostability. Extremophiles 7, 111–122. doi: 10.1007/s00792-002-0302-7
Christodoulou, E., and Vorgias, C. E. (2002). The thermostability of DNA-binding protein HU from mesophilic, thermophilic, and extreme thermophilic bacteria. Extremophiles 6, 21–31. doi: 10.1007/s007920100235
Claret, L., and Rouvière-Yaniv, J. (1996). Regulation of HU alpha and HU beta by CRP and FIS in Escherichia coli. J. Mol. Biol. 263, 126–139. doi: 10.1006/jmbi.1996.0564
Croinin, O., Carroll, R. K., Kelly, A., and Dorman, C. J. (2006). Roles for DNA supercoiling and the Fis protein in modulating expression of virulence genes during intracellular growth of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 62, 869–882. doi: 10.1111/j.1365-2958.2006.05416.x
Dame, R. T., Wyman, C., and Goosen, N. (2000). H-NS mediated compaction of DNA visualised by atomic force microscopy. Nucleic Acids Res. 28, 3504–3510. doi: 10.1093/nar/28.18.3504
de la Campa, A. G., del Solar, G. H., and Espinosa, M. (1990). Initiation of replication of plasmid pLS1. The initiator protein RepB acts on two distant DNA regions. J. Mol. Biol. 213, 247–262. doi: 10.1016/S0022-2836(05)80188-3
de la Campa, A. G., Ferrándiz, M. J., Martín-Galiano, A. J., García, M. T., and Tirado-Vélez, J. M. (2017). The transcriptome of Streptococcus pneumoniae induced by local and global changes in supercoiling. Front. Microbiol. 8:1447. doi: 10.3389/fmicb.2017.01447
de los Rios, S., and Perona, J. J. (2007). Structure of the Escherichia coli leucine-responsive regulatory protein Lrp reveals a novel octameric assembly. J. Mol. Biol. 366, 1589–1602. doi: 10.1016/j.jmb.2006.12.032
de Vries, R. (2010). DNA condensation in bacteria: interplay between macromolecular crowding and nucleoid proteins. Biochimie 92, 1715–1721. doi: 10.1016/j.biochi.2010.06.024
Dillon, S. C., and Dorman, C. J. (2010). Bacterial nucleoid-associated proteins, nucleoid structure and gene spression. Nat. Rev. Microbiol. 8, 185–195. doi: 10.1038/nrmicro2261
Dorman, C. J. (2013). Co-operative roles for DNA supercoiling and nucleoid-associated proteins in the regulation of bacterial transcription. Biochem. Soc. Trans. 41, 542–547. doi: 10.1042/BST20120222
Dorman, C. J. (2014). Function of nucleoid-associated proteins in chromosome structuring and transcriptional regulation. J. Mol. Microbiol. Biotechnol. 24, 316–331. doi: 10.1159/000368850
Drlica, K., and Rouvière-Yaniv, J. (1987). Histone like proteins of bacteria. Microbiol. Rev. 51, 301–319.
Fenoll, A., Muñoz, R., García, E., and de la Campa, A. G. (1994). Molecular basis of the optochin-sensitive phenotype of pneumococcus: characterization of the genes encoding the F0 complex of the Streptococcus pneumoniae and Streptococcus oralis H+-ATPases. Mol. Microbiol. 12, 587–598. doi: 10.1111/j.1365-2958.1994.tb01045.x
Ferrándiz, M. J., and de la Campa, A. G. (2014). The fluoroquinolone levofloxacin triggers the transcriptional activation of iron transport genes that contribute to cell death in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 58, 247–257. doi: 10.1128/AAC.01706-13
Ferrándiz, M. J., Martín-Galiano, A. J., Arnanz, C., Camacho-Soguero, I., Tirado-Vélez, J. M., and de la Campa, A. G. (2016). An increase in negative supercoiling in bacteria reveals topology-reacting gene clusters and a homeostatic response mediated by the DNA topoisomerase I gene. Nucleic Acids Res. 44, 7292–7303. doi: 10.1093/nar/gkw602
Ferrándiz, M. J., Martín-Galiano, A. J., Schvartzman, J. B., and de la Campa, A. G. (2010). The genome of Streptococcus pneumoniae is organized in topology-reacting gene clusters. Nucleic Acids Res. 38, 3570–3581. doi: 10.1093/nar/gkq106
García, M. T., Blazquez, M. A., Ferrándiz, M. J., Sanz, M. J., Silva-Martín, N., Hermoso, J. A., et al. (2011). New alkaloid antibiotics that target the DNA topoisomerase I of Streptococcus pneumoniae. J. Biol. Chem. 286, 6402–6413. doi: 10.1074/jbc.M110.148148
Ghosh, S., and Grove, A. (2004). Histone-like protein HU from Deinococcus radiodurans binds preferentially to four-way DNA junctions. J. Mol. Biol. 337, 561–571. doi: 10.1016/j.jmb.2004.02.010
Hołówka, J., Trojanowski, D., Ginda, K., Wojtaś, B., Gielniewski, B., Jakimowicz, D., et al. (2017). HupB is a bacterial nucleoid-associated protein with an indispensable eukaryotic-like tail. mBio 8:e01272-17. doi: 10.1128/mBio.01272-17
Kahramanoglou, C., Seshasayee, A. S., Prieto, A. I., Ibberson, D., Schmidt, S., Zimmermann, J., et al. (2011). Direct and indirect effects of H-NS and Fis on global gene expression control in Escherichia coli. Nucleic Acids Res. 39, 2073–2091. doi: 10.1093/nar/gkq934
Kamashev, D., and Rouviere-Yaniv, J. (2000). The histone-like protein HU binds specifically to DNA recombination and repair intermediates. EMBO J. 19, 6527–6535. doi: 10.1093/emboj/19.23.6527
Kamau, E., Tsihlis, N. D., Simmons, L. A., and Grove, A. (2005). Surface salt bridges modulate the DNA site size of bacterial histone-like HU proteins. Biochem. J. 390, 49–55. doi: 10.1042/BJ20050274
Kasho, K., Fujimitsu, K., Matoba, T., Oshima, T., and Katayama, T. (2014). Timely binding of IHF and Fis to DARS2 regulates ATP-DnaA production and replication initiation. Nucleic Acids Res. 42, 13134–13149. doi: 10.1093/nar/gku1051
Kelley, L. A., Mezulis, S., Yates, C. M., Wass, M. N., and Sternberg, M. J. E. (2015). The Phyre2 web portal for protein modelling, prediction and analysis. Nat. Protoc. 10, 845–858. doi: 10.1038/nprot.2015.053
Kim, D. H., Im, H., Jee, J. G., Jang, S. B., Yoon, H. J., Kwon, A. E., et al. (2014). β-arm flexibility of HU from Staphylococcus aureus dictates the DNA binding and recognition mechanism. Acta Crystallogr. D Biol. Crystallogr. 70, 3273–3289. doi: 10.1107/S1399004714023931
Kloosterman, T. G., van der Kooi-Pol, M. M., Bijlsma, J. J., and Kuipers, O. P. (2007). The novel transcriptional regulator SczA mediates protection against Zn2+ stress by activation of the Zn2+-resistance gene czcD in Streptococcus pneumoniae. Mol. Microbiol. 65, 1049–1063. doi: 10.1111/j.1365-2958.2007.05849.x
Lacks, S. A., López, P., Greenberg, B., and Espinosa, M. (1986). Identification and analysis of genes for tetracycline resistance and replication functions in the broad-host-range plasmid pLS1. J. Mol. Biol. 192, 753–765. doi: 10.1016/0022-2836(86)90026-4
Liu, D., Yumoto, H., Hirota, K., Murakami, K., Takahashi, K., Hirao, K., et al. (2008a). Histone-like DNA binding protein of Streptococcus intermedius induces the expression of pro-inflammatory cytokines in human monocytes via activation of ERK1/2 and JNK pathways. Cell Microbiol. 10, 262–276. doi: 10.1111/j.1462-5822.2007.01040.x
Liu, D., Yumoto, H., Murakami, K., Hirota, K., Ono, T., Nagamune, H., et al. (2008b). The essentiality and involvement of Streptococcus intermedius histone-like DNA-binding protein in bacterial viability and normal growth. Mol. Microbiol. 68, 1268–1282. doi: 10.1111/j.1365-2958.2008.06232.x
Luijsterburg, M. S., Noom, M. C., Wuite, G. J., and Dame, R. T. (2006). The architectural role of nucleoid-associated proteins in the organization of bacterial chromatin: a molecular perspective. J. Struct. Biol. 156, 262–272. doi: 10.1016/j.jsb.2006.05.006
Luijsterburg, M. S., White, M. F., van Driel, R., and Dame, R. T. (2008). The major architects of chromatin: architectural proteins in bacteria, archaea and eukaryotes. Crit. Rev. Biochem. Mol. Biol. 43, 393–418. doi: 10.1080/10409230802528488
Lynch, J. P. III, and Zhanel, G. G. (2009). Streptococcus pneumoniae: epidemiology, risk factors, and strategies for prevention. Semin. Respir. Crit. Care Med. 30, 189–209. doi: 10.1055/s-0029-1202938
Martín-Galiano, A. J., Yuste, J., Cercenado, M. I., and de la Campa, A. G. (2014). Inspecting the potential physiological and biomedical value of 44 conserved uncharacterised proteins of Streptococcus pneumoniae. BMC Genomics 15:652. doi: 10.1186/1471-2164-15-652
Martín-Parras, L., Lucas, I., Martínez-Robles, M. L., Hernández, P., Krimer, D. B., Hyrien, O., et al. (1998). Topological complexity of different populations of pBR322 as visualized by two-dimensional agarose gel electrophoresis. Nucleic Acids Res. 26, 3424–3432. doi: 10.1093/nar/26.14.3424
Mc Govern, V., Higgins, N. P., Chiz, R. S., and Jaworski, A. (1994). H-NS over-expression induces an artificial stationary phase by silencing global transcription. Biochimie 76, 1019–1029. doi: 10.1016/0300-9084(94)90026-4
Menzel, R., and Gellert, M. (1983). Regulation of the genes for E. coli DNA gyrase: homeostatic control of DNA supercoiling. Cell 34, 105–113. doi: 10.1016/0092-8674(83)90140-X
Micka, B., and Marahiel, M. A. (1992). The DNA-binding protein HBsu is essential for normal growth and development in Bacillus subtilis. Biochimie 74, 641–650. doi: 10.1016/0300-9084(92)90136-3
Mobegi, F. M., van Hijum, S. A., Burghout, P., Bootsma, H. J., de Vries, S. P., van der Gaast-de Jongh, C. E., et al. (2014). From microbial gene essentiality to novel antimicrobial drug targets. BMC Genomics 15:958. doi: 10.1186/1471-2164-15-958
Mukherjee, A., Sokunbi, A. O., and Grove, A. (2008). DNA protection by histone-like protein HU from the hyperthermophilic eubacterium Thermotoga maritima. Nucleic Acids Res. 36, 3956–3968. doi: 10.1093/nar/gkn348
Murphy, L. D., and Zimmerman, S. B. (1995). Condensation and cohesion of lambda DNA in cell extracts and other media: implications for the structure and function of DNA in prokaryotes. Biophys. Chem. 57, 71–92. doi: 10.1016/0301-4622(95)00047-2
Nieto, C., Puyet, A., and Espinosa, M. (2001). MalR-mediated regulation of the Streptococcus pneumoniae malMP operon at promoter PM. Influence of a proximal divergent promoter region and competition between MalR and RNA polymerase proteins. J. Biol. Chem. 276, 14946–14954. doi: 10.1074/jbc.M010911200
Oberto, J., Nabti, S., Jooste, V., Mignot, H., and Rouviere-Yaniv, J. (2009). The HU regulon is composed of genes responding to anaerobiosis, acid stress, high osmolarity and SOS induction. PLoS. One 4:e4367. doi: 10.1371/journal.pone.0004367
O’Neil, P., Lovell, S., Mehzabeen, N., Battaile, K., and Biswas, I. (2016). Crystal structure of histone-like protein from Streptococcus mutans refined to 1.9 A resolution. Acta Crystallogr. F Struct. Biol. Commun. 72, 257–262. doi: 10.1107/S2053230X1600217X
Ouafa, Z. A., Reverchon, S., Lautier, T., Muskhelishvili, G., and Nasser, W. (2012). The nucleoid-associated proteins H-NS and FIS modulate the DNA supercoiling response of the pel genes, the major virulence factors in the plant pathogen bacterium Dickeya dadantii. Nucleic Acids Res. 40, 4306–4319. doi: 10.1093/nar/gks014
Pato, M. L., and Banerjee, M. (1996). The mu strong gyrase-binding site promotes efficient synapsis of the prophage termini. Mol. Microbiol. 22, 283–292. doi: 10.1046/j.1365-2958.1996.00115.x
Pruss, G. J., Manes, S. H., and Drlica, K. (1982). Escherichia coli DNA topoisomerase I mutants: increased supercoiling is corrected by mutations near gyrase genes. Cell 31, 35–42. doi: 10.1016/0092-8674(82)90402-0
Sagi, D., Friedman, N., Vorgias, C., Oppenheim, A. B., and Stavans, J. (2004). Modulation of DNA conformations through the formation of alternative high-order HU-DNA complexes. J. Mol. Biol. 341, 419–428. doi: 10.1016/j.jmb.2004.06.023
Song, J. H., Ko, K. S., Lee, J. Y., Baek, J. Y., Oh, W. S., Yoon, H. S., et al. (2005). Identification of essential genes in Streptococcus pneumoniae by allelic replacement mutagenesis. Mol. Cells 19, 365–374.
Stinson, M. W., McLaughlin, R., Choi, S. H., Juarez, Z. E., and Barnard, J. (1998). Streptococcal histone-like protein: primary structure of hlpA and protein binding to lipoteichoic acid and epithelial cells. Infect. Immun. 66, 259–265.
Swinger, K. K., Lemberg, K. M., Zhang, Y., and Rice, P. A. (2003). Flexible DNA bending in HU-DNA cocrystal structures. EMBO J. 22, 3749–3760. doi: 10.1093/emboj/cdg351
Swinger, K. K., and Rice, P. A. (2004). IHF and HU: flexible architects of bent DNA. Curr. Opin. Struct. Biol. 14, 28–35. doi: 10.1016/j.sbi.2003.12.003
Tanaka, H., Goshima, N., Kohno, K., Kano, Y., and Imamoto, F. (1993). Properties of DNA-binding of HU heterotypic and homotypic dimers from Escherichia coli. J. Biochem. 113, 568–572. doi: 10.1093/oxfordjournals.jbchem.a124084
Tanaka, I., Appelt, K., Dijk, J., White, S. W., and Wilson, K. S. (1984). 3-A resolution structure of a protein with histone-like properties in prokaryotes. Nature 310, 376–381. doi: 10.1038/310376a0
Thanassi, J. A., Hartman-Neumann, S. L., Dougherty, T. J., Dougherty, B. A., and Pucci, M. J. (2002). Identification of 113 conserved essential genes using a high-throughput gene disruption system in Streptococcus pneumoniae. Nucleic Acids Res. 30, 3152–3162. doi: 10.1093/nar/gkf418
Thaw, P., Sedelnikova, S. E., Muranova, T., Wiese, S., Ayora, S., Alonso, J. C., et al. (2006). Structural insight into gene transcriptional regulation and effector binding by the Lrp/AsnC family. Nucleic Acids Res. 34, 1439–1449. doi: 10.1093/nar/gkl009
Travers, A., and Muskhelishvili, G. (2007). A common topology for bacterial and eukaryotic transcription initiation? EMBO Rep. 8, 147–151. doi: 10.1038/sj.embor.7400898
Tse-Dinh, Y. C. (1985). Regulation of the Escherichia coli DNA topoisomerase I gene by DNA supercoiling. Nucleic Acids Res. 13, 4751–4763. doi: 10.1093/nar/13.13.4751
van Noort, J., Verbrugge, S., Goosen, N., Dekker, C., and Dame, R. T. (2004). Dual architectural roles of HU: formation of flexible hinges and rigid filaments. Proc. Natl. Acad. Sci. U.S.A. 101, 6969–6974. doi: 10.1073/pnas.0308230101
van Opijnen, T., Bodi, K. L., and Camilli, A. (2009). Tn-seq: high-throughput parallel sequencing for fitness and genetic interaction studies in microorganisms. Nat. Methods 6, 767–772. doi: 10.1038/nmeth.1377
van Opijnen, T., and Camilli, A. (2012). A fine scale phenotype-genotype virulence map of a bacterial pathogen. Genome Res. 22, 2541–2551. doi: 10.1101/gr.137430.112
Verhagen, L. M., de Jonge, M. I., Burghout, P., Schraa, K., Spagnuolo, L., Mennens, S., et al. (2014). Genome-wide identification of genes essential for the survival of Streptococcus pneumoniae in human saliva. PLoS One 9:e89541. doi: 10.1371/journal.pone.0089541
Wada, M., Kano, Y., Ogawa, T., Okazaki, T., and Imamoto, F. (1988). Construction and characterization of the deletion mutant of hupA and hupB genes in Escherichia coli. J. Mol. Biol. 204, 581–591. doi: 10.1016/0022-2836(88)90357-9
Wang, X., Montero Llopis, P., and Rudner, D. Z. (2013). Organization and segregation of bacterial chromosomes. Nat. Rev. Genet. 14, 191–203. doi: 10.1038/nrg3375
Weitao, T., Nordstrom, K., and Dasgupta, S. (2000). Escherichia coli cell cycle control genes affect chromosome superhelicity. EMBO Rep. 1, 494–499. doi: 10.1093/embo-reports/kvd106
Winters, B. D., Ramasubbu, N., and Stinson, M. W. (1993). Isolation and characterization of a Streptococcus pyogenes protein that binds to basal laminae of human cardiac muscle. Infect. Immun. 61, 3259–3264.
World Health Organization (WHO) (2007). Pneumococcal Conjugate Vaccine for Childhood Immunization-WHO. Available at: http://www.who.int/iris/handle/10665/240901
Zechiedrich, E. L., Khodursky, A. B., Bachellier, S., Schneider, R., Chen, D., Lilley, D. M., et al. (2000). Roles of topoisomerases in maintaining steady-state DNA supercoiling in Escherichia coli. J. Biol. Chem. 275, 8103–8113. doi: 10.1074/jbc.275.11.8103
Zimmerman, S. B. (1993). Macromolecular crowding effects on macromolecular interactions: some implications for genome structure and function. Biochim. Biophys. Acta 1216, 175–185. doi: 10.1016/0167-4781(93)90142-Z
Keywords: histone-like protein, HU, Streptococcus pneumoniae, supercoiling, nucleoid
Citation: Ferrándiz M-J, Carreño D, Ayora S and de la Campa AG (2018) HU of Streptococcus pneumoniae Is Essential for the Preservation of DNA Supercoiling. Front. Microbiol. 9:493. doi: 10.3389/fmicb.2018.00493
Received: 05 December 2017; Accepted: 02 March 2018;
Published: 19 March 2018.
Edited by:
Marc Bramkamp, Ludwig-Maximilians-Universität München, GermanyReviewed by:
Morten Kjos, Norwegian University of Life Sciences, NorwayJolanta Zakrzewska-Czerwinska, University of Wrocław, Poland
Copyright © 2018 Ferrándiz, Carreño, Ayora and de la Campa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María-José Ferrándiz, mjferrandiz@isciii.es
† Present address: David Carreño, Department of Genetics, University of Leicester, Leicester, United Kingdom