 Peter-Louis Plaumann
Peter-Louis Plaumann Johannes Schmidpeter
Johannes Schmidpeter Marlis Dahl
Marlis Dahl Leila Taher
Leila Taher Christian Koch
Christian Koch- 1Division of Biochemistry, Department of Biology, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany
- 2Division of Bioinformatics, Department of Biology, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany
The hemibiotrophic plant pathogen Colletotrichum higginsianum infects Brassicaceae and in combination with Arabidopsis thaliana, represents an important model system to investigate various ecologically important fungal pathogens and their infection strategies. After penetration of plant cells by appressoria, C. higginsianum establishes large biotrophic primary hyphae in the first infected cell. Shortly thereafter, a switch to necrotrophic growth occurs leading to the invasion of neighboring cells by secondary hyphae. In a forward genetic screen for virulence mutants by insertional mutagenesis, we identified mutants that penetrate the plant but show a defect in the passage from biotrophy to necrotrophy. Genome sequencing and pulsed-field gel electrophoresis revealed that two mutants were lacking chromosome 11, encoding potential pathogenicity genes. We established a chromosome loss assay to verify that strains lacking this small chromosome abort infection during biotrophy, while their ability to grow on artificial media was not affected. C. higginsianum harbors a second small chromosome, which can be lost without effects on virulence or growth on agar plates. Furthermore, we found that chromosome 11 is required to suppress Arabidopsis thaliana plant defense mechanisms dependent on tryptophan derived secondary metabolites.
Introduction
Pathogenic fungi employ different strategies for causing disease. Biotrophic fungi like the rusts or powdery mildew fungi show a narrow host range and highly adapted genomes showing gene expansions of pathogenicity related genes while having a reduced repertoire of genes involved in primary and secondary metabolism. This specializes them to keep the infected host cells alive and suppress plant defense (Spanu et al., 2010; Duplessis et al., 2011; Raffaele and Kamoun, 2012). Extra- and intracellular effector proteins play a central role in regulating these interactions (Stergiopoulos and de Wit, 2009). Both effector-triggered susceptibility and effector-triggered immunity leave their marks in the genomes of pathogenic fungi which often show species-specific gene duplications and genome expansions involving effector functions required for virulence (Raffaele and Kamoun, 2012; Jones et al., 2016). Some effectors have been genetically identified and the recent genome analysis of many filamentous pathogens has potentially provided the full repertoire of virulence genes and effectors proteins (Raffaele and Kamoun, 2012; Moller and Stukenbrock, 2017). However, genetic insights into the function of individual pathogenicity determinants are frequently lacking because loss of function mutants often show no pathogenicity phenotype. This is usually attributed to functional redundancy of effector gene functions (Koeck et al., 2011).
The genomic analysis of fungal and oomycete pathogens has also led to the identification of genomic regions encoding high numbers of potential effector genes (Dong et al., 2015). Such regions, like the pathogenicity related gene clusters in Ustilago maydis (Kamper et al., 2006), are often sparsely covered with housekeeping genes (Raffaele and Kamoun, 2012). Magnaporthe strains, for example, may differ in the presence or absence of regions as large as 1.7 mb (Yoshida et al., 2009), while Verticillium dahliae contains a 4 mb lineage specific region (de Jonge et al., 2013). In the case of cereal rust fungi it was shown that a 2.5 mb large genomic region harboring Avr genes can be altered by unequal recombination (Chen et al., 2017), not only leading to genetic variations but also as a mechanism for breaking resistance. Chromosomal variations have been particularly well documented by comparative genomics (Dong et al., 2015) and are reflecting the adaptation to environmental cues and host defense mechanisms (Seidl and Thomma, 2014; Jones et al., 2016). Such regions of chromosomal polymorphisms can be very large and may even encompass whole chromosomes as in the case of Fusarium pathogens that harbor host range specific chromosomes (Ma et al., 2010). Such chromosomes may even become dispensable for growth under saprophytic conditions (Beadle et al., 2003) as is seen in Alternaria (Tsuge et al., 2013), Nectria haematococca (VanEtten et al., 1998) or in Leptosphaeria maculans (Balesdent et al., 2013). Dispensable chromosomes or supernumerary chromosomes, often also referred to as B-chromosomes (Jones, 1991) and originally known from the analysis of the rye plant genome, may be a hallmark of quickly evolving genomes (Camacho et al., 2000). 14 species of fungi have been reported to contain dispensable chromosomes (D’Ambrosio et al., 2017) including Colletotrichum gloeosporioides (Masel et al., 1996). A role for these in pathogenicity has not been identified in Colletotrichum species so far. The recent analysis of the C. higginsianum genome, in particular the assembly of complete chromosomes (O’Connell et al., 2012; Dallery et al., 2017), now allows to map virulence functions to individual genomic regions. C. higginsianum harbors 12 chromosomes, two much smaller than the others with less than 1000 kb DNA in size.
Colletotrichum higginsianum pursues a hemibiotrophic lifestyle (O’Connell et al., 2004). After penetration of the host plant with the help of appressoria, it forms primary biotrophic hyphae which have a characteristic large and bulbous appearance. The biotrophic stage during the infection in which the host cell remains alive, is confined to the first infected cell. After 2–3 days the pathogen forms filamentous secondary hyphae and switches its lifestyle and nutritional strategy. Then, necrotrophic hyphae colonize and kill the neighboring tissue (O’Connell et al., 2012). The switch to necrotrophic growth is accompanied with characteristic changes in gene expression and with the induction of genes encoding specific effector proteins (Kleemann et al., 2012). The signals orchestrating this developmental switch are largely unknown.
Together with Arabidopsis thaliana, C. higginsianum is a prominent model system for the analysis of hemibiotrophic plant pathogen interactions (O’Connell and Panstruga, 2006). Forward genetic screens, expression analysis and functional characterization of candidate genes have identified an array of pathogenicity determinants and effectors in C. higginsianum (Kleemann et al., 2008, 2012; Huser et al., 2009; O’Connell et al., 2012; Liu et al., 2013; Korn et al., 2015; Schmidpeter et al., 2017). We have previously isolated a series of Colletotrichum higginsianum mutants with different characteristic virulence defects on the host plant Arabidopsis thaliana (Korn et al., 2015; Schmidpeter et al., 2017). Some mutants penetrated the host successfully but were arrested during the biotrophic phase of infection. For two of these virulence mutants, we here report that the defect is due to the loss of the small chromosome 11. Colletotrichum higginsianum harbors two small chromosomes. Whereas chromosome 11 is critically important for virulence, we find that chromosome 12 can be lost without affecting pathogenicity.
Materials and Methods
Fungal and Bacterial Strains
Agrobacterium tumefaciens strain AGL1 (AGLO recA::bla pTiBo542ΔT Mop+ CbR) was used for fungal transformation as described (Korn et al., 2015). Colletotrichum higginsianum WT strain MAFF 305635 (O’Connell et al., 2004) (CY7444) was from the Ministry of Agriculture, Forest and Fisheries collection, Japan. All other C. higginsianum (Ch) strains used in this study are listed in the Supplementary Data S1.
Arabidopsis Lines
Arabidopsis thaliana Col-0 was obtained from the local seed collection. Mutant A. thaliana lines were congenic to Col-0 unless specifically noted. pen2-2 (Bednarek et al., 2009) was obtained from P. Schulze-Lefert (Cologne). All other mutant lines were obtained from L. Voll (Marburg). The cyp79b2/cyp79b3 double mutant (Zhao et al., 2002) was originally obtained from Tamara Gigolashvili. pad3-1 was originally obtained from Jane Glazebrook (Glazebrook et al., 1997). NahG (Lawton et al., 1995) was originally obtained from Yves Marco. sid2 was obtained from the NASC seed collection (N542603, SALK_042603) and screened for homozygosity. cerk1-2 (Miya et al., 2007) was obtained from the GABI-KAT seed collection (GK-096F09) and screened for homozygosity. The starch free p-pgm mutant line was originally obtained from Caroline Dean and was in the Ler-0 background.
Media and Cultivation Methods
Escherichia coli, A. tumefaciens, and C. higginsianum strains were cultured and transformed as described (Korn et al., 2015). For the selection of fungal transformants nourseothricin was used at 100 μg/ml. Hygromycin was used at 72 U/ml. Bialaphos-resistant transformants were selected on Czapek Dox plates supplemented with 2.64 g/l ammonium sulfate and were containing 20 μg/ml bialaphos. 100 μg/ml cefotaxime was added to transformation plates for selecting against A. tumefaciens. A. thaliana plants were grown for 2 weeks under short-day conditions (8 h of light, 22°C/16 h of dark, 20°C) and then transferred to growth chambers with 12 h light (22°C)/12 h dark (19°C) cycles (120 μE/m2s; 70% humidity). After 3 weeks (5 weeks in total), the plants were infected with C. higginsianum. For quantification of fungal biomass in Arabidopsis mutant lines, plants were grown for 1 additional week at 12 h light/12 h dark (6 weeks in total). Infections with C. higginsianum and induction of appressoria were performed as described (Schmidpeter et al., 2017). For all transformed C. higginsianum strains 2–4 independent original transformants were analyzed.
Standard Techniques and DNAs
DNA manipulations, PCR reactions and plasmid DNA isolations followed standard protocols as described (Korn et al., 2015). Plasmid constructions and oligonucleotide sequences are listed in the Supplementary Data S1.
Microscopy and Histochemical Staining
Histochemical samples or samples containing fluorescent reporter proteins were stained and analyzed by microscopy as described (Korn et al., 2015) with the following modifications. Quantification of trypan blue stained fungal infection structures was performed with 2 infected plants per strain at 3 dpi. Three leaves were analyzed per plant. At least 600 appressoria were counted per leaf. For the analysis of in planta appressoria formation, calcofluor staining was performed 18 h post infection. Leaf sections were cut out without the midrib and placed on the abaxial surface on ethanol soaked whatman paper (3 MM) to remove chlorophyll. After 60 min, the sections were placed on water soaked whatman paper and incubated for additional 20 min. Subsequently, sections were placed on glass slides and 100 μl calcofluor-white solution was added (0.1 mg/ml). Epifluorescence was detected with an excitation band pass filter (340–380 nm) and observed with a long pass filter (LP425 nm). Quantification was performed with 2 sections of 2 infected plants per strain. Detection of reactive oxygen species was performed as described (Korn et al., 2015) with the following modifications. Two leaves each of two infected A. thaliana plants per fungal strain were harvested after 4 days. The staining solution was supplemented with 0.05% (v/v) Tween 20. After 18 h incubation in the dark, the staining solution was replaced by destaining solution (ethanol:acetic acid:glycerol = 3:1:1) and boiled for 15 min at 95°C. After mounting with water, the leaves were viewed with bright-field microscopy.
Pulsed-Field Gel Electrophoresis (PFGE)
Pulsed-field gel electrophoresis was performed on a Hoefer HG 1000 HULA unit using 1% agarose (Agarose Pulsed-Field, Roth) and 0.5x TBE buffer. Gels were pre-run at 10°C for 2 h at 140 V without alternating the electric field, then for 36 h with alternating angles of 120°. The switch time for alternating the electric field was increased from 60 to 120 s over the course of the run. Yeast CHEF DNA size markers (BioRad) were used as standards. For documentation, gels were stained with ethidium bromide (0.5 mg/l) for 30 min. Plugs containing fungal conidia were prepared following published protocols (Maringele and Lydall, 2006). Freshly grown conidia were suspended in water containing 0.005% NaN3 and washed twice with H2O afterward. 1 × 107 conidia were collected by centrifugation, resuspended in 300 μl SCE buffer (1 M sorbitol, 100 mM sodium-citrate, 60 mM EDTA, pH 7, 60 mM 2-mercaptoethanol) and incubated at 37°C. After adding 50 μl enzyme solution [10 mg/ml Zymolyase T20 (Seikagaku) in SCE buffer], cells were gently mixed with 350 μl low melting point agarose (Bio&Sell) preheated to 50°C. After pipetting up and down twice, the mixture was spread on a casting form, generating 1.5 mm thick plugs. Plugs cut to the appropriate size were incubate in 400 μl SCE with 10 mg/ml Zymolyase T20 for 2 h at 37°C. After washing twice in PFGE buffer (10 mM pH 7.6, 50 mM EDTA) and once in PK buffer (100 mM EDTA, 0.2% sodium desoxycholate, 1% sodium N-laurylsarcosine, pH 8), plugs were incubated in 400 μl PK buffer containing 1 mg/ml Proteinase K at 50°C overnight (18 h). The following day, plugs were washed twice in PFGE buffer and stored in the buffer at 4°C until use.
DNA Sequencing
Genomic DNA sequencing was performed by a commercial provider using chromosomal DNA from Colletotrichum higginsianum strain MAFF 305635 and the mutant derivatives vir-49 and vir-51. High molecular weight DNA was prepared as described (Korn et al., 2015). Genomic sequencing was performed on an Illumina MiSeq platform using DNA fragmented to a size between 600 and 900 bp. The raw read files (.fastq accessions: SAMN08226879, SAMN08226880, SAMN08226881) underwent quality control analysis using FastQC.1 The quality-checked forward and reverse reads were trimmed separately with cutadapt (v1.14)2 with following parameters: -u 5 -u -5 -q 25 –trim-n –max-n 5 –minimum-length = 80 (forward reads) and -u 5 -u -25 -q 25 –trim-n –max-n 5 –minimum-length = 80 (reverse reads). Only read pairs where both the forward and reversed read fulfilled the criteria were kept for further analysis. Reads were aligned as single reads using Bowtie2 (v2.2.9) (Langmead and Salzberg, 2012) with default parameters to the reference assembly of strain IMI 349063 (accession: GCA_001672515.1) and deposited into the sequence read archive (SRA) at NCBI (accession number of bioproject: PRJNA427316). Raw library sizes were 13,696,134 for WT, 7,246,183 for vir-49 and 6,367,668 for vir-51. Read lengths were 266 bp for WT, 250 bp for vir-49, and 259 bp for vir-51. This corresponds to a coverage of approximately 72x for WT, 36x for vir-49 and 33x for vir-51 (Supplementary Tables S1, S2). For further analysis see the Supplementary Data S2.
Transformation Protocol for the Selection of Homologous Recombinants
Colletotrichum higginsianum strain MAFF 305635 was transformed as described (Korn et al., 2015) with the following modifications to identify homologous recombinants. Transformation plates were incubated for 16 days to induce sufficient conidiation on the primary transformation plates. Conidia were rinsed off the transformation plates with 3 ml H2O and further propagated on oatmeal-agar (OMA) plates for 1 week to generate a population of transformed conidia. The conidia were again rinsed off with water and diluted to 3 × 102 spores/ml. Hundred microliter of this dilution (n = 30 cells) was plated on minimal medium plates (20 μg/ml bialaphos and 100 μg/ml cefotaxime). Two days later, single colonies were picked and further propagated on 12-well OMA plates for 6 days. Subsequently, individual clones were analyzed on antibiotic containing media to identify transformants having undergone homologous recombination which should be resistant to bialaphos but sensitive to nourseothricin (Supplementary Figure S1). To verify homologous recombination, DNAs from such transformants were tested by PCR using primers diagnostic for integration in chromosome 11 (3′ homology region: CK3831/CK5328; 5′ homology region: CK4042/CK5327) or for chromosome 12 (3′ homology region: CK3831/CK5330; 5′ homology region: CK4042/CK5329).
Sorting of mCherry Negative Cells
Strains expressing cytosolic mCherry from a reporter gene inserted on either chromosome 11 (chr11::mCherry) or chromosome 12 (chr12::mCherry) were grown for 10 days on OMA plates under standard conditions and then kept at 8°C for 1 day. Prior to FACS analysis, cells were stained with the vital dye CellTrace CFSE (Thermo Fisher, C34570). 5 × 107 conidia were dissolved in 1 ml staining solution (1:1000 dilution in 1xPBS). After 20 min in the dark, 5 ml 1xPBS 1% BSA were added. After 5 additional minutes, conidia were spun down at 2000 rpm for 2 min and resuspended in 1 ml PBS. Prior to flow cytometry, the cell suspension was diluted 10-fold. Cell sorting was performed using a Beckman & Coulter Astrios platform. Sorted cells (mCherry negative conida) were plated out on PDA plates without antibiotics. After 2 days, single colonies were isolated and propagated on 12-well OMA plates for 7 days. To test for the presence of the particular chromosome, we isolated genomic DNA as described (Korn et al., 2015) and conducted PCR for genes located on chromosome 11 (CK5230/CK5231 and CK5192/CK5193) or chromosome 12 (CK5196/CK5197 and CK5304/CK5305), respectively. To validate the result, we performed pulsed-field gel electrophoresis as described above.
Assessment of Fungal Proliferation in Plant Tissue via Quantitative PCR
To assess fungal proliferation in Arabidopsis leaves, we performed quantitative PCR on fungal genomic DNA. For each fungal strain, we infected 5 plants. After 90–96 h, 5 samples per fungal strain were taken by pooling leaf punches from different plants, harvesting a total of 1.5 cm2 of infected leaf area per sample in liquid nitrogen. To extract total genomic DNA, we used the Qiagen DNeasy Kit. Quantification of fungal DNA was subsequently analyzed with an Agilent AriaMX Real-Time PCR System using Promega’s GoTaq® qPCR Master Mix and primers for the C. higginsianum TrpC gene (Ch TrpC fw and ChTrpC rev). All samples were measured as 3 technical replicates. The relative quantity of fungal DNA was calculated as described (Rieu and Powers, 2009) and normalized to the leaf sample area.
Extraction of Total RNA From Infected Plant Tissue
For the extraction of total RNA from infected plant tissue, three samples were harvested for each fungal strain and frozen in liquid nitrogen. For each sample three fully expanded leaves from independent plants were pooled and ground up in liquid nitrogen. For the isolation of total RNA, leaf tissue was frozen and ground in liquid nitrogen. The resulting powder was suspended in 3 ml of RNA isolation working solution (4 M guanidinium thiocyanate, 25 mM sodium acetate, 0.5% N-lauroylsarcosine, 0.7% mercaptoethanol, pH 7, freshly mixed with equal amounts of water saturated phenol). After thawing, the homogenate was transferred to two reaction tubes (2 ml) each containing 0.3 ml of chloroform:isoamylalcohol (24:1) and vortexed. After incubation for 30 min on ice, tubes were centrifuged for 15 min at 11.000 rpm and 4°C. The aqueous phase was extracted once more with phenol (0.7 ml phenol + 0.3 ml chloroform:isoamylalcohol) and once with chloroform:isoamylalcohol. Subsequently the volume of the upper aqueous phase was determined, 1/20 volume 1 M acetic acid and 1 volume of ethanol was added. After mixing, the pecipitated RNA was collected by centrifugation for 8 min at 13.000 rpm and 4°C. The supernatant was removed and the resulting pellet was washed with 1 ml 3 M sodium acetate pH 6. The two pellets of each sample were combined into one reaction tube by pipetting and pelleted for 10 min at 13.000 rpm and 4°C. The resulting pellet was washed twice with 80% ethanol. The RNA pellet was dried and resuspended in H2O at 60°C for 5 min. The concentration was estimated photometrically (NanoDrop, Thermo Fisher). For cDNA synthesis, 1 μg RNA was digested with DNAse I (DNAse I, Thermo Fisher) and subsequently used for the reverse transcriptase reaction (RevertAid Reverse Transcriptase, Thermo Fisher) as described (Korn et al., 2015). The cDNA was used for assessment of the plant defense reaction using DNA primers (Supplementary Data S1) specific for different pathogenesis-related genes (PR-genes).
Results
Virulence Mutants vir-49 and vir-51 Lack a Whole Chromosome and Arrest Early in the Infection Process
In a previous forward genetic screen for virulence mutants of C. higginsianum by insertional mutagenesis (Korn et al., 2015) we identified in total 75 mutants with diverse defects in the interaction with the model plant A. thaliana. Virulence mutants vir-49 and vir-51 showed a similar virulence phenotype characterized by their failure to effectively form necrotrophic secondary hyphae. Their ability to penetrate the A. thaliana host appeared normal, suggesting that they have some defect in switching from biotrophy to necrotrophic growth or fail to overcome host plant defense. Infection symptoms of Col-0 plants with these mutants (vir-49 and vir-51) can be seen in Figure 1A. Three days post infection (3 dpi), both strains showed normal penetration rates and formed apparently normal looking bulbous primary hyphae. In contrast, the wild type (WT) already started to form necrotrophic secondary hyphae at 3 dpi. This difference was even more pronounced after 4 days. While the wild type had completely switched to necrotrophy and established a massive hyphal network in the infected host tissue, mutants vir-49 and vir-51 remained arrested in the stage of primary hyphae. Occasionally, some hyphae underwent a morphologic switch but developed no further (Figure 1A, lowest panel – vir-49). We additionally observed intense trypan blue staining of plant cells at spots with fungal entry, suggesting necrosis of the infected cells. Macroscopically, the mutants showed only very mild symptoms and never the strong necrotic lesions typical for the wild type. We failed to genetically link any T-DNA insertion to the observed phenotype in the two mutants by genome walker PCR (Siebert et al., 1995; Korn et al., 2015) and therefore decided to perform whole genome sequencing of the mutants to identify the mutations responsible for the phenotype. DNAs from the mutants and the parental wild type (MAFF 305635) were sequenced on an Illumina MiSeq (Supplementary Data S2). After assembling the reads to the published reference genome (Dallery et al., 2017), we found to our surprise that both mutant strains were lacking most sequence data from chromosome 11. The number of reads from the mutant DNAs that were mapped to chromosome 11 was 10-fold lower than for wild type DNA and most reads mapped to transposable elements (Supplementary Tables S1, S2). This strongly suggested that the mutant phenotype was caused by complete loss of this chromosome rather than by DNA insertions (see Supplementary Data S2 and Supplementary Table S3 for details).
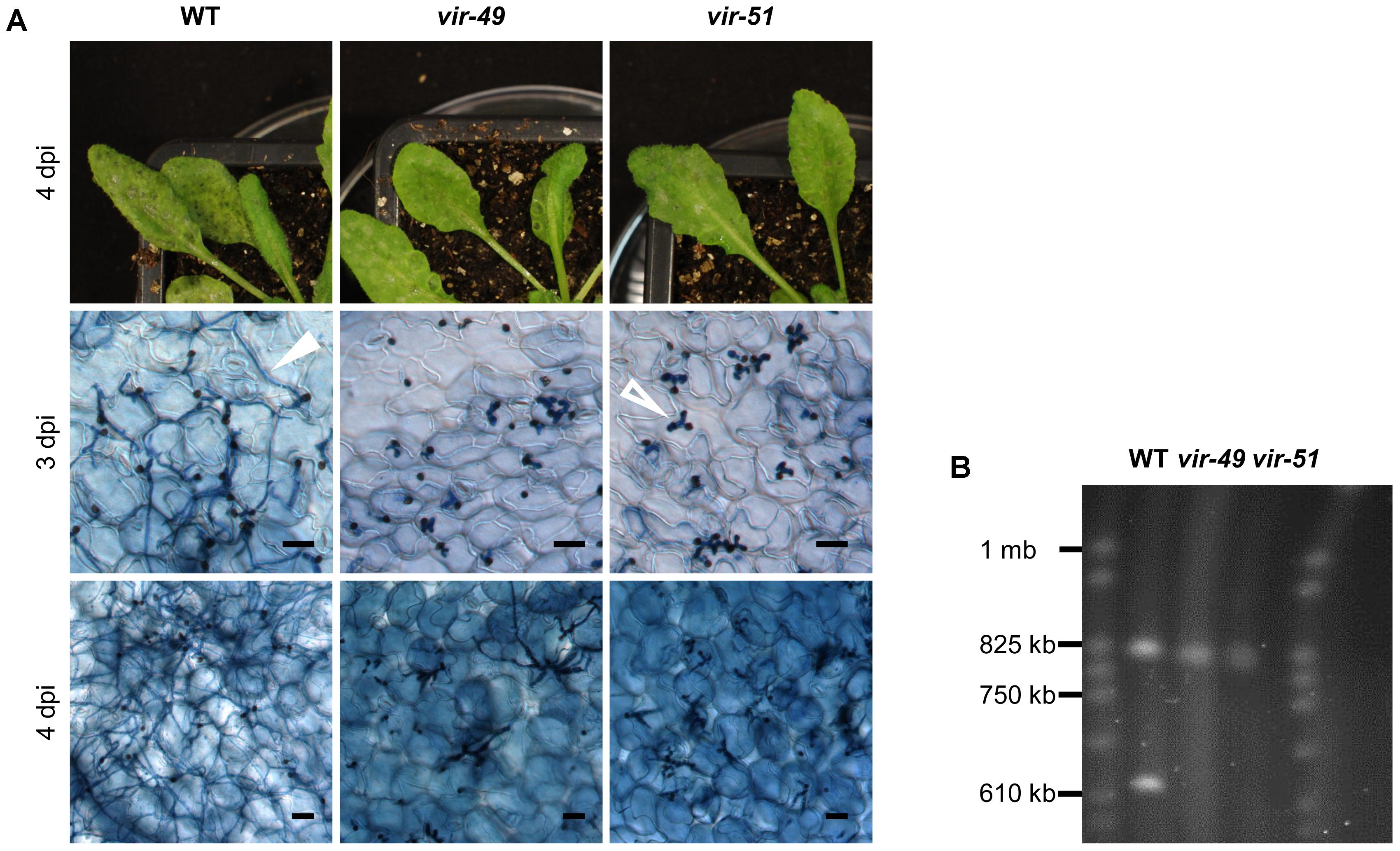
FIGURE 1. T-DNA insertion mutants vir-49 and vir-51 arrest early during infection. (A) A. thaliana Col-0 plants were infected with C. higginsianum WT and the mutant strains vir-49 (CY5976) and vir-51 (CY6030). The upper panel shows macroscopic symptoms at 4 dpi. The lower panels show microscopic images of trypan blue stained A. thaliana leaves at 3 and 4 dpi, respectively. Unfilled arrow: biotrophic primary hyphae; filled arrow: necrotrophic secondary hyphae. Scale bars = 20 μm. (B) Pulsed-field gel electrophoresis (PFGE) of small C. higginsianum chromosomes. C. higginsianum wild type (CY7444) and mutant conidia from strains vir-49 and vir-51 were analyzed on a Hoefer Hula Gel 1000 unit. Running conditions were chosen to separate chromosomes between 0.2 and 2 mb. S. cerevisiae chromosomes were used as a size marker (CHEF DNA Size Marker).
We tested this further by pulsed-field gel electrophoresis (PFGE) in order to validate the absence of this particular chromosome. C. higginsianum harbors 12 chromosomes. While chromosomes 1–10 have sizes between 3 and 6 mb, chromosomes 11 and 12 are smaller than 1 mb (Dallery et al., 2017). Using running conditions that allow to separate small chromosomes, the two small chromosomes were detectable in the WT strain (Figure 1B). Vir-49 and vir-51, however, were clearly lacking one of the small chromosomes with a size of approximately 620 kb. This matches the published size of chromosome 11 (646 kb) (Dallery et al., 2017). We also found that the other small chromosome was present in all three strains and showed a size of around 800 kb. In the published reference sequence of isolate IMI 349063, this small chromosome 12 only has a size of 600 kb. This also demonstrated that the MAFF 305635 isolate used for our studies contains some genomic differences to the reference isolate (Supplementary Data S2). While genome sequencing and PFGE analysis clearly demonstrated the absence of chromosome 11 in the two analyzed mutants, this did not prove that chromosomal loss is responsible for the observed virulence phenotype in vir-49 and vir-51. We therefore used wild type C. higginsianum strains to isolate spontaneous variants that were nullisomic for chromosome 11.
A Chromosome Loss Assay for Chromosome 11
To associate the absence of chromosome 11 with the phenotype of the analyzed vir mutants, we established a chromosome loss assay for the generation of congenic strains with and without chromosome 11. We designed an mCherry expression cassette conferring bialaphos resistance flanked by regions of the locus CH63R_14406 located on the left arm of chromosome 11 (Figure 2A). Upon homologous recombination, this locus is replaced with an mCherry gene expressed from the ChTef1 promotor. A second selectable marker (nourseothricin resistance) outside the region of homology allowed to distinguish ectopic insertions from homologous recombinants after transformation of C. higginsianum WT (CY 7444) by Agrobacterium tumefaciens mediated transformation (ATMT) (Supplementary Data S1). Approximately 5 percent of the bialaphos-resistant transformants were negative for the nourseothricin marker (Supplementary Figure S1) and were also tested positive for homologous recombination by PCR on single colonies (not shown). The resulting chr11::mCherry strains showed strong red fluorescence in the conidial cytoplasm (Figure 2A).
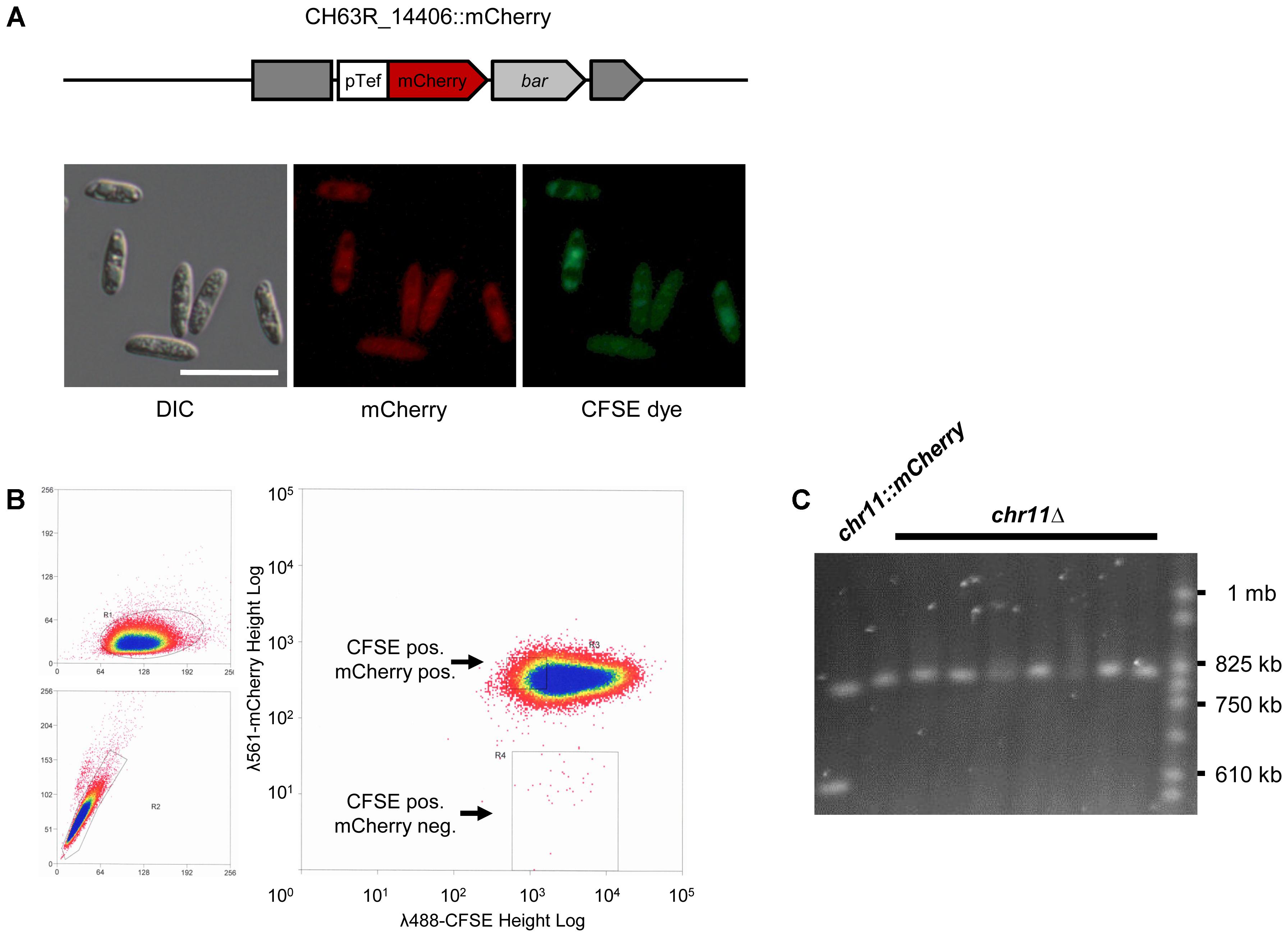
FIGURE 2. A chromosome loss assay using a targeted insertion of an mCherry gene. (A) Schematic representation of the CH63R_14406 locus on chromosome 11 after insertion of an mCherry reporter cassette by homologous recombination (strain CY7466). The mCherry gene was driven by a strong promotor (pTef). The bar gene conferring bialaphos resistance was used as a selectable marker. The dark gray blocks represent the disrupted CH63R_14406 locus. The lower images show the signals of mCherry in the conidial cytoplasm and of the fluorescent cell tracer CFSE in conidia of this strain. DIC, differential interference contrast, scale bar = 20 μm. (B) Flow cytometry of chr11::mCherry strain (CY7466) used to sort for mCherry negative conidia. We found a loss rate of 3 × 10-4 for chromosome 11. Left: forward and side scatter of cells. FSC-Height/ SSC-Height (top) and SSC-Height/SSC-Area (bottom). Right: fluorescent signals of CFSE at λ = 488 nm (x-axis) and mCherry at λ = 561 (y-axis). Scales are logarithmic and were adjusted to fit the range of fluorescence in the sample. The region gated for isolating mCherry neg. cells is shown as a rectangle. (C) Pulsed-field gel electrophoresis of chromosomes from parental chr11::mCherry conidia and from randomly chosen single colonies of sorted mCherry negative cells. Yeast chromosomes were used as a size marker (CHEF DNA Size Marker).
The spontaneous loss of chromosome 11 in the tagged strain should be accompanied with the loss of mCherry fluorescence. We therefore used flow cytometry with subsequent cell sorting to isolate individual mCherry negative conidia. For gating the correct population, cells were counterstained with the green fluorescent vital stain CFSE (Figure 2A). FACS analysis showed that besides a large, homogenous population of double stained conidia, a very small fraction of CFSE positive, mCherry negative cells could be identified (Figure 2B). These were isolated by cell sorting and plated for single colonies. PCR screening showed that approximately 90 percent of mCherry negative clones had lost genes located on chromosome 11 (CH63R_14508 and CH63R_14384) (not shown). The complete loss of chromosome 11 in these isolates (chr11Δ) was validated by PFGE for randomly picked colonies (Figure 2C) and by PCR analysis of five different loci separated by approximately 100 kb on chromosome 11 (Supplementary Figure S1). Interestingly, loss of chromosome 11 could be detected without extensive subculturing of the original chr11::mCherry strains.
Chromosome 11 of C. higginsianum Is Essential for Virulence on A. thaliana but Does Not Affect Vegetative Growth
To validate the connection between loss of chromosome 11 and loss of virulence, we infected A. thaliana Col-0 plants with variants that have lost chromosome 11 (chr11Δ), the corresponding parental strain chr11::mCherry and C. higginsianum wild type (Figure 3A). There were no obvious differences between infections with either the wild type or the tagged chr11::mCherry strain. At 3 dpi both strains had switched to necrotrophic growth. At 4 dpi, the infected leaves were abounded with secondary hyphae and the plants showed strong macroscopic symptoms. The chr11Δ variant, however, showed a defect closely resembling the virulence phenotype originally observed for vir-49 and vir-51 mutants (Figure 3A). Three days after infection the fungus had penetrated the plant cells and established normal looking primary hyphae but showed no visible switch in hyphal morphology. Four days post infection almost all hyphae remained stuck as large bulbous hyphae. Only few (<3%) primary hyphae of the chr11Δ strain produced filamentous secondary hyphae. Identical results were found for several independently isolated chr11Δ strains (see below and not shown).
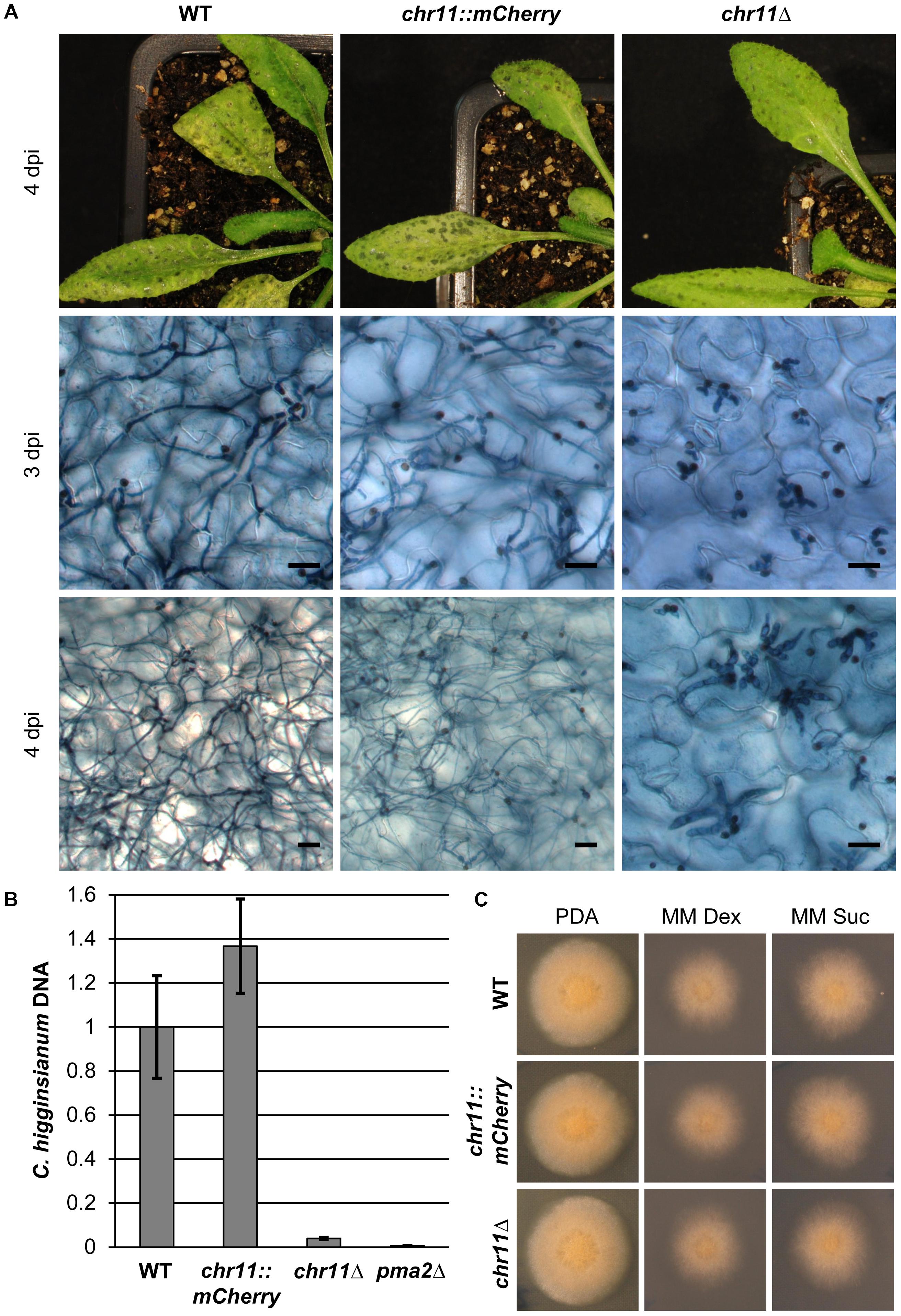
FIGURE 3. Strains lacking chromosome 11 arrest in the post penetration stage of infection. (A) A. thaliana Col-0 plants were infected with C. higginsianum WT, a strain lacking chromosome 11 (CY7548) (chr11Δ) and the respective mCherry tagged parental strain (CY7466) (chr11::mCherry). The upper panel shows macroscopic symptoms at 4 dpi. The two lower panels show microscopic images of trypan blue stained A. thaliana leaves at 3 and 4 dpi, respectively. Scale bars = 20 μm. (B) Relative amount of fungal DNA from infected host tissue at 4 dpi. A pma2Δ mutant strain (CY6153) which only forms appressoria and does not penetrate the plant was used as additional control. DNA amount was assessed via qPCR, corrected for leaf area and normalized to WT infection. Samples of five biological replicates per strain were taken after 90 h. Error bars show the standard error for each data set. (C) Vegetative growth of the respective strains on potato dextrose agar (PDA) and minimal media with dextrose (MM Dex) or sucrose (MM Suc).
We performed quantitative PCR on fungal genomic DNA of infected leaves approximately 4 days post infection to quantify fungal proliferation in planta (Figure 3B). As expected for an aborted infection, leaves infected with chr11Δ showed 25 times less fungal DNA than leaves infected with the wild type. In order to further evaluate the limited proliferation of chr11Δ cells, we compared it to a penetration defective C. higginsianum mutant lacking a virulence specific proton pump (pma2Δ) that replicates twice during appressoria formation but never enters the host cell (Korn et al., 2015). The chr11Δ mutant generated approximately 6.5 times more DNA during infection compared to pma2Δ. The wild type, however, generated approximately 160 times more DNA in the host tissue than the penetration mutant (Figure 3B). In summary, our data clearly demonstrate that infection was aborted early during infection after few replication cycles in the absence of chromosome 11.
To more carefully investigate if the chr11Δ strains show additional phenotypes we analyzed the formation of appressoria in vitro, i.e., on petri dishes and in planta but did not see any difference between chr11Δ and the WT C. higginsianum strain. Importantly, the number of appressoria that successfully penetrated the plant cells during infection was unaltered in the chr11Δ mutants (Supplementary Figure S2). In addition, we noticed some atypical lesions on leaves 4 days after infection which stained heavily with trypan blue which may indicate host cell necrosis, i.e., defense responses (Supplementary Figure S3). Such areas were absent in WT infections. Moreover, we observed that the staining of large, bulbous hyphae of chr11Δ strains at 4 dpi was less intense and the hyphae were more vacuolated compared to the bulbous hyphae after 3 days (Supplementary Figure S3B). This may suggest that the hyphae are no longer viable at this point of the infection.
Finally, the loss of chromosome 11 had no visible effect on the vegetative fitness or colony morphology on rich or minimal media (Figure 3C). The functions of genes on chromosome 11 therefore appear to be confined to specific roles during the infection process.
Altered Host Response in the Absence of Chromosome 11
We observed atypical lesions on A. thaliana leaves infected with chr11Δ strains. These areas were macroscopically visible in stained samples (Figure 4A) and distributed on the whole leaf beneath spots with signs of fungal infection. As these areas contained very little fungal material, they most probably correspond to dead Arabidopsis cells which also get stained with trypan blue (Keogh et al., 1980; Pogany et al., 2009). The lesions are reminiscent of a hypersensitive response of A. thaliana which may fail to be suppressed by the pathogen when genes from chromosome 11 are missing.
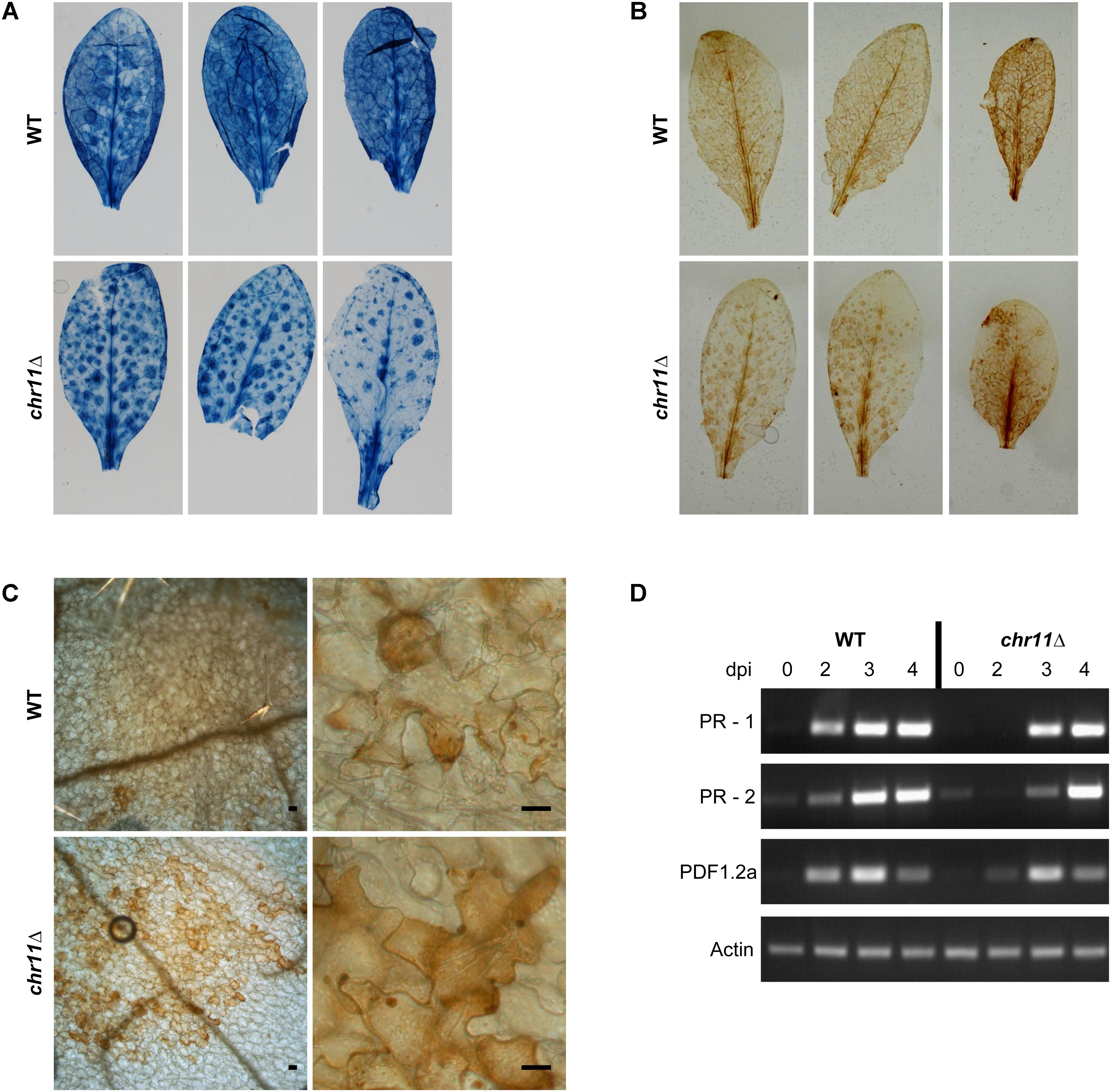
FIGURE 4. Plant response to C. higginsianum lacking chromosome 11. (A) A. thaliana Col-0 plants were infected with C. higginsianum WT (CY7444) and chr11Δ mutant (CY7548). One representative leaf each from 3 infected plants per strain were harvested at 4 dpi and stained with trypan blue to stain fungal cells and dead plant tissue. After destaining, macroscopic photos were taken. (B) Macroscopic images of 3,3′-diaminobenzidine (DAB) stained leaves at 4 dpi for visualization of reactive oxygen species (ROS). (C) Microscopic images of representative DAB stained leaves at 4 dpi. Scale bars = 20 μm. (D) RT-PCR of plant PR genes at different time points during the infection of chr11Δ and the C. higginsianum wild type.
To characterize this further, we performed ROS staining using 3,3′-diaminobenzidine (Figures 4B,C). At 4 dpi, we found the macroscopic DAB staining of leaves infected with chr11Δ to show the same pattern as the staining of dead plant cells with trypan blue. Microscopically, we observed clusters of cells with stronger DAB staining in the chr11Δ infection which could be assigned to the macroscopically visible areas. In contrast, leaves infected with C. higginsianum wild type were already fully colonized by the fungus at this time point with many dead plant cells and less DAB staining. The observed and altered host response against chr11Δ suggested that the inability of chr11Δ mutants to switch to necrotic growth is due to successful plant defense rather than a consequence of a defective fungal regulation. We also analyzed the expression of plant pathogenesis-related genes by RT-PCR at different time points during the infection with chr11Δ and the C. higginsianum wild type (Figure 4D). Both fungal strains induced expression of the salicylic acid induced genes PR-1 and PR-2 (Penninckx et al., 1996). Also the plant defensin PDF1.2a which is induced by the jasmonic acid pathway (Uknes et al., 1992) was induced by both strains. However, the three genes appear to be induced earlier in the wild type infection.
Plant Tryptophan P450 Monooxygenases cyp79b2 and cyp79b3 Are Required to Prevent Infection by C. higginsianum Mutants Lacking Chromosome 11
Having observed an altered host reaction against strains lacking chromosome 11 and C. higginsianum wild type, we tested a collection of A. thaliana mutants with defects in different layers of plant defense. If the phenotype of chr11Δ strains were due to the inability to suppress plant defense, we hypothesized that such strains should regain virulence in the absence of critical plant defense pathways.
We therefore infected the A. thaliana double mutant cyp79b2/cyp79b3 (Figure 5). This hypersusceptible mutant cannot convert tryptophan to indole-3-acetaldoxime (IAOX) and is therefore lacking a number of defense related metabolic products like indole glucosinolates and camalexin. This mutant is highly susceptible to various fungal pathogens including Colletotrichum species (Bednarek et al., 2009; Sanchez-Vallet et al., 2010; Hiruma et al., 2013). Already 3 days post infection the C. higginsianum WT strain produced necrotrophic hyphae in large areas. Four days post infection, the tissue was massively colonized by the WT and acervuli were detected. The chr11Δ strain not only established primary hyphae after 3 days, but at 4 dpi the strain also colonized large parts of the host tissue with necrotrophic secondary hyphae, indicating that chr11Δ strains can be virulent on A. thaliana when plant defense is impaired (Figure 5A and Supplementary Figure S4).
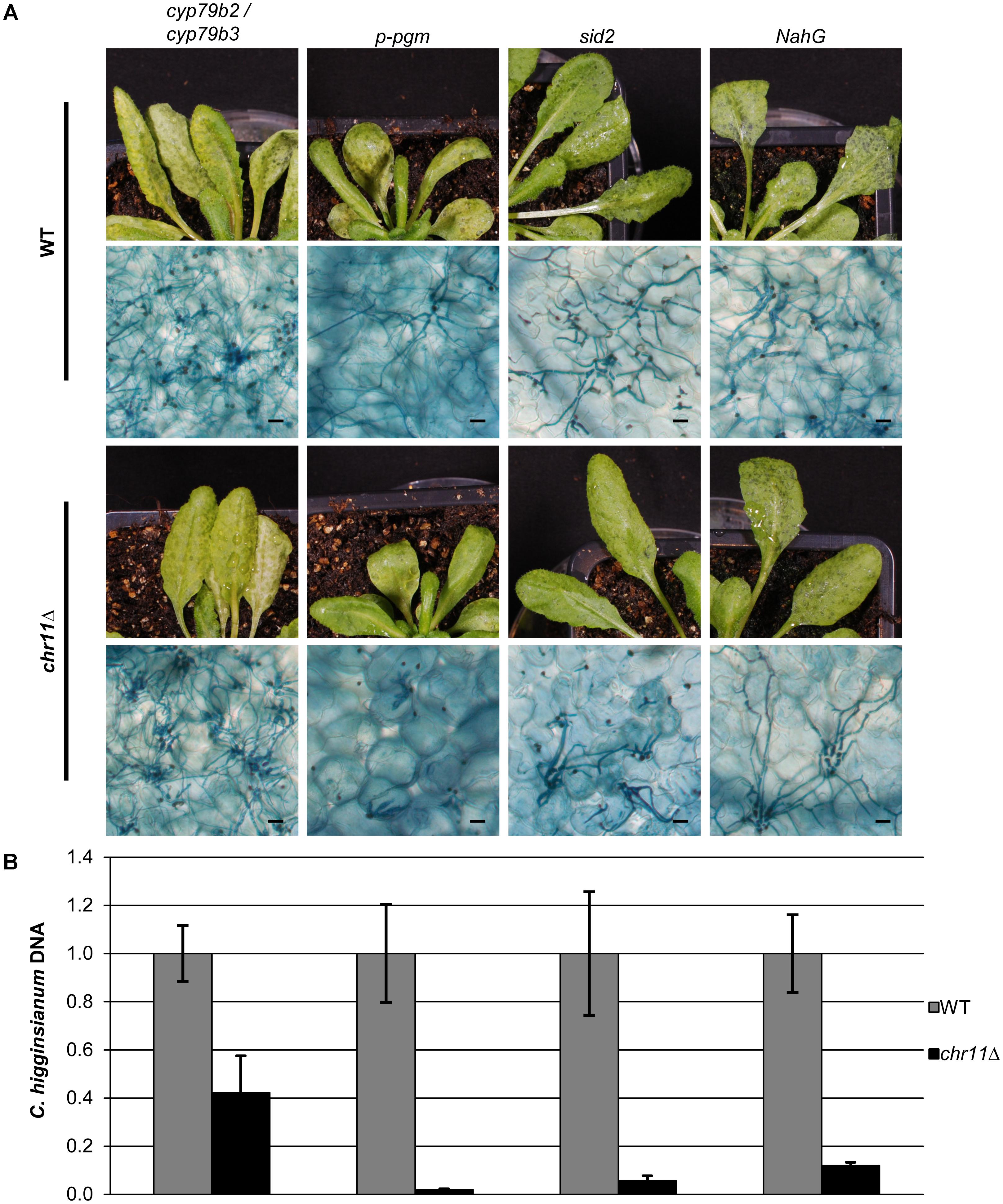
FIGURE 5. Chromosome 11 is required to suppress post penetration plant defense. (A) Macroscopic and microscopic symptoms of WT (upper panel) and chr11Δ (lower panel) infection of the analyzed mutant lines (p-pgm, sid2, NahG, cyp79b2/cyp79b3) after 90 hpi. Scale bars = 20 μm. (B) C. higginsianum WT and chr11Δ strains were used to infect different A. thaliana mutants. Relative amount of fungal DNA from infected A. thaliana lines (p-pgm, sid2, NahG, cyp79b2/cyp79b3) was analyzed at 90 hpi. DNA amount was assessed via qPCR, corrected for leaf area and normalized to Ch WT infection of each mutant line. Samples of five biological replicates per strain were taken. Error bars show the standard error for each data set.
To exclude that the colonization of A. thaliana cyp79b2/cyp79b3 double mutants by chr11Δ was due to pleiotropic defects of the plants, we tested additional A. thaliana mutants and quantified the virulence of C. higginsianum wild type and the chr11Δ strain via quantitative PCR. Infections of transgenic NahG plants, which do not accumulate salicylic acid (Delaney et al., 1994), sid2 plants which are defective for salicylic acid signaling and infections of the starch free p-pgm mutant which is more susceptible toward C. higginsianum were compared with cyp79b2/cyp79b3 mutant plants (Figure 5). While the chr11Δ strain colonized the cyp79b2/cyp79b3 successfully and generated more than 40% of the fungal DNA observed in infections with C. higginsianum wild type, the chr11Δ strain was unable to establish secondary hyphae on the p-pgm mutant and generated only approx. 2% of the fungal biomass observed for the C. higginsianum wild type (Figure 5B). Given the fact that the p-pgm line is generally more susceptible to C. higginsianum but refractory to infection with chr11Δ, this strongly suggests that the colonization of cyp79b2 / cyp79b3 double mutants by chr11Δ is due to a specific defense defect and not due to pleiotropic effects. On sid2 mutants and the NahG lines the C. higginsianum strain lacking chromosome 11 showed very limited proliferation compared to WT C. higginsianum (6–12% of WT). Consistent with that, only a few chr11Δ primary hyphae switched to necrotrophic hyphae and their growth remained locally restricted (Figure 5A). This may indicate that genes on chromosome 11 also effect SA-mediated defense.
We also analyzed additional defense mutants (Supplementary Figure S4). The cerk1-2 mutant, defective for a pattern recognition receptor (PRR) sensing chitin (Miya et al., 2007), showed no increased susceptibility when infected with the chr11Δ strain. The same was observed for the preinvasion defense defective mutant pen2-2 (Bednarek et al., 2009). In addition, we also analyzed the camalexin defective pad3 mutant (Narusaka et al., 2004) but did not observe significant colonization by the chr11Δ strain.
Chromosome 12 Is Not Required for Virulence on A. thaliana
Colletotrichum higginsianum’s two small chromosomes 11 and 12 have similar properties. Both are very small, harbor many transposable elements and encode potential effector genes (Dallery et al., 2017). We therefore also generated strains that had spontaneously lost chromosome 12, using the chromosome loss assay described above but with an mCherry gene integrated at the CH63R_14534 locus from chromosome 12 (Figure 6 and Supplementary Figure S5). Similar to the chr11Δ strains, chr12Δ strains were readily obtained and were not impaired in growth on synthetic media (Figure 6C).
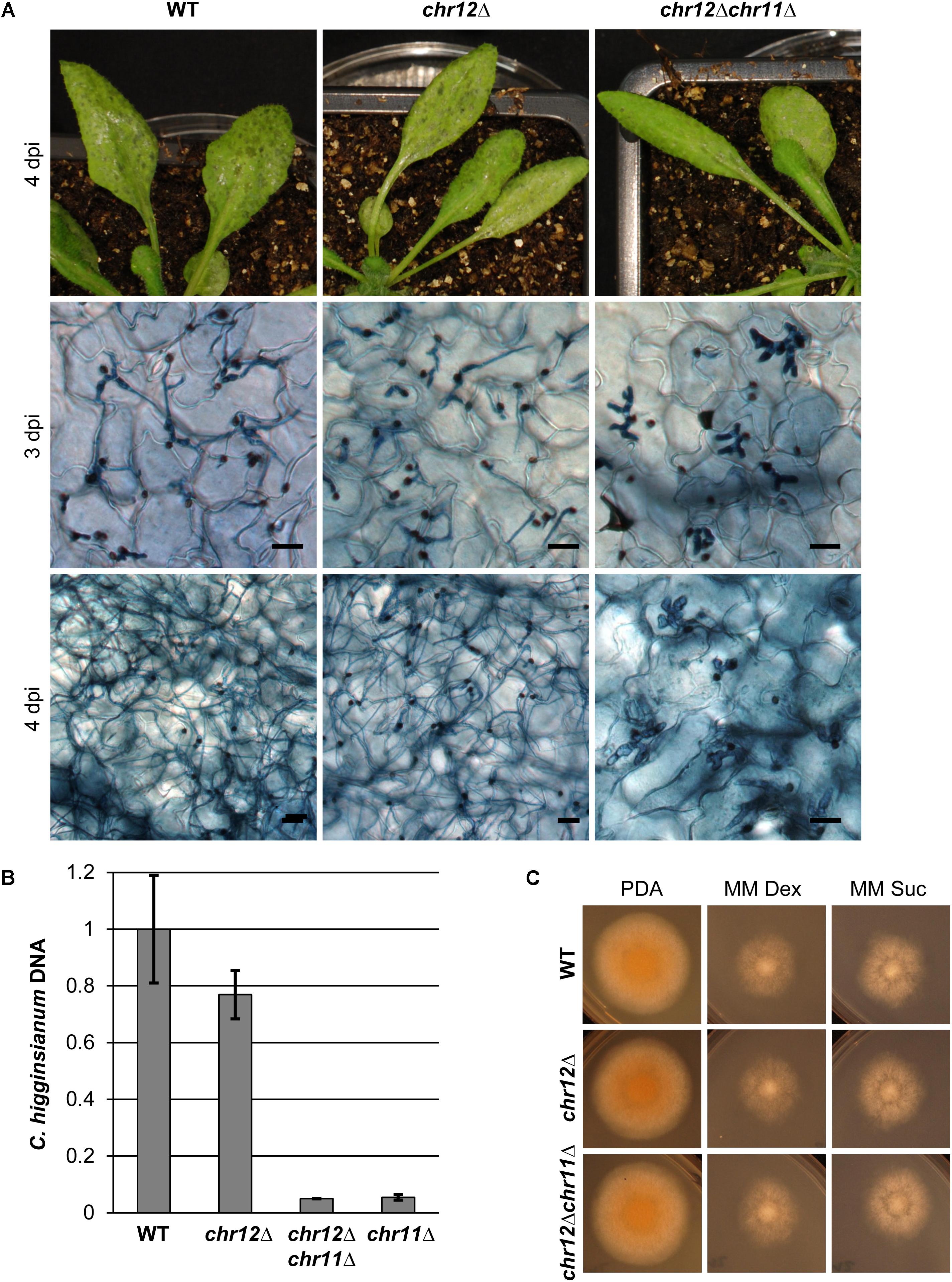
FIGURE 6. Chromosome 12 is not required for virulence on A. thaliana. (A) A. thaliana Col-0 plants were infected with C. higginsianum WT, a strain lacking chromosome 12 (CY7554) (chr12Δ) and a strain lacking both small chromosomes (CY7589) (chr12Δchr11Δ). The upper panel shows macroscopic symptoms at 4 dpi. The two lower panels show microscopic images of trypan blue stained A. thaliana leaves at 3 and 4 dpi, respectively. Scale bars = 20 μm. (B) Relative amount of fungal DNA from infected host tissue at 4 dpi. The DNA amount was assessed via qPCR, corrected for leaf area and normalized to WT infection. Samples of four biological replicates per strain were taken at 4 dpi. Error bars show the standard error for each data set. (C) Vegetative growth of the respective strains on potato dextrose agar (PDA) and minimal media with dextrose or sucrose.
In contrast to chr11Δ strains, however, we did not detect any obvious differences in virulence between C. higginsianum wild type and the chromosome 12 loss mutant chr12Δ (Figure 6A). Both established biotrophic primary hyphae in the host tissue at 3 dpi and showed a morphological switch to necrotrophy in some areas. Four days after infection, leaves showed heavy macroscopic symptoms in both cases. Microscopically, colonization with necrotrophic secondary hyphae was similar for both strains.
We also generated a double mutant lacking both small chromosomes by using chr12Δ as the parental strain for the tagging of chromosome 11 and for the subsequent isolation of mCherry negative variants lacking both chromosomes (Supplementary Figure S5). Again, such chr12Δchr11Δ strains were easily obtained by cell sorting and their growth on synthetic media was comparable to WT (Figure 6C). In virulence assays, the phenotype of the double mutant resembled the virulence defects of the chr11Δ in all regards (Figure 6A), suggesting that the additional loss of genes on chromosome 12 does not enhance the virulence defect of C. higginsianum lacking chromosome 11. We also performed quantitative PCR on fungal genomic DNA of infected leaves 4 days post infection to assess the fungal proliferation in planta (Figure 6B). As observed in microscopy, there was no significant difference between the wild type and the chr12Δ mutant and no enhanced phenotype in the double mutant (chr12Δchr11Δ) compared to chr11Δ.
Differences Between C. higginsianum Strains
As the karyotype of the MAFF 305635 strain showed differences to strain IMI 349063 regarding the size of chromosomes 11 and 12 (see above), we compared the entire genome sequences by mapping MiSeq illumina reads of the MAFF 305635 WT strain to the reference genome of IMI 349063 (Supplementary Data S2). The PFGE analysis (Figure 1 and data not shown) of the MAFF 305635 isolate showed that chromosome 12 was ∼200 kb larger than reported for the reference strain, while chromosome 11 lacked roughly 30–50 kb relative to the reference sequence from IMI 349063. The sequence alignment to the reference confirmed this difference as no reads from the MAFF 305635 strain mapped to annotated genes in the region from position 355,533 to position 406,259 corresponding to a 50 kb region between gene CH63R_14468 and gene CH63R_14477 of the reference genome (Supplementary Data S2). Some small additional regions in which the two strains differ were identified by looking for segments of low coverage in the MAFF 305635 sequencing (Supplementary Tables S2, S3 and Supplementary Data S2). This analysis identified genes potentially lacking in MAFF 305635, while the absence of a complete genome assembly for the MAFF 305635 strain did not allow to identify genes specific to MAFF 305635.
In order to evaluate how divergent the two C. higginsianum isolates are, we analyzed differences in the coding regions between the reference assembly and the MAFF 305635 isolate using the variant caller SnpEfff (Supplementary Data S2 and Supplementary Table S4). Most of the genome was very similar to the reference and showed about 1 SNP per 1 kb of coding region (Table 1 and Supplementary Table S4). This variation is very similar to the variations found between pairs different isolates of the human fungal pathogen Aspergillus fumigatus (1 SNP/kb coding) (Takahashi-Nakaguchi et al., 2015).
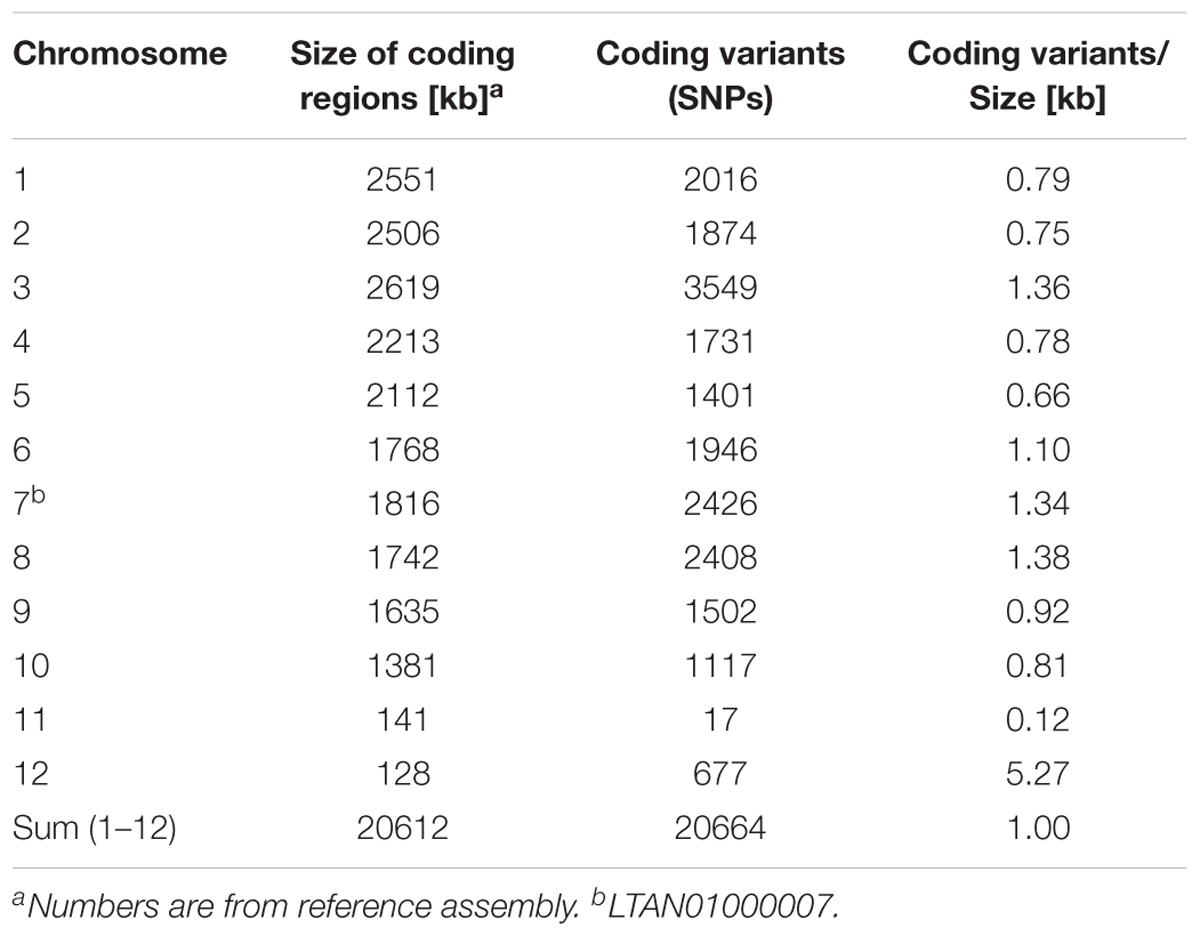
TABLE 1. Sequence variations in coding regions of MAFF 305635 and IMI 349063.
Two large regions in the genome showed notable differences to this picture. One region corresponds to chromosome 11 and was more conserved between IMI 349063 and MAFF 305635 with a total of only 17 SNPs in all coding regions between the strains (0.12 SNPs/kb). The other region corresponds to chromosome 12 which showed approximately 5 SNPs per 1 kb coding region. This may indicate that chromosome 11 is more conserved than the rest of the genome and may have only recently been acquired, while genes on chromosome 12 may have undergone significant, possibly, strain specific changes in the two isolates.
Discussion
Dispensable chromosomes have been described in diverse organisms. In some cases, in particular in sexual species, it has been argued that they may selectively provide advantages and/or adaptation to special conditions (Jones, 1991). In fungi, dispensable chromosomes have been reported in several species, in particular in Fusarium solani (Miao et al., 1991). Later lineage-specific (LS) chromosomes were found in other pathogenic fungi, for instance Alternaria alternata (Hatta et al., 2002) or in Fusarium oxysporum (Ma et al., 2010; Vlaardingerbroek et al., 2016a,b). Dispensable chromosomes often differ from the core chromosomes of the particular organisms in their size, gene content or sequence characteristics (Moller and Stukenbrock, 2017). In Colletotrichum species dispensable chromosomes have been noticed in C. gloeosporioides (Masel et al., 1996) and it was reported that C. higginsianum and C. graminicola harbor small mini chromosomes (Taga et al., 2015; Dallery et al., 2017) but no roles in virulence have been reported for these small chromosomes.
We here show that the loss of a dispensable chromosome in C. higginsianum has severe consequences on its ability to cause disease while having no effects on vegetative growth. The presence of this chromosome seems to provide a specific advantage for C. higginsianum as a pathogen. In particular, it enables the fungus to suppress post invasion plant defense mechanisms. We also found that a second dispensable mini chromosome can be lost during saprophytic growth without any effect on growth or pathogenicity. The two dispensable chromosomes are much smaller than the other 10 chromosomes which range from 3 to 6 mb (O’Connell et al., 2012). Chromosomes 11 and 12 account for only 3% of the genome, covering approximately 1% of annotated genes but are enriched for genes encoding putative secreted effector proteins (Dallery et al., 2017).
How Often Does C. higginsianum Lose Mini Chromosomes?
Our chromosome loss assay was based on the spontaneous loss of tagged chromosomes 11 or 12, respectively, during mitotic division and growth on agar plates. Vegetatively grown cultures of chr11::mCherry and chr12::mCherry produce conidia lacking the respective chromosome at a frequency of more than 1 × 10-4. This rate is remarkably high compared to the numbers reported for Fusarium where the loss rate of a lineage specific LS chromosome using a similar experimental approach was approximately 3 × 10-5 (Ma et al., 2010; Vlaardingerbroek et al., 2016b). This frequency may be directly related to chromosome size as chromosomes 11 and 12 from C. higginsianum are much smaller than the LS chromosomes of Fusarium. Chromosome loss can only be observed and studied when the respective chromosome is not essential or important for growth. Therefore, documented frequencies of chromosome loss usually are rare. But there are numbers for mitotic stability of centromeric plasmids and chromosomes in yeast. Yeast plasmids with replication origins and centromeres are lost at very high frequencies of up to 1 × 10-2. For linear yeast chromosomes, the segregation frequencies appear to be size dependent with artificial mini chromosomes (less than 200 kb) being lost at a rate of about 1 × 10-3 while larger chromosomes (e.g., ChrVII, 1091 kb) are 100 times more stable (Hartwell et al., 1982; Hieter et al., 1985). In light of these numbers, a specific active mechanism may not be required to explain the high frequency of mini chromosome loss in C. higginsianum. This may also explain why we found loss of chromosome 11 in two independently isolated vir mutants. The high spontaneous rate of chromosome loss for C. higginsianum mini chromosomes further suggests that chromosome loss was not dependent on the transformation by ATMT during the original insertional mutagenesis (Korn et al., 2015). Chromosomes that are not essential can be lost by three possible mechanisms when checkpoint mechanisms are overridden. First, a late replicating chromosome with few origins of replication may become left behind. Second, a failure in chromosome cohesion during mitosis might lead to non-disjunction. Finally, meiotic cohesion problems may lead to chromosome loss. All of these mechanisms may be more error prone when chromosomes are small, i.e., have fewer replication origins (ARS sequences) and fewer sites of cohesion or meiotic chiasmata. Error prone non-disjunction may further be enhanced in systems where the number of microtubules is limiting as was suggested for yeast (Hieter et al., 1985). In fact, yeast artificial chromosomes with sizes close to chromosomes 11 and 12 from C. higginsianum have been reported to be unstable. It is, however, also quite possible that the rate of chromosome loss we observed is neither specific to mini chromosomes nor a selected feature of C. higginsianum but occurs in all systems, yet without being noticed as the resulting cells may simply die or may have no obvious phenotype.
Why Should Chromosomes 11 and 12 Be Mitotically Instable?
If the loss of mini chromosomes were harmful one might expect such small chromosomes to fuse with others in order to gain stability. It is therefore tempting to speculate that, rather than being a problem, their instability is a feature that is selected for by small chromosome size. Chromosome loss may be the simplest mechanism to escape those host specific defense mechanisms that depend on the presence of corresponding avirulence genes in the pathogen. Loss of function mutations in pathogen effector genes have indeed been shown to be a key mechanism to escape plant defense (Jones and Dangl, 2006). Since C. higginsianum belongs to fungi that reproduce mostly asexual, the loss of mini chromosomes may be a dead end unless they are regained by cell fusion, conidial anastomosis or other mechanisms (Seidl and Thomma, 2014). Indeed, such events have been described in the genus of Colletotrichum. Conidial anastomosis and parasexual recombination was shown for Colletotrichum lindemuthianum (Ishikawa et al., 2012) and the transfer of supernumerary chromosomes between two biotypes was observed in Colletotrichum gloeosporioides (He et al., 1998). The loss and parasexual gain of dispensable mini chromosomes may be an alternative resource of expanding the genetic repertoire quite similar to bacterial episomal plasmids. Such recent gain of extra chromosomes may be a means for the fast adaptation to changing environmental situations and for the adaptation to changing hosts and hosts evolving novel resistance mechanisms.
Sequence Variations in Chromosomes 11 and 12
The evolutionary history of individual genomic regions is thought to be reflected in coding sequence variations as well as genomic and genetic rearrangements (Dong et al., 2015; Moller and Stukenbrock, 2017). Most of the core genome of C. higginsianum shows a variation rate in coding regions of 1 SNP/kb between the two different C. higgisianum isolates analyzed. This is similar to the variation seen when different Verticillium dahliae strains were compared (de Jonge et al., 2013; Faino et al., 2016) and comparable to the situation for different genetically isolated yeast isolates (Peter et al., 2018). In contrast, the conservation between coding regions on chromosome 11 is well above average (Table 1), suggesting that the encoded genes are subject to purifying selection. It is striking that genes on chromosome 11 show such little variability, since the two compared isolates originate from very different locations. The reference isolate IMI 349063 was isolated in Trinidad and Tobago while MAFF 305635 used here originates from Japan. For genes on chromosome 12 the situation is exactly the opposite, significant variation in their coding regions is observed and is 5 times higher than for the rest of the genome (Table 1). This may indicate that chromosome 12 has undergone recent changes, possibly reflecting some adaptation to the different environments. We also noticed that according to the data reported for gene expression in planta (Dallery et al., 2017) genes from chromosome 12 contribute only very little to fungal gene expression during infection of A. thaliana. It is also noteworthy that chromosome 11 may be variable with regard to gene number because a region of 50 kb from the reference appeared to be absent from the isolate used in this study. This region encompasses nine genes (CH63R_14468 to CH63R_14477) including five genes annotated as hypothetical protein and two genes encoding C14 peptidases.
Loss of Chromosome 11 Leads to Increased Success of Post Penetration Resistance
Our results clearly showed that the loss of chromosome 11 leads to a specific defect in suppression of plant host response mechanisms, because typical signs of necrotrophic colonization are restored when tryptophan derived secondary metabolites are missing. We excluded any indirect pleiotropic effect by showing that starch free A. thaliana p-pgm plants in the Ler-0 background which are much more susceptible to C. higginsianum infection (Engelsdorf et al., 2013) remain fully resistant to chr11Δ strains.
Which specific plant defense mechanisms are no longer suppressed when chromosome 11 is missing? A. thaliana uses different lines of defense to provide resistance toward hemibiotrophic fungi. Most important are: Salicylic acid (SA) and Jasmonic acid (JA) induced by PAMP (pathogen-associated molecular pattern) recognition (Narusaka et al., 2004; Liu et al., 2007), the phytoalexin camalexin (Narusaka et al., 2004) and tryptophan derived secondary metabolites including indole glucosinolates (Hiruma et al., 2013).
We analyzed a subset of these pathways by infecting the respective Arabidopsis defense mutants. We found no effect on the ability of chr11Δ to colonize Arabidopsis when the chitin PAMP receptor CERK1 is missing, which fits to the finding that CERK1 plays a minor role in defense against C. higginsianum (Miya et al., 2007). Most likely, other chitin receptors or other PAMPs are more important for C. higginsianum detection. However, the existence of chitin masking LysM domain proteins in C. higginsianum suggests that chitin is important (Takahara et al., 2016; Dallery et al., 2017). In addition, chitin deacetylation and dynamic changes in cell wall components may be important to hamper chitin detection (Sanchez-Vallet et al., 2015) as it is known for Magnaporthe (Fujikawa et al., 2012). Both SA and JA are known to be upregulated upon infection with pathogens as a part of the innate immune system and to trigger a specific set of pathogenesis-related (PR) genes (Thomma et al., 1998). Infection with C. higginsianum leads to induction of PR transcripts, like SA-dependent PR1 and PR2 while PDF1.2 is induced by JA-dependent signaling (Narusaka et al., 2004; Liu et al., 2007). We found that ectopic NahG expression and to a lesser extent sid2 mutations which both result in reduced SA levels, allow locally limited colonization by chr11Δ strains suggesting that genes on chromosome 11 may also interfere with SA-mediated defense responses. For Pseudomonas syringae and the oomycete Peronospora parasitica, it was reported that NahG mutants affect susceptibility more strongly than the sid2 mutant (Nawrath and Métraux, 1999) which fits our observations. However, the spreading of the chr11Δ strain was very limited and not comparable to the C. higginsianum WT (Figure 5). The double mutant cyp79b2/cyp79b3, defective for two monooxygenases was the only Arabidopsis line susceptible to chr11Δ. We therefore assume any of the several metabolites downstream of their product indole-3-acetaldoxime (IAOx) to be a key component in the defense against C. higginsianum. IAOx is the starting component of several biochemical pathways including auxin, camalexin and indole glucosinolates. Although camalexin was reported to play an important role in the defense against C. higginsianum (Narusaka et al., 2004), mutants defective for this compound (pad3) were not more susceptible to the chr11Δ strain. The same was true for pen2-2, a preinvasion defense defective mutant involved in indole glucosinolate biosynthesis (Bednarek et al., 2009). Either IAOx synthesis itself or other tryptophan derived secondary metabolites must therefore be targeted by genes on chromosome 11 to successfully suppress infection by C. higginsianum. Our fortuitous finding of strains lacking chromosome 11 should now allow to identify the relevant genes.
Potential Pathogenicity Genes on Chromosome 11
The genome sequencing of C. higginsianum (Dallery et al., 2017) revealed shared characteristics for chromosome 11 and 12 which differ from the core genome. Both display a lower gene density, are significantly enriched with transposable elements and have a higher AT-content. Most interestingly, both are also enriched in genes encoding potential effector proteins and proteins of unknown function. We did not observe any virulence defects in chr12Δ strains, suggesting no contribution in the pathogenicity against A. thaliana. In contrast, we found chromosome 11 to be directly associated with virulence, in particular in the suppression of post invasion defense mechanisms, indicating that a certain subset of genes from chromosome 11 have critical functions in manipulating plant host responses. In total, there are 7 candidate effector genes (ChEC) on chromosome 11 which may contribute to fungal virulence. ChEC7 is duplicated on both arms of the chromosome. This duplicated region harbors three genes within 7 kb all of which have homologs in other species. ChEC12 and ChEC12a exhibit high similarity. Both genes are expressed during the biotrophic phase. ChEC21 and ChEC24 are not species specific genes and are expressed during the initial phases of infection. Interestingly, ChEC116 is expressed during the whole infection and is one of the most highly expressed genes in the biotrophic phase of C. higginsianum (Dallery et al., 2017).
Moreover, there are in total five genes annotated as Nudix domain-containing proteins which code for hydrolases of nucleoside diphosphate linked compounds and which have been suggested to have important function in plant pathogens (Dong and Wang, 2016). A Nudix domain-containing protein has been reported to be involved in the switch from biotrophy to necrotrophy in C. truncatum (Bhadauria et al., 2013).
In summary, chromosome 11 harbors a number of potential genes which can now be selectively analyzed to evaluate their possible function in suppressing post invasion host defense.
Data Availability Statement
The genome data for this study can be found in the sequence read archive (SRA) at NCBI (accession number of bioproject: PRJNA427316 [https://www.ncbi.nlm.nih.gov/bioproject/PRJNA427316]).
Author Contributions
CK conceived the project. P-LP, MD, and JS performed the experiments. CK, P-LP, and MD analyzed the experimental data. LT and CK performed the bioinformatic analysis. P-LP and CK wrote the manuscript. All authors edited the manuscript.
Funding
Genome sequencing was funded by the Universitätsbund Erlangen.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Lars Voll for discussions and helpful suggestions. The generous help of Volker Huss teaching us the art of PFGE and providing a suitable electrophoresis unit is gratefully acknowledged. We thank Richard O’Connell for sharing strains and sequence information. We further thank Thomas Winkler for helping us with cell sorting and the many fruitful discussions. And we thank G. Mertens for making possible what seemed impossible.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01005/full#supplementary-material
Footnotes
References
Balesdent, M. H., Fudal, I., Ollivier, B., Bally, P., Grandaubert, J., Eber, F., et al. (2013). The dispensable chromosome of Leptosphaeria maculans shelters an effector gene conferring avirulence towards Brassica rapa. New Phytol. 198, 887–898. doi: 10.1111/nph.12178
Beadle, J., Wright, M., McNeely, L., and Bennett, J. W. (2003). Electrophoretic karyotype analysis in fungi. Adv. Appl. Microbiol. 53, 243–270. doi: 10.1016/S0065-2164(03)53007-6
Bednarek, P., Piślewska-Bednarek, M., Svatoš, A., Schneider, B., Doubský, J., Mansurova, M., et al. (2009). A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 323, 101–106. doi: 10.1126/science.1163732
Bhadauria, V., Banniza, S., Vandenberg, A., Selvaraj, G., and Wei, Y. (2013). Overexpression of a novel biotrophy-specific Colletotrichum truncatum effector, CtNUDIX, in hemibiotrophic fungal phytopathogens causes incompatibility with their host plants. Eukaryot. Cell 12, 2–11. doi: 10.1128/EC.00192-12
Camacho, J. P., Sharbel, T. F., and Beukeboom, L. W. (2000). B-chromosome evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 355, 163–178. doi: 10.1098/rstb.2000.0556
Chen, J., Upadhyaya, N. M., Ortiz, D., Sperschneider, J., Li, F., Bouton, C., et al. (2017). Loss of AvrSr50 by somatic exchange in stem rust leads to virulence for Sr50 resistance in wheat. Science 358, 1607–1610. doi: 10.1126/science.aao4810
Dallery, J. F., Lapalu, N., Zampounis, A., Pigne, S., Luyten, I., Amselem, J., et al. (2017). Gapless genome assembly of Colletotrichum higginsianum reveals chromosome structure and association of transposable elements with secondary metabolite gene clusters. BMC Genomics 18:667. doi: 10.1186/s12864-017-4083-x
D’Ambrosio, U., Alonso-Lifante, M. P., Barros, K., Kovarik, A., Mas de Xaxars, G., and Garcia, S. (2017). B-chrom: a database on B-chromosomes of plants, animals and fungi. New Phytol. 216, 635–642. doi: 10.1111/nph.14723
de Jonge, R., Bolton, M. D., Kombrink, A., van den Berg, G. C., Yadeta, K. A., and Thomma, B. P. (2013). Extensive chromosomal reshuffling drives evolution of virulence in an asexual pathogen. Genome Res. 23, 1271–1282. doi: 10.1101/gr.152660.112
Delaney, T. P., Uknes, S., Vernooij, B., Friedrich, L., Weymann, K., Negrotto, D., et al. (1994). A central role of salicylic acid in plant disease resistance. Science 266, 1247–1250. doi: 10.1126/science.266.5188.1247
Dong, S., Raffaele, S., and Kamoun, S. (2015). The two-speed genomes of filamentous pathogens: waltz with plants. Curr. Opin. Genet. Dev. 35, 57–65. doi: 10.1016/j.gde.2015.09.001
Dong, S., and Wang, Y. (2016). Nudix effectors: a common weapon in the arsenal of plant pathogens. PLoS Pathog. 12:e1005704. doi: 10.1371/journal.ppat.1005704
Duplessis, S., Cuomo, C. A., Lin, Y. C., Aerts, A., Tisserant, E., Veneault-Fourrey, C., et al. (2011). Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Natl. Acad. Sci. U.S.A. 108, 9166–9171. doi: 10.1073/pnas.1019315108
Engelsdorf, T., Horst, R. J., Pröls, R., Pröschel, M., Dietz, F., Hückelhoven, R., et al. (2013). Reduced carbohydrate availability enhances the susceptibility of arabidopsis toward Colletotrichum higginsianum. Plant Physiol. 162, 225–238. doi: 10.1104/pp.112.209676
Faino, L., Seidl, M. F., Shi-Kunne, X., Pauper, M., van den Berg, G. C., Wittenberg, A. H., et al. (2016). Transposons passively and actively contribute to evolution of the two-speed genome of a fungal pathogen. Genome Res. 26, 1091–1100. doi: 10.1101/gr.204974.116
Fujikawa, T., Sakaguchi, A., Nishizawa, Y., Kouzai, Y., Minami, E., Yano, S., et al. (2012). Surface alpha-1,3-glucan facilitates fungal stealth infection by interfering with innate immunity in plants. PLoS Pathog. 8:e1002882. doi: 10.1371/journal.ppat.1002882
Glazebrook, J., Zook, M., Mert, F., Kagan, I., Rogers, E. E., Crute, I. R., et al. (1997). Phytoalexin-deficient mutants of Arabidopsis reveal that PAD4 encodes a regulatory factor and that four PAD genes contribute to downy mildew resistance. Genetics 146, 381–392.
Hartwell, L. H., Dutcher, S. K., Wood, J. S., and Garvik, B. (1982). The fidelity of mitotic chromosome reproduction in S. cerevisiae. Rec. Adv. Yeast Mol. Biol. 1, 28–38.
Hatta, R., Ito, K., Hosaki, Y., Tanaka, T., Tanaka, A., Yamamoto, M., et al. (2002). A conditionally dispensable chromosome controls host-specific pathogenicity in the fungal plant pathogen Alternaria alternata. Genetics 161, 59–70.
He, C. Z., Rusu, A. G., Poplawski, A. M., Irwin, J. A., and Manners, J. M. (1998). Transfer of a supernumerary chromosome between vegetatively incompatible biotypes of the fungus Colletotrichum gloeosporioides. Genetics 150, 1459–1466.
Hieter, P., Mann, C., Snyder, M., and Davis, R. W. (1985). Mitotic stability of yeast chromosomes: a colony color assay that measures nondisjunction and chromosome loss. Cell 40, 381–392. doi: 10.1016/0092-8674(85)90152-7
Hiruma, K., Fukunaga, S., Bednarek, P., Piślewska-Bednarek, M., Watanabe, S., Narusaka, Y., et al. (2013). Glutathione and tryptophan metabolism are required for Arabidopsis immunity during the hypersensitive response to hemibiotrophs. Proc. Natl. Acad. Sci. U.S.A. 110, 9589–9594. doi: 10.1073/pnas.1305745110
Huser, A., Takahara, H., Schmalenbach, W., and O’Connell, R. (2009). Discovery of pathogenicity genes in the crucifer anthracnose fungus Colletotrichum higginsianum, using random insertional mutagenesis. Mol. Plant Microbe Interact. 22, 143–156. doi: 10.1094/MPMI-22-2-0143
Ishikawa, F. H., Souza, E. A., Shoji, J. Y., Connolly, L., Freitag, M., Read, N. D., et al. (2012). Heterokaryon incompatibility is suppressed following conidial anastomosis tube fusion in a fungal plant pathogen. PLoS One 7:e31175. doi: 10.1371/journal.pone.0031175
Jones, J. D., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Jones, J. D., Vance, R. E., and Dangl, J. L. (2016). Intracellular innate immune surveillance devices in plants and animals. Science 354:aaf6395. doi: 10.1126/science.aaf6395
Kamper, J., Kahmann, R., Bolker, M., Ma, L. J., Brefort, T., Saville, B. J., et al. (2006). Insights from the genome of the biotrophic fungal plant pathogen Ustilago maydis. Nature 444, 97–101. doi: 10.1038/nature05248
Keogh, R. C., Deverall, B. J., and Mcleod, S. (1980). Comparison of histological and physiological responses to Phakopsora Pachyrhizi in resistant and susceptible soybean. Mycol. Res. 74, 329–333. doi: 10.1016/S0007-1536(80)80163-X
Kleemann, J., Rincon-Rivera, L. J., Takahara, H., Neumann, U., Ver Loren van Themaat, E., van der Does, H. C., et al. (2012). Sequential delivery of host-induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum. PLoS Pathog. 8:e1002643. doi: 10.1371/journal.ppat.1002643
Kleemann, J., Takahara, H., Stuber, K., and O’Connell, R. (2008). Identification of soluble secreted proteins from appressoria of Colletotrichum higginsianum by analysis of expressed sequence tags. Microbiology 154(Pt 4), 1204–1217. doi: 10.1099/mic.0.2007/014944-0
Koeck, M., Hardham, A. R., and Dodds, P. N. (2011). The role of effectors of biotrophic and hemibiotrophic fungi in infection. Cell. Microbiol. 13, 1849–1857. doi: 10.1111/j.1462-5822.2011.01665.x
Korn, M., Schmidpeter, J., Dahl, M., Muller, S., Voll, L. M., and Koch, C. (2015). A genetic screen for pathogenicity genes in the hemibiotrophic fungus Colletotrichum higginsianum identifies the plasma membrane proton pump Pma2 required for host penetration. PLoS One 10:e0125960. doi: 10.1371/journal.pone.0125960
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Lawton, K., Weymann, K., Friedrich, L., Vernooij, B., Uknes, S., and Ryals, J. (1995). Systemic acquired resistance in Arabidopsis requires salicylic acid but not ethylene. Mol. Plant Microbe Interact. 8, 863–870. doi: 10.1094/Mpmi-8-0863
Liu, G., Kennedy, R., Greenshields, D. L., Peng, G., Forseille, L., Selvaraj, G., et al. (2007). Detached and attached Arabidopsis leaf assays reveal distinctive defense responses against hemibiotrophic Colletotrichum spp. Mol. Plant Microbe Interact. 20, 1308–1319. doi: 10.1094/MPMI-20-10-1308
Liu, L., Zhao, D., Zheng, L., Hsiang, T., Wei, Y., Fu, Y., et al. (2013). Identification of virulence genes in the crucifer anthracnose fungus Colletotrichum higginsianum by insertional mutagenesis. Microb. Pathog. 64, 6–17. doi: 10.1016/j.micpath.2013.06.001
Ma, L. J., van der Does, H. C., Borkovich, K. A., Coleman, J. J., Daboussi, M. J., Di Pietro, A., et al. (2010). Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 464, 367–373. doi: 10.1038/nature08850
Maringele, L., and Lydall, D. (2006). Pulsed-field gel electrophoresis of budding yeast chromosomes. Methods Mol. Biol. 313, 65–73. doi: 10.1385/1-59259-958-3:065
Masel, A. M., He, C. Z., Poplawski, A. M., Irwin, J. A. G., and Manners, J. M. (1996). Molecular evidence for chromosome transfer between biotypes of Colletotrichum gloeosporioides. Mol. Plant Microbe Interact. 9, 339–348. doi: 10.1094/Mpmi-9-0339
Miao, V. P., Covert, S. F., and Vanetten, H. D. (1991). A fungal gene for antibiotic-resistance on a dispensable (B) chromosome. Science 254, 1773–1776. doi: 10.1126/science.1763326
Miya, A., Albert, P., Shinya, T., Desaki, Y., Ichimura, K., Shirasu, K., et al. (2007). CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 104, 19613–19618. doi: 10.1073/pnas.0705147104
Moller, M., and Stukenbrock, E. H. (2017). Evolution and genome architecture in fungal plant pathogens. Nat. Rev. Microbiol. 15, 756–771. doi: 10.1038/nrmicro.2017.76
Narusaka, Y., Narusaka, M., Park, P., Kubo, Y., Hirayama, T., Seki, M., et al. (2004). RCH1, a locus in Arabidopsis that confers resistance to the hemibiotrophic fungal pathogen Colletotrichum higginsianum. Mol. Plant Microbe Interact. 17, 749–762. doi: 10.1094/MPMI.2004.17.7.749
Nawrath, C., and Métraux, J.-P. (1999). Salicylic acid induction–deficient mutants of Arabidopsis express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell 11, 1393–1404. doi: 10.1105/tpc.11.8.1393
O’Connell, R., Herbert, C., Sreenivasaprasad, S., Khatib, M., Esquerré-Tugayé, M.-T., and Dumas, B. (2004). A novel Arabidopsis-Colletotrichum pathosystem for the molecular dissection of plant-fungal interactions. Mol. Plant Microbe Interact. 17, 272–282. doi: 10.1094/MPMI.2004.17.3.272
O’Connell, R. J., and Panstruga, R. (2006). Tête à tête inside a plant cell: establishing compatibility between plants and biotrophic fungi and oomycetes. New Phytol. 171, 699–718. doi: 10.1111/j.1469-8137.2006.01829.x
O’Connell, R. J., Thon, M. R., Hacquard, S., Amyotte, S. G., Kleemann, J., Torres, M. F., et al. (2012). Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 44, 1060–1065. doi: 10.1038/ng.2372
Penninckx, I. A., Eggermont, K., Terras, F. R., Thomma, B. P., De Samblanx, G. W., Buchala, A., et al. (1996). Pathogen-induced systemic activation of a plant defensin gene in Arabidopsis follows a salicylic acid-independent pathway. Plant Cell 8, 2309–2323. doi: 10.1105/tpc.8.12.2309
Peter, J., De Chiara, M., Friedrich, A., Yue, J. X., Pflieger, D., Bergstrom, A., et al. (2018). Genome evolution across 1,011 Saccharomyces cerevisiae isolates. Nature 556, 339–344. doi: 10.1038/s41586-018-0030-5
Pogany, M., von Rad, U., Grun, S., Dongo, A., Pintye, A., Simoneau, P., et al. (2009). Dual roles of reactive oxygen species and NADPH oxidase RBOHD in an Arabidopsis-Alternaria pathosystem. Plant Physiol. 151, 1459–1475. doi: 10.1104/pp.109.141994
Raffaele, S., and Kamoun, S. (2012). Genome evolution in filamentous plant pathogens: why bigger can be better. Nat. Rev. Microbiol. 10, 417–430. doi: 10.1038/nrmicro2790
Rieu, I., and Powers, S. J. (2009). Real-time quantitative RT-PCR: design, calculations, and statistics. Plant Cell 21, 1031–1033. doi: 10.1105/tpc.109.066001
Sanchez-Vallet, A., Mesters, J. R., and Thomma, B. P. (2015). The battle for chitin recognition in plant-microbe interactions. FEMS Microbiol. Rev. 39, 171–183. doi: 10.1093/femsre/fuu003
Sanchez-Vallet, A., Ramos, B., Bednarek, P., López, G., Piślewska-Bednarek, M., Schulze-Lefert, P., et al. (2010). Tryptophan-derived secondary metabolites in Arabidopsis thaliana confer non-host resistance to necrotrophic Plectosphaerella cucumerina fungi. Plant J. 63, 115–127. doi: 10.1111/j.1365-313X.2010.04224.x
Schmidpeter, J., Dahl, M., Hofmann, J., and Koch, C. (2017). ChMob2 binds to ChCbk1 and promotes virulence and conidiation of the fungal pathogen Colletotrichum higginsianum. BMC Microbiol. 17:22. doi: 10.1186/s12866-017-0932-7
Seidl, M. F., and Thomma, B. P. (2014). Sex or no sex: evolutionary adaptation occurs regardless. Bioessays 36, 335–345. doi: 10.1002/bies.201300155
Siebert, P. D., Chenchik, A., Kellogg, D. E., Lukyanov, K. A., and Lukyanov, S. A. (1995). An improved PCR method for walking in uncloned genomic DNA. Nucleic Acids Res. 23, 1087–1088. doi: 10.1093/nar/23.6.1087
Spanu, P. D., Abbott, J. C., Amselem, J., Burgis, T. A., Soanes, D. M., Stuber, K., et al. (2010). Genome expansion and gene loss in powdery mildew fungi reveal tradeoffs in extreme parasitism. Science 330, 1543–1546. doi: 10.1126/science.1194573
Stergiopoulos, I., and de Wit, P. J. (2009). Fungal effector proteins. Annu. Rev. Phytopathol. 47, 233–263. doi: 10.1146/annurev.phyto.112408.132637
Taga, M., Tanaka, K., Kato, S., and Kubo, Y. (2015). Cytological analyses of the karyotypes and chromosomes of three Colletotrichum species, C. orbiculare, C. graminicola and C. higginsianum. Fungal Genet. Biol. 82, 238–250. doi: 10.1016/j.fgb.2015.07.013
Takahara, H., Hacquard, S., Kombrink, A., Hughes, H. B., Halder, V., Robin, G. P., et al. (2016). Colletotrichum higginsianum extracellular LysM proteins play dual roles in appressorial function and suppression of chitin-triggered plant immunity. New Phytol. 211, 1323–1337. doi: 10.1111/nph.13994
Takahashi-Nakaguchi, A., Muraosa, Y., Hagiwara, D., Sakai, K., Toyotome, T., Watanabe, A., et al. (2015). Genome sequence comparison of Aspergillus fumigatus strains isolated from patients with pulmonary aspergilloma and chronic necrotizing pulmonary aspergillosis. Med. Mycol. 53, 353–360. doi: 10.1093/mmy/myv003
Thomma, B. P., Eggermont, K., Penninckx, I. A., Mauch-Mani, B., Vogelsang, R., Cammue, B. P., et al. (1998). Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. U.S.A. 95, 15107–15111. doi: 10.1073/pnas.95.25.15107
Tsuge, T., Harimoto, Y., Akimitsu, K., Ohtani, K., Kodama, M., Akagi, Y., et al. (2013). Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 37, 44–66. doi: 10.1111/j.1574-6976.2012.00350.x
Uknes, S., Mauch-Mani, B., Moyer, M., Potter, S., Williams, S., Dincher, S., et al. (1992). Acquired resistance in Arabidopsis. Plant Cell 4, 645–656. doi: 10.1105/tpc.4.6.645
VanEtten, H., Jorgensen, S., Enkerli, J., and Covert, S. F. (1998). Inducing the loss of conditionally dispensable chromosomes in Nectria haematococca during vegetative growth. Curr. Genet. 33, 299–303. doi: 10.1007/s002940050340
Vlaardingerbroek, I., Beerens, B., Rose, L., Fokkens, L., Cornelissen, B. J., and Rep, M. (2016a). Exchange of core chromosomes and horizontal transfer of lineage-specific chromosomes in Fusarium oxysporum. Environ. Microbiol. 18, 3702–3713. doi: 10.1111/1462-2920.13281
Vlaardingerbroek, I., Beerens, B., Schmidt, S. M., Cornelissen, B. J., and Rep, M. (2016b). Dispensable chromosomes in Fusarium oxysporum f. sp. lycopersici. Mol. Plant Pathol. 17, 1455–1466. doi: 10.1111/mpp.12440
Yoshida, K., Saitoh, H., Fujisawa, S., Kanzaki, H., Matsumura, H., Yoshida, K., et al. (2009). Association genetics reveals three novel avirulence genes from the rice blast fungal pathogen Magnaporthe oryzae. Plant Cell 21, 1573–1591. doi: 10.1105/tpc.109.066324
Keywords: Arabidopsis, plant defense, protein effectors, chromosome loss, B-chromosomes, virulence
Citation: Plaumann P-L, Schmidpeter J, Dahl M, Taher L and Koch C (2018) A Dispensable Chromosome Is Required for Virulence in the Hemibiotrophic Plant Pathogen Colletotrichum higginsianum. Front. Microbiol. 9:1005. doi: 10.3389/fmicb.2018.01005
Received: 16 March 2018; Accepted: 30 April 2018;
Published: 18 May 2018.
Edited by:
Jens Staal, Ghent University, BelgiumReviewed by:
Richard John O’Connell, Institut National de la Recherche Agronomique (INRA), FranceRoger W. Innes, Indiana University Bloomington, United States
Copyright © 2018 Plaumann, Schmidpeter, Dahl, Taher and Koch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Koch, christian.koch@fau.de