 Nina Schleimer1
Nina Schleimer1 Ursula Kaspar1Mike Drescher1Jochen Seggewiß2Christof von Eiff1†
Ursula Kaspar1Mike Drescher1Jochen Seggewiß2Christof von Eiff1† Richard A. Proctor3
Richard A. Proctor3 Georg Peters1
Georg Peters1 André Kriegeskorte1†
André Kriegeskorte1† Karsten Becker1*
Karsten Becker1*- 1Institute of Medical Microbiology, University Hospital Münster, Münster, Germany
- 2Institute of Human Genetics, University Hospital Münster, Münster, Germany
- 3Departments of Medical Microbiology/Immunology and Medicine, University of Wisconsin School of Medicine and Public Health, Madison, WI, United States
Staphylococcal small-colony variants (SCVs) are invasive and persistent due to their ability to thrive intracellularly and to evade the host immune response. Thus, the course of infections due to this phenotype is often chronic, relapsing, and therapy-refractory. In order to improve treatment of patients suffering from SCV-associated infections, it is of major interest to understand triggers for the development of this phenotype, in particular for strains naturally occurring in clinical settings. Within this study, we comprehensively characterized two different Staphylococcus aureus triplets each consisting of isogenic strains comprising (i) clinically derived SCV phenotypes with auxotrophy for unsaturated fatty acids, (ii) the corresponding wild-types (WTs), and (iii) spontaneous in vitro revertants displaying the normal phenotype (REVs). Comparison of whole genomes revealed that clinical SCV isolates were closely related to their corresponding WTs and REVs showing only seven to eight alterations per genome triplet. However, both SCVs carried a mutation within the energy-coupling factor (ECF) transporter-encoding ecf module (EcfAA’T) resulting in truncated genes. In both cases, these mutations were shown to be naturally restored in the respective REVs. Since ECF transporters are supposed to be essential for optimal bacterial growth, their dysfunction might constitute another mechanism for the formation of naturally occurring SCVs. Another three triplets analyzed revealed neither mutations in the EcfAA’T nor in other FASII-related genes underlining the high diversity of mechanisms leading to the fatty acid-dependent phenotype. This is the first report on the ECF transporter as genetic basis of fatty acid–auxotrophic staphylococcal SCVs.
Introduction
As an integral part of the normal bacterial life cycle and the infection process, the formation of the small-colony variant (SCV) phenotype enables staphylococcal cells to adapt to an intracellular lifestyle protecting them against the host defense system and antimicrobial therapy resulting in persistent, relapsing, and often therapy-refractory chronic infections (Tuchscherr et al., 2010, 2011; Kriegeskorte et al., 2011, 2014b; Edwards, 2012; Kahl et al., 2016).
Two major types of SCVs have been most frequently associated with clinical cases (Proctor et al., 2006, 2014): (i) electron transport-deficient SCVs, which are typically recovered from patients suffering from chronic osteomyelitis and/or treated with aminoglycosides and found to exhibit auxotrophies for hemin, menadione, or thiamine, respectively (Proctor et al., 1995; von Eiff et al., 1997b; Kohler et al., 2008) and (ii) thymidine-auxotrophic SCVs with thymidylate biosynthesis defects recovered from cystic fibrosis patients especially after long-term trimethoprim–sulfamethoxazole treatment (Kahl et al., 1998; Kriegeskorte et al., 2014a). Based on in vitro generation of deletion mutants and sequencing, the genetic basis of only a small fraction of these SCV phenotypes could be clarified so far discovering defects in genes including aroB, aroD, hemA-D, hemG, hemH, menA-F, and thyA, respectively (von Eiff et al., 1997b; Bates et al., 2003; Schaaff et al., 2003; Chatterjee et al., 2008; Lannergård et al., 2008; Köser et al., 2012; Hammer et al., 2013; Dean et al., 2014; Painter et al., 2015; Cao et al., 2017; Zhang et al., 2017). However, for clinically derived SCVs, only mutations of the hemG, menB, menC, menE, menF, and thyA genes were identified. Besides these intensively studied mechanisms of SCV formation, less is known on SCVs dependent on unsaturated fatty acids or other compounds (Sherris, 1952; Slifkin et al., 1971; Kaplan and Dye, 1976; Gómez-González et al., 2010; Lin et al., 2016). Hitherto, the fatty acid metabolism-linked genes accC, accD, fabF, fabI (eventually combined with fabD), and plsX have been associated with the phenotype switch of fatty acid-auxotrophic SCVs, with fabF mutation being the only one found in a clinical isolate (Parsons et al., 2011, 2013, 2014; Lin et al., 2016; Bazaid et al., 2018). Moreover, the underlying auxotrophism could be elucidated only for a part of the SCVs (Garcia et al., 2013), which further complicates the identification of potential genes. As an example, a mutation in relA was identified as potential trigger for the phenotype switch (Gao et al., 2010).
Here, we comprehensively analyzed two different triplets of isogenic S. aureus isolates, each comprising a clinically derived, fatty acid–auxotrophic SCV phenotype, its corresponding wild-type (WT) strain, both sampled in parallel, and a spontaneous in vitro revertant (REV) displaying the normal phenotype. In particular, the unsaturated fatty acid-based auxotrophism of SCVs was characterized and the phenotype switch was assessed by a whole-genome sequencing (WGS) approach. WGS revealed SCV formation-associated mutations within the energy-coupling factor (ECF) transporter-encoding ecf module (EcfAA’T) for both triplets and, in one triplet, an alteration within the Agr system most likely responsible for the decreased hemolytic activity displayed by the SCV and REV.
Materials and Methods
Bacterial Strains and Culture Conditions
Clinical S. aureus WT isolates and their corresponding SCVs were recovered in parallel from patients of the University Hospital Münster in Germany suffering from different infections (Table 1 and Supplementary Table 1 in the Supplementary Material). Strains were cultivated and grown on Columbia blood agar (BBLTM Columbia agar with 5% sheep blood, Becton Dickinson, Franklin Lakes, NJ, United States) at 37°C, then frozen and stored at -80°C until testing. Isolates exhibiting the following characteristics were identified as SCVs: (i) pinpoint colonies on Columbia blood agar (Becton Dickinson, Franklin Lakes, NJ, United States) after 24–72 h of incubation, (ii) reduced hemolytic activity, and (iii) decreased pigmentation. Corresponding REVs exhibiting the normal phenotype (Becker et al., 2006; Proctor et al., 2006) emerged spontaneously from subcultured SCV isolates after several in vitro passages on Columbia blood agar (Becton Dickinson, Franklin Lakes, NJ, United States).
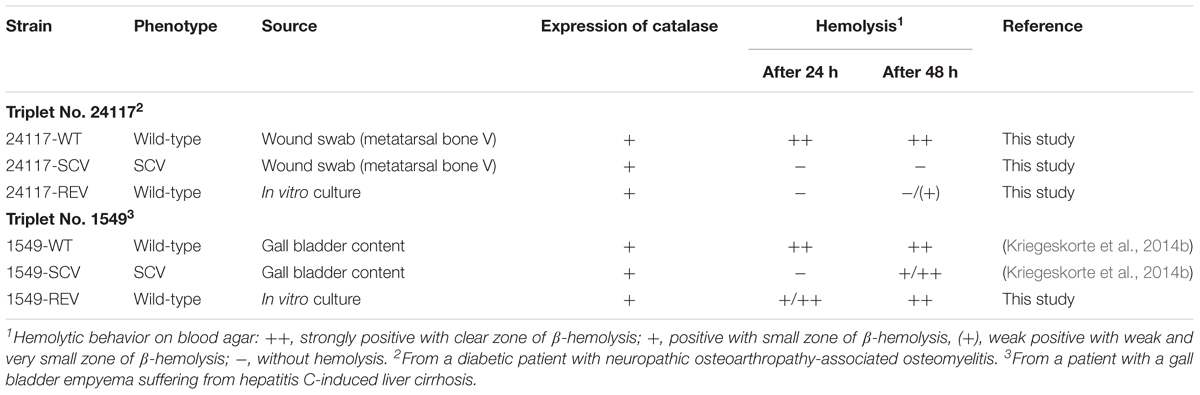
TABLE 1. Characteristics of the strain triplets analyzed in this study.
To monitor possible spontaneous reversions of the SCVs into the WT, every step of the cultivation procedures and the inoculum preparations were performed exclusively on solid media. SCVs were tested for reversion by subculturing of individual colonies onto Columbia blood agar (Becton Dickinson, Franklin Lakes, NJ, United States) under non-stress conditions (37°C, 24–48 h). The ability of the SCV colonies to revert to normal-sized colonies was judged visually regarding change in size and hemolysis behavior. Colonies that matched the SCV criteria underwent several (at least 10) passages of subculturing on solid media.
Differences in colony sizes were assessed on solid media by measuring the size of 50 single colonies on Columbia blood agar after 24 h of incubation. To analyze the results of colony sizes, statistical analyses were performed applying non-parametric tests using the Kruskal–Wallis test, with p = 0.05 set for statistical significance.
DNA Manipulations
Unless otherwise stated, all DNA manipulations were carried out following standard procedures and manufacturer’s recommendations. Chromosomal DNA from S. aureus cells was extracted after lysostaphin treatment (20 μg/mL, 1 h, 37°C) (Wak-Chemie Medical, Steinbach, Germany) using the PrestoSpin D kit (Molzym, Bremen, Germany). Plasmid DNA was isolated with the Qiagen Plasmid Mini kit (Qiagen, Venlo, Netherlands). PCRs were performed using oligonucleotides listed in Table 2 and Taq DNA Polymerase (Segenetic, Borken, Germany). Standard-PCR conditions consist of 5-min initial denaturation step at 95°C followed by 35 cycles of (i) denaturation at 95°C for 30 s, (ii) annealing at 65°C for 30 s, and (iii) extension at 72°C for 3 min. The final extension was performed at 72°C for 7 min. PCR products were analyzed by agarose gel electrophoresis and purified using the QIAquick PCR Purification kit (Qiagen, Venlo, Netherlands). All PCR amplicons and constructed mutants were analyzed by Sanger sequencing (Eurofins Genomics, Ebersberg, Germany).

TABLE 2. Oligonucleotides used in this study.
S. aureus Knockout Mutants
The ΔthyA mutant was constructed as previously published using the vector pBT9-thyA::ermB that was transformed by electroporation into clinical 1549-WT applying the standard protocol (Kriegeskorte et al., 2014a). Further cultivation and integration (first recombination) of pBT9-thyA::ermB into the genome of 1549-WT were performed as described (Kriegeskorte et al., 2014a) with the exceptions of using brain heart infusion broth (BHI, Merck, Darmstadt, Germany) supplemented with erythromycin (2.5 μg/mL), chloramphenicol (10 μg/mL), and thymidine (100 μg/mL). Integration was verified by PCR of selected colonies (Supplementary Table 2). For resolution (second recombination), an overnight culture of verified 1549-pBT9-thyA::ermB were then grown in BHI (Merck, Darmstadt, Germany) containing erythromycin and thymidine at 25°C. Further procedure and selection of the mutants was performed as described elsewhere (Kriegeskorte et al., 2014a) with the exceptions for using erythromycin at a concentration of 2.5 μg/mL and susceptibility disks (Oxoid, Hampshire, United Kingdom) impregnated with 10 μL of thymidine (10 mg/mL) for supplementation of Mueller-Hinton agar (MHA, Merck, Darmstadt, Germany). Deletion of thyA was verified via PCR amplification followed by sequencing (Eurofins Genomics). Furthermore, a ΔhemB mutant of the clinical S. aureus A3878-WT and a ΔmenD mutant derived from the laboratory strain COL, both constructed as previously published (von Eiff et al., 2006; Kriegeskorte et al., 2011), were also included in the experiments (Table 3).

TABLE 3. Genetically defined SCVs used as positive controls in auxotrophism studies.
Screening for Alterations Within the ecf Module and the FASII Pathway Genes
In order to screen further strain triplets (listed in Supplementary Table 1) for alterations within the ecf module, amplification of the module was performed by standard PCR with annealing at 55°C and oligonucleotides ecf-F and ecf-R. For sequencing (Eurofins Genomics), oligonucleotides listed in Table 2 were used. Alterations within the FASII pathway genes were screened by amplification of genes by standard PCR with annealing at 55°C and extension for 1 min for genes accB, fabG, fabI, and fabZ and extension for 1.5 min for genes accA, accC, accD, fabD, fabF, fabH, and plsX followed by Sanger sequencing (Eurofins Genomics, oligonucleotides listed in Table 2).
Genotyping by PFGE
Clonal relationship within the strain triplets for the corresponding WT, SCV, and REV strains was confirmed by SmaI macrorestriction analyses of total bacterial DNA followed by resolving the digests using pulsed-field gel electrophoresis (PFGE) as previously described (Goering and Winters, 1992; von Eiff et al., 1997a). Instead of tryptic soy broth (TSB), BHI broth (Merck, Darmstadt, Germany) was used in order to optimize growth conditions for SCVs. Strains were considered clonally identical if less than two bands varied on the gel according to the published guidelines (Tenover et al., 1995).
MLST, Spa Typing, and Microarray Analysis
MLST was done based on the WGS data applying the PubMLST database and software available on PubMLST.org/saureus/ (Jolley and Maiden, 2010). Typing of the spa gene was performed with spaTyper 1.0 available from the Center for Genomic Epidemiology homepage (Bartels et al., 2014) also using whole-genome data. Verification of MLST and spa typing and determination of regulatory and hemolysin genes were performed at the genetic level using DNA microarray analyses (IdentiBAC Microarray, Alere, Jena, Germany).
Auxotrophism Studies
Auxotrophism testing was performed in triplicate on surface-dried, in particular condensation water-free MHA (Merck, Darmstadt, Germany). For further evaluation of growth, samples were adjusted to McFarland 0.5 (in 0.9% NaCl), diluted (1:1,000 for WTs and 1:100 for SCVs), and an amount of 100 μL was streaked on unsupplemented MHA (Merck, Darmstadt, Germany) and incubated for 24–48 h at 37°C. For evaluation of an underlying auxotrophism, samples were adjusted and streaked on MHA (Merck, Darmstadt, Germany) as described and a maximum of three blank antimicrobial susceptibility disks was laid on top of the MHA (Merck, Darmstadt, Germany) surface. Disks were impregnated with 10 μL of each of the solutions listed in Supplementary Table 3. Auxotrophism of the SCVs was presumed if a growth-promoting effect became exclusively visible only surrounding the impregnated disks after incubation for 24–48 h at 37°C. As positive controls for hemin-, menadione-, and thymidine-auxotrophy, ΔhemB, ΔmenD, and ΔthyA mutants, respectively, were included in all experiments (Table 3). For mutants, MHA (Merck, Darmstadt, Germany) supplemented with erythromycin at 2.5 μg/mL was used.
Further Characterization of Strains
Hemolysis activity was examined by culturing the strains on Columbia blood agar (Becton Dickinson, Franklin Lakes, NJ, United States) for 24 and 48 h. Hemolysis was considered as strongly positive (++) if showing a clear zone of β-hemolysis, positive (+) if showing a weak and small zone of β-hemolysis, and negative (-) when no hemolysis could be detected. The presence of catalase was confirmed using 3% hydrogen peroxide (Merck, Darmstadt, Germany).
Whole-Genome Sequencing of Triplets
For the PacBio RS II sequencing platform (Pacific Biosciences, Menlo Park, CA, United States), genomic DNAs of WTs, SCVs, and REVs were extracted after lysostaphin treatment (20 μg/mL, 1 h, 37°C) (Wak-Chemie Medical, Steinbach, Germany) using the Genomic-tip 20/G kit (Qiagen, Venlo, Netherlands). This was followed by sequencing on the Pacific Biosciences RS II instrumentation with use of P6 DNA polymerase with C4 chemistry (P6-C4), 110 pM of complexed 20 kb-SMRTbell library, and 240 min continuous movie collection. Initial de novo assembly of reads was performed using the HGAP3 (Chin et al., 2013) v 2.3.0 assembler. Assembly coverages were ranging between 96.77× and 607.93×, with 126,769–153,216 mapped reads and a mean read length from 11,158 to 12,746 bp (N50, 16,228–19,044 bp). Assembled genomes were annotated via the GenDB pipeline (Meyer et al., 2003) and BLAST+ 2.7.1 (Zhang et al., 2000). After sequence alignment of the three phenotypes with Mauve 2.4.0 (Darling et al., 2004) (RRID:SCR_012852) and Lasergene 12 (DNASTAR, Madison, WI, United States), detected differences between the phenotypes were verified applying standard PCR followed by Sanger sequencing. For detailed analysis of detected differences, SnapGene 4.0.6 (GSL Biotech, Chicago, IL, United States; available at snapgene.com; RRID:SCR_015052) was used. The Staphylococcal regulatory RNAs Database (SRD) (Sassi et al., 2015) was used for detection of small non-protein-coding RNAs (npcRNAs; often referred to as “non-coding RNA”) within the ecf module.
Availability of Supporting Data
The genome sequences of the sequenced strains were deposited in the European Nucleotide Archive ENA (Accession No. LT992434-LT992436 for triplet 1549 and LT996889-LT996891 for triplet 24117, respectively).
Results
Phenotypic, Biochemical, and Molecular Characterization
WTs and REVs exhibited a normal phenotype after 24 h (Figure 1) and SCVs demonstrated a significantly reduced colony size on Columbia blood agar (Becton Dickinson, Franklin Lakes, NJ, United States) after 24 h of incubation (Figures 1A,B). Their phenotypes were also stable after 48 h of incubation (Figure 2). WTs and REVs displayed hemolysis after 48 h with 24117-REV showing only a very weak hemolytic activity after 48 h of incubation. 1549-SCV exhibited hemolysis only after 48 h of incubation, whereas 24117-SCV was not hemolytic (Table 1 and Supplementary Figure 1). When cultivated on MHA (not supplemented), 1549-SCV was not able to grow after 48 h of incubation, whereas 24117-SCV exhibited no visible growth after 24 h but grew in micro-colonies after 48 h of incubation (Figure 2). Both SCVs were catalase-positive as their corresponding WTs and REVs (Table 1).
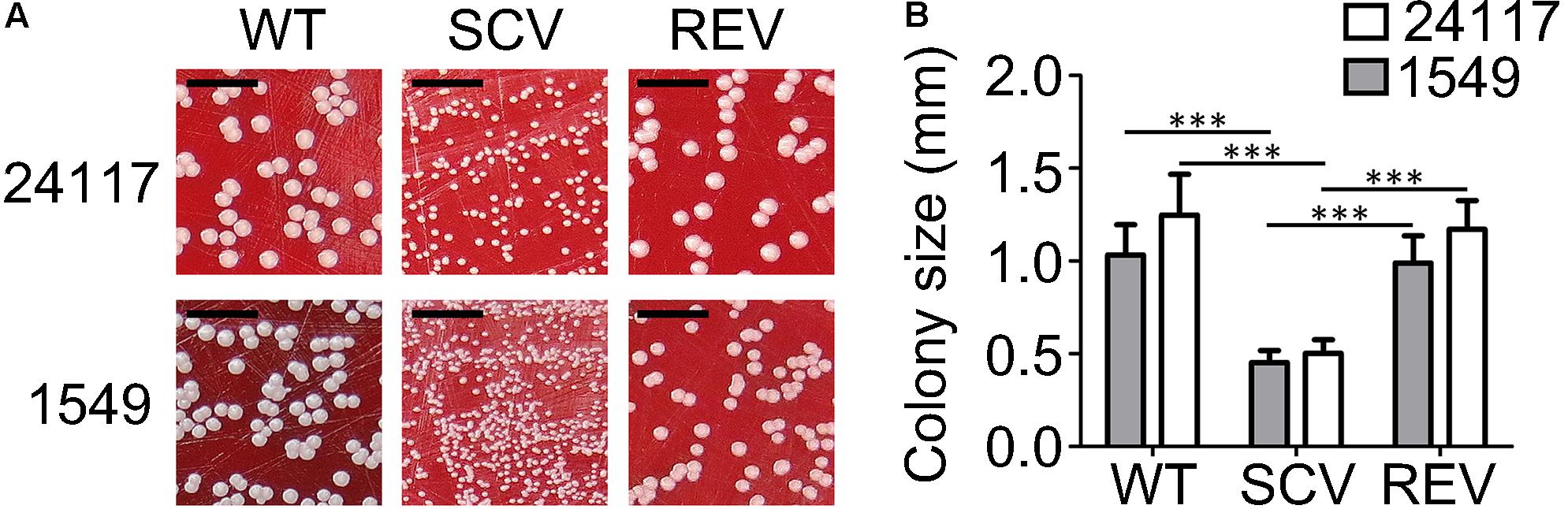
FIGURE 1. (A) Clinical S. aureus strain triplets 1549 and 24117 each comprising isogenic wild-type (WT), small-colony variant (SCV), and revertant (REV) phenotypes after 24 h of incubation at 37°C on Columbia blood agar with 5% sheep blood; scale bar indicates 5 mm. (B) Colony sizes of WTs, SCVs, and REVs after 24 h incubation on Columbia blood agar at 37°C; ∗∗∗P ≤ 0.001.
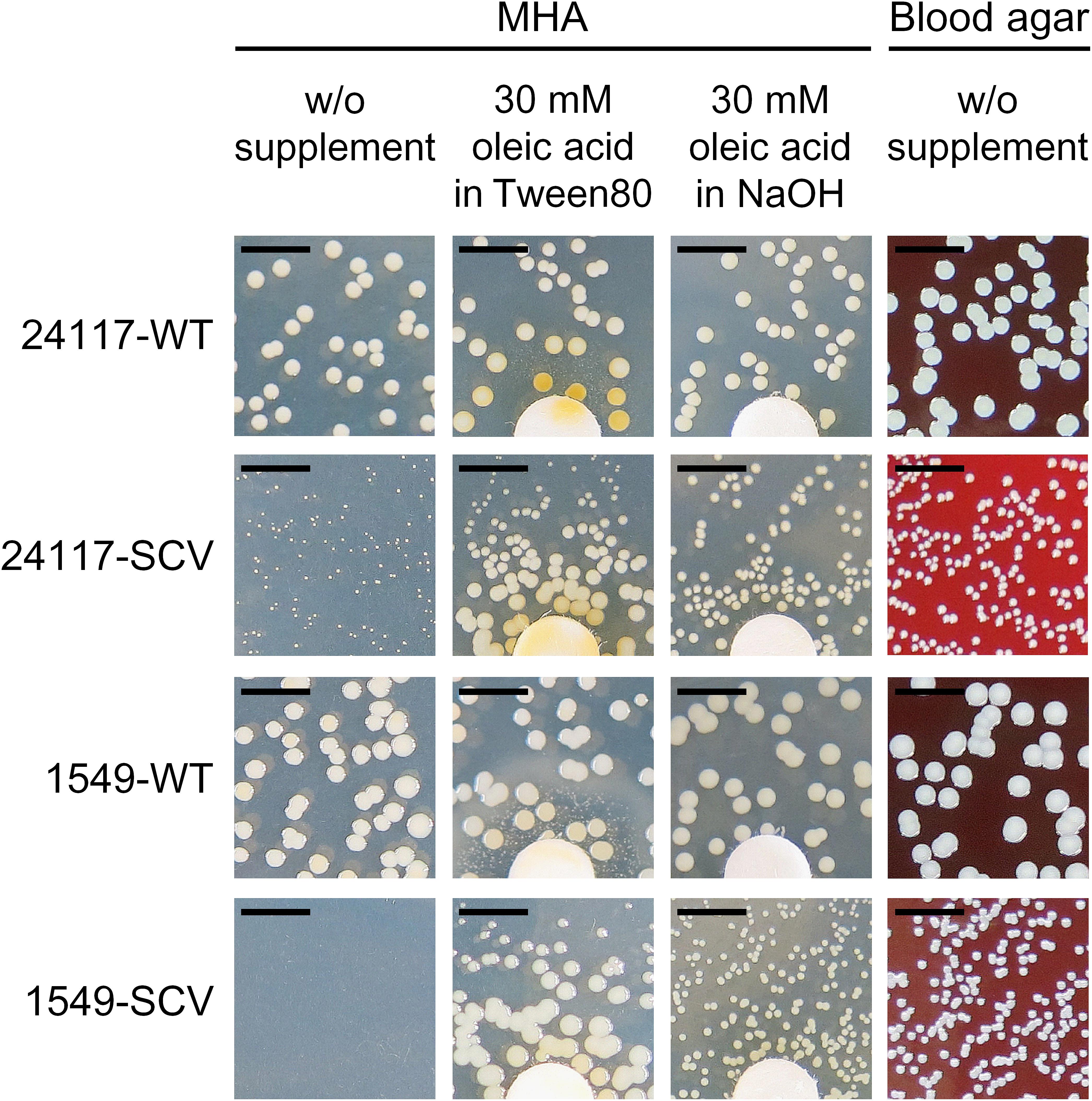
FIGURE 2. Phenotype of clinical S. aureus strain pairs after 48 h of incubation at 37°C on MHA with and without supplementation with oleic acid and on Columbia blood agar with 5% sheep blood; scale bar indicates 5 mm; diameter of the discs is 6.4 mm.
Pulsed-field gel electrophoresis fragment patterns of each strain triplet were identical or varied in only one band (data not shown). MLST and spa typing revealed that all phenotypes of triplet 1549 belonged to a single locus variant of ST45 and spa type t015 and the phenotypes of triplet 24117 belonged to ST15 and spa type t084, respectively. All results were confirmed by microarray analysis that further revealed agr type 1 for triplet 1549, and type 2 for triplet 24117, respectively (Supplementary Table 4). Virulence and regulatory profiles of each strain triplet were evaluated with microarray and were identical between the three phenotypes. WGS data were consistent with these findings (Supplementary Tables 4, 5).
Auxotrophism
Supplementation with hemin, menadione, or thymidine had no growth-supporting effect on the SCVs (data not shown), as shown as an example for the respective knockout mutant SCVs (Supplementary Figure 2). Supplementation with oleic acid solved in Tween 80, a synthetic ester from polyethoxylated sorbitan and oleic acid, restored the normal growth phenotype of both SCVs (Figure 2). Therefore, SCVs were further tested for auxotrophy for oleic acid and/or polyethylene oxide (PEG) and sorbitan. Auxotrophy was detected for oleic acid solved in NaOH (Figure 2), but not for PEG or sorbitan.
Whole-Genome Sequencing of Strain Triplets
Genome comparison of the three phenotypes of triplet 24117 revealed a total of seven alterations (point and frameshift mutations, Table 4): Due to a deletion of two nucleotides, 24117-SCV exhibited a frameshift mutation in the gene ecfT, which is part of the ecf module and encodes the ECF transporter transmembrane protein EcfT. This mutation caused a premature termination of the gene 19 amino acids downstream of the mutation locus resulting in a truncated protein with only 133 instead of 268 amino acids. While this mutation was still present in the 24117-REV, this strain showed an additional suppressor mutation (one nucleotide deletion) directly upstream of the first mutation locus resulting in the original WT open-reading frame with only one missing amino acid and a conservative amino acid exchange (Y→F). The genomic arrangement of the genes belonging to the ecf module in the genome of triplet 24117 is shown in Figure 3.
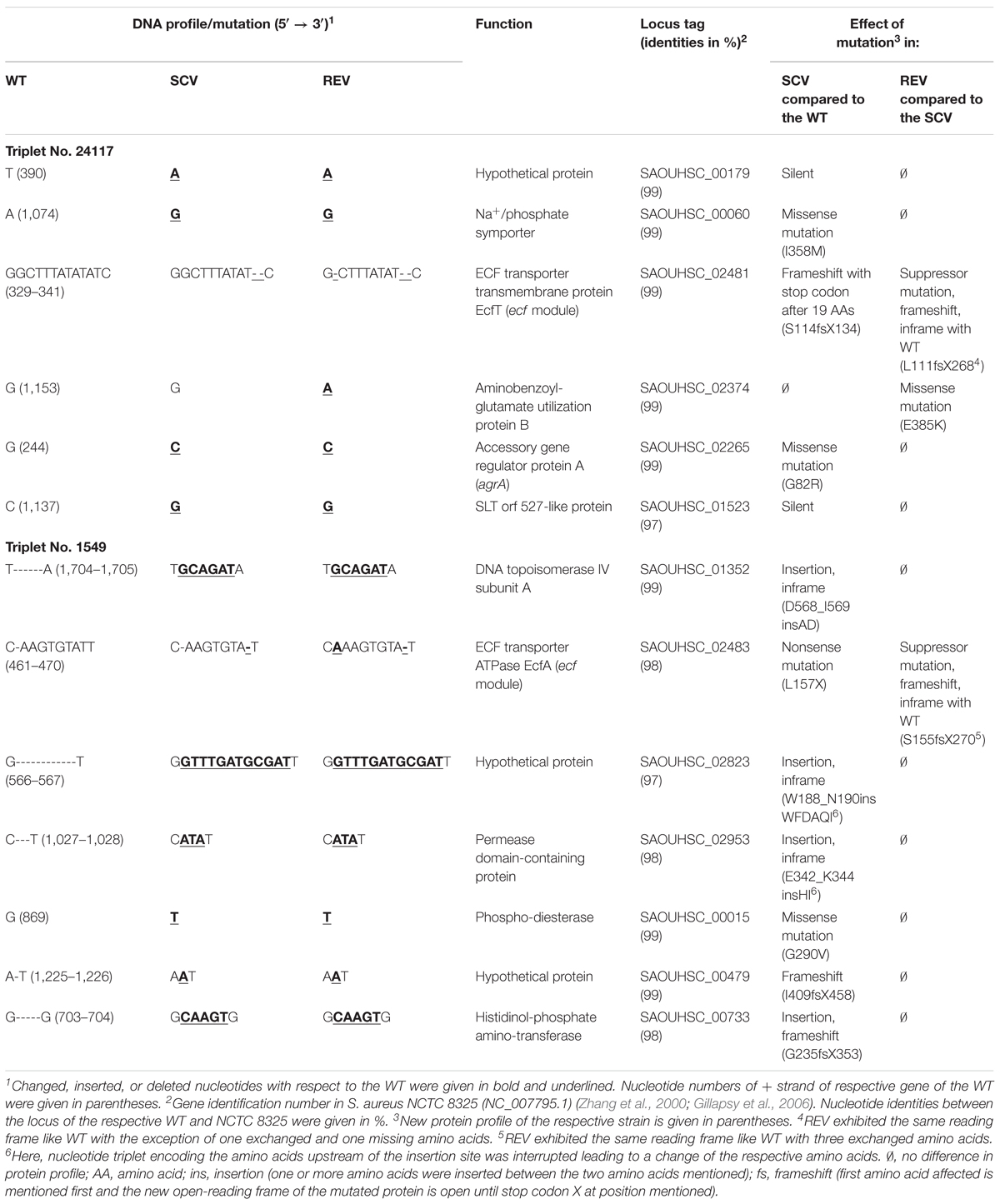
TABLE 4. Genetic alterations between the three phenotypes of two S. aureus strain triplets detected with a whole-genome sequencing approach.
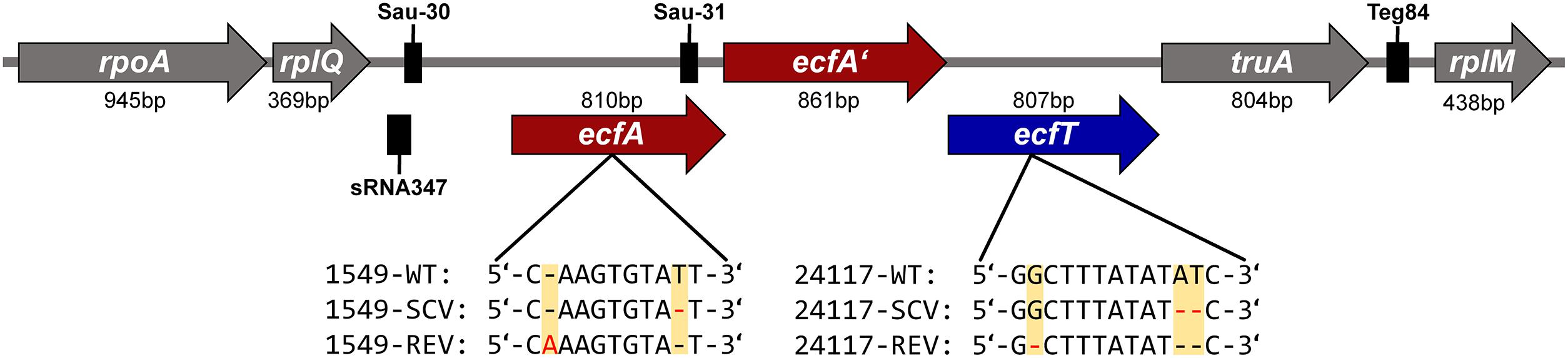
FIGURE 3. Gene arrangement of the ecf modules of S. aureus strain triplets 1549 and 24117. The mutation locus within the corresponding strain and the resulting DNA profiles of the respective phenotypes are indicated. Black boxes represent small non-protein-coding RNAs Sau-30 (63 nucleotides), Sau-31 (65 nucleotides), Teg84 (79 nucleotides), and sRNA347 (91 nucleotides) (Abu-Qatouseh et al., 2010; Beaume et al., 2010; Howden et al., 2013). Among the ecf modules of the WTs of 1549 and 24117, there were 137 mismatches, 1 insertion (1 nucleotide), and 1 deletion (1 nucleotide). ecfA and ecfA’, ECF transporter ATPases; ecfT, ECF transporter; rplM, 50S ribosomal protein L13; rplQ, 50S ribosomal protein L17; rpoA, DNA-directed RNA polymerase, alpha subunit; and truA, tRNA pseudouridine synthase A.
Besides these alterations, four other mutations occurring between 24117-WT and 24117-SCV were detected that were still present in 24117-REV without any additional suppressor mutation. All of them constituted point mutations with two of them being silent. Two other point mutations were functional missense mutations in the genes coding for a Na+/phosphate symporter (I358M) and the accessory gene regulator protein A (agrA) (G82R), a part of global virulence regulator agr. Another missense mutation could only be detected in 24117-REV, namely in a gene coding for the aminobenzoyl-glutamate utilization protein B (E385K).
WGS analysis of the triplet 1549 revealed eight alterations with seven of them occurring between 1549-WT and 1549-SCV (Table 4). Similar to the other strain triplet, 1549-SCV exhibited a nucleotide deletion in the ecf module. Here, the gene ecfA encoding for the ECF transporter ATP-binding protein EcfA was affected by a nonsense mutation resulting in a truncated EcfA protein in 1549-SCV with 156 instead of 269 amino acids. Again, similar to 24117-REV, 1549-REV exhibited this mutation as well as a suppressor mutation (one nucleotide insertion) directly upstream of the mutation locus resulting in a restored ecfA gene of 269 nucleotides, but with three amino acid exchanges. The genomic arrangement of the genes belonging to the ecf module was identical to strain triplet 1549 (Figure 3). In each case, the ecf module consisted of two adjacent genes encoding ecf transporter ATP-binding proteins EcfA and EcfA’ directly upstream of the ecfT gene. A sequence alignment of the ecf modules of 1549-WT and 24117-WT revealed 137 mismatches, one insertion (one nucleotide) and one deletion (one nucleotide). Within the ecfA gene, the sequence of the npcRNA Sau-31 (Abu-Qatouseh et al., 2010) could be identified exhibiting one and two nucleotide exchanges in triplets 1549 and 24117, respectively, when compared to NCTC 8325. Furthermore, for both triplets, 348 and 380 nucleotides upstream of ecfA, the npcRNA sequences Sau-30 (Abu-Qatouseh et al., 2010) and sRNA347 (Howden et al., 2013) could be found (Figure 3). Compared to NCTC 8325, there were three nucleotide exchanges in Sau-30 in triplet 1549 and one in triplet 24117, whereas sequences encoding sRNA347 were identical to NCTC 8325.
All other nucleotide alterations between 1549-WT and 1549-SCV could also be identified in 1549-REV without any suppressor mutation and comprise a nucleotide exchange that resulted in a conservative missense mutation in a phosphodiesterase (G290V). Furthermore, a total of five loci showed insertions of a number of 1–12 nucleotides in the genome of 1549-SCV resulting in (i) a frameshift within a hypothetical protein, (ii) an insertion of two amino acids followed by a frameshift within a histidinol-phosphate aminotransferase, and (iii) three different inframe insertions in the DNA topoisomerase IV subunit A, within a permease domain-containing protein and another hypothetical protein, respectively (Table 4).
Screening for Alterations Within the ecf Module and the FASII Pathway Genes
Another three isogenic strain triplets comprising SCVs with dependency on oleic acid were screened for alterations within genes of the ecf module and genes linked to the FASII pathway. For all genes analyzed, mutations between WT and SCV phenotypes could not be detected.
Discussion
For clinical SCVs, only few underlying genetic alterations for distinct auxotrophisms were hitherto identified, whereas the genetic basis for the phenotypic switch remains unclear for the majority of clinical SCVs (von Eiff et al., 1997b; Schaaff et al., 2003; Chatterjee et al., 2008; Lannergård et al., 2008; Abu-Qatouseh et al., 2010; Köser et al., 2012; Lin et al., 2016).
The SCVs investigated here did not show any of the well-characterized auxotrophies for hemin, menadione, and/or thymidine, but dependency on monounsaturated oleic acid. 1549-SCV was originally described as heme auxotroph (Kriegeskorte et al., 2014b), which becomes explainable due to the use of Tween 80 as solving reagent for hemin in earlier studies since this dispersing agent contains a complex mixtures of polyoxyethylene ethers with approximately 70% oleic acid as part of its fatty acid composition. While 1549-SCV and 1549-WT were recovered from the fat digesting bile of a gall bladder content, there is no obvious explanation for the fatty acid-auxotrophy of 24117-SCV. Already in the first descriptions of fatty acid-auxotrophic S. aureus SCVs, it has been suspected that dependency on fatty acids might be due to defects in bacterial lipid synthesis accompanied by impaired electron transport (Kaplan and Dye, 1976). In 2016, Lin et al. (2016) detected a conservative point mutation within the fatty acid synthesis (FASII) pathway gene fabF encoding the β-ketoacyl-ACP synthase for a fatty acid-dependent S. aureus SCV when compared to the related WT. Recently, triclosan-selected SCVs were shown to exhibit mutations within fabI, eventually combined with a mutation within fabD (Bazaid et al., 2018).
Using a WGS approach, we revealed alternate genetic variations between parental WTs, their corresponding SCVs, and the reverted normal-growing REVs that were not directly linked to the FASII pathway. In particular, both SCVs exhibited mutations of genes located within the ecf module. These mutations induced transcriptional stops, which resulted in significantly truncated ECF proteins in the SCVs and, most likely, with loss of function. However, in both cases, the respective mutation was still present in the normal-growing REV, but was almost fully restored by a compensatory intragenic suppressor mutation directly upstream. Suppressor mutations, which are defined as second mutations that counteracts the effects of first original mutations and, thereby, resulting in a restored phenotype (Michels, 2002), were shown to occur at higher rates than true reversions (Levin et al., 2000; Poon et al., 2005; Lannergård et al., 2008).
ECF transporters are part of the large ABC-transporter family and mediate the uptake of essential vitamins and metal ions in many prokaryotes, thus being necessary for cellular growth and metabolism, in particular for those bacteria lacking the pathways for folate, biotin, and thiamin biosynthesis, respectively (Konings, 2006; Slotboom, 2014).
The ECF transporter types hitherto described comprise three components: (i) a membrane-embedded, substrate-binding protein (S component, EcfS), (ii) an energy-coupling element consisting of one or two cytosolic ATP-binding proteins (EcfA and EcfA’), and (iii) a transmembrane transport protein (EcfT) (Figure 3). There are two types of ECF transporters with type-1 transporters encoding all components in the same module and being specific for only one substrate and type-2 having the EcfAA’T module encoded in one module but the genes encoding for different exchangeable EcfS are scattered around the chromosome (Slotboom, 2014). For S. aureus, less data are available for this transporter family. However, according to the WGS data, it can be concluded that the ECF transporters analyzed here belong to the type-2 transporters, as no genes encoding for EcfS were found in close proximity up- or downstream of the genes ecfA, ecfA’, and ecfT. In fact, for both S. aureus strain triplets, the gene arrangement within the ecf modules was identical to an ecf module type described in Bacillus subtilis (Rodionov et al., 2009).
To date, 21 different EcfS have been identified, among these pantothenic acid, also called vitamin B5 (Rodionov et al., 2009; Slotboom, 2014). This vitamin is necessary for the biosynthesis of the ubiquitous coenzyme A (CoA). CoA is essential for biosynthesis of fatty acids (Begley et al., 2001) and a lack in CoA levels will thus inevitably lead to complications in fatty acid synthesis. Moreover, CoA plays also a key role in the energy production of the cell. Once in the form of acetyl-CoA, it enters the TCA cycle and the electrons obtained are used during oxidative phosphorylation for the generation of ATP. For 1549-SCV and other clinical and genetically defined SCVs, a down-regulation of TCA cycle activity could already be shown (Kohler et al., 2003; Chatterjee et al., 2005, 2007; Gaupp et al., 2010; Kriegeskorte et al., 2014b). Therefore, for both SCVs, one can speculate that the mutations within the ecf module might be also the cause for a reduced uptake of vitamin B5, resulting in insufficient amounts of intracellular CoA. Thus, mutations within the ecf module may lead to both the impaired FASII metabolism and the downregulated TCA cycle as explanation for the slow growth of the analyzed SCVs.
While being essential in fatty acid synthesis, CoA is not needed for the incorporation of fatty acids into the membrane in the case of S. aureus (Parsons et al., 2011, 2014). Uptake of supplemented oleic acid may therefore restore membrane synthesis in oleic acid-auxotrophic SCVs. Accordingly, higher amounts of CoA are available for entering the TCA cycle and contributing to the reversion of the phenotype. Furthermore, bacilli deficient for pantothenate were found to be limited in growth (Baigori et al., 1991) and their defective uptake of glutamic acid was shown to be reversible by supplementation of unsaturated fatty acids (Holden and Bunch, 1972).
Since for type-2 ECF transporters multiple EcfS can use the same EcfAA’T module to form an active transporter complex (Rodionov et al., 2009), there might be also a lack of other substrates being responsible for the SCV phenotype such as riboflavin, niacin, and biotin. Riboflavin (vitamin B2), an essential component of the basic metabolism, represents a precursor of coenzyme flavin adenine dinucleotie (FAD) (Vitreschak et al., 2002; LeBlanc et al., 2017). Niacin (vitamin B3), a component of NAD, as well as biotin (vitamin B7) are also known to be required or to constitute stimulatory factors for growth of S. aureus (Peterson and Peterson, 1945). Besides CoA, NAD and FAD are also important cofactors needed in the TCA cycle and, thus, maybe also implicated in growth characteristics of the tested SCVs.
Staphylococcal SCVs not defective for fatty acids may also exhibit alterations in ecf modules mediating the uptake of other B-group vitamins. Auxotrophy for thiamine (vitamin B1) was already identified being responsible for SCVs (Ziv and Sompolinsky, 1976; Acar et al., 1978). The same is probably the case for the ECF substrate folate (vitamin B9). In a previous study, we could show a down-regulation of a protein cluster involved in the folate metabolism for a clinically derived SCV (Kriegeskorte et al., 2011).
By contrast, other SCVs dependent on oleic acid screened in this study showed neither mutations within the ecf module nor alterations in genes linked to the FASII pathway. This was also the case for genes accC, accD, and plsX for which genetically defined knockout mutants were already proven to exhibit auxotrophy for fatty acids (Table 5; Parsons et al., 2011, 2013, 2014). This underlines the high diversity of potential mutation loci leading to the phenotype switch. Furthermore, high reversion rates of clinical SCVs may indicate phenotypical heterogeneity based on genetic alterations being not yet detectable (Avery, 2006). Besides the genomic background, regulation processes, e.g., via differentially expressed npcRNAs are also involved in SCV formation (Abu-Qatouseh et al., 2010). ECF transporter can be coupled with npcRNAs (riboswitches) (Rodionov et al., 2009). It was previously demonstrated that npcRNA Sau-31 is developmentally regulated and not expressed in a clinical SCV during stationary phase (Abu-Qatouseh et al., 2010) indicating potential impairment of ECF substrate uptake in the according SCV. This might be a hint for the involvement of these npcRNAs in the SCV phenotype switch.
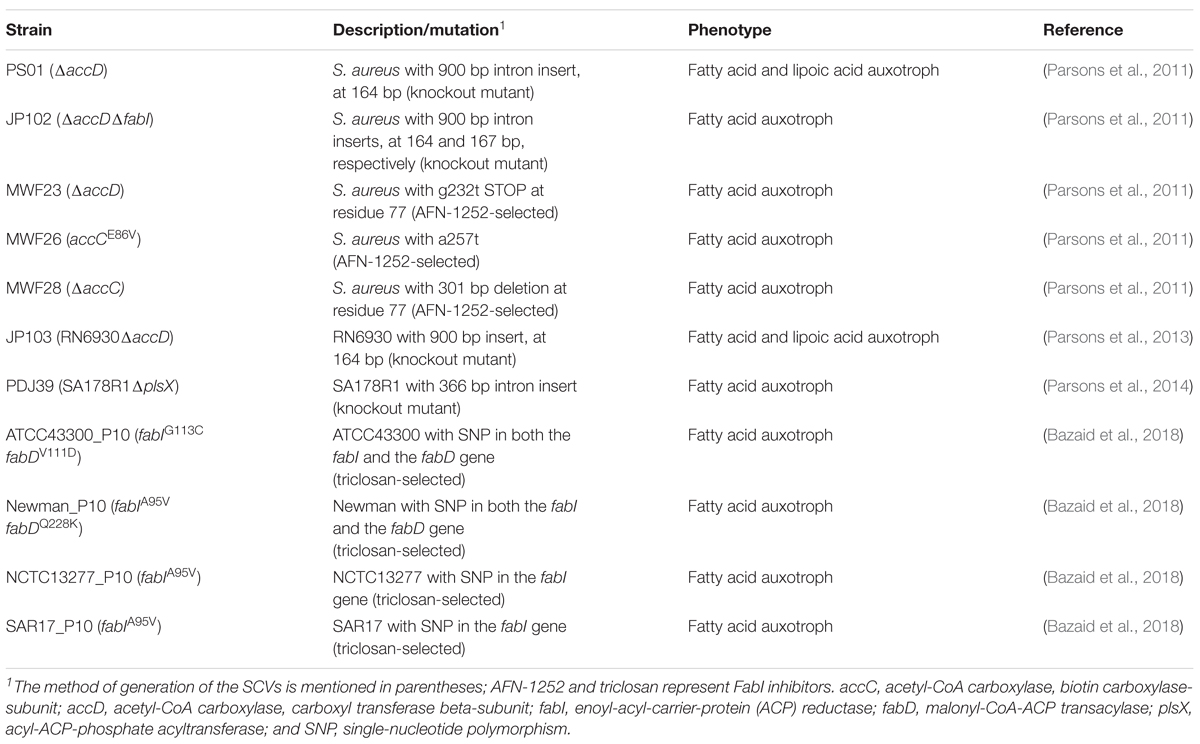
TABLE 5. Genetically defined and FabI inhibitor selected strains with defects in fatty acid incorporation or biosynthesis (FASII).
By WGS, we were furthermore able to find probable genetic drivers for the variable hemolysis behavior between the REV phenotypes of the two strain triplets. Whereas 1549-REV showed a normally restored β-hemolysis after 48 h of incubation at 37°C, 24117-REV only exhibited weak hemolysis under the same conditions. Sequence analysis of the strain triplet 24117 revealed no alterations in the genes hla, hlb, hld, and hlg (encoding for α-, β-, δ-, and γ-hemolysin), but a mutation in the gene agrA occurring in 24117-SCV and 24117-REV. This mutation leads to a non-conservative amino acid exchange (G82R) in AgrA, part of the global virulence regulator agr (Table 4). AgrA is responsible for the activation of agr promotor P3 and therefore for the transcription of RNAIII (Novick et al., 1993), which also encodes the agr-regulated δ-hemolysin hld (Janzon et al., 1989). RNAIII stimulates the translational regulation of proteolytic enzymes and several exotoxins like hla encoded α-hemolysin (Morfeldt et al., 1995) and its downregulation is shown to be a characteristic trait in SCVs (Proctor et al., 2014). The accompanying loss of hemolytic activity was recently shown to be caused by a point mutation within the agrC gene of S. aureus resulting in an amino acid substitution that leads to a destabilization of the AgrC–AgrA interaction (Mairpady Shambat et al., 2016). Moreover, it was already shown that another mutation in agrA is responsible for the non-hemolytic phenotype of laboratory strain RN4220 due to defective translation of hla and hld (Traber and Novick, 2006). Accordingly, in 24117-REV, the mutated agrA may most likely contributed to the poor hemolysis.
Comparison of whole genomes of the included strains revealed the close relationship between the isogenic, but phenotypically different strains showing only seven to eight alterations per strain triplet. The number of the genetic events was found to be similar to other WGS approaches tracking the mutability of isogenic S. aureus strains. However, in many cases, isogenic strains were isolated before and after extensive chemotherapy resulting in approx. 2–30 mutations most likely driven by adaptation to the antibiotics and by pathogenesis mechanisms (Mwangi et al., 2007; Lannergård et al., 2011; Peleg et al., 2012; Lin et al., 2016).
Our study emphasized that WGS-based identification of all mutations leading to a phenotype switch may prove workable, while generation of knockout mutants can be expedient only for analyzing one or a few genes of interest. Furthermore, if the gene affected is not directly linked to the respective auxotrophism, knockout mutant generation is ineffectual. This should be considered especially for undefined auxotrophies.
Conclusion
This study demonstrated that the genetic background of SCVs is highly diverse and that the detection of underlying genes inducing the phenotypic switch is mandatory for a better understanding of this phenotype. Furthermore, the study reveals a novel dynamic process of phenotype switching between naturally occurring SCVs and REVs displaying the normal phenotype. WGS seems to be the most reliable and efficient tool for detecting the underlying genetic mechanisms responsible for the SCV formation and the way back to the WT phenotype. Alterations within ECF transporters as detected here display further drivers for the phenotype switch from WT to SCV by limitation of cellular nutrient uptake, thus leading to a downregulation of the TCA cycle activity. Moreover, this study underlines the importance of including clinically derived strains when studying the genetic background of phenotypic variation.
Author Contributions
KB designed the study concept. AK performed initial studies. NS designed the experiments, performed laboratory work, evaluated the data, drafted, and wrote the manuscript. UK contributed in data evaluation and writing the manuscript. MD performed cloning experiments. JS provided scientific support regarding whole-genome sequencing. RP, GP, and CE provided scientific support regarding SCVs and data interpretation. All authors have read and approved the final draft of the article.
Funding
This work was supported in part by the Deutsche Forschungsgemeinschaft (DFG) within the SPP1316 to KB (BE 2546/1-2).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank the Technology Development group, in particular Robert Sebra, from the Icahn Institute for Genomics and Multiscale Biology at the Icahn School of Medicine at Mount Sinai for PacBio sequencing and assembly and Christian Ruckert from the Institute of Genomics at the University Hospital of Münster for bioinformatical support. Furthermore, we are grateful to the GenDB support team for technical assistance and access to resources financially supported by the BMBF [FKZ 031A533] within the de.NBI network. Special thanks to Melanie Bach and Daniela Kuhn for excellent technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01863/full#supplementary-material
References
Abu-Qatouseh, L., Chinni, S., Seggewiß, J., Proctor, R. A., Brosius, J., Rozhdestvensky, T. S., et al. (2010). Identification of differentially expressed small non-protein-coding RNAs in Staphylococcus aureus displaying both the normal and the small-colony variant phenotype. J. Mol. Med. 88, 565–575. doi: 10.1007/s00109-010-0597-2
Acar, J. F., Goldstein, F. W., and Lagrange, P. (1978). Human infections caused by thiamine- or menadione-requiring Staphylococcus aureus. J. Clin. Microbiol. 8, 142–147.
Avery, S. V. (2006). Microbial cell individuality and the underlying sources of heterogeneity. Nat. Rev. Microbiol. 4, 577–587. doi: 10.1038/nrmicro1460
Baigori, M., Grau, R., Morbidoni, H. R., and de Mendoza, D. (1991). Isolation and characterization of Bacillus subtilis mutants blocked in the synthesis of pantothenic acid. J. Bacteriol. 173, 4240–4242. doi: 10.1128/JB.173.13.4240-4242.1991
Bartels, M. D., Petersen, A., Worning, P., Nielsen, J. B., Larner-Svensson, H., Johansen, H. K., et al. (2014). Comparing whole-genome sequencing with sanger sequencing for spa typing of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 52, 4305–4308. doi: 10.1128/JCM.01979-14
Bates, D. M., von Eiff, C., McNamara, P. J., Peters, G., Yeaman, M. R., Bayer, A. S., et al. (2003). Staphylococcus aureus menD and hemB mutants are as infective as the parent strains, but the menadione biosynthetic mutant persists within the kidney. J. Infect. Dis. 187, 1654–1661. doi: 10.1086/374642
Bazaid, A. S., Forbes, S., Humphreys, G. J., Ledder, R. G., O’Cualain, R., and McBain, A. J. (2018). Fatty acid supplementation reverses the small colony variant phenotype in triclosan-adapted Staphylococcus aureus: genetic, proteomic and phenotypic analyses. Sci. Rep. 8:3876. doi: 10.1038/s41598-018-21925-6
Beaume, M., Hernandez, D., Farinelli, L., Deluen, C., Linder, P., Gaspin, C., et al. (2010). Cartography of methicillin-resistant S. aureus transcripts: detection, orientation and temporal expression during growth phase and stress conditions. PLoS One 5:e10725. doi: 10.1371/journal.pone.0010725
Becker, K., Al Laham, N., Fegeler, W., Proctor, R. A., Peters, G., and von Eiff, C. (2006). Fourier-transform infrared spectroscopic analysis is a powerful tool for studying the dynamic changes in Staphylococcus aureus small-colony variants. J. Clin. Microbiol. 44, 3274–3278. doi: 10.1128/JCM.00847-06
Begley, T. P., Kinsland, C., and Strauss, E. (2001). The biosynthesis of coenzyme A in bacteria. Vitam. Horm. 61, 157–171. doi: 10.1016/S0083-6729(01)61005-7
Cao, S., Huseby, D. L., Brandis, G., and Hughes, D. (2017). Alternative evolutionary pathways for drug-resistant small colony variant mutants in Staphylococcus aureus. mBio 8:e00358-17. doi: 10.1128/mBio.00358-17
Chatterjee, I., Becker, P., Grundmeier, M., Bischoff, M., Somerville, G. A., Peters, G., et al. (2005). Staphylococcus aureus Clpc is required for stress resistance, aconitase activity, growth recovery, and death. J. Bacteriol. 187, 4488–4496. doi: 10.1128/JB.187.13.4488-4496.2005
Chatterjee, I., Herrmann, M., Proctor, R. A., Peters, G., and Kahl, B. C. (2007). Enhanced post-stationary-phase survival of a clinical thymidine-dependent small-colony variant of Staphylococcus aureus results from lack of a functional tricarboxylic acid cycle. J. Bacteriol. 189, 2936–2940. doi: 10.1128/JB.01444-06
Chatterjee, I., Kriegeskorte, A., Fischer, A., Deiwick, S., Theimann, N., Proctor, R. A., et al. (2008). In vivo mutations of thymidylate synthase (encoded by thyA) are responsible for thymidine dependency in clinical small-colony variants of Staphylococcus aureus. J. Bacteriol. 190, 834–842. doi: 10.1128/JB.00912-07
Chin, C.-S., Alexander, D. H., Marks, P., Klammer, A. A., Drake, J., Heiner, C., et al. (2013). Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 10, 563–569. doi: 10.1038/nmeth.2474
Darling, A. C. E., Mau, B., Blattner, F. R., and Perna, N. T. (2004). Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 14, 1394–1403. doi: 10.1101/gr.2289704
Dean, M. A., Olsen, R. J., Long, S. W., Rosato, A. E., and Musser, J. M. (2014). Identification of point mutations in clinical Staphylococcus aureus strains that produce small-colony variants auxotrophic for menadione. Infect. Immun. 82, 1600–1605. doi: 10.1128/IAI.01487-13
Edwards, A. M. (2012). Phenotype switching is a natural consequence of Staphylococcus aureus replication. J. Bacteriol. 194, 5404–5412. doi: 10.1128/JB.00948-12
Gao, W., Chua, K., Davies, J. K., Newton, H. J., Seemann, T., Harrison, P. F., et al. (2010). Two novel point mutations in clinical Staphylococcus aureus reduce linezolid susceptibility and switch on the stringent response to promote persistent infection. PLoS Pathog. 6:e1000944. doi: 10.1371/journal.ppat.1000944
Garcia, L. G., Lemaire, S., Kahl, B. C., Becker, K., Proctor, R. A., Denis, O., et al. (2013). Antibiotic activity against small-colony variants of Staphylococcus aureus: review of in vitro, animal and clinical data. J. Antimicrob. Chemother. 68, 1455–1464. doi: 10.1093/jac/dkt072
Gaupp, R., Schlag, S., Liebeke, M., Lalk, M., and Götz, F. (2010). Advantage of upregulation of succinate dehydrogenase in Staphylococcus aureus biofilms. J. Bacteriol. 192, 2385–2394. doi: 10.1128/JB.01472-09
Gillapsy, A. F., Worrell, V., Orvis, J., Roe, B. A., Dyer, D. W., and Iandolo, J. J. (2006). “The Staphylococcus aureus NCTC 8325 Genome,” in Gram-Positive Pathogens, 2nd Edn, eds V. A. Fischetti, R. Novick, J. J. Ferreti, D. A. Portnoy, and J. I. Rood (Washington, DC: ASM Press), 381–412. doi: 10.1128/9781555816513.ch32
Goering, R. V., and Winters, M. A. (1992). Rapid method for epidemiological evaluation of gram-positive cocci by field inversion gel electrophoresis. J. Clin. Microbiol. 30, 577–580.
Gómez-González, C., Acosta, J., Villa, J., Barrado, L., Sanz, F., Orellana, M. A., et al. (2010). Clinical and molecular characteristics of infections with CO2-dependent small-colony variants of Staphylococcus aureus. J. Clin. Microbiol. 48, 2878–2884. doi: 10.1128/JCM.00520-10
Hammer, N. D., Reniere, M. L., Cassat, J. E., Zhang, Y., Hirsch, A. O., Indriati Hood, M., et al. (2013). Two heme-dependent terminal oxidases power Staphylococcus aureus organ-specific colonization of the vertebrate host. mBio 4:e00241-13. doi: 10.1128/mBio.00241-13
Holden, J. T., and Bunch, J. M. (1972). stimulation by fatty acids of amino acid accumulation in pantothenic acid depleted Lactobacillus plantarum. Biochem. Biophys. Res. Commun. 46, 437–442. doi: 10.1016/S0006-291X(72)80157-8
Howden, B. P., Beaume, M., Harrison, P. F., Hernandez, D., Schrenzel, J., Seemann, T., et al. (2013). Analysis of the small RNA transcriptional response in multidrug-resistant Staphylococcus aureus after antimicrobial exposure. Antimicrob. Agents Chemother. 57, 3864–3874. doi: 10.1128/AAC.00263-13
Janzon, L., Löfdahl, S., and Arvidson, S. (1989). Identification and nucleotide sequence of the delta-lysin gene, hld, adjacent to the accessory gene regulator (agr) of Staphylococcus aureus. Mol. Gen. Genet. 219, 480–485. doi: 10.1007/BF00259623
Jolley, K. A., and Maiden, M. C. (2010). BIGSdb: scalable analysis of bacterial genome variation at the population level. BMC Bioinformatics 11:595. doi: 10.1186/1471-2105-11-595
Kahl, B., Herrmann, M., Everding, A. S., Koch, H. G., Becker, K., Harms, E., et al. (1998). Persistent infection with small colony variant strains of Staphylococcus aureus in patients with cystic fibrosis. J. Infect. Dis. 177, 1023–1029. doi: 10.1086/515238
Kahl, B. C., Becker, K., and Löffler, B. (2016). Clinical significance and pathogenesis of staphylococcal small colony variants in persistent infections. Clin. Microbiol. Rev. 29, 401–427. doi: 10.1128/CMR.00069-15
Kaplan, M. L., and Dye, W. (1976). Growth requirements of some small-colony-forming variants of Staphylococcus aureus. J. Clin. Microbiol. 4, 343–348.
Kohler, C., von Eiff, C., Liebeke, M., McNamara, P. J., Lalk, M., Proctor, R. A., et al. (2008). A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus. J. Bacteriol. 190, 6351–6364. doi: 10.1128/JB.00505-08
Kohler, C., von Eiff, C., Peters, G., Proctor, R. A., Hecker, M., and Engelmann, S. (2003). Physiological characterization of a heme-deficient mutant of Staphylococcus aureus by a proteomic approach. J. Bacteriol. 185, 6928–6937. doi: 10.1128/JB.185.23.6928-6937.2003
Konings, W. N. (2006). Microbial transport: adaptations to natural environments. Antonie Van Leeuwenhoek 90, 325–342. doi: 10.1007/s10482-006-9089-3
Köser, C. U., Holden, M. T. G., Ellington, M. J., Cartwright, E. J. P., Brown, N. M., Ogilvy-Stuart, A. L., et al. (2012). Rapid whole-genome sequencing for investigation of a neonatal MRSA outbreak. N. Engl. J. Med. 366, 2267–2275. doi: 10.1056/NEJMoa1109910
Kriegeskorte, A., Block, D., Drescher, M., Windmüller, N., Mellmann, A., Baum, C., et al. (2014a). Inactivation of thyA in Staphylococcus aureus attenuates virulence and has a strong impact on metabolism and virulence gene expression. mBio 5:e01447-14. doi: 10.1128/mBio.01447-14
Kriegeskorte, A., Grubmüller, S., Huber, C., Kahl, B. C., von Eiff, C., Proctor, R. A., et al. (2014b). Staphylococcus aureus small colony variants show common metabolic features in central metabolism irrespective of the underlying auxotrophism. Front. Cell. Infect. Microbiol. 4:141. doi: 10.3389/fcimb.2014.00141
Kriegeskorte, A., König, S., Sander, G., Pirkl, A., Mahabir, E., Proctor, R. A., et al. (2011). Small colony variants of Staphylococcus aureus reveal distinct protein profiles. Proteomics 11, 2476–2490. doi: 10.1002/pmic.201000796
Lannergård, J., Cao, S., Norström, T., Delgado, A., Gustafson, J. E., and Hughes, D. (2011). Genetic complexity of fusidic acid-resistant small colony variants (SCV) in Staphylococcus aureus. PLoS One 6:e28366. doi: 10.1371/journal.pone.0028366
Lannergård, J., von Eiff, C., Sander, G., Cordes, T., Seggewiß, J., Peters, G., et al. (2008). Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 52, 4017–4022. doi: 10.1128/AAC.00668-08
LeBlanc, J. G., Chain, F., Martín, R., Bermúdez-Humarán, L. G., Courau, S., and Langella, P. (2017). Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 16:79. doi: 10.1186/s12934-017-0691-z
Levin, B. R., Perrot, V., and Walker, N. (2000). Compensatory mutations, antibiotic resistance and the population genetics of adaptive evolution in bacteria. Genetics 154, 985–997.
Lin, Y.-T., Tsai, J.-C., Yamamoto, T., Chen, H.-J., Hung, W.-C., Hsueh, P.-R., et al. (2016). Emergence of a small colony variant of vancomycin-intermediate Staphylococcus aureus in a patient with septic arthritis during long-term treatment with daptomycin. J. Antimicrob. Chemother. 71, 1807–1814. doi: 10.1093/jac/dkw060
Mairpady Shambat, S., Siemens, N., Monk, I. R., Mohan, D. B., Mukundan, S., Krishnan, K. C., et al. (2016). A point mutation in AgrC determines cytotoxic or colonizing properties associated with phenotypic variants of ST22 MRSA strains. Sci. Rep. 6:31360. doi: 10.1038/srep31360
Meyer, F., Goesmann, A., McHardy, A. C., Bartels, D., Bekel, T., Clausen, J., et al. (2003). GenDB-an open source genome annotation system for prokaryote genomes. Nucleic Acids Res. 31, 2187–2195. doi: 10.1093/nar/gkg312
Michels, C. A. (ed.) (2002). “Suppression Analysis,” in Genetic Techniques for Biological Research. Hoboken, NJ: John Wiley & Sons, Ltd, 91–98. doi: 10.1002/0470846623.ch8
Morfeldt, E., Taylor, D., von Gabain, A., and Arvidson, S. (1995). Activation of alpha-toxin translation in Staphylococcus aureus by the trans-encoded antisense RNA, RNAIII. EMBO J. 14, 4569–4577.
Mwangi, M. M., Wu, S. W., Zhou, Y., Sieradzki, K., de Lencastre, H., Richardson, P., et al. (2007). Tracking the in vivo evolution of multidrug resistance in Staphylococcus aureus by whole-genome sequencing. Proc. Natl. Acad. Sci. U.S.A. 104, 9451–9456. doi: 10.1073/pnas.0609839104
Novick, R. P., Ross, H. F., Projan, S. J., Kornblum, J., Kreiswirth, B., and Moghazeh, S. (1993). Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. EMBO J. 12, 3967–3975.
Painter, K. L., Strange, E., Parkhill, J., Bamford, K. B., Armstrong-James, D., and Edwards, A. M. (2015). Staphylococcus aureus adapts to oxidative stress by producing H2O2-resistant small-colony variants via the SOS response. Infect. Immun. 83, 1830–1844. doi: 10.1128/IAI.03016-14
Parsons, J. B., Frank, M. W., Jackson, P., Subramanian, C., and Rock, C. O. (2014). Incorporation of extracellular fatty acids by a fatty acid kinase-dependent pathway in Staphylococcus aureus. Mol. Microbiol. 92, 234–245. doi: 10.1111/mmi.12556
Parsons, J. B., Frank, M. W., Rosch, J. W., and Rock, C. O. (2013). Staphylococcus aureus fatty acid auxotrophs do not proliferate in mice. Antimicrob. Agents Chemother. 57, 5729–5732. doi: 10.1128/AAC.01038-13
Parsons, J. B., Frank, M. W., Subramanian, C., Saenkham, P., and Rock, C. O. (2011). Metabolic basis for the differential susceptibility of Gram-positive pathogens to fatty acid synthesis inhibitors. Proc. Natl. Acad. Sci. U.S.A. 108, 15378–15383. doi: 10.1073/pnas.1109208108
Peleg, A. Y., Miyakis, S., Ward, D. V., Earl, A. M., Rubio, A., Cameron, D. R., et al. (2012). Whole genome characterization of the mechanisms of daptomycin resistance in clinical and laboratory derived isolates of Staphylococcus aureus. PLoS One 7:e28316. doi: 10.1371/journal.pone.0028316
Peterson, W. H., and Peterson, M. S. (1945). Relation of bacteria to vitamin and other growth factors. Bacteriol. Rev. 9, 49–109.
Poon, A., Davis, B. H., and Chao, L. (2005). The coupon collector and the suppressor mutation: estimating the number of compensatory mutations by maximum likelihood. Genetics 170, 1323–1332. doi: 10.1534/genetics.104.037259
Proctor, R. A., Kriegeskorte, A., Kahl, B. C., Becker, K., Löffler, B., and Peters, G. (2014). Staphylococcus aureus Small Colony Variants (SCVs): a road map for the metabolic pathways involved in persistent infections. Front. Cell. Infect. Microbiol. 4:99. doi: 10.3389/fcimb.2014.00099
Proctor, R. A., van Langevelde, P., Kristjansson, M., Maslow, J. N., and Arbeit, R. D. (1995). Persistent and relapsing infections associated with small-colony variants of Staphylococcus aureus. Clin. Infect. Dis. 20, 95–102. doi: 10.1093/clinids/20.1.95
Proctor, R. A., von Eiff, C., Kahl, B. C., Becker, K., McNamara, P., Herrmann, M., et al. (2006). Small colony variants: a pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat. Rev. Microbiol. 4, 295–305. doi: 10.1038/nrmicro1384
Rodionov, D. A., Hebbeln, P., Eudes, A., ter Beek, J., Rodionova, I. A., Erkens, G. B., et al. (2009). A novel class of modular transporters for vitamins in prokaryotes. J. Bacteriol. 191, 42–51. doi: 10.1128/JB.01208-08
Sassi, M., Augagneur, Y., Mauro, T., Ivain, L., Chabelskaya, S., Hallier, M., et al. (2015). SRD: a Staphylococcus regulatory RNA database. RNA 21, 1005–1017. doi: 10.1261/rna.049346.114
Schaaff, F., Bierbaum, G., Baumert, N., Bartmann, P., and Sahl, H.-G. (2003). Mutations are involved in emergence of aminoglycoside-induced small colony variants of Staphylococcus aureus. Int. J. Med. Microbiol. 293, 427–435. doi: 10.1078/1438-4221-00282
Sherris, J. C. (1952). Two small colony variants of Staph. aureus isolated in pure culture from closed infected lesions and their carbon dioxide requirements. J. Clin. Pathol. 5, 354–355. doi: 10.1136/jcp.5.4.354
Slifkin, M., Merkow, L. P., Kreuzberger, S. A., Engwall, C., and Pardo, M. (1971). Characterization of CO2 dependent microcolony variants of Staphylococcus aureus. Am. J. Clin. Pathol. 56, 584–592. doi: 10.1093/ajcp/56.5.584
Slotboom, D. J. (2014). Structural and mechanistic insights into prokaryotic energy-coupling factor transporters. Nat. Rev. Microbiol. 12, 79–87. doi: 10.1038/nrmicro3175
Tenover, F. C., Arbeit, R. D., Goering, R. V., Mickelsen, P. A., Murray, B. E., Persing, D. H., et al. (1995). Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 33, 2233–2239.
Traber, K., and Novick, R. (2006). A slipped-mispairing mutation in AgrA of laboratory strains and clinical isolates results in delayed activation of agr and failure to translate δ- and α-haemolysins. Mol. Microbiol. 59, 1519–1530. doi: 10.1111/j.1365-2958.2006.04986.x
Tuchscherr, L., Heitmann, V., Hussain, M., Viemann, D., Roth, J., von Eiff, C., et al. (2010). Staphylococcus aureus small-colony variants are adapted phenotypes for intracellular persistence. J. Infect. Dis. 202, 1031–1040. doi: 10.1086/656047
Tuchscherr, L., Medina, E., Hussain, M., Völker, W., Heitmann, V., Niemann, S., et al. (2011). Staphylococcus aureus phenotype switching: an effective bacterial strategy to escape host immune response and establish a chronic infection. EMBO Mol. Med. 3, 129–141. doi: 10.1002/emmm.201000115
Vitreschak, A. G., Rodionov, D. A., Mironov, A. A., and Gelfand, M. S. (2002). Regulation of riboflavin biosynthesis and transport genes in bacteria by transcriptional and translational attenuation. Nucleic Acids Res. 30, 3141–3151. doi: 10.1093/nar/gkf433
von Eiff, C., Bettin, D., Proctor, R. A., Rolauffs, B., Lindner, N., Winkelmann, W., et al. (1997a). Recovery of small colony variants of Staphylococcus aureus following gentamicin bead placement for osteomyelitis. Clin. Infect. Dis. 25, 1250–1251. doi: 10.1086/516962
von Eiff, C., Heilmann, C., Proctor, R., Woltz, C., Peters, G., and Götz, F. (1997b). A site-directed Staphylococcus aureus hemB mutant is a small-colony variant which persists intracellularly. J. Bacteriol. 179, 4706–4712.
von Eiff, C., McNamara, P., Becker, K., Bates, D., Lei, X.-H., Ziman, M., et al. (2006). Phenotype microarray profiling of Staphylococcus aureus menD and hemB mutants with the small-colony-variant phenotype. J. Bacteriol. 188, 687–693. doi: 10.1128/JB.188.2.687-693.2006
Zhang, P., Wright, J. A., Osman, A. A., and Nair, S. P. (2017). An aroD ochre mutation results in a Staphylococcus aureus small colony variant that can undergo phenotypic switching via two alternative mechanisms. Front. Microbiol. 8:1001. doi: 10.3389/fmicb.2017.01001
Zhang, Z., Schwartz, S., Wagner, L., and Miller, W. (2000). A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 7, 203–214. doi: 10.1089/10665270050081478
Keywords: Staphylococcus aureus, small-colony variants (SCVs), fatty acid-auxotrophy, whole-genome sequencing, energy-coupling factor transporter (EcfAA’T), phenotype switch
Citation: Schleimer N, Kaspar U, Drescher M, Seggewiß J, von Eiff C, Proctor RA, Peters G, Kriegeskorte A and Becker K (2018) The Energy-Coupling Factor Transporter Module EcfAA’T, a Novel Candidate for the Genetic Basis of Fatty Acid-Auxotrophic Small-Colony Variants of Staphylococcus aureus. Front. Microbiol. 9:1863. doi: 10.3389/fmicb.2018.01863
Received: 10 April 2018; Accepted: 25 July 2018;
Published: 14 August 2018.
Edited by:
Martin G. Klotz, Washington State University Tri-Cities, United StatesReviewed by:
Stephen Peter Kidd, The University of Adelaide, AustraliaIlana Kolodkin-Gal, Weizmann Institute of Science, Israel
Alex Van Belkum, BioMérieux, France
Copyright © 2018 Schleimer, Kaspar, Drescher, Seggewiß, von Eiff, Proctor, Peters, Kriegeskorte and Becker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karsten Becker, kbecker@uni-muenster.de
†Present address: Christof von Eiff, Pfizer Pharma GmbH, Berlin, Germany André Kriegeskorte, GSK Vaccines GmbH Marburg, Marburg, Germany