 Emanuel Bojórquez-Quintal1
Emanuel Bojórquez-Quintal1 Camilo Escalante-Magaña2
Camilo Escalante-Magaña2 Ileana Echevarría-Machado2
Ileana Echevarría-Machado2 Manuel Martínez-Estévez2*
Manuel Martínez-Estévez2*- 1CONACYT-Laboratorio de Análisis y Diagnóstico del Patrimonio, El Colegio de Michoacán, La Piedad, Mexico
- 2Unidad de Bioquímica y Biología Molecular de Plantas, Centro de Investigación Científica de Yucatán, Mérida, Mexico
Aluminum (Al) is the most abundant metal in the earth’s crust, but its availability depends on soil pH. Despite this abundance, Al is not considered an essential element and so far no experimental evidence has been put forward for a biological role. In plants and other organisms, Al can have a beneficial or toxic effect, depending on factors such as, metal concentration, the chemical form of Al, growth conditions and plant species. Here we review recent advances in the study of Al in plants at physiological, biochemical and molecular levels, focusing mainly on the beneficial effect of Al in plants (stimulation of root growth, increased nutrient uptake, the increase in enzyme activity, and others). In addition, we discuss the possible mechanisms involved in improving the growth of plants cultivated in soils with acid pH, as well as mechanisms of tolerance to the toxic effect of Al.
Introduction
Acid soils, also called ultisols or oxisols, have a pH of 5.5 or lower; they are widely distributed in tropical and subtropical regions, constituting approximately 30% of the total area of the planet and 50% of the arable land in the world, as well as providing between 25 and 80% of vegetable production (Sade et al., 2016). Soil acidification can occur due to natural and/or anthropogenic processes (Figure 1). Most acid soils occur in the tropics and subtropics, where acidification is a natural process. This situation can be worsened by environmental contamination through the use of fertilizers and acidifying substances, as well as the use of fossil energy sources such as coal and oil which release nitrogen dioxide (NO2) and sulfur dioxide (SO2) into the atmosphere and which, in the presence of oxidizing agents, give rise to nitric acid (HNO3) and sulfuric acid (H2SO4), thereby increasing the precipitation of acid rain and acidification of the bodies of water and the soil. Moreover, organic material in decomposition, imbalance in N, S, and C cycles, excessive uptake of cations over anions and the uptake of nitrogen by leguminous crops all increase the concentration of H+ and reduce soil pH (Figure 1). Acid soils are characterized by a deficiency in nutrients and toxicity by metals, such as manganese (Mn), iron (Fe) and Al; with toxicity by Al being the main factor limiting plant growth in acid soils (Kochian et al., 2004; Gupta et al., 2013; Bose et al., 2015).
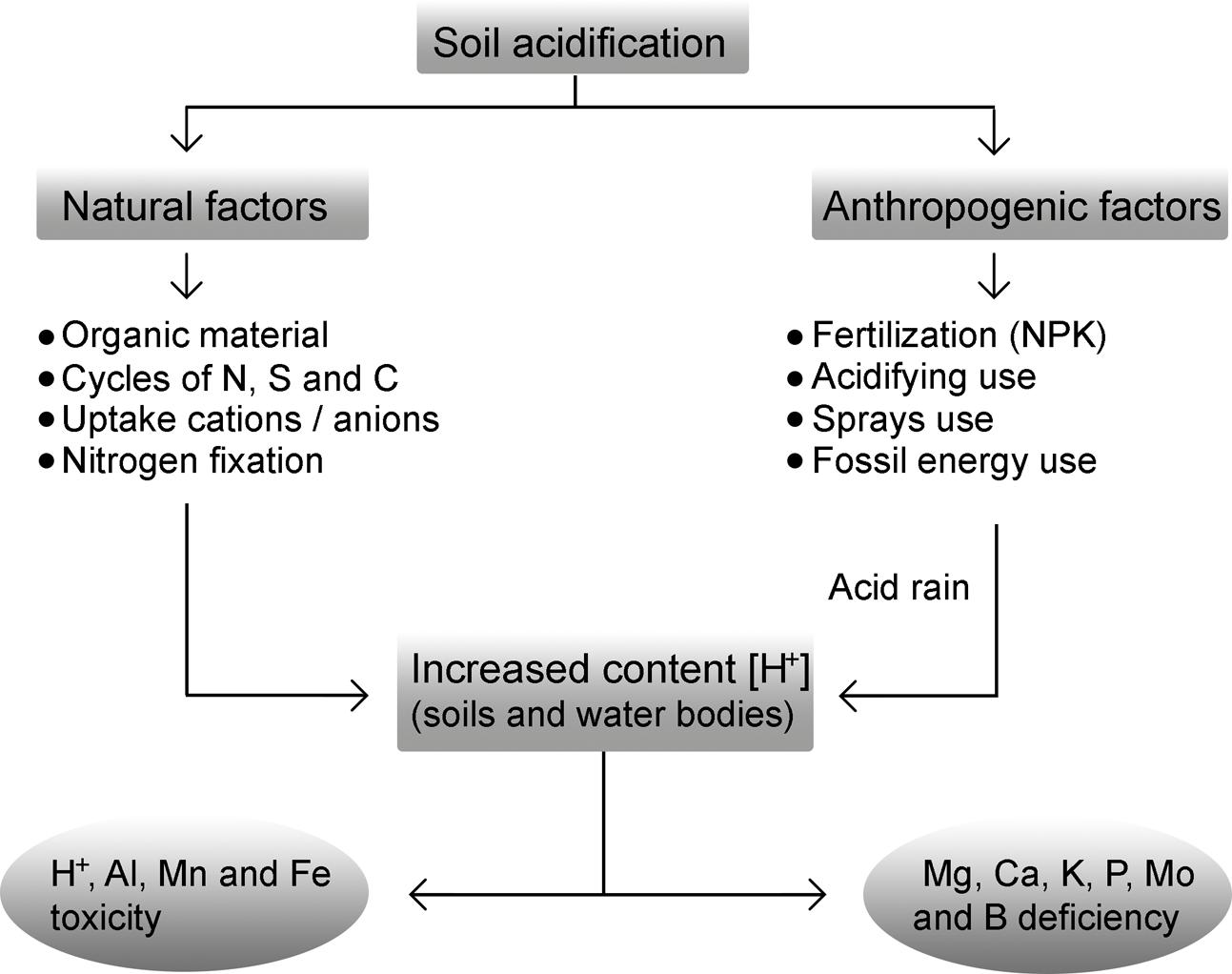
FIGURE 1. Soil acidification is a phenomenon determined by natural and anthropogenic factors. The decomposition of the organic matter, the imbalance of the N, S, and C cycles, the excess in the cation uptake on anions and the N fixation by the legumes influence the concentration of protons [H+] in the soil solution. Anthropogenic factors such as the use of fertilizers (nitrogen, phosphorus, and potassium, NPK), the use of acidifiers and aerosols (H2S, H2SO4, HF, and Cl2), and the emission of gasses (CO2, NO2, and SO2) into the atmosphere by use of fossil energy give rise to environmental pollution. Such molecules find their way into soil and water bodies in the form of acid rain, causing acidification of soils and the release of Al ions in a form easily absorbed by the plant root system, which is extremely toxic. Also, nutrient deficiency (P, Mg, and K) and toxicity by other metals (Mn and Fe) may occur.
Aluminum is the most abundant metal on earth and it is the third most abundant element (after oxygen and silicon) in the earth’s crust, representing approximately 8.1% of its content in weight (Figure 2A). Despite being ubiquitous and available during the life cycle of plants, Al has no specific biological function (Poschenrieder et al., 2008). Organisms are not usually exposed to relevant concentrations of Al in the soil as it is mainly found in the form of a mineral (aluminosilicates and aluminum oxides); however, in aqueous solutions and at different pH, Al hydrolyzes water molecules to form aluminum hydroxide (Figure 2A). Total Al concentration in the soil and the speciation of Al depend on the pH and the chemical environment of the solution (Kisnieriené and Lapeikaité, 2015). The toxic effect of the different forms of Al (speciation) on plant growth diminishes in the following order: Al, Al(OH)2+, Al(OH)2+, Al(OH)4- (Figure 2B). At a low pH (about 4.3) trivalent aluminum (Al3+) is the most abundant form and has the greatest impact on plant growth. In contrast, precipitated or chelated Al with organic compounds is not toxic for plants (Nogueirol et al., 2015).
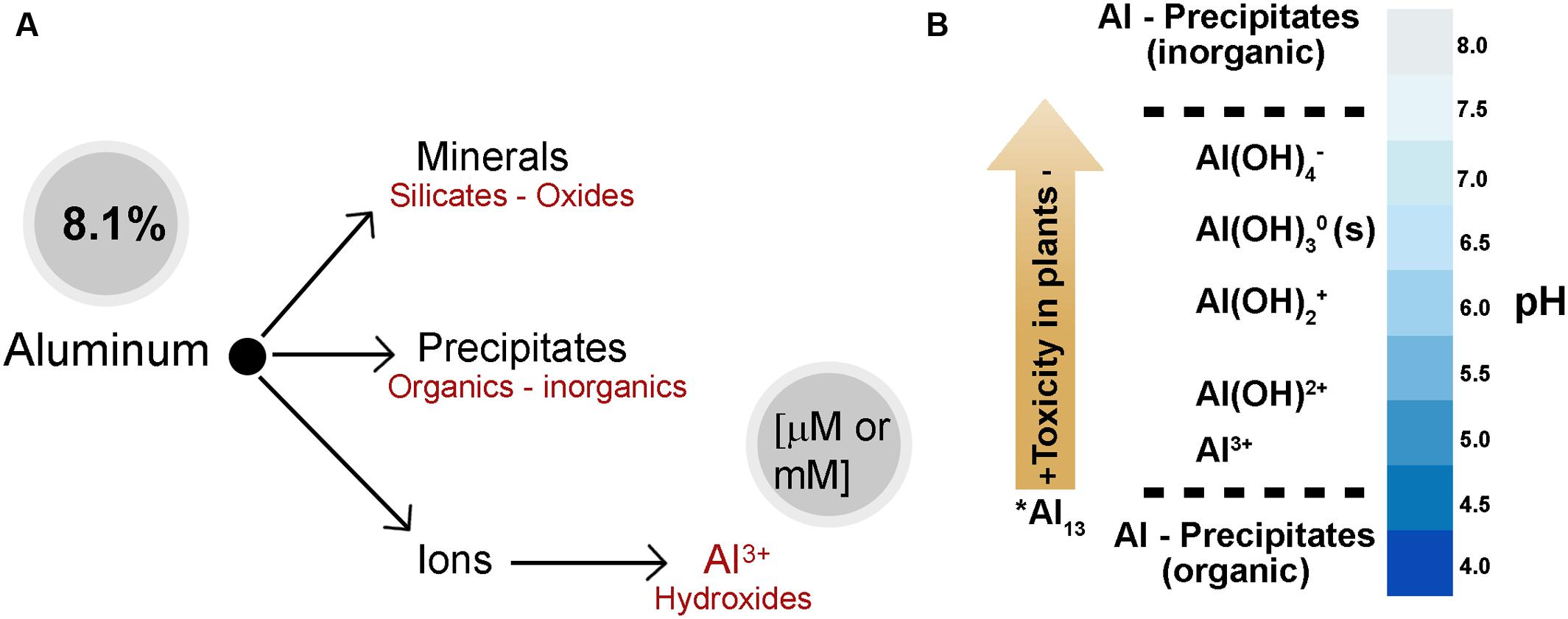
FIGURE 2. Aluminum abundance and speciation in the earth’s crust. (A) Different forms of Al in the soil and water. Aluminum is mainly found in the mineral form (aluminosilicates and aluminum oxides). In addition, Al can be found as precipitates or conjugated organic and inorganic, and molecular ions depending on the soil pH. (B) Al-speciation in soil solution. Al concentration and the speciation of Al depend on the pH and the chemical environment of the soil solution. However, a very toxic polynuclear Al species (∗Al13) depends on the total concentration of Al. Molecular aluminum (mononuclear) exists as hydroxyaluminum: Al/Al(H2O)63+, AlOH2+, Al(OH)2+, Al(OH)3 y Al(OH)4- Trivalent aluminum (Al) is the most abundant form and has the greatest impact on plant growth at pH < 5. At pH > 5–6, the dominant species are AlOH2+ and Al(OH)2+, which are not as toxic to plants as Al. When the pH is neutral, Al(OH)3 or gibbsite occurs; however, it is non-toxic and relatively insoluble. Aluminate, Al(OH)4-, is the dominant specie when the pH is alkaline (pH > 7). (Kinraide, 1991; Delhaize and Ryan, 1995; Brautigan et al., 2012; Hagvall et al., 2015; Kisnieriené and Lapeikaité, 2015).
It was recognized for the first time, over 100 years ago, that concentrations of soluble Al increase in acid soils (Veitch, 1904) and that this soluble Al is toxic for plant growth, the main effect of Al toxicity being inhibition of root growth (Daikuhara, 1914; Miyake, 1916; Magistad, 1925; Kopittke et al., 2016). Surprisingly, stimulation of root growth is one of the beneficial effects of Al. The impact of Al on plant growth, both toxic and beneficial, depends on the concentration of the metal and varies according to the plant species, which includes the genotype within the same species, physiological age, growth conditions and the duration of exposure to the metal. According to Barcelo and Poschenrieder (2002) and Zhou et al. (2011), with respect to root growth, three responses can be observed depending on the concentration of Al. (1) Root growth is not affected at low concentrations of Al; however, at a higher concentration it is diminished; (2) root growth is stimulated at low concentrations of Al, but it is affected at high concentrations; (3) inhibition of root growth at low concentrations or short time periods, but little or no effect at high concentrations or long time periods. A fourth response is presented in genotypes where root growth is not affected even at very high concentrations of Al, indicating that different plant species differ in their response mechanisms to stress by Al at cellular and tissue levels, as well as whole-plant level.
Beneficial Effect of Aluminum in Plants
Since Maze (1915) and Stoklasa (1922) reported, for the first time, on the possible role of Al in plant growth and development, considerable interest has been shown in studies on the beneficial effect of Al on plants. In recent years, an increasing number of articles dealing with this topic have been published. However, to date, no evidence has been put forward as to the essentiality of this metal. Although the exact mechanism causing the beneficial effect of Al is still unknown, a few possible mechanisms have been suggested to explain it (Figure 3).
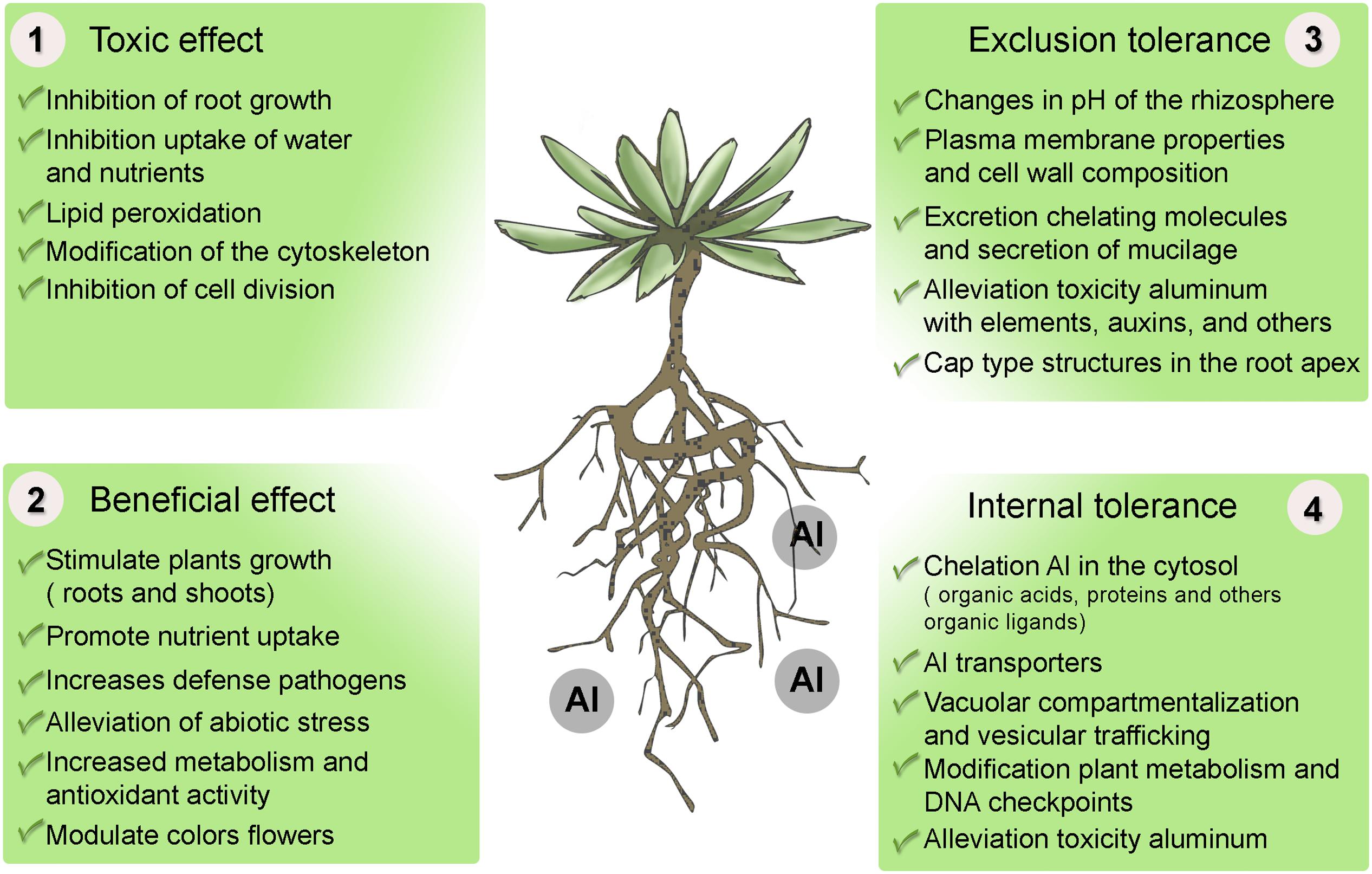
FIGURE 3. Effect of aluminum on plants and mechanisms of tolerance to stress by aluminum. (1) Toxicity of Al in plants. (2) Beneficial effect of Al in some taxas, mainly species adapted to acid soils. (3) Mechanisms of exclusion, resistance or alleviation of Al uptake, and (4) Mechanisms of internal tolerance to stress by Al in plants.
Plant Growth Stimulation by Aluminum
Growth stimulation induced by Al has been observed frequently in native plants, or plants which have adapted to acid soils when Al is mainly administered at low concentrations (Yoshii, 1937; Osaki et al., 1997; Pilon-Smits et al., 2009). In the case of the Tabebuia chrysantha tree, low levels of Al stimulated the synthesis of root biomass; in contrast, a high level of Al had an inhibitory effect (Rehmus et al., 2014). According to Poschenrieder et al. (2015) two patterns of growth stimulation resulting from the administration of Al can be observed in plants: a transitory increase (short term) in growth, observed mainly in laboratory studies, and a permanent increase in productivity induced by Al in the highly tolerant plants. For example, prevention of H+ toxicity and an increase in root elongation induced by Al is a transitory effect which occurs for short periods (Kinraide et al., 1992; Llugany et al., 1995; Kinraide, 1998). However, in an in vitro culture of coffee (Coffea arabica) seedlings, growth stimulation of the primary root induced by Al occurred from day 10 to day 30 of culture (Bojórquez-Quintal et al., 2014). In Betula pendula and Quercus serrata trees, growth increase (roots and leaves) induced by Al has been observed in the long term, even after 28 days and 17 months, respectively (Kidd and Proctor, 2000; Tomioka and Takenaka, 2007). Similar results were reported in Conostegia xalapensis after 24 days of treatment with 0.5 and 1 mM of Al. The treatment with Al increased root biomass and the number of lateral roots. C. xalapensis is a shrub, hyperaccumulator of Al, which is common to Mexico and Central America, and colonizes pollution-perturbed areas. A possible role as an indicator species for toxicity and contamination of Al has been suggested for this species (González-Santana et al., 2012).
Camellia sinensis, Miscanthus sinensis, Quercus serrata, and Melastoma malabathricum are tropical species, hyperaccumulators of Al, which grow in acid soils (Yoshii, 1937; Osaki et al., 1997; Ghanati et al., 2005; Tomioka et al., 2005). In tea (C. sinensis) and Q. serrata, it is well known that Al potentializes biomass growth, root elongation and both short and long term proliferation of lateral roots (Ghanati et al., 2005; Tomioka et al., 2005; Mukhopadyay et al., 2012; Hajiboland et al., 2013b; Xu et al., 2016). In the case of tea, it has been demonstrated that root growth is stimulated in the presence of Al, while in the absence of the metal, growth of the root and the plant is delayed (Tsuji et al., 1994; Fung et al., 2008). In M. malabathricum it has been suggested that Al is essential for the growth of this plant; in the absence of the metal, chlorosis, morphological changes and leaf curling have been observed (Watanabe et al., 2006). The Melastoma plant secretes mucilage in the roots in order to accumulate Al in soils with poor availability of this element and the accumulation of Al increases the growth of roots and shoots (Watanabe et al., 2005, 2008a,b). Morphological changes in the root, such as thickening, white coloration and elongation have also been reported in C. sinensis, Q. acutissima, Cinnamomum camphora, Eucalyptus viminalis, Q. serrata, M. malabathricum, and Symplocos paniculata in the presence of Al (Oda, 2003; Ghanati et al., 2005; Watanabe et al., 2006; Schmitt et al., 2016a).
Despite the fact that the beneficial effect of Al in plants has been reported mainly in woody species adapted to acid soils (Osaki et al., 1997; Hajiboland et al., 2013b), there are reports available on species of economic importance, such as rice (Oryza sativa) and corn (Zea mays) (Watanabe et al., 1997; Famoso et al., 2011; Wang et al., 2015b). In rice varieties tolerant to Al, an increase in root growth has been observed in the presence of 160 and 200 μM of Al (Famoso et al., 2011; Moreno-Alvarado et al., 2017). The administration of a low dosage of Al in corn inhibited root growth, but increased leaf growth (Wang et al., 2015b). Similarly, an increase in the Al content of horticultural species, such as turnip (Brassica rapa) and the legume Glycine max increases elongation and activity of the root (Rufty et al., 1995; Yu et al., 2011).
Aluminum can also stimulate the growth of other organisms. In marine environments, Al increases the growth and biomass accumulation of phytoplankton, mainly diatoms (Saçan et al., 2007; Golding et al., 2015; Zhou L. et al., 2016). In a mutant of Saccharomyces cerevisiae (zrt1Δ), deficient in zinc uptake, the addition of Al restored growth to a level comparable to that of the wild strain and promoted the uptake of Zn (Tamura and Yoshimura, 2008). In microorganisms such as Frankia (a nitrogen-fixing bacteria), an increase in growth (in vitro) was observed with the addition of 500 μM of Al; similarly, it was possible to observe that Al prevented the inhibitory effect of the acidic pH (Igual and Dawson, 1999). Aluminum is also involved in bone formation in animal cells (Quarles et al., 1988, 1989). It has been observed that the addition of Al increases the proliferation and differentiation of human and chicken osteoblasts (Lau et al., 1991). Moreover, it has been demonstrated that Al induces the synthesis of DNA in osteoblasts (Quarles et al., 1994) and acts as mitogen in epithelial cells of mice (Jones et al., 1986).
Aluminum Promotes Nutrient Uptake
One of the possible reasons explaining the stimulation of plant growth induced by Al is the promotion of nutrient uptake. In hyperaccumulator plants, Al can stimulate or have no effect on essential nutrient uptake (Malta et al., 2016). In different plant species, nitrogen (N), phosphorous (P), and potassium (K) uptake has been considered the mechanism responsible for the stimulation of root growth induced by Al (Osaki et al., 1997). In tea plants, the stimulation of root growth has been explained as a consequence of an increased uptake of some macronutrients (Fung et al., 2008). In M. malabathricum, Q. serrata and tea the uptake and accumulation of P in the roots and leaves of the plants increased in the presence of Al. It has been suggested that the stimulation of root growth and the increase in P could be due to precipitation of the Al-P complex on the root surface and/or in the Donnan free space (apoplast) and in some way the plants use the precipitated P (Konishi, 1992; Osaki et al., 1997; Tomioka et al., 2005). Similarly, Al stimulates alkaline phosphatase activity and organic P uptake in the marine diatom Thalassiosira weissflogii (Zhou L. et al., 2016).
In Q. serrata, stimulation of root growth was associated with the activation of the nitrate reductase and the increase in NO3- uptake (Tomioka et al., 2007, 2012). In species of the genus Symplocos there is a positive correlation between the calcium (Ca) uptake and the level of Al (Schmitt et al., 2016a). Similarly, in C. arabica roots an increase in the content of K and Ca was observed with the concentration of Al which stimulated primary root growth (Bojórquez-Quintal et al., 2014). All the data presented suggest that Al can induce the expression or activity of transport proteins (channels and transporters) and change the membrane potential and proton flux (H+) which promotes the fluxes of nutrients in the plants. There is evidence in wheat roots (Triticum aestivum) that Al increased the extrusion of H+ and the polarity of the membrane, which could be associated with a greater nutrient uptake and growth increase (Kinraide et al., 1992; Kinraide, 1993). Moreover, in Arabidopsis thaliana, Al induces depolarization of the membrane, K and H+ influx, and reduces K efflux (Bose et al., 2010a,b). Magnesium (Mg) can prevent the toxic effect of Al on plants (Bose et al., 2011). In a mutant of Arabidopsis (alr-140), an increase in influx and intracellular content of Mg induced by Al has been observed; it has also been suggested that Al can activate channels and Mg transporters in Al-resistant plants (Bose et al., 2013).
Aluminum Prevents Biotic and Abiotic Stress
The beneficial elements, including Al, can increase tolerance to abiotic stress (ion toxicity and nutrient deficiency) and resistance to biotic stress (herbivores and pathogens) (Kaur et al., 2016). Pilon-Smits et al. (2009) mention that plants which are hyperaccumulators of Al (1 g Al kg-1 dry weight) can use this metal in their tissues to discourage herbivores, as was observed in the application of Al to prevent herbivory in Festuca arundinacea (Potter et al., 1996). Due to the fact that Al can be toxic for some pathogenic microorganisms, a number of salts containing Al have been used to control diseases caused by fungi in crops of carrot (Daucus carota) and potato (Solanum tuberosum) (Kolaei et al., 2013). In S. tuberosum, treatment with Al increased resistance to the oomycete Phytophthora infestans and to the pathogenic fungus Thielaviopsis basicola Ferraris. The addition of Al inhibited the germination of spores and fungus growth (Meyer et al., 1994; Andrivon, 1995). The protective capacity of Al against P. infestans was associated with the accumulation of H2O2 in the roots and the activation of the acquired systemic response depending on salicylic acid and nitric oxide (Arasimowicz-Jelonek et al., 2014).
Aluminum can prevent the effects of H+ toxicity and that of different elements when found in excess. The increase in root biomass of tree species after treatment with Al has been associated with the lessening of H+ toxicity (Thornton et al., 1986, 1989). This mechanism has also been suggested in wheat, Japanese radish (Raphanus sativus var. Longipinnatus) and pea plants (Pisum sativum), but not in Q. serrata (Kinraide et al., 1992; Kinraide, 1993; Tomioka et al., 2005). In acid soils, the availability of Al, iron (Fe) and manganese (Mn) is high; thus, the plants growing in these soils can present toxicity of these metals (Figure 1). An excess of Fe induces the production of reactive oxygen species (ROS), leading to the interruption of several cellular functions. In plants of M. malabathricum and C. sinensis, growth stimulation is accompanied by the prevention of iron toxicity. In these species, Al prevents bronzing of the leaves due to the toxic effect of Fe and also reduces the Fe content in leaves and roots (Watanabe et al., 2006; Watanabe and Osaki, 2009; Hajiboland et al., 2013a).
In some plant species, Al can prevent the Mn toxicity (Yang et al., 2009; Wang et al., 2015e). In rice, the prevention of Mn toxicity induced by Al can be attributed to the reduced metal accumulation in the shoots as a result of the decrease in Mn uptake in the roots. The reduction of Mn uptake in the roots was a consequence of changes in the membrane potential. In addition, Al brought about an increase in insoluble Mn in the root and changed the properties of junction to the cell wall, making Mn less available in the rice roots (Wang et al., 2015e). Aluminum can also help to detoxify fluoride (F) in tea plants by forming Al-F compounds. In the absence of the metal, the tea plants were sensitive to F (Yang Y. et al., 2016). Aluminum is also able to prevent toxicity caused by other elements, particularly P, zinc (Zn) and copper (Cu) (Asher, 1991). Also, Al improves plant growth under nutrient deficiency; for example, supplementation with Al enhanced the growth of tea plants deficient in boron (B). Under B deficiency conditions, Al was able to positively regulate the metabolism of N and carbon (C) and antioxidant defense activity, while increasing the uptake and transportation of B in tea plants (Hajiboland et al., 2014, 2015). Under limiting conditions of P, Al stimulates P uptake in microalgae (Zhou L. et al., 2016).
Beneficial Effects of Aluminum on Plant Metabolism
Investigation on the beneficial effect of Al has focused mainly on physiological aspects such as the promotion of nutrient uptake; however, recent advances have suggested the possible role of Al in plant metabolism (Xu et al., 2016). In tea plants, Al is retained in the apoplast of leaf epidermal cells as a mechanism of Al-resistance (Tolrà et al., 2011). In the aerial part of the plant, the stimulatory effect of Al has been attributed to the increase in photosynthesis and the activation of antioxidant defense (Hajiboland et al., 2013b). Similarly, in roots and cell suspensions of tea plants, Al induces the activation of antioxidant enzymes, increases the integrity of the membrane and reduces lignification and aging, which could be a possible reason for the stimulation of root growth (Ghanati et al., 2005). In C. xalapensis plants treated with Al, the increase in biomass and in the number of lateral roots coincided with greater activity of the glutathione reductase and superoxide dismutase, at low levels of ROS (González-Santana et al., 2012). In Q. serrata, stimulation of root growth by Al is associated with an increase in the activation of nitrate reductase and the photosynthetic rate (Tomioka and Takenaka, 2007; Tomioka et al., 2007, 2012; Moriyama et al., 2016).
The beneficial effect of Al on plants has also been associated with the regulation of C and N metabolism. The shrub M. malabathricum changes the metabolism of organic acids (OA) in the absence or presence of Al; more specifically, it increases the synthesis of citrate and diminishes that of malate (Watanabe and Osaki, 2001), suggesting the possible role of Al in the regulation of the expression and activation of genes and proteins associated with the biosynthesis of OA with respect to growth stimulation. In M. malabathricum, citrate is necessary for the transportation of Al from the roots to aerial part of the plant (Watanabe et al., 2005). In tea plant roots, it has been suggested that the secretion of caffeine induced by Al can increase root growth by inhibiting callus deposits (Morita et al., 2011). Also, Al positively regulates the biosynthetic pathway of caffeine in suspension cells of C. arabica. This regulation may be accomplished by activation of a signal transduction pathway through Ca and ROS (Pech-Ku et al., 2017). There is also evidence indicating that Al increases the content of chlorophyll, carotenoids, sugars, amin oacids such as proline and cysteine, hormones, and metabolites of the Shikimico acid pathway in woody and crop plants (Hajiboland et al., 2013b; Moriyama et al., 2016; Xu et al., 2016; Moreno-Alvarado et al., 2017). In fact, glucose has been suggested as an energy source which is a key element in the promotion of root growth, in the presence of Al. Moreover, glucose and abscisic acid (ABA) may participate as signaling molecules which promote the root growth induced by Al, acting on the metabolism of N and C (Moriyama et al., 2016). For instance, Al increases the concentration of soluble sugars and differentially regulates the expression of NAC transcription factors, which in turn may enhance growth and biomass production in rice plants (Moreno-Alvarado et al., 2017).
In corn, stimulation of leaf growth has been associated with greater protein synthesis (Wang et al., 2015b). In coffee seedlings, the activity of phospholipase C, an enzyme which participates in signal transduction, was increased in the concentration of Al where it was possible to observe stimulation and inhibition of root growth, suggesting a possible participation in the signal-transduction pathway mediated by inositol phosphate in the beneficial and toxic effect of Al in plants (Bojórquez-Quintal et al., 2014). It has also been suggested that the beneficial effect of Al on root growth and morphological changes is associated with changes in the levels of plant growth regulators (PGR). Aluminum is capable of inducing, directly or indirectly, synthesis or transportation of PGR. In the conifer Picea abies, an increase in the levels of indole-acetic acid (IAA), cytokinins and gibberellins has been reported in the roots in the presence of Al; these changes are in correlation with the morphological changes and the stimulation of lateral roots close to the root apex. The formation of new roots in Q. acutissima, C. camphora, and Q. serrata was accelerated by the Al treatment and correlated with low concentrations of IAA and an increase in the concentration of cytokinins and ABA in the roots (Oda, 2003; Moriyama et al., 2016).
Aluminum Modulates Floral Colors
Aluminum plays an important role in the color changes of some hyperaccumulator plants. In hydrangea (Hydrangea macrophylla), Al and pH are important factors for the change of color in the sepals, from red to blue (Ito et al., 2009; Schreiber et al., 2011). The blue coloring is formed from the union of Al with metalloanthocyanins, delphinidin 3-glucoside and 5-O-caffeoylquinic acid (Oyama et al., 2015; Kodama et al., 2016). In the genus Camellia, the purple color of Camellia japonica flowers (cv. Sennen-fujimurasaki) and the intense yellow tone of Camellia chrysanthaes are generated by the chelation of the Al ions with anthocyanins and flavonoids (quercetin), respectively (Tanikawa et al., 2008, 2016). Anthropologically, the leaves and bark of species of the genus Simplocos are used by weavers as mordant in textile dyeing due to their high content of Al (Schmitt et al., 2016b).
Aluminum Toxicity
Aluminum is also the primary factor in reducing crop yields in acid soils (Ma et al., 2002; Kochian et al., 2005). The initial and most dramatic Al toxicity symptom is the inhibition of root elongation (Delhaize and Ryan, 1995). Because Al is a highly reactive element, there are innumerable mechanisms of toxicity involving the cell wall (Jones et al., 2006; Zhang et al., 2007) and plasma membrane, where it can modify its structure, as well as the nearby ionic medium to wall, both disturb the transport of ions and cause an improper balance of nutrients (Bose et al., 2011). Also, Al can affect the constituents’ symplast (calmodulin) (Tokizawa et al., 2015), apoplast (pectin matrix) (Eticha et al., 2005; Delhaize et al., 2007) and DNA in cells of plant roots (Kochian et al., 2004; Sade et al., 2016).
Aluminum is widely reported as toxic to most plants (Sade et al., 2016). There are a number of symptoms caused by aluminum toxicity in plants (Figure 3), all associated with severe changes in the root system (Kopittke and Blamey, 2016). Al interferes with cell division at the root apex and lateral roots, increases the rigidity of the cell wall by cross-linking of pectins and reduces DNA replication because of increased rigidity of the double helix (Zhang et al., 2014; Eekhout et al., 2017). Moreover, Al induces a series of cellular toxic changes concerning cell division and nucleolus, and localization and expression of the nucleolar proteins such as fibrillarin (Zhang et al., 2014). Aluminum tends to bond with phosphorus (P) in a less available and insoluble form in soils and plant roots, thereby creating a P deficiency for plant growth. Aluminum also decreases root respiration, interferes with enzymes governing the deposition of polysaccharides in cell walls, decreases the synthesis and transport of cytokinins, and modifies the structure and function of plasma membranes which interfere with the uptake, transport, and use of multiple elements (Ca, Mg, P, and K), as well as water uptake by roots plants (Foy, 1974; Foy et al., 1978; Kochian et al., 2004).
In plants these symptoms are linked to disorders that are generally divided into two categories: (1) long-term responses, requiring hours to develop, and (2) short-term responses that can be measured within a few minutes or even a few seconds after exposure to Al (Taylor, 1988; Simoes et al., 2012). The first signs of these responses related to Al toxicity have been observed after one hour (Ownby and Popham, 1989). Moreover, the most important response to the application of Al seems to be short term interruption of Ca influx through the plasma membrane (Jones et al., 1998; Panda and Matsumoto, 2007).
Aluminum and other metals are non-biodegradable, they remain in the environment and are able to circulate in the food chain, posing a serious threat not only to plants but also to animals and humans (Jackson and Huang, 1983; Exley, 2003; Ashenef, 2014). For example, tea plants contain a substantial amount of Al in leaves, and thus it is present in tea leaf infusions. Although only a small proportion of Al is available for absorption in the gastrointestinal tract and the renal excretion of Al is fairly effective, this metal can cause serious problems or possible health risks in humans (Exley, 2003; Mehra and Baker, 2007). High Al content in the human body has been hypothesized as having possible links with various diseases, such as encephalopathy, dementia, Alzheimer’s disease, osteomalacia fractures and high levels of bone Al; therefore tea-drinkers should be forewarned of the risks (Anitha and Rao, 2002; Mehra and Baker, 2007; Ashenef, 2014).
Mechanisms of Tolerance to Aluminum
Plants have developed different mechanisms of tolerance to counteract the toxic effects of Al. These mechanisms can be divided into two forms (Figure 3): mechanisms of exclusion or resistance to Al, the function being to avoid or reduce the entrance of Al to the cell; and mechanisms of internal tolerance which compartmentalize Al in vacuoles or stabilize them in order to inhibit its toxicity.
Tolerance to Aluminum – Mechanisms of Exclusion
Changes in the Rhizosphere pH
A small increment in the rhizospheres pH reduces the solubility, activity, toxicity and content of Al in plants through exclusion of the metal in the root apoplast (Yang Y. et al., 2011). The Arabidopsis alr-104 mutant Al-resistant and other plant species increase the apoplastic pH through the H+ and NH4+ influx and the efflux of OA in the root apex, in the presence of Al (Foy et al., 1965; Degenhardt et al., 1998; Larsen et al., 1998; Houman et al., 2009; Bose et al., 2010b; Wang et al., 2015a). In squash (Cucurbita pepo) and wheat, changes in rhizosphere pH and resistance to stress due to Al are regulated by H+-ATPasa of the plasma membrane (Ahn et al., 2001, 2002; Yang Y. et al., 2011). OA can also increase or reduce rhizosphere pH in the presence of other metals, as well as Al (Kochian et al., 2004; Zeng et al., 2008; Javed et al., 2013). In the case of tea, the Al uptake induces H+ efflux from the roots, suggesting a mechanism of soil acidification (Wan et al., 2012). Aluminum plays a positive role in growth increase in tea (Konishi et al., 1985), particularly in the promotion of root elongation. Soil acidification might maintain the solubility and Al uptake in tea. In contrast, acidification of the rhizosphere increases toxicity and metal accumulation in Al-sensitive species (Houman et al., 2009).
Changes in Cell Wall Composition and Plasma Membrane Properties
The root cell wall is the main binding site of Al and thus is the target of Al toxicity and exclusion in plants (Horst et al., 2010; Yang J.L. et al., 2011; Kopittke et al., 2015; Liu S. et al., 2016). At present, physiological, biochemical and molecular evidence is available which has demonstrated that modification of the cell wall composition plays an important role in the resistance to Al (Levesque-Tremblay et al., 2015; Che et al., 2016; Zhang et al., 2016). The increase in the polysaccharide content of the cell wall induced by Al can reduce water and nutrient uptake, as well as cellular wall elasticity (Nguyen et al., 2005). Among the cereals, rice is the crop with most tolerance to Al (Yang et al., 2008; Famoso et al., 2010). In rice cultivars which differ in their Al-resistance, a positive correlation between polysaccharide content (pectin and hemicellulose) in the root apex and the accumulation of Al has been observed, indicating the importance of the cell wall composition in the exclusion of Al (Yang et al., 2008). The pectin content and its degree of methylation in the cell wall contribute to the differences in resistance to Al (Eticha et al., 2005; Yang J.L. et al., 2011). In Arabidopsis and other plant species, the regulation of genes and expansin enzymes, pectin methyltransferases and xiloglucano endohydrolases reduce binding and accumulation of Al in the root apex by changing the load and the porosity of the cell wall (Yang J.L. et al., 2011; Zhu et al., 2014; Zhang et al., 2016). The transporter ABC formed by the complex STAR1/STAR2 transport UDP-glucose and it can modify the cell wall under stress by Al (Huang et al., 2009).
The physicochemical and physiological properties of the plasma membrane also affect tolerance of the plants to Al. The lipid composition of the plasma membrane (PM) has an important role in Al-tolerance (Wagatsuma et al., 2015). The phospholipids create a negative charge on the surface of the PM and increase the sensitivity to Al, as a result of the union of the metal to the PM. In rice and timber species (M. malabathricum and M. cajuputi), Al-tolerance increased in response to a decrease in the proportion of phospholipids of the root cell PM (Maejima and Watanabe, 2014). Similarly, the sterol content also plays an important role in the tolerance to stress by Al. High sterol content in combination with low contents of phospholipids is a common strategy for Al-tolerance in different plant species (Khan et al., 2009; Wagatsuma et al., 2015). It has also been suggested that a small peptide, anchoring the PM, could prevent Al-influx in root cells through bonding with cation and thus contribute to the resistance in rice (Xia et al., 2013).
Excretion of Chelating Molecules and Mucilage Secretion
The release of OA in the root is the Al exclusion mechanism most widely described in plants (Kochian et al., 2015), with a wide natural variation in different crops, mainly in cereals (Brunner and Sperisen, 2013; Schroeder et al., 2013). The organic anions, malate, citrate and oxalate are secreted by the root and chelate Al in a non-toxic Al-OA complex, protecting the root apex and permitting it to grow. Malate and citrate are ubiquitous in all plant cells given that they are part of the tricarboxylic acid (TCA) cycle in the mitochondria, while oxalate participates in the regulation of Ca and the detoxification of metals (Brunner and Sperisen, 2013). Evidence in different species of cultivable plants and timber trees indicates that OA efflux confers resistance to Al (Delhaize et al., 1993; Brunner and Sperisen, 2013; Yang et al., 2013).
In a wheat genotype resistant to Al (Delhaize et al., 1993), the first gene of a malate transporter activated by Al (TaALMT1) was cloned, which codifies an anionic channel activated by Al that facilitates the malate efflux from the root. TaALMT1 was the first gene of resistance to Al cloned in plants (Sasaki et al., 2004). To date, several ALMT1 type genes have been isolated in different plant species (AtALMT1, Hoekenga et al., 2006; BnALMT1 and BnALMT2, Ligaba et al., 2006; HvALMT1, Gruber et al., 2010; ZmALMT2, Ligaba et al., 2012). The heterologous expression of TaALMT1 in barley (Hordeum vulgare) and tobacco (Nicotiana tabacum) allowed the malate efflux and an increase in Al-resistance (Delhaize et al., 2004; Zhang et al., 2008). On the other hand, the citrate efflux is mediated by the family of MATE proteins (Multidrug and toxic compound extrusion). MATE genes have been cloned and identified in Arabidopsis (AtMATE, Liu et al., 2009), sorghum (SbMATE, Magalhaes et al., 2007), barley (HvAACT1-HvMATE1, Furukawa et al., 2007), wheat (TaMATE1, García-Oliveira et al., 2014), poplar (PtMATE1, Grisel et al., 2010), rice (OsFRD1; Yokosho et al., 2011), rice bean (VuMATE1, Yang X.Y. et al., 2011), corn (ZmMATE1-ZmMATE2, Maron et al., 2010, 2013) and soybean (GmMATE1-GmMATE117, Liu J. et al., 2016). The plasma membrane H+-ATPase plays an important role in OA exudation in the root by the transporters ALMT and MATE (Yu W. et al., 2016); however, in cluster roots of white lupin (Lupinus albus), the citrate exudation induced by combined treatment with P-deficiency and Al is independent of H+-ATPase PM activity and dependent on K efflux (Zeng H. et al., 2013). To date, the molecular components of oxalate efflux have not been identified. In tea plants and other woody species such as poplar (Populus tremula) and buckwheat (Fagopyrum esculentum), oxalate is an important element in the detoxification of Al in the root (Qin et al., 2007; Morita et al., 2008; Wang et al., 2015a).
In addition to the OA, the exudation of other organic compounds in the root has been suggested for the chelation of Al (Kochian et al., 2015); however, very little is known of their mechanism of action. Studies on Eucalyptus camaldulensis have revealed secretion in the root of a ligand with low molecular weight binding to Al (Tahara et al., 2008, 2009). In tea plants, an increase in the release of caffeine, a phenolic compound, has been observed in response to exposition to Al (Morita et al., 2011). Other ligands released in the root include the phenolic compounds (catechol, catechin, and quercetin), flavonoids, succinate, phosphates, UDP-glucose and polysaccharides in the form of mucilage (Kidd et al., 2001; Winkel-Shirley, 2002; Zheng et al., 2005; Huang et al., 2009; Osawa et al., 2011; Kochian et al., 2015). In fact, root mucilage plays an important role as a mechanism of resistance to metals (Morel et al., 1986). Mucilage is a gelatinous material consisting of polysaccharides of high molecular weight which are exuded from the most external layers of the root apex. Due to the fact that the mucilage contains uronic acid and pectin, the carboxil groups of this acid and of the pectin can ligate metallic cations such as Al (Watanabe et al., 2008a,b). In cowpea (Vigna unguiculata), wheat and corn a strong binding of mucilage-Al has been reported and this bond is not toxic for the plant (Horst et al., 1982; Archambault et al., 1996; Li et al., 2000).
CAP Type Structures and Border Cells in the Radical Apex
The function of the root cap or calyptra and the border cells is to protect the mother cells of the root apex from microbes and soil stresses, and also to receive and transmit environmental signals which will ultimately determine root growth (Endo et al., 2011; Kumpf and Nowack, 2015; Hawes et al., 2016). In many plant species, the root cap and border cells produce and exude mucilage, rich in polysaccharides, which can bind to metallic cations (Horst et al., 1982; Cai et al., 2013; Kumpf and Nowack, 2015). In genotypes of bean (Phaseolus vulgaris), barley, soybean and castor (Ricinus communis), resistant to Al, the exclusion of the metal is associated with the immobilization and detoxification of Al with the mucilage secreted by the root cap and border cells (Miyasaka and Hawes, 2001; Zhu et al., 2003; Cai et al., 2013; Silva et al., 2014). In contrast, the mucilage secreted by the root cap of M. malabathricum increases the accumulation of Al in the plant (Watanabe et al., 2008a,b). In pea (Pisum sativum) plants, it has been observed that removal of the border cells increases the sensitivity and absorption of Al, indicating the important role of the border cells as mechanisms of Al exclusion (Yu et al., 2009a; Yang J. et al., 2016). Furthermore, in Acacia mangium, a legume tropical forest tree known to be resistant to Al, the root apex was found to be surrounded by a cap type structure, the purpose of which is to protect the root from the flexion induced by Al (Endo et al., 2009, 2011).
Alleviation of Aluminum-Induced Toxicity
Recent evidence has demonstrated that the exogenous addition and availability of certain elements prevents Al toxicity in plants. In rice, ammonium (NH4+) reduces the accumulation of Al in the root as a consequence of pH changes induced by NH4+ uptake and the direct competition of Al and NH4+ for the cell wall binding sites (Wang et al., 2015d). Similarly, the exogenous addition of Si in corn prevents the inhibition of root elongation and callous deposition through the formation of hydroxy aluminosilicates in the root apex (Wang et al., 2004). In pea and rice, the application of Si reduced the content of Al in the roots, stem and leaves (Singh et al., 2011; Shen et al., 2014). In addition to being cofactor of many enzymes and a central component of chlorophyll, Mg also prevents metal phytotoxicity, including Al. High concentrations of Mg (mM) prevent Al toxicity by competing in uptake and interaction with the binding sites in the cell wall and the plasma membrane (Bose et al., 2011; Rengel et al., 2015). In the cytosol, Mg (μM) can increase the biosynthesis of OA and induce the activity of the H+-ATPasa by phosphorylation to increase the resistance to Al (Bose et al., 2013; Rengel et al., 2015). Prevention of Al toxicity by supplementation with B has been reported in a large number of plants. Boron can act synergically with Ca and prevent binding of Al to the cell wall (Hossain et al., 2005; Yu et al., 2009b; Horst et al., 2010). Recent evidence in pomelo (Citrus grandis) suggests that B prevents Al-toxicity by regulating different genes associated with modification of the cell wall, cellular transport, metabolism, signal transduction and antioxidant activity (Wang et al., 2015c; Zhou et al., 2015). Similarly, the addition of P prevents the effect of Al-toxicity on root growth and photosynthetic machinery of C. grandis (Jianq et al., 2009). The P-deficiency also reduces Al-toxicity by changing the properties of the plasma membrane and cell wall, while high P increases the toxicity of the metal, possibly through the precipitation of Al-P on the root surface (Maejima et al., 2014; Shao et al., 2015).
Hormones and polyamines play an important role in the tolerance to stress by Al. The exogenous addition of auxins (indole-acetic acid, IAA) reduces the accumulation of Al in the root apex in wheat (Wang et al., 2013c) and alleviates the Al-induced inhibition of root growth in lucerne (Medicago sativa) (Wang S. et al., 2016). Indole-acetic acid stimulates the exudation of citrate by the positive regulation of the GmMATE transporter and increases the activity of the PM H+-ATPasa by phosphorylation. Both processes participate in resistance to stress by Al through chelation of Al-citrate and pH changes in the rhizosphere (Wang S. et al., 2016; Wang P. et al., 2016). Furthermore, the overexpression of a transport auxin efflux (OsPin2) prevents rigidity of the cell wall, Al-binding to the cell wall and oxidative damage induced by Al through the transport of auxins and H+ (Wu et al., 2014). The polyamine putrescine (Put) and nitric oxide are also involved in the modulation of citrate secretion from roots of red bean (Wang et al., 2013a). The Al-induced root inhibition could be alleviated by Put through decreased ethylene production (Yu Y. et al., 2016). Putrescine has been identified as an important signaling molecule involved in Al tolerance in plants (Chen et al., 2008; Wang et al., 2013b; Yu Y. et al., 2016).
Some factors of abiotic stress such as drought and hypoxia can indirectly reduce the accumulation of Al and its effect on plants. In bean, drought stress changes the porosity of the cell wall and reduces the Al-binding (Zhang et al., 2016); on the other hand, hypoxic stress in barley prevents Al-toxicity in roots by increasing the antioxidant capacity and reducing K efflux induced by ROS (Ma et al., 2016). Mycorrhiza association can also prevent Al-toxicity in acid soils (Seguel et al., 2013).
Tolerance to Aluminum- Mechanisms of Internal Tolerance
Chelation of Aluminum in the Cytosol (Organic Acids and Other Organic Ligands)
The mechanism of internal detoxification of Al involves the chelation of metal with OA and subsequent sequestration into the vacuole. Many tolerant plants, including the hyperaccumulators of Al, use the OA for the sequestration of Al in the cytosol of the root cells and also to remobilize or to translocate Al toward the shoots. Fagopyrum esculentum uses oxalate for the internal and external chelation of Al (Ma et al., 1998; Ma and Hiradate, 2000; Shen et al., 2002; Wang et al., 2015a). Oxalate is the predominant ligand in the root cytosol of tea and forms Al-oxalate compounds (Morita et al., 2004); however, Al is translocated toward the shoots in the form of Al-citrate or Al-malate (Morita et al., 2004, 2008). A mechanism similar to that of tea is observed in buckwheat (Wang et al., 2015a). In the shrub M. malabathricum, intracellular Al binds to citrate, and the Al-citrate compound is transported to the shoots; once in the leaves, the citrate is substituted by oxalate to form Al-oxalate (1:3) which is less toxic (Watanabe and Osaki, 2001). In Camellia oleifera Al is transported via phloem; however, it is not known if it is in the form of an Al-OA compound (Zeng Q.L. et al., 2013).
In addition to the OAs, the phenolic compounds can bind to Al and form a complex in the cytosol. For example, in tea plants, catechin forms a complex with Al (Nagata et al., 1992). In the sepals of H. macrophylla Al can bind to chlorogenic acid (3-caffeoylquinic acid) and delphinidin a 3-glucoside for stabilization of Al and color change in the flowers (Ma et al., 1997, 2001). In C. camphora, the accumulation of proanthocyanidin in the epidermal cells of the root apex is induced by Al; however, there is no evidence of its binding to metal (Osawa et al., 2011). It has also been suggested that the hydroxamates can bind to Al in the root (Poschenrieder et al., 2005). In the sap of tea leaves, the Al-F compound has been identified as a mechanism of tolerance to F (Yang Y. et al., 2016).
Aluminum Transporters in the Plasma Membrane and Vacuolar Compartmentalization
Transportation through biological membranes requires transport proteins. In plants, Al transportation through the plasma membrane and the vacuole tonoplast has not been widely studied. However, it has been reported that the ABC transporters, transporters of binding to ATP, AtABCI16/AtALS3, AtABCI17/AtSATR1/AtALS1 and OsALS1, as well as the transporters Nrat1 (Nramp Family) contribute to the detoxification of Al in plants (Xia et al., 2010; Hwang et al., 2016). AtALS3 is a partial, type ABC transporter which is located in the PM of the epidermal cells of the root cortex, and in phloem cells throughout the plant. It is believed that AtASL3 distributes Al inside the plant far from the root apex (sensitive to Al), by transporting Al directly or bound to a ligand (Kang et al., 2011; Ryan et al., 2011). To date, it is unknown if AtASL3 is a transporter of Al influx or efflux (Delhaize et al., 2012). However, it has been suggested that it might participate in Al efflux from the root after absorption of the metal (Larsen et al., 2005). In fact, the Al efflux has been proposed as an exclusion mechanism (Arunakumara et al., 2013). Mutation of the transporter AtALS3 results in hypersensitivity to Al and an increase in the accumulation of the metal in the roots of Arabidopsis (Larsen et al., 1997, 2005). An ALS3 gene with characteristics similar to those of AtASL3 has been identified in poplar (P. tremula); however, it has been suggested that it participates in internal tolerance of Al (Grisel et al., 2010; Brunner and Sperisen, 2013). AtALS1 is also a partial, type ABC protein and probably participates in intracellular transportation of Al in vacuoles of root cells and vascular cells of the plant (Larsen et al., 2007; Delhaize et al., 2012). OsALS1 is an ortholog of AtALS1 in rice and its expression is induced rapidly and specifically by Al. OsALS1 is located in the tonoplast of root cells and participates in the compartmentalization of Al in the vacuoles, which is required for the internal detoxification of Al in rice (Huang et al., 2009, 2012).
The Nrat1 transporter is a member of the Nramp family of transporters located in the PM of all root apex cells, except in the epidermal cells (Xia et al., 2010). The Nrat1 gene is positively regulated by the transcription factor responsive to Al, ART1 in rice (Yamaji et al., 2009). It has been demonstrated that the protein Nrat1 exhibits the activity of Al transportation, but not for divalent metals such as Fe, Mn, and Cd or Al-citrate compounds (Xia et al., 2010, 2011). Moreover, it has been suggested that Nrat1 is required for detoxification of Al in the cell wall as it reduces metal levels through Al-influx to the root cells and their subsequent compartmentalization in the vacuole, possibly by OsALS1 (Xia et al., 2010, 2011, 2014; Huang et al., 2012; Li et al., 2014). Given that the mutant nrat1 presents increased sensitivity to Al and the over-expression of Nrat1 in yeast, rice and Arabidopsis increases Al uptake (Xia et al., 2010, 2011; Li et al., 2014), the Nrat1 gene or its orthologs can be useful tools to enhance Al tolerance in different plant species.
In H. macrophylla, transporters have been identified [members of the aquaporin (AQP) family and anion permeases], located in the tonoplast and plasma membrane, which must work as Al transporters (Negishi et al., 2012, 2013). Recently, Wang et al. (2017) reported that NIP1;2, a plasma membrane-localized member of the Arabidopsis nodulin 26-like intrinsic protein (NIP) subfamily of the AQP family, facilitates Al-malate transport from the root cell wall into the root symplasm, with subsequent Al xylem loading and root-to-shoot translocation, which are critical steps in an internal Al tolerance mechanism in A. thaliana. Surprisingly, NIP1;2 facilitates the transport of Al-malate, but not Al3+ ions. Hence the coordinated function of NIP1;2 and ALMT1 are required for Al uptake, translocation, and tolerance in Arabidopsis (Hoekenga et al., 2006; Wang et al., 2017). Moreover, a recent report mentions that endocytic vesicular traffic can contribute to the internalization of Al in the root apex of rice. The overexpression of OsPIN2 in rice increases tolerance to Al through the internalization of Al via vesicular traffic (Wu et al., 2015).
Modification of Plant Metabolism and DNA Checkpoints
In plants, the mechanisms of exclusion and tolerance to Al are intimately related to mitochondrial activity, mitochondrial metabolism and OA transportation (Nunes-Nesi et al., 2014). The cycle of TCA is inducible by Al, and intervenes in the biosynthesis of OA to chelate Al in the apoplast or cytosol. In Brassica napus, Al induces enzyme activity of OA anabolism and catabolism, as well as the accumulation of OA (Ligaba et al., 2004). Overexpression of the enzymes citrate synthase, malate dehydrogenase and pyruvate phosphate dikinase confers resistance to Al by increasing the synthesis and exudation of OA (Deng et al., 2009; Han et al., 2009; Trejo-Téllez et al., 2010; Wang et al., 2010). In wheat varieties tolerant to Al, exposure to Al positively regulates the expression of mitochondrial ATP synthase and vacuolar H+-ATPasa, suggesting an increase in metabolic activity in the mitochondria and pH changes in the cytosol (Hamilton et al., 2001). Therefore, the increase in mitochondrial biochemical activity is important for the synthesis of OA under stress by Al. In fact, the lack of correlation between OA exudation and resistance to Al in corn, soybean and buckwheat (Piñeros et al., 2005; Nian et al., 2005; Zheng et al., 2005) has suggested that the exudation of OA is a result of the biochemical reactions required for the tolerance to Al (Nunes-Nesi et al., 2014).
Alternative metabolic routes of cellular respiration also participate in tolerance to stress by Al. Under adverse conditions, where carbohydrates are scarce, plants can metabolize proteins and lipids as alternative substrates for cellular respiration (Araújo et al., 2011). Similarly, the overexpression of mitochondrial alternative oxidase (AOX) increases the respiration and reduces the oxidative stress induced by Al in the mitochondria (Panda et al., 2013). Lou et al. (2016) suggested that formate accumulation is involved in both H+ and Al-induced root growth inhibition in rice bean (Vigna umbellate). The overexpression of the formate dehydrogenase (FDH) gene of rice bean (VuFDH) in tobacco (N. tabacum) results in decreased sensitivity to Al and H+ stress due to less production of format in the transgenic tobacco lines under Al and H+ stresses. These findings suggest a possible new route toward the improvement of plant performance in acidic soils, where Al toxicity and H+ stress coexist (Lou et al., 2016). In tomato (Solanum lycopersicum) roots proteins have been identified which play an important role in Al exclusion and tolerance (Zhou S. et al., 2016). Gallagher et al. (1980) evaluated nitrate reductase activity and reported an increase in the activity of this enzyme in crops which are tolerant to Al. Toxicity by Al is also associated with the metabolism of nitrogen (N) (Foy and Fleming, 1982). Transcriptome analyses in lucerne roots reveal candidate Al-stress-responsive genes involved in ribosome, protein biosynthesis, TCA cycle, membrane transport (organic, small molecules and ions) and hormonal regulation. However, the ribosome protein genes was the pathway with the largest numbers of genes differentially up-regulated, which suggested a high biological importance for ribosomal genes and an alternative in response to Al stress in plants (Liu et al., 2017). Also, NAC genes have been postulated to play pivotal roles in plants exposed to Al (Moreno-Alvarado et al., 2017).
Recently identified Arabidopsis mutants with increased Al tolerance provide evidence of DNA as one of the main targets of Al (Eekhout et al., 2017). Al treatment results in binding of Al to the negative charges of the phosphodiester backbone DNA. In fact, nuclei have been reported to accumulate Al even in the presence of low environmental concentrations. Binding Al to DNA might possibly alter DNA topology from the B-DNA to Z-DNA conformation, resulting in increased DNA rigidity that leads to difficulty in unwinding during DNA replication and susceptibility of DNA to endogenous mutagens (Anitha and Rao, 2002; Hu et al., 2016). Also, Al stress gives rise to changes in the localization and expression of the nucleolar proteins and inhibition of DNA synthesis, as well as promoting DNA fragmentation and the generation of micronuclei (Zhang et al., 2014; Sade et al., 2016; Eekhout et al., 2017). In Arabidopsis plants, this Al- induced DNA damage triggered the activation of a cell cycle arrest causing root growth inhibition, at least partly (Rounds and Larsen, 2008). Eekhout et al. (2017) suggested the modification of DNA checkpoints to confer Al-tolerance through DNA repair with cell-cycle progression. The DNA damage response (DDR) pathway maintains genome integrity under adverse conditions that affect DNA replication. The DDR pathway introduces a transient cell-cycle arrest during the process of DNA repair, through the coordinated expression of DNA repair and cell-cycle inhibitory genes, thus ensuring that both the daughter cells inherit a complete and error free copy of the genome. This insight could lead the way for novel strategies to generate Al-tolerant crop plants (Hu et al., 2016; Eekhout et al., 2017).
Conclusion
Much interest has been shown recently in the use of biostimulants and stimulants in agriculture with the aim of increasing root growth, nutrient uptake and tolerance to stress in plants. Plant biostimulants or agricultural stimulants include microorganisms and a diversity of substances, among which are the beneficial elements (Calvo et al., 2014). Aluminum (Al), cobalt (Co), selenium (Se), sodium (Na), and silicon (Si) are considered to be beneficial elements for plants, given that, despite the fact that they are not required by all plants, they can promote growth and are essential for certain plant taxa, depending on the environmental conditions, concentration of the element and plant species. These elements can also increase tolerance to abiotic stress (drought, salinity, high temperatures, cold, UV light, toxicity or nutrient deficiency) as well as biotic stress (pathogens and herbivores) when administered at low concentrations. However, it is important to know the range of concentration in which a beneficial element becomes lethal, in particular with respect to its use as a fertilizer to increase the production of crops under conditions of stress and/or improve the nutritional value of food plants (Pilon-Smits et al., 2009; Kaur et al., 2016).
Aluminum stimulates growth in plants of economic importance such as the tea shrub and can maintain or fix floral colors, as in the case of hydrangeas; therefore its application as a biostimulant of a desirable response in these plants is feasible (Fang et al., 2014). However, high concentrations of Al can pose a serious threat to agricultural production due to inhibition of root elongation and plant growth through a diversity of mechanisms with the participation of Al, including interactions in the symplast, plasma membrane and the cell wall (Kochian et al., 2015). In addition to the plants, Al can also cause serious problems in the nervous system, lungs and kidneys of human beings. Tea is an important dietetic source of Al for human beings. There is a great need to understand how environmental factors can have an influence on the accumulation of Al in tea leaves in order to create strategies for the reduction of Al uptake in tea plants (de Silva et al., 2016). A number of studies have suggested that people who are exposed to high levels of Al can develop Alzheimer’s disease, encephalopathy and dementia, among other diseases (Anitha and Rao, 2002; Mehra and Baker, 2007; Ashenef, 2014).
The root system is complex and a wide variety of root phenotypes have been identified which contribute to the adaptation to toxicity by Al (Rao et al., 2016). A greater root surface induced by Al can increase the uptake of water and nutrients by plants (Hajiboland et al., 2013b), mainly in conditions of stress caused by drought, salinity and nutrient deficiency. Further studies on the use of low concentrations of Al to prevent the effect of different conditions of stress must be considered.
Author Contributions
EB-Q designed, planned, wrote, and checked the manuscript. CE-M wrote the manuscript. IE-M checked the manuscript. MM-E group leader, wrote and checked the manuscript.
Funding
Recent studies carried out by our group have been financially supported by the Mexico’s National Council of Science and Technology (CONACYT; grant 166621).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
We are grateful to Mexico’s National Council of Science and Technology (CONACYT) for supporting our studies through the Grant 166621.
References
Ahn, S. J., Sivaguru, M., Chung, G. C., Rengel, Z., and Matsumoto, H. (2002). Aluminium-induced growth inhibition is associated with impaired efflux and influx of H+ across the plasma membrane in root apices of squash (Cucurbita pepo). J. Exp. Bot. 53, 1959–1966. doi: 10.1093/jxb/erf049
Ahn, S. J., Sivaguru, M., Osawa, H., Chung, G. C., and Matsumoto, H. (2001). Aluminum inhibits the H+-ATPase activity by permanently altering the plasma membrane surface potentials in squash roots. Plant Physiol. 126, 1381–1390. doi: 10.1104/pp.126.4.1381
Andrivon, D. (1995). Inhibition by aluminum of mycelia growth and of sporangial production and germination in Phytophthora infestans. Eur. J. Plant Pathol. 101, 527–533. doi: 10.1007/BF01874477
Anitha, S., and Rao, K. S. J. (2002). The complexity of aluminum-DNA interactions: relevance to Alzheimer’s and other neurological diseases. Struct. Bond. 104, 79–97. doi: 10.1007/3-540-45425-X_3
Arasimowicz-Jelonek, M., Floryszak-Wieczorek, J., Drzewiecka, K., Chmielowska-Ba, K. J., Abramowski, D., and Izbianska, K. (2014). Aluminum induces cross-resistance of potato to Phytophthora infestans. Planta 239, 679–694. doi: 10.1007/s00425-013-2008-8
Araújo, W. L., Tohge, T., Ishizaki, K., Leaver, C. J., and Fernie, A. R. (2011). Protein degradation- an alternative respiratory substrate for stressed plants. Trends Plant Sci. 16, 489–498. doi: 10.1016/j.tplants.2011.05.008
Archambault, D. J., Zhang, G., and Taylor, G. J. (1996). Accumulation of Al in root mucilage of an Al-resistant and an Al-sensitive cultivar of wheat. Plant Physiol. 112, 1471–1478. doi: 10.1104/pp.112.4.1471
Arunakumara, K. K. I. U., Walpola, B. C., and Yoon, M.-H. (2013). Aluminum toxicity and tolerance mechanism in cereals and legumes- a review. J. Korean Soc. Appl. Biol. Chem. 56, 1–9. doi: 10.1007/s13765-012-2314-z
Ashenef, A. (2014). Essential and toxic metals in tea (Camellia sinensis) imported and produced in Ethiopia. Food Addit. Contam. Part B 7, 30–36. doi: 10.1080/19393210.2013.831951
Asher, C. J. (1991). “Beneficial elements, functional nutrients, and possible new essential elements,” in Micronutrients in Agriculture, 2nd Edn, eds J. J. Mortvedt, F. R. Cox, L. M. Shuman, and R. M. Welch (Madison WI: Soil Science Society-America), 703–723.
Barcelo, J., and Poschenrieder, C. (2002). Fast root growth responses, root exudates, and internal detoxification as clues to the mechanisms of aluminium toxicity and resistance: a review. Environ. Exp. Bot. 48, 75–92. doi: 10.1016/S0098-8472(02)00013-8
Bojórquez-Quintal, J. E. A., Sánchez-Cach, L. A., Ku-González, A., de los Santos-Briones, C., Medina-Lara, M. F., Echevarría-Machado, I., et al. (2014). Differential effects of aluminum on in vitro primary root growth, nutrient content and phospholipase C activity in coffee seedlings (Coffea arabica). J. Inorg. Biochem. 134, 39–48. doi: 10.1016/j.jinorgbio.2014.01.018
Bose, J., Babourina, O., Ma, Y., Zhou, M., Shabala, S., and Rengel, Z. (2015). “Specificity of ion uptake and homeostasis maintenance during acid and aluminium stresses,” in Aluminum Stress Adaptation in Plants. Signaling Communication in Plants, Vol. 24, eds S. K. Panda and F. Baluska (Cham: Springer International Publishing), 229–251. doi: 10.1007/978-3-319-19968-9_12
Bose, J., Babourina, O., and Rengel, Z. (2011). Role of magnesium in alleviation of aluminium toxicity in plants. J. Exp. Bot. 62, 2251–2264. doi: 10.1093/jxb/erq456
Bose, J., Babourina, O., Shabala, S., and Rengel, Z. (2010a). Aluminium induced ion transport in Arabidopsis: the relationship between Al tolerance and root ion flux. J. Exp. Bot. 61, 3163–3175. doi: 10.1093/jxb/erq143
Bose, J., Babourina, O., Shabala, S., and Rengel, Z. (2010b). Aluminum dependent dynamics of ion transport in Arabidopsis: specificity of low pH and aluminum responses. Physiol. Plant. 139, 401–412. doi: 10.1111/j.1399-3054.2010.01377.x
Bose, J., Babourina, O., Shabala, S., and Rengel, Z. (2013). Low-pH and aluminum resistance in Arabidopsis correlates with high cytosolic magnesium content and increased magnesium uptake by plant roots. Plant Cell Physiol. 54, 1093–1104. doi: 10.1093/pcp/pct064
Brautigan, D. J., Rengasamy, P., and Chittleborough, D. J. (2012). Aluminium speciation and phytotoxicity in alkaline soils. Plant Soil 360, 187–196. doi: 10.1007/s11104-012-1232-5
Brunner, I., and Sperisen, C. (2013). Aluminum exclusion and aluminum tolerance in woody plants. Front. Plant Sci. 4:172. doi: 10.3389/fpls.2013.00172
Cai, M., Wang, N., Xing, C., Wang, F., Wu, K., and Du, X. (2013). Immobilization of aluminum with mucilage secreted by root cap and root border cells is related to aluminum resistance in Glycine max L. Environ. Sci. Pollut. Res. 20, 8924–8933. doi: 10.1007/s11356-013-1815-6
Calvo, P., Nelson, L., and Kloepper, J. W. (2014). Agricultural uses of plant biostimulants. Plant Soil 383, 3–41. doi: 10.1007/s11104-014-2131-8
Che, J., Yamaji, N., Shen, R. F., and Ma, J. F. (2016). An Al-inducible expansin gene, OsEXPA10 is involved in root cell elongation of rice. Plant J. 88, 132–142. doi: 10.1111/tpj.13237
Chen, W., Xu, C., Zhao, B., Wang, X., and Wang, Y. (2008). Improved Al tolerance of saffron (Crocus sativus L.) by exogenous polyamines. Acta Physiol. Plant. 30, 121–127. doi: 10.1007/s11738-007-0100-z
Daikuhara, G. (1914). “Üeber saure mineralböden,” in Agricultural Experiment Station. The Bulletin of the Imperial Central Agricultural Experiment Stations Japan, Vol. 2 (Tokyo: Nishigahara Station), 18.
de Silva, J., Tuwei, G., and Zhao, F.-J. (2016). Environmental factors influencing aluminium accumulation in tea (Camellia sinensis L.). Plant Soil 400, 223–230. doi: 10.1007/s11104-015-2729-5
Degenhardt, J., Larsen, P. B., Howell, S. H., and Kochian, L. V. (1998). Aluminium resistance in the Arabidopsis mutant alr-104 is caused by an aluminium induced increase in rhizosphere pH. Plant Physiol. 117, 19–27. doi: 10.1104/pp.117.1.19
Delhaize, E., Craig, S., Beaton, C. D., Bennet, R. J., Jagadish, V. C., and Randall, P. J. (1993). Aluminum tolerance in wheat (Triticum aestivum L.) I. Uptake and distribution of aluminum in root apices. Plant Physiol. 103, 685–693. doi: 10.1104/pp.103.3.695
Delhaize, E., Gruber, B. D., and Ryan, P. R. (2007). The roles of organic anion permeases in aluminium resistance and mineral nutrition. FEBS Lett. 581, 2255–2262. doi: 10.1016/j.febslet.2007.03.057
Delhaize, E., Ma, J. F., and Ryan, P. R. (2012). Transcriptional regulation of aluminium tolerance genes. Trends Plant Sci. 17, 341–348. doi: 10.1016/j.tplants.2012.02.008
Delhaize, E., and Ryan, P. R. (1995). Aluminum toxicity and tolerance in plants. Plant Physiol. 107, 315–321. doi: 10.1104/pp.107.2.315
Delhaize, E., Ryan, P. R., Hebb, D. M., Yamamoto, Y., Sasaki, T., and Matsumoto, H. (2004). Engineering high-level aluminum tolerance in barley with the ALMT1 gene. Proc. Natl. Acad. Sci. U.S.A. 101, 15249–15254. doi: 10.1073/pnas.0406258101
Deng, W., Luo, K., Li, Z., Yang, Y., Hu, N., and Wu, Y. (2009). Overexpression of Citrus junos mitochondrial citrate synthase gene in Nicotiana benthamiana confers aluminum tolerance. Planta 230, 355–365. doi: 10.1007/s00425-009-0945-z
Eekhout, T., Larsen, P., and De Veylder, L. (2017). Modification of DNA checkpoints to confer aluminum tolerance. Trends Plant Sci. 22, 102–105. doi: 10.1016/j.tplants.2016.12.003
Endo, I., Tange, T., and Osawa, H. (2009). “The protective role of a unique tissue detachment on root cap cells in a strong aluminum-resistant tree Acacia mangium,” in Proceedings of the International Plant Nutrition Colloquium XVI, (Davis, CA: University of California).
Endo, I., Tange, T., and Osawa, H. (2011). A cell-type-specific defect in border cell formation in the Acacia mangium root cap developing an extraordinary sheath of sloughed-off cells. Ann. Bot. 108, 279–290. doi: 10.1093/aob/mcr139
Eticha, D., Stass, A., and Horst, W. J. (2005). Cell-wall pectin and its degree of methylation in the maize root-apex: significance for genotypic differences in aluminium resistance. Plant Cell Environ. 28, 1410–1420. doi: 10.1111/j.1365-3040.2005.01375.x
Famoso, A. N., Clark, R. T., Shaff, J. E., Craft, E., McCouch, S. R., and Kochian, L. V. (2010). Development of a novel aluminum tolerance phenotyping platform used for comparisons of cereal aluminum tolerance and investigations into rice aluminum tolerance mechanisms. Plant Physiol. 153, 1678–1691. doi: 10.1104/pp.110.156794
Famoso, A. N., Zhao, K., Clark, R. T., Tung, C.-W., Wright, M. H., Bustamante, C., et al. (2011). Genetic architecture of aluminum tolerance in rice (Oryza sativa) determined through genome-wide association analysis and QTL mapping. PLOS Genet. 7:e1002221. doi: 10.1371/journal.pgen.1002221
Fang, X.-M., Chen, F.-S., Hu, X.-F., Yuan, P.-C., Li, J., and Chen, X. (2014). Aluminum and nutrient interplay across an age-chronosequence of tea plantations within a hilly red soil farm of subtropical China. J. Soil Sci. Plant Nutr. 60, 448–459. doi: 10.1080/00380768.2014.912950
Foy, C. D. (1974). “Effects of aluminum on plant growth,” in The Plant Root and Its Environment, ed. E. W. Carson (Charlottesville, VA: University of Virginia), 601–642.
Foy, C. D., Burn, G. R., Brown, J. C., and Fleming, A. L. (1965). Differential aluminum tolerance of two wheat varieties associated with plant-induced pH changes around their roots. Soil Sci. Soc. Am. Proc. 29, 64–67. doi: 10.2136/sssaj1965.03615995002900010019x
Foy, C. D., Chaney, R. L., and White, M. C. (1978). The physiology of metal toxicity in plants. Annu. Rev. Plant Physiol. 29, 511–566. doi: 10.1146/annurev.pp.29.060178.002455
Foy, C. D., and Fleming, A. L. (1982). Aluminium tolerance of two wheat cultivars related to nitrate reductase activities. J. Plant. Nutr. 5, 1313–1333. doi: 10.1080/01904168209363064
Fung, K. F., Carr, H. P., Zhang, J., and Wong, M. H. (2008). Growth and nutrient uptake of tea under different aluminium concentrations. J. Sci. Food Agric. 88, 1582–1591. doi: 10.1002/jsfa.3254
Furukawa, J., Yamaji, N., Wang, H., Mitani, N., Murata, Y., Sato, K., et al. (2007). An aluminum-activated citrate transporter in barley. Plant Cell Physiol. 48, 1081–1091. doi: 10.1093/pcp/pcm091
Gallagher, L. W., Soliman, K. M., Qualset, C. O., Huffaker, R. C., and Rains, D. W. (1980). Major gene control of nitrate reductase activity in common wheat. Crop Sci. 20, 717–721. doi: 10.2135/cropsci1980.0011183X002000060010x
García-Oliveira, A., Martins-Lopes, P., Tolrà, R., Poschenrieder, C., Tarquis, M., Guedes-Pinto, H., et al. (2014). Molecular characterization of the citrate transporter gene TaMATE1 and expression analysis of upstream genes involved in organic acid transport under Al stress in bread wheat (Triticum aestivum). Physiol. Plant. 152, 441–452. doi: 10.1111/ppl.12179
Ghanati, F., Morita, A., and Yokota, H. (2005). Effects of aluminium on the growth of tea plant and activation of antioxidant system. Plant Soil 276, 133–141. doi: 10.1007/s11104-005-3697-y
Golding, L. A., Angel, B. M., Batley, G. E., Apte, S. C., Krassoi, R., and Doyle, C. J. (2015). Derivation of a water quality guideline for aluminium in marine waters. Environ. Toxicol. Chem. 34, 141–151. doi: 10.1002/etc.2771
González-Santana, I. H., Márquez-Guzmán, J., Cram-Heydrich, S., and Cruz-Ortega, R. (2012). Conostegia xalapensis (Melastomataceae): an aluminum accumulator plant. Physiol. Plant. 144, 134–145. doi: 10.1111/j.1399-3054.2011.01527.x
Grisel, N., Zoller, S., Künzli-Gontarczyk, M., Lampart, T., Münsterkötter, M., Brunner, I., et al. (2010). Transcriptome responses to aluminum stress in roots of aspen (Populus tremula). BMC Plant Biol. 10:185. doi: 10.1186/1471-2229-10-185
Gruber, B. D., Ryan, P. R., Richardson, A. E., Tyerman, S. D., Ramesh, S., Hebb, D. M., et al. (2010). HvALMT1 from barley is involved in the transport of organic anions. J. Exp. Bot. 61, 1455–1467. doi: 10.1093/jxb/erq023
Gupta, N., Gaurav, S. S., and Kumar, A. (2013). Molecular basis of aluminum toxicity in plants: a review. Am. J. Plant Sci. 4, 21–37. doi: 10.4236/ajps.2013.412A3004
Hagvall, K., Persson, P., and Karlsson, T. (2015). Speciation of aluminum in soils and stream waters: The importance of organic matter. Chem. Geol. 417, 32–43. doi: 10.1016/j.chemgeo.2015.09.012
Hajiboland, R., Bahrami-Rad, S., and Bastani, S. (2014). Aluminum alleviates boron-deficiency induced growth impairment in tea plants. Biol. Plant. 58, 717–724. doi: 10.1007/s10535-014-0425-6
Hajiboland, R., Barceló, J., Poschenrieder, C., and Tolrá, R. (2013a). Amelioration of iron toxicity: a mechanism for aluminum-induced growth stimulation in tea plants. J. Inorg. Biochem. 128, 183–187. doi: 10.1016/j.jinorgbio.2013.07.007
Hajiboland, R., Bahrami Rad, S., Barceló, J., and Poschenrieder, C. (2013b). Mechanisms of aluminum-induced growth stimulation in tea (Camellia sinensis). J. Plant Nutr. Soil Sci. 176, 616–625. doi: 10.1002/jpln.201200311
Hajiboland, R., Bastani, S., Bahrami-Rad, S., and Poschenrieder, C. (2015). Interactions between aluminum and boron in tea (Camellia sinensis) plants. Acta Physiol. Plant. 37, 54. doi: 10.1007/s11738-015-1803-1
Hamilton, C. A., Good, A. G., and Taylor, G. J. (2001). Induction of vacuolar ATPase and mitochondrial ATP synthase by aluminum in an aluminum-resistant cultivar of wheat. Plant Physiol. 125, 2068–2077.
Han, Y., Zhang, W., Zhang, B., Zhang, S., Wang, W., and Ming, F. (2009). One novel mitochondrial citrate synthase from Oryza sativa L. can enhance aluminum tolerance in transgenic tobacco. Mol. Biotechnol. 42, 299–305. doi: 10.1007/s12033-009-9162-z
Hawes, M. C., McLain, J., Ramirez-Andreotta, M., Curlango-Rivera, G., Flores-Lara, Y., and Brigham, L. A. (2016). Extracellular trapping of soil contaminants by root border cells: new insights into plant defense. Agronomy. 6, 5. doi: 10.3390/agronomy6010005
Hoekenga, O., Maron, L. G., Pineros, M. A., and Cancado, P. (2006). AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 103, 9738–9743. doi: 10.1073/pnas.0602868103
Horst, W. J., Wagner, A., and Maschner, H. (1982). Mucilage protects root meristem from aluminum injury. Z. Pflanzenphysiol. 105, 435–444. doi: 10.1016/S0044-328X(82)80041-X
Horst, W. J., Wang, Y., and Eticha, D. (2010). The role of the root apoplast in aluminum-induced inhibition of root elongation and in aluminum resistance of plants: a review. Ann. Bot. 106, 185–197. doi: 10.1093/aob/mcq053
Hossain, A. K. M. Z., Md Asgar, A., Hossain, M. A., Tosaki, T., Koyama, H., and Hara, T. (2005). Boron-calcium synergically alleviates aluminum toxicity in wheat plants (Triticum aestivum L.). Soil Sci. Plant Nutr. 51, 43–49. doi: 10.1111/j.1747-0765.2005.tb00005.x
Houman, Y., Tahara, K., Shinmachi, F., Noguchi, A., Satohiko, S., and Hasegawa, I. (2009). “Differences of growth response to aluminum excess of two Melaleuca trees differing in aluminum resistance,” in Proceedings of the International Plant Nutrition Colloquium XVI, (Davis, CA: University of California).
Hu, Z., Cools, T., and De Veylder, L. (2016). Mechanisms used by plants to cope with DNA damage. Annu. Rev. Plant Biol. 67, 439–462. doi: 10.1146/annurev-arplant-043015-111902
Huang, C. F., Yamaji, N., Chen, Z., and Ma, J. F. (2012). A tonoplast-localized half-size ABC transporter is required for internal detoxification of aluminum in rice. Plant J. 69, 857–867. doi: 10.1111/j.1365-313X.2011.04837.x
Huang, C. F., Yamaji, N., Mitani, N., Yano, M., Nagamura, Y., and Ma, J. F. (2009). A bacterial-type ABC transporter is involved in aluminum tolerance in rice. Plant Cell 21, 655–667. doi: 10.1105/tpc.108.064543
Hwang, J.-U., Song, W.-Y., Hong, D., Ko, D., Yamaoka, Y., Jang, S., et al. (2016). Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol. Plant. 9, 338–355. doi: 10.1016/j.molp.2016.02.003
Igual, J. M., and Dawson, J. O. (1999). Stimulatory effects of aluminum on in vitro growth of Frankia. Can. J. Bot. 77, 1321–1326. doi: 10.1139/b99-061
Ito, D., Shinkai, Y., Kato, Y., Kondo, T., and Yoshida, K. (2009). Chemical studies on different color development in blue-and red-colored sepal cells of Hydrangea macrophylla. Biosci. Biotechnol. Biochem. 73, 1054–1059. doi: 10.1271/bbb.80831
Jackson, M. L., and Huang, P. M. (1983). Aluminum of acid soils in the food chain and senility. Sci. Total Environ. 28, 269–276.
Javed, M. T., Stoltz, E., Lindberg, S., and Greger, M. (2013). Changes in pH and organic acids in mucilage of Eriophorum angustifolium roots after exposure to elevated concentrations of toxic elements. Environ. Sci. Pollut. Res. Int. 20, 1876–1880. doi: 10.1007/s11356-012-1413-z
Jianq, H. X., Tanq, N., Zheng, J. G., Li, Y., and Chen, L. S. (2009). Phosphorus alleviates aluminum-induced inhibition of growth and photosynthesis in Citrus grandis seedlings. Physiol. Plant. 137, 298–311. doi: 10.1111/j.1399-3054.2009.01288.x
Jones, D. L., Blancaflor, E. B., Kochian, L. V., and Gilroy, S. (2006). Spatial coordination of aluminium uptake, production of reactive oxygen species, callose production and wall rigidification in maize roots. Plant Cell Environ. 29, 1309–1318. doi: 10.1111/j.1365-3040.2006.01509.x
Jones, D. L., Kochian, L. V., and Gilroy, S. (1998). Aluminum induces a decrease in cytosolic calcium concentration in BY-2 tobacco cell cultures. Plant Physiol. 116, 81–89.
Jones, T. R., Antonetti, D. L., and Reid, T. W. (1986). Aluminum ions stimulate mitosis in murine cells in tissue culture. J. Cell Biochem. 30, 31–39. doi: 10.1002/jcb.240300105
Kang, J., Park, J., Choi, H., Burla, B., Kretzschmar, T., Lee, Y., et al. (2011). Plant ABC transporters. Arabidopsis Book. 9:e0153. doi: 10.1199/tab.0153
Kaur, S., Kaur, N., Siddique, K. H. M., and Nayyar, H. (2016). Beneficial elements for agricultural crops and their functional relevance in defense against stresses. Arch. Agron. Soil Sci. 62, 905–920. doi: 10.1080/03650340.2015.1101070
Khan, M. S. H., Tawaraya, K., Sekimoto, H., Koyama, H., Kobayashi, Y., Murayama, T., et al. (2009). Relative abundance of delta5-sterols in plasma membrane lipids of root-tip cells correlates with aluminum tolerance of rice. Physiol. Plant. 135, 73–83. doi: 10.1111/j.1399-3054.2008.01175.x
Kidd, P. S., Llugany, M., Poschenrieder, C., Gunsé, B., and Barceló, J. (2001). The role of root exudates in aluminium resistance and silicon-induced amelioration of aluminium toxicity in three varieties of maize (Zea mays L.). J. Exp. Bot. 52, 1339–1352.
Kidd, P. S., and Proctor, J. (2000). Effects of aluminium on the growth and mineral composition of Betula pendula Roth. J. Exp. Bot. 51, 1057–1066.
Kinraide, T. B. (1991). Identity of the rhizotoxic aluminum species. Plant Soil. 134, 167–178. doi: 10.1007/BF00010729
Kinraide, T. B. (1993). Aluminum enhancement of plant growth in acid rooting media – a case of reciprocal alleviation of toxicity by two toxic cations. Physiol. Plant. 88, 619–625. doi: 10.1111/j.1399-3054.1993.tb01380.x
Kinraide, T. B. (1998). Three mechanisms for the calcium alleviation of mineral toxicities. Plant Physiol. 118, 513–520.
Kinraide, T. B., Ryan, P. R., and Kochian, L. V. (1992). Interactive effects of Al, H+, and other cations on root elongation considered in terms of cell-surface electrical potential. Plant Physiol. 99, 1461–1468.
Kisnieriené, V., and Lapeikaité, I. (2015). When chemistry meets biology: the case of aluminium- a review. Chemija 26, 148–158.
Kochian, L. V., Hoekenga, O. A., and Pineros, M. A. (2004). How do crop plants tolerate acid soils?-mechanisms of aluminium tolerance and phosphorous efficiency. Annu. Rev. Plant Biol. 55, 459–493. doi: 10.1146/annurev.arplant.55.031903.141655
Kochian, L. V., Pineros, M. A., and Hoekenga, O. A. (2005). The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant Soil 274, 175–195. doi: 10.1007/s11104-004-1158-7
Kochian, L. V., Piñeros, M. A., Liu, J., and Magalhaes, J. V. (2015). Plant adaptation to acid soils: the molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 66, 571–598. doi: 10.1146/annurev-arplant-043014-114822
Kodama, M., Tanabe, Y., and Nakayama, M. (2016). Analyses of coloration-related components in Hydrangea sepals causing color variability according to soil conditions. Hortic. J. 85, 372–379. doi: 10.2503/hortj.MI-131
Kolaei, E. A., Cenatus, C., Tweddell, R. J., and Avis, T. J. (2013). Antifungal activity of aluminium-containing salts against the development of carrot cavity and potato dry rot. Ann. Appl. Biol. 163, 311–317. doi: 10.1111/aab.12056
Konishi, S., Miyamoto, S., and Taki, T. (1985). Stimulatory effects of aluminium on tea plants grown under low and high phosphorus supply. J. Soil Sci. Plant Nutr. 31, 361–368. doi: 10.1080/00380768.1985.10557443
Kopittke, P. M., and Blamey, F. P. C. (2016). Theoretical and experimental assessment of nutrient solution composition in short-term studies of aluminium rhizotoxicity. Plant Soil 406, 311–326. doi: 10.1007/s11104-016-2890-5
Kopittke, P. M., Menzies, N. W., Wang, P., and Blamey, F. P. C. (2016). Kinetic and nature of aluminium rhizotoxic effects: a review. J. Exp. Bot. 67, 4451–4467. doi: 10.1093/jxb/erw233
Kopittke, P. M., Moore, K. L., Lombi, E., Gianoncelli, A., Ferguson, B. J., Blamey, F. P. C., et al. (2015). Identification of the primary lesion of toxic aluminum in plant roots. Plant Physiol. 167, 1402–1411. doi: 10.1104/pp.114.253229
Kumpf, R. P., and Nowack, M. K. (2015). The root cap: a short story of life and death. J. Exp. Bot. 66, 5651–5662. doi: 10.1093/jxb/erv295
Larsen, P. B., Cancel, J., Rounds, M., and Ochoa, V. (2007). Arabidopsis ALS1 encodes a root tip and stele localized half type ABC transporter required for root growth in an aluminum toxic environment. Planta 225, 1447–1458. doi: 10.1007/s00425-006-0452-4
Larsen, P. B., Degenhardt, J., Tai, C. Y., Stenzler, L. M., Howell, S. H., and Kochian, L. V. (1998). Aluminum-resistant Arabidopsis mutants that exhibit altered patterns of aluminum accumulation and organic acid release from roots. Plant Physiol. 117, 9–18. doi: 10.1104/pp.117.1.9
Larsen, P. B., Geisler, M. J. B., Jones, C. A., Williams, K. M., and Cancel, J. D. (2005). ALS3 encodes a phloem-localized ABC transporter-like protein that is required for aluminum tolerance in Arabidopsis. Plant J. 41, 353–363. doi: 10.1111/j.1365-313X.2004.02306
Larsen, P. B., Kochian, L. V., and Howell, S. H. (1997). Al inhibits both shoot development and root growth in als3, an Al-sensitive Arabidopsis mutant. Plant Physiol. 114, 1207–1214.
Lau, K.-H. W., Yoo, A., and Wang, S. P. (1991). Aluminum stimulates the proliferation and differentiation of osteoblasts in vitro by a mechanism that is different from fluoride. Mol. Cell Biochem. 105, 93–105. doi: 10.1007/BF00227749
Levesque-Tremblay, G., Pelloux, J., Braybrook, S. A., and Müller, K. (2015). Tunning of pectin methylesterification: consequences for cell wall biomechanics and development. Planta 242, 791–811. doi: 10.1007/s00425-015-2358-5
Li, J. Y., Liu, J., Dong, D., Jia, X., McCouch, S. R., and Kochian, L. V. (2014). Natural variation underlies alterations in Nramp aluminum transporter (NRAT1) expression and function that play a key role in rice aluminum tolerance. Proc. Natl. Acad. Sci. U.S.A. 111, 6503–6508. doi: 10.1073/pnas.1318975111
Li, X. F., Ma, J. F., Hiradate, S., and Matsumoto, H. (2000). Mucilage strongly bind aluminum but does not prevent roots from aluminum injury in Zea may. Physiol. Plant. 108, 152–160. doi: 10.1034/j.1399-3054.2000.108002152.x
Ligaba, A., Katsuhura, M., Ryan, P. R., Shibasaka, M., and Matsumoto, H. (2006). The BnALMT1 and BnALMT2 genes from rape encode aluminum-activated malate transporters that enhance the aluminum resistance of plant cells. Plant Physiol. 142, 1294–1303.
Ligaba, A., Maron, L., Shaff, J., Kochian, L. V., and Piñeros, M. (2012). Maize ZmALMT2 is a root anion transporter that mediates constitutive root malate efflux. Plant Cell Environ. 35, 1185–1200. doi: 10.1111/j.1365-3040.2011.02479.x
Ligaba, A., Shen, H., Shibata, K., Yamamoto, Y., Tanakamaru, S., and Matsumoto, H. (2004). The role of phosphorus in aluminum-induced citrate and malate exudation in rape (Brassica napus L.). Physiol. Plant. 120, 575–584. doi: 10.1111/j.0031-9317.2004.0290
Liu, J., Magalhaes, J. V., Shaff, J., and Kochian, L. V. (2009). Aluminum-activated citrate and malate transporters from the MATE and ALMT families function independently to confer Arabidopsis aluminum tolerance. Plant J. 57, 389–399. doi: 10.1111/j.1365-313X.2008.03696
Liu, S., Gao, H., Wu, X., Fang, Q., Chen, L., Zhao, F.-J., et al. (2016). Isolation and characterization of an aluminum-resistant mutant in rice. Rice 9, 60. doi: 10.1186/s12284-016-0132-3
Liu, J., Li, Y., Wang, W., Gai, J., and Li, Y. (2016). Genome-wide analysis of MATE transporters and expression patterns of a subgroup of MATE genes in response to aluminum toxicity in soybean. BMC Genomics 17:223. doi: 10.1186/s12864-016-2559-8
Liu, W., Xiong, C., Yan, L., Zhang, Z., Ma, L., Wang, Y., et al. (2017). Transcriptome analyses reveal candidate genes potentially involved in Al stress response in alfalfa. Front. Plant Sci. 8:26. doi: 10.3389/fpls.2017.00026
Llugany, M., Poschenrieder, C., and Barceló, J. (1995). Monitoring of aluminium-induced inhibition of root elongation in four maize cultivars differing in tolerance to aluminium-induced inhibition of root elongation in four maize cultivars differing in tolerance to aluminium and proton toxicity. Physiol. Plant. 93, 265–271. doi: 10.1111/j.1399-3054.1995.tb02227.x
Lou, H. Q., Gong, Y. L., Fan, W., Xu, J. M., Liu, Y., Cao, M. J., et al. (2016). A formate dehydrogenase confers tolerance to aluminum and low pH. Plant Physiol. 171, 294–305. doi: 10.1104/pp.16.01105
Ma, J. F., and Hiradate, S. (2000). Form of aluminium for uptake and translocation in buckwheat (Fagopyrum esculentum Moench). Planta 211, 355–360. doi: 10.1007/s004250000292
Ma, J. F., Hiradate, S., and Matsumoto, H. (1998). High aluminum resistance in buckwheat: II. Oxalic acid detoxifies aluminum internally. Plant Physiol. 117, 753–759. doi: 10.1104/pp.117.3.753
Ma, J. F., Hiradate, S., Nomoto, K., Iwashita, T., and Matsumoto, H. (1997). Internal detoxification mechanism of Al in hydrangea: identification of Al form in the leaves. Plant Physiol. 113, 1033–1039.
Ma, J. F., Ryan, P. R., and Delhaize, E. (2001). Aluminum tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 6, 273–278.
Ma, Q., Rengel, Z., and Kuo, J. (2002). Aluminum toxicity in rye (Secale cereale): root growth and dynamics of cytoplasmic Ca in intact root tips. Ann. Bot. 89, 241–244. doi: 10.1093/aob/mcf017
Ma, Y., Zhu, M., Shabala, L., Zhou, M., and Shabala, S. (2016). Conditioning of roots with hypoxia increases aluminum and acid stress tolerance by mitigating activation of K+ efflux channels by ROS in Barley: Insights into croos-tolerance mechanisms. Plant Cell Physiol. 57, 160–173. doi: 10.1093/pcp/pcv175
Maejima, E., and Watanabe, T. (2014). Proportion of phospholipids in the plasma membrane is an important factor in Al tolerance. Plant Signal. Behav. 9, e29277.
Maejima, E., Watanabe, T., Osaki, M., and Wagatsuma, T. (2014). Phosphorus deficiency enhances aluminum tolerance of rice (Oryza sativa) by changing the physicochemical characteristics of root plasma membranes and cell walls. J. Plant Physiol. 171, 9–15. doi: 10.1016/j.jplph.2013.09.012
Magalhaes, J. V., Liu, J., Guimaraes, C. T., Lana, U. G., Alves, V. M., Wang, Y. H., et al. (2007). A gene in the multidrug and toxic compound extrusion (MATE) family confers aluminum tolerance in sorghum. Nat. Genet. 39, 1156–1161. doi: 10.1038/ng2074
Magistad, O. C. (1925). The aluminum content of the soil solution and its relation to soil reaction and plant growth. Soil Sci. 20, 181–226.
Malta, P. G., Arcanjo-Silva, S., Ribeiro, C., Campos, N. V., and Alves-Azevedo, A. (2016). Rudgea viburnoides (Rubiaceae) overcomes the low soil fertility of the Brazilian Cerrado and hyperaccumulates aluminum in cell walls and chloroplast. Plant Soil 408, 369–384. doi: 10.1007/s11104-016-2926-x
Maron, L., Piñeros, M. A., Guimarães, C., Magalhaes, J., Pleiman, J., Mao, C., et al. (2010). Two functionally distinct members of the MATE (multidrug and toxic compound extrusion) family of transporters potentially underlie two major Al tolerance QTL in maize. Plant J. 61, 728–740. doi: 10.1111/j.1365-313X.2009.04103.x
Maron, L. G., Guimarães, C. T., Kirst, M., Albert, P. S., Birchler, J. A., Bradbury, P., et al. (2013). Aluminum tolerance is associated with higher MATE1 gene copy-number in maize. Proc. Natl. Acad. Sci. U.S.A. 110, 5241–5246. doi: 10.1073/pnas.1220766110
Maze, P. (1915). Determination des Elements inineraux rates ilfcessair’es an developpement du maïs. Conlpt. Fend. Acad. Sci. Paris 160, 211.
Mehra, A., and Baker, C. L. (2007). Leaching and bioavailability of aluminium, copper and manganese from tea (Camellia sinensis). Food Chem. 100, 1456–1463. doi: 10.1016/j.foodchem.2005.11.038
Meyer, J. R., Shew, H. D., and Harrison, U. J. (1994). Inhibition of germination and growth of Thielaviopsis basicola by aluminum. Phytopathology 84, 598–602.
Miyake, K. (1916). The toxic action of soluble aluminium salts upon the growth of the rice plant. J. Biol. Chem. 25, 23–28.
Miyasaka, S. C., and Hawes, M. C. (2001). Possible role of root border cells in detection and avoidance of aluminum toxicity. Plant Physiol. 125, 1978–1987. doi: 10.1104/pp.125.4.1978
Morel, J. L., Mench, M., and Guckert, A. (1986). Measurement of Pb2+, Cu2+ and Cd2+ binding with mucilage exudates from maize (Zea mays L.) roots. Biol. Fertil. Soils 2, 29–34. doi: 10.1007/BF00638958
Moreno-Alvarado, M., García-Morales, S., Trejo-Téllez, L. I., Hidalgo-Contreras, J. V., and Gómez-Merino, F. C. (2017). Aluminum enhances growth and sugar concentration, alters macronutrient status and regulates the expression of NAC transcription factors in rice. Front. Plant Sci. 8:73. doi: 10.3389/fpls.2017.00073
Morita, A., Horie, H., Fujii, Y., Takatsu, S., Watanabe, N., Yagi, A., et al. (2004). Chemical forms of aluminum in xylem sap of tea plants (Camellia sinensis L.). Phytochemistry 65, 2775–2780. doi: 10.1016/j.phytochem.2004.08.043
Morita, A., Yanagisawa, O., Maeda, S., Takatsu, S., and Ikka, T. (2011). Tea plant (Camellia sinensis L.) roots secrete oxalic acid and caffeine into medium containing aluminum. Soil Sci. Plant Nutr. 57, 796–802. doi: 10.1080/00380768.2011.629176
Morita, A., Yanagisawa, O., Takatsu, S., Maeda, S., and Hiradate, S. (2008). Mechanism for the detoxification of aluminum in roots of tea plant (Camellia sinensis (L.) Kuntze). Phytochemistry 69, 147–153. doi: 10.1016/j.phytochem.2007.06.007
Moriyama, U., Tomioka, R., Kojima, M., Sakakibara, H., and Takenaka, C. (2016). Aluminum effect on starch, soluble sugar, and phytohormone in roots of Quercus serrata Thunb seedlings. Trees 30, 405–413. doi: 10.1007/s00468-015-1252-x
Mukhopadyay, M., Bantawa, P., Das, A., Sarkar, B., Bera, B., Ghosh, P., et al. (2012). Changes of growth, photosynthesis and alteration of leaf antioxidative defence system of tea [Camellia sinensis (L.) O. Kuntze] seedlings under aluminum stress. BioMetals 25, 1141–1154. doi: 10.1007/s10534-012-9576-0
Nagata, T., Hayatsu, M., and Kosuge, N. (1992). Identification of aluminium forms in tea leaves by 27Al NMR. Phytochemistry 31, 1215–1218. doi: 10.1016/0031-9422(92)80263-E
Negishi, T., Oshima, K., Hattori, M., Kanai, M., Mano, S., Nishimura, M., et al. (2012). Tonoplast- and plasma membrane localized aquaporin-family transporters in blue hydrangea sepals of aluminum hyperaccumulating plant. PLOS ONE 7:e43189. doi: 10.1371/journal.pone.0043189
Negishi, T., Oshima, K., Hattori, M., and Yoshida, K. (2013). Plasma membrane-localized Al-transporter from blue hydrangea sepals is a member of the anion permease family. Genes Cells 18, 341–352. doi: 10.1111/gtc.12041
Nguyen, N. T., Dudzinski, M., Mohapatra, P. K., and Fujita, K. (2005). Distribution of accumulated aluminum and changes in cell wall polysaccharides in Eucalyptus camaldulensis and Melaleuca cajuputi under aluminum stress. Soil Sci Plant Nutr. 51, 737–740. doi: 10.1111/j.1747-0765.2005.tb00103.x
Nian, H., Yang, Z., Huang, H., Yan, X., and Matsumoto, H. (2005). Citrate secretion induced by aluminum stress may not be a key mechanism responsible s for differential aluminum tolerance of some soybean genotypes. J. Plant Nutr. 27, 2047–2066. doi: 10.1081/PLN-200030112
Nogueirol, R. C., Monteiro, F. A., and Azevedo, R. A. (2015). Tropical soils cultivated with tomato: fractionation and speciation of Al. Environ. Monit. Assess. 187, 160. doi: 10.1007/s10661-015-4366-0
Nunes-Nesi, A., Brito, D. S., Inostroza-Blancheteau, C., Fernie, A. R., and Araújo, W. L. (2014). The complex role of mitocondrial metabolism in plant aluminum resistance. Trends Plant Sci. 19, 399–407. doi: 10.1016/j.tplants.2013.12.006
Oda, A. (2003). Physiological and Ecological Study About Effect of Acid Rain on Forest. Effect of Aluminum Concentration on Growth and Endogenous Growth Regulatory Substances of Woody Plant Seedlings (in Japanese). Doctoral dissertation, Tottori University, Tottori.
Osaki, M. T., Watanabe, T., and Tadano, T. (1997). Beneficial effect of aluminum on growth of plants adapted to low pH soils. Soil Sci. Plant Nutr. 43, 551–563. doi: 10.1080/00380768.1997.10414782
Osawa, H., Endo, I., Hara, Y., Matsushima, Y., and Tange, T. (2011). Transient proliferation of proanthocyanidin accumulating cells on the epidermal apex contributes to highly aluminum-resistant root elongation in camphor tree. Plant Physiol. 155, 433–446. doi: 10.1104/pp.110.166967
Ownby, J. D., and Popham, H. R. (1989). Citrate reverses the inhibition of wheat root growth caused by aluminum. J. Plant Physiol. 135, 588–591. doi: 10.1016/S0176-1617(11)80641-3
Oyama, K., Yamada, T., Ito, D., Kondo, T., and Yoshida, K. (2015). Metal complex pigment involved in the blue sepal color development of Hydrangea. J. Agric. Food Chem. 63, 7630–7635. doi: 10.1021/acs.jafc.5b02368
Panda, S. K., and Matsumoto, H. (2007). Molecular physiology of aluminum toxicity and tolerance in plants. Bot. Rev. 73, 326–347.
Panda, S. K., Sahoo, L., Katsuhara, M., and Matsumoto, H. (2013). Overexpression of alternative oxidase gene confers aluminum tolerance by altering the respiratory capacity and the response to oxidative stress in tobacco cells. Mol. Biotechnol. 54, 551–563. doi: 10.1007/s12033-012-9595-7
Pech-Ku, R., Muñoz-Sánchez, A., Monforte-González, M., Vázquez-Flota, F., Rodas-Junco, B. A., González-Mendoza, V. M., et al. (2017). Relationship between aluminum stress and caffeine biosynthesis in suspension cells of Coffea arabica L. J. Inorg. Biochem. doi: 10.1016/j.jinorgbio.2017.07.032 [Epub ahead of print],
Pilon-Smits, E. A. H., Quinn, C. F., Tapken, W., Malagoli, M., and Schiavon, M. (2009). Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 12, 267–274. doi: 10.1016/j.pbi.2009.04.009
Piñeros, M. A., Shaff, J. E., Manslank, H. S., Alves, V. M. C., and Kochian, L. V. (2005). Aluminum resistance in maize cannot be solely explained by root organic acid exudation. A comparative physiological study. Plant Physiol. 137, 231–241. doi: 10.1104/pp.104.047357
Poschenrieder, C., Gunsé, B., Corrales, I., and Barceló, J. (2008). A glance into aluminum toxicity and resistance in plants. Sci. Total Environ. 400, 356–368. doi: 10.1016/j.scitotenv.2008.06.003
Poschenrieder, C., Tolrà, R., Hajiboland, R., Arroyave, C., and Barceló, J. (2015). “Mechanisms of hyper-resistance and hyper-tolerance to aluminum in plants,” in Aluminum Stress Adaptation in Plants, eds S. Panda and F. Baluška (Cham: Springer International Publishing), 81–98.
Poschenrieder, C., Tolra, R. P., and Barcelo, J. (2005). A role for cyclic hydroxamates in aluminium resistance in maize? J. Inorg. Biochem. 99, 1830–1836. doi: 10.1016/j.jinorgbio.2005.05.017
Potter, D. A., Powell, A. J., Spicer, P. G., and Williams, D. W. (1996). Cultural practices affect root feeding white grubs (Coleoptera: Scarabaeidae) in turfgrass. J. Econ. Entomol. 89, 156–164. doi: 10.1093/jee/89.1.156
Qin, R., Hirano, Y., and Brunner, I. (2007). Exudation of organic acid anions from poplar roots after exposure to Al, Cu, and Zn. Tree Physiol. 27, 313–320. doi: 10.1093/treephys/27.2.313
Quarles, L. D., Gitelman, H. J., and Drezner, M. K. (1988). Induction of de novo bone formation in the beagle. A novel effect of aluminum. J. Clin. Invest. 81, 1056–1066. doi: 10.1172/JCI113417
Quarles, L. D., Gitelman, H. J., and Drezner, M. K. (1989). Aluminum-induced de novo bone formation in the beagle. A parathyroid hormone-dependent event. J. Clin. Invest. 83, 1644–1650. doi: 10.1172/JCI114063
Quarles, L. D., Hartle, J. E. I., Middleton, J. P., Zhang, J., Arthur, J. M., and Raymond, J. R. (1994). Aluminum-induced DNA synthesis in osteoblast: mediated by a G protein coupled cation sensing mechanism. J. Cell Biochem. 56, 106–117. doi: 10.1002/jcb.240560115
Rao, I. M., Miles, J. W., Beebe, S. E., and Horst, W. J. (2016). Root adaptations to soils with low fertility and aluminium toxicity. Ann. Bot. 118, 593–605. doi: 10.1093/aob/mcw073
Rehmus, A., Bigalke, M., Valarezo, C., Mora-Castillo, J., and Wilcke, W. (2014). Aluminum toxicity to tropical montane forest tree seedlings in southern Ecuador: response of biomass and plant morphology to elevated Al concentrations. Plant Soil 382, 301–315. doi: 10.1007/s11104-014-2110-0
Rengel, Z., Bose, J., Chen, Q., and Tripathi, B. N. (2015). Magnesium alleviates plant toxicity of aluminium and heavy metals. Crop Pasture Sci. 66, 1298–1307. doi: 10.1071/CP15284
Rounds, M. A., and Larsen, P. B. (2008). Aluminum-dependent root-growth inhibition in Arabidopsis results from AtATR-regulated cell-cycle arrest. Curr. Biol. 18, 1495–1500. doi: 10.1016/j.cub.2008.08.050
Rufty, T. W., Mackown, C. T., Lazof, D. B., and Carter, T. E. (1995). Effects of aluminium on nitrate uptake and assimilation. Plant Cell Environ. 18, 1325–1331. doi: 10.1111/j.1365-3040.1995.tb00192.x
Ryan, P. R., Tyerman, S. D., Sasaki, T., Furuichi, T., Yamamoto, Y., Zhang, W. H., et al. (2011). The identification of aluminium-resistance genes provides opportunities for enhancing crop production on acid soils. J. Exp. Bot. 62, 9–20. doi: 10.1093/jxb/erq272
Saçan, M. T., Oztay, F., and Bolkent, S. (2007). Exposure of Dunaliella tertiolecta to lead and aluminum: toxicity and effects on ultrastructure. Biol. Trace Elem. Res. 120, 264–272. doi: 10.1007/s12011-007-8016-4
Sade, H., Meriga, B., Surapu, V., Gadi, J., Sunita, M. S. L., Suravajhala, P., et al. (2016). Toxicity and tolerance of aluminium in plants: tailoring plants to suit to acid soils. Biometals 29, 187–210. doi: 10.1007/s10534-016-9910-z
Sasaki, T., Yamamoto, Y., Ezaki, B., Katsuhara, M., Ahn, S. J., Ryan, P. R., et al. (2004). A wheat gene encoding an aluminum-activated malate transporter. Plant J. 37, 645–653. doi: 10.1111/j.1365-313X.2003.01991.x
Schmitt, M., Boras, S., Tjoa, A., Watanabe, T., and Jansen, S. (2016a). Aluminium accumulation and intra-tree distribution patterns in three arbor aluminosa (Symplocos) species from Central Sulawesi. PLOS ONE 11:e0149078. doi: 10.1371/journal.pone.0149078
Schmitt, M., Watanabe, T., and Jansen, S. (2016b). The effects of aluminium on plant growth in a temperate and deciduous aluminium accumulating species. AOB Plants 8:plw065. doi: 10.1093/aobpla/plw065
Schreiber, H. D., Jones, A. H., Lariviere, C. M., Mayhew, K. M., and Cain, J. B. (2011). Role of aluminum in red-to-blue color changes in Hydrangea macrophylla sepals. Biometals 24, 1005–1015. doi: 10.1007/s10534-011-9458-x
Schroeder, J. I., Delhaize, E., Frommer, W. B., Guerinot, M. L., Harrison, M. J., Herrera-Estrella, L., et al. (2013). Using membrane transporters to improve crops for sustainable food production. Nature 497, 60–66. doi: 10.1038/nature11909
Seguel, A., Cumming, J. R., Klugh-Stewart, K., Cornejo, P., and Borie, F. (2013). The role of arbuscular mycorrhizas in decreasing aluminium phytoxicity in acidic soils: a review. Mycorrhiza 23, 167–183. doi: 10.1007/s00572-013-0479-x
Shao, J. F., Che, J., Chen, R. F., Ma, J. F., and Shen, R. F. (2015). Effect of in planta phosphorus on aluminum-induced inhibition of root elongation in wheat. Plant Soil 395, 307–315. doi: 10.1007/s11104-015-2566-6
Shen, R., Ma, J., Kyo, M., and Iwashita, T. (2002). Compartmentation of aluminium in leaves of an Al-accumulator, Fagopyrum esculentum Moench. Planta 215, 394–398. doi: 10.1007/s00425-002-0763-z
Shen, X., Xiao, X., Dong, Z., and Chen, Y. (2014). Silicon effects on antioxidative enzymes and lipid peroxidation in leaves and roots of peanut under aluminum stress. Acta Physiol. Plant. 36, 3063–3069. doi: 10.1007/s11738-014-1676-8
Silva, G. E. A., Ramos, F. T., de Faria, A. P., and França, M. G. C. (2014). Seeds’ physicochemical traits and mucilage protection against aluminum effect during germination and root elongation as important factors in a biofuel seed crop (Ricinus communis). Environ. Sci. Pollut. Res. 21, 11572–11579. doi: 10.1007/s11356-014-3147-6
Simoes, C. C., Melo, J. O., Magalhaes, J. V., and Guimaraes, C. T. (2012). Genetic and molecular mechanisms of aluminum tolerance in plants. Genet. Mol. Res. 11, 1949–1957. doi: 10.4238/2012.July.19.14
Singh, V. P., Tripathi, D. K., Kumar, D., and Chauhan, D. K. (2011). Influence of exogenous silicon addition on aluminium tolerance in rice seedlings. Biol. Trace Elem. Res. 144, 1260–1274. doi: 10.1007/s12011-011-9118-6
Stoklasa, J. (1922). Über die Verbreitung des Aluminiums in der Natur und Seine Bedeutung Beim bau- und Betriebsstoffwechsel der Pflanzen. Jena: Fisher Scientific.
Tahara, K., Hashida, K., Ohara, S., Kojima, K., and Shinohara, K. (2009). “Aluminum detoxifying compounds in roots of Eucalyptus camaldulensis,” in Proceedings of the International Plant Nutrition Colloquium XVI, (Davis, CA: University of California).
Tahara, K., Norisada, M., Yamanoshita, T., and Kojima, K. (2008). Role of binding ligands in aluminum resistance of Eucalyptus camaldulensis and Melaleuca cajuputi. Plant Soil 302, 175–187. doi: 10.1007/s11104-0079464-5
Tamura, S., and Yoshimura, E. (2008). Promotion of Zn2+ uptake by Al3+ in a Saccharomyces cerevisiae mutant that lacks the ZRT1 gene encoding a high-affinity Zn transporter. Biol. Trace Elem. Res. 124, 262–268. doi: 10.1007/s12011-008-8145-4
Tanikawa, N., Inoue, H., and Nakayama, M. (2016). Aluminum ions are involved in purple flower coloration in Camellia japonica ‘Sennen-fujimurasaki’. Hortic. J. 85, 331–339. doi: 10.2503/hortj.MI-114
Tanikawa, N., Kashiwabara, T., Hokura, A., Abe, T., Shibata, M., and Nakayama, M. (2008). A peculiar yellow flower coloration of camellia using aluminum-flavonoid interaction. J. Jpn. Soc. Hortic. Sci. 77, 402–407. doi: 10.2503/jjshs1.77.402
Taylor, G. J. (1988). The physiology of aluminum tolerance in higher plants. Commun. Soil Sci. Plant Anal. 19, 1179–1194. doi: 10.1080/00103628809368004
Thornton, F. C., Schaedle, M., and Ryanal, D. J. (1986). Effect of aluminum on the growth of sugar maple in solution culture. Can. J. For. Res. 16, 892–896. doi: 10.1139/x86-159
Thornton, F. C., Schaedle, M., and Ryanal, D. J. (1989). Tolerance of red oak and American and European beech seedlings to aluminum. J. Environ. Qual. 18, 541–545. doi: 10.2134/jeq1989.00472425001800040027x
Tokizawa, M., Kobayashi, Y., Saito, T., Kobayashi, M., Iuchi, S., Nomoto, M., et al. (2015). Sensitive to proton rhizotoxicity1, calmodulin binding transcription activator2, and other transcription factors are involved in aluminum-activated malate transporter1 expression. Plant Physiol. 167, 991–1003. doi: 10.1104/pp.114.256552
Tolrà, R., Vogel-Mikus, K., Hajiboland, R., Kump, P., Pongrac, P., Kaulich, B., et al. (2011). Localization of aluminium in tea (Camellia sinensis (L.) O. Kuntze) leaves using low energy X-ray fluorescence spectro-microscopy. J. Plant Res. 124, 165–172. doi: 10.1007/s10265-010-0344-3
Tomioka, R., Oda, A., and Takenaka, C. (2005). Root growth enhancement by rhizospheric aluminum treatment in Quercus serrata Thunb. seedlings. J. For. Res. 10, 319–324. doi: 10.1007/s10310-005-0152-0
Tomioka, R., and Takenaka, C. (2007). Enhancement of root respiration and photosynthesis in Quercus serrata Thunb. seedlings by long-term aluminum treatment. Environ. Sci. 14, 141–148.
Tomioka, R., Takenaka, C., Maeshima, M., Tezuka, T., Kojima, M., and Sakakibara, H. (2012). Stimulation of root growth induced by aluminum in Quercus serrata Thunb. Is related to activity of nitrate reductase and maintenance of IAA concentration in roots. Am. J. Plant Sci. 3, 1619–1624. doi: 10.4236/ajps.2012.311196
Tomioka, R., Uchida, A., Takenaka, C., and Tezuka, T. (2007). Effect of aluminum on nitrate reductase and photosynthetic activities in Quercus serrata seedlings. Environ Sci. 14, 157–165. doi: 10.4236/ajps.2012.311196
Trejo-Téllez, L., Stenzel, R., Gomez-Merino, F., and Schmitt, J. (2010). Transgenic tobacco plants overexpressing pyruvate phosphate dikinase increase exudation of organic acids and decrease accumulation of aluminum in the roots. Plant Soil 326, 187–198. doi: 10.1007/s11104-009-9994-0
Tsuji, M., Kuboi, T., and Konishi, S. (1994). Stimulatory effects of aluminum on the growth of cultured roots of tea. Soil Sci. Plant Nutr. 40, 471–476. doi: 10.1080/00380768.1994.10413324
Veitch, F. P. (1904). Comparison of methods for the estimation of soil acidity. J. Am. Chem. Soc. 26, 637–662. doi: 10.1021/ja01996a005
Wagatsuma, T., Khan, M. S., Watanabe, T., Maejima, E., Sekimoto, H., Yokota, T., et al. (2015). Higher sterol content regulated by CYP51 with concomitant lower phospholipid contents in membranes is a common strategy for aluminium tolerance in several plant species. J. Exp. Bot. 66, 907–918. doi: 10.1093/jxb/eru455
Wan, Q., Xu, R.-K., and Li, X.-H. (2012). Proton release from tea plant (Camellia sinensis L.) roots induced by Al(III) under hydroponic conditions. Soil Res. 50, 482–488.
Wang, H., Chen, R. F., Iwashita, T., Shen, R. F., and Ma, J. F. (2015a). Physiological characterization of aluminum tolerance and accumulation in tartary and wild buckwheat. New Phytol. 205, 273–279. doi: 10.1111/nph.13011
Wang, L., Fan, X.-W., Pan, J.-L., Huang, Z.-B., and Li, Y.-Z. (2015b). Physiological characterization of maize tolerance to low dose of aluminum, highlighted by promoted leaf growth. Planta 242, 1391–1403. doi: 10.1007/s00425-015-2376-3
Wang, L.-Q., Yang, L.-T., Guo, P., Zhou, X.-X., Ye, X., Chen, E.-J., et al. (2015c). Leaf cDNA-AFLP analysis reveals novel mechanisms for boron-induced alleviation of aluminum-toxicity in Citrus grandis seedlings. Ecotoxicol. Environ. Saf. 120, 349–359. doi: 10.1016/j.ecoenv.2015.06.009
Wang, W., Zhao, X. Q., Chen, R. F., Dong, X. Y., Lan, P., Ma, J. F., et al. (2015d). Altered cell wall properties are responsible for ammonium-reduced aluminium accumulation in rice roots. Plant Cell Environ. 38, 1382–1390. doi: 10.1111/pce.12490
Wang, W., Zhao, X. Q., Hu, Z. M., Shao, J. F., Che, J., Chen, R. F., et al. (2015e). Aluminium alleviates manganese toxicity to rice by decreasing root symplastic Mn uptake and reducing availability to shoots of Mn stored in roots. Ann. Bot. 116, 237–246. doi: 10.1093/aob/mcv090
Wang, H., Huang, J., Liang, W., Liang, X., and Bi, Y. (2013a). Involvement of putrescine and nitric oxide in aluminum tolerance by modulating citrate secretion from roots of red kidney bean. Plant Soil 366, 479–490. doi: 10.1007/s11104-012-1447-5
Wang, H., Liang, W., and Huang, J. (2013b). Putrescine mediates aluminum tolerance in red kidney bean by modulating aluminum-induced oxidative stress. Crop Sci. 53, 2120–2128. doi: 10.2135/cropsci2013.04.0215
Wang, Q., Nian, F., Zhao, L., Li, F., Yang, H., and Yang, Y. (2013c). Exogenous indole-3-acetic acid could reduce the accumulation of aluminum in root apex of wheat (Triticum aestivum L.) under Al stress. J. Soil Plant Nutr. 13, 534–543. doi: 10.4067/S0718-95162013005000042
Wang, Q.-F., Zhao, Y., Yi, Q., Li, K.-Z., Yu, Y.-X., and Chen, L.-M. (2010). Overexpression of malate dehydrogenase in transgenic tobacco leaves: enhanced malate synthesis and augmented Al-resistance. Acta Physiol. Plant. 32, 1209–1220. doi: 10.1007/s11738-010-0522-x
Wang, S., Ren, X., Huang, B., Wang, G., Zhou, P., and An, Y. (2016). Aluminium-induced reduction of plant growth in alfalfa (Medicago sativa) is mediated by interrupting auxin transport and accumulation in roots. Sci. Rep. 6:30079. doi: 10.1038/srep30079
Wang, P., Yu, W., Zhang, J., Rengel, Z., Xu, J., Han, Q., et al. (2016). Auxin enhances aluminium-induced citrate exudation through upregulation of GmMATE and activation of the plasma membrane H+-ATPase in soybean roots. Ann. Bot. 118, 933–940. doi: 10.1093/aob/mcw133
Wang, Y., Li, R., Li, D., Jia, X., Zhou, D., Li, J., et al. (2017). NIP1;2 is a plasma membrane-localized transporter mediating aluminum uptake, translocation, and tolerance in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 114, 5047–5052. doi: 10.1073/pnas.1618557114
Wang, Y., Stass, A., and Horts, W. J. (2004). Apoplastic binding of aluminum is involved in silicon-induced amelioration of aluminum toxicity in maize. Plant Physiol. 136, 3762–3770. doi: 10.1104/pp.104.045005
Watanabe, T., Jansen, S., and Osaki, M. (2005). The beneficial effect of aluminium and the role of citrate in Al accumulation in Melastoma malabathricum. New Phytol. 165, 773–780. doi: 10.1111/j.1469-8137.2004.01261.x
Watanabe, T., Jansen, S., and Osaki, M. (2006). Al-Fe interactions and growth enhancement in Melastoma malabathricum and Miscanthus sinensis dominating acid sulphate soils. Plant Cell Environ. 29, 2124–2132. doi: 10.1111/j1365-3040.2006.001586.x
Watanabe, T., Misawa, S., Hiradate, S., and Osaki, M. (2008a). Characterization of root mucilage from Melastoma malabathricum, with emphasis on its roles in aluminium accumulation. New Phytol. 178, 581–589. doi: 10.1111/j.1469-8137.2008.02397.x
Watanabe, T., Misawa, S., Hiradate, S., and Osaki, M. (2008b). Root mucilage enhances aluminum accumulation in Melastoma malabathricum, an aluminum accumulator. Plant Signal. Behav. 3, 603–605. doi: 10.4161/psb.3.8.6356
Watanabe, T., and Osaki, M. (2001). Influence of aluminum and phosphorus on growth and xylem sap composition in Melastoma malabathricum L. Plant Soil 237, 63–70. doi: 10.1023/A:1013395814958
Watanabe, T., and Osaki, M. (2009). “Possible reasons why aluminum is a beneficial element for Melastoma malabathricum, an aluminum accumulator,” in Proceedings of the International Plant Nutrition Colloquium XVI, (Davis, CA: University of California).
Watanabe, T., Osaki, M., and Tadano, T. (1997). Aluminum-induced growth stimulation in relation to calcium, magnesium, and silicate nutrition in Melastoma malabathricum L. Soil Sci. Plant Nutr. 43, 827–837. doi: 10.1080/00380768.1997.10414649
Winkel-Shirley, B. (2002). Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 5, 218. doi: 10.1016/S1369-5266(02)00256-X
Wu, D., Shen, H., Yokawa, K., and Baluška, F. (2014). Alleviation of aluminium-induced cell rigidity by overexpression of OsPIN2 in rice roots. J. Exp. Bot. 65, 5305–5315. doi: 10.1093/jxb/eru292
Wu, D., Shen, H., Yokawa, K., and Baluška, F. (2015). Overexpressing OsPIN2 enhances aluminium internalization by elevating vesicular trafficking in rice root apex. J. Exp. Bot. 66, 6791–6801. doi: 10.1093/jxb/erv385
Xia, J., Yamaji, N., Che, J., Shen, R. F., and Ma, J. F. (2014). Differential expression of Nrat1 is responsible for Al-tolerance QTL on chromosome 2 in rice. J. Exp. Bot. 65, 4297–4304. doi: 10.1093/jxb/eru201
Xia, J., Yamaji, N., Kasai, T., and Ma, J. F. (2010). Plasma membrane-localized transporter for aluminum in rice. Proc. Natl. Acad. Sci. U.S.A. 107, 18381–18385. doi: 10.1073/pnas.1004949107
Xia, J., Yamaji, N., and Ma, J. F. (2011). Further characterization of an aluminum influx transporter in rice. Plant Signal. Behav. 6, 160–163. doi: 10.4161/psb.6.1.14319
Xia, J., Yamaji, N., and Ma, J. F. (2013). A plasma membrane-localized small peptide is involved in rice aluminum tolerance. Plant J. 76, 345–355. doi: 10.1111/tpj.12296
Xu, Q., Wang, Y., Ding, Z., Song, L., Li, Y., Ma, D., et al. (2016). Aluminum induced metabolic responses in two tea cultivars. Plant Physiol. Biochem. 101, 162–172. doi: 10.1016/j.plaphy.2016.02.001
Yamaji, N., Huang, C. F., Nagao, S., Yano, M., Sato, Y., Nagamura, Y., et al. (2009). A zinc finger transcription factor ART1 regulates multiple genes implicated in aluminum tolerance in rice. Plant Cell 21, 3339–3349. doi: 10.1105/tpc.109.070771
Yang, J., Qu, M., Fang, J., Shen, R. F., Feng, Y. M., Liu, J. Y., et al. (2016). Alkali-soluble pectin is the primary target of aluminum immobilization in root border cells of pea (Pisum sativum). Front. Plant Sci 7:1297. doi: 10.3389/fpls.2016.01297
Yang, J. L., Chen, W. W., Chen, L. Q., Qin, C., Jin, C. W., Shi, Y. Z., et al. (2013). The 14-3-3 protein GENERAL REGULATORY FACTOR11 (GRF11) acts downstream of nitric oxide to regulate iron acquisition in Arabidopsis thaliana. New Phytol. 197, 815–824. doi: 10.1111/nph.12057
Yang, J. L., Li, Y. Y., Zhang, Y. J., Zhang, S. S., Wu, Y. R., Wu, P., et al. (2008). Cell wall polysaccharides are specifically involved in the exclusion of aluminum from the rice root apex. Plant Physiol. 146, 602–611. doi: 10.1104/pp.107.111989
Yang, J. L., Zhu, X. F., Peng, Y. X., Zheng, C., Li, G. X., Liu, Y., et al. (2011). Cell wall hemicellulose contributes significantly to aluminum adsorption and root growth in Arabidopsis. Plant Physiol. 155, 1885–1892. doi: 10.1104/pp.111.172221
Yang, X. Y., Yang, J. L., Zhou, Y., Piñeros, M. A., Kochian, L. V., Li, G. X., et al. (2011). A de novo synthesis citrate transporter, Vigna umbellata multidrug and toxic compound extrusion, implicated in Al activated citrate efflux in rice bean (Vigna umbellata) root apex. Plant Cell Environ. 34, 2138–2148. doi: 10.1111/j.1365-3040.2011.02410.x
Yang, Y., Liu, Y., Huang, C.-F., de Silva, J., and Zhao, F.-J. (2016). Aluminium alleviates fluoride toxicity in tea (Camellia sinensis). Plant Soil 402, 179–190. doi: 10.1007/s11104-015-2787-8
Yang, Y., Wang, Q. L., Geng, M. L., Guo, Z. H., and Zhao, Z. (2011). Rhizosphere pH difference regulated by plasma membrane H+-ATPase is related do differential Al-tolerance of two wheat cultivars. Plant Soil Environ. 57, 201–206.
Yang, Z. B., You, J. F., Xu, M. Y., and Yang, Z. M. (2009). Interaction between aluminum toxicity and manganese toxicity in soybean (Glycine max). Plant Soil 319, 277–289. doi: 10.1007/s11104-008-9869-9
Yokosho, K., Yamaji, N., and Ma, J. F. (2011). An Al-inducible MATE gene is involved in external detoxification of Al in rice. Plant J. 68, 1061–1069. doi: 10.1111/j.1365-313X.2011.04757.x
Yoshii, Y. (1937). Aluminium requirements of solfatara plants. Bot. Mag. 51, 262–270. doi: 10.15281/jplantres1887.51.262
Yu, H. N., Liu, P., Wang, Z. Y., Chen, W. R., and Xu, G. D. (2011). The effect of aluminum treatments on the root growth and cell ultrastructure of two soybean genotypes. Crop Prot. 30, 323–328. doi: 10.1016/j.cropro.2010.11.024
Yu, M., Shen, R., Liu, J., Chen, R., Xu, M., Yong, Y., et al. (2009a). The role of root border cells in aluminum resistance of pea (Pisum sativum) grown in mist culture. J. Plant Nutr. Soil Sci. 172, 528–534. doi: 10.1002/jpln.200800039
Yu, M., Shen, R., Xiao, H., Xu, M., Wang, H., Wang, H., et al. (2009b). Boron alleviates aluminum toxicity in pea (Pisum sativum). Plant Soil 314, 87–98. doi: 10.1007/s11104-008-9708-z
Yu, W., Kan, Q., Zhang, J., Zeng, B., and Chen, Q. (2016). Role of the plasma membrane H+-ATPase in the regulation of organic acid exudation under aluminum toxicity and phosphorus deficiency. Plant Signal. Behav. 11:e1106660. doi: 10.1080/15592324.2015.1106660
Yu, Y., Jin, C., Sun, C., Wang, J., Ye, Y., Zhou, W., et al. (2016). Inhibition of ethylene production by putrescine alleviates aluminium-induced root inhibition in wheat plants. Sci. Rep. 6:18888. doi: 10.1038/srep18888
Zeng, F., Chen, S., Miao, Y., Wu, F., and Zhang, G. (2008). Changes of organic acid exudation and rhizosphere pH in rice plants under chromium stress. Environ. Pollut. 155, 284–289. doi: 10.1016/j.envpol.2007.11.019
Zeng, H., Feng, X., Wang, B., Zhu, Y., Shen, Q., and Xu, G. (2013). Citrate exudation induced by aluminum is independent of plasma membrane H+-ATPase activity and coupled with potassium efflux from cluster roots of phosphorus-deficient white lupin. Plant Soil 366, 389–400. doi: 10.1007/s1104-012-1445-7
Zeng, Q. L., Chen, R. F., Zhao, X. Q., Shen, R. F., Noguchi, A., Shinmachi, F., et al. (2013). Aluminum could be transported via phloem in Camellia oleifera Abel. Tree Physiol. 33, 96–105. doi: 10.1093/treephys/tps117
Zhang, B., Liu, P., Yang, Y. S., and Xu, G. D. (2007). Effect of Al in soil on photosynthesis and related morphological and physiological characteristics of two soybean genotypes. Bot. Stud. 48, 435–444.
Zhang, H., Jiang, Z., Qin, R., Zhang, H., Zou, J., Jiang, W., et al. (2014). Accumulation and cellular toxicity of aluminum in seedling of Pinus massoniana. BMC Plant Biol. 14:264. doi: 10.1186/s12870-014-0264-9
Zhang, M., Ma, Y., Horst, W. J., and Yang, Z.-B. (2016). Spatial-temporal analysis of polyethylene glycol-reduced aluminium accumulation and xyloglucan endotransglucosylase action in root tips of common bean (Phaseolus vulgaris). Ann. Bot. 118, 1–9. doi: 10.1093/aob/mcw062
Zhang, W. H., Ryan, P. R., Sasaki, T., Yamamoto, Y., Sullivan, W., and Tyerman, S. D. (2008). Characterization of the TaALMT1 protein as an Al3+-activated anion channel in transformed tobacco (Nicotiana tabacum L.) cells. Plant Cell Physiol. 49, 1316–1330. doi: 10.1093/pcp/pcn107
Zheng, S. J., Yang, J. L., He, Y. F., Yu, X. H., Zhang, L., You, J. F., et al. (2005). Immobilization of aluminum with phosphorus in roots is associated with high aluminum resistance in buckwheat. Plant Physiol. 138, 297–303. doi: 10.1104/pp.105.059667
Zhou, G., Delhaize, E., Zhou, M., and Ryan, P. R. (2011). “Biotechnological solutions for enhancing the aluminium resistance of crop plants,” in Abiotic Stress in Plants-Mechanisms and Adaptations, eds A. Shanker and B. Venkateswarlu (Rijeka: InTech), doi: 10.5772/25187
Zhou, L., Tan, Y., Huang, L., and Wang, W.-X. (2016). Enhanced utilization of organic phosphorus in a marine diatom Thalassiosira weissflogii: a possible mechanism for aluminum effect under P limitation. J. Exp. Mar. Biol. Ecol. 478, 77–85. doi: 10.1016/j.jembe.2016.02.009
Zhou, S., Okekeogbu, I., Sangireddy, S., Ye, Z., Li, H., Bhatti, S., et al. (2016). Proteome modification in tomato plants upon long-term aluminum treatment. J. Proteome Res. 15, 1670–1684. doi: 10.1021/acs.jproteome.6b00128
Zhou, X.-X., Yang, L.-T., Qi, Y.-P., Guo, P., and Chen, L.-S. (2015). Mechanisms on boron-induced alleviation of aluminum-toxicity in Citrus grandis seedlings at a transcriptional level revealed by cDNA-AFLP analysis. PLOS ONE 10:e0115485. doi: 10.1371/journal.pone.0115485
Zhu, M. Y., Ahn, S., and Matsumoto, H. (2003). Inhibition of growth and development of root border cells in wheat by Al. Physiol. Plant. 117, 359–367. doi: 10.1034/j.1399-3054.2003.00036.x
Zhu, X. F., Wan, J. X., Sun, Y., Shi, Y. Z., Braam, J., Li, G. X., et al. (2014). Xyloglucan endotransglucosylase-hydrolase17 interacts with xyloglucan endotransglucosylase-hydrolase31 to confer xyloglucan endotransglucosylase action and affect aluminum sensitivity in Arabidopsis. Plant Physiol. 165, 1566–1574. doi: 10.1104/pp.114.243790
Keywords: acid soils, aluminum, aluminum toxicity, beneficial effect of aluminum, mechanisms of tolerance, metal, plant growth stimulation
Citation: Bojórquez-Quintal E, Escalante-Magaña C, Echevarría-Machado I and Martínez-Estévez M (2017) Aluminum, a Friend or Foe of Higher Plants in Acid Soils. Front. Plant Sci. 8:1767. doi: 10.3389/fpls.2017.01767
Received: 30 April 2017; Accepted: 27 September 2017;
Published: 12 October 2017.
Edited by:
Rudra Deo Tripathi, National Botanical Research Institute (CSIR), IndiaReviewed by:
Frantisek Baluska, University of Bonn, GermanyFernando Carlos Gómez-Merino, Colegio de Postgraduados, Mexico
Copyright © 2017 Bojórquez-Quintal, Escalante-Magaña, Echevarría-Machado and Martínez-Estévez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuel Martínez-Estévez, luismanh@cicy.mx