 Jiming Liu1,2,3†
Jiming Liu1,2,3† Shilun Gao1,2†Yuanyuan Xu1,2Mianzhi Wang1,2
Shilun Gao1,2†Yuanyuan Xu1,2Mianzhi Wang1,2 Jia Jun Ngiam3
Jia Jun Ngiam3 Nicholas Cho Rui Wen3Joan Jong Jing Yi3Xuehuang Weng4
Nicholas Cho Rui Wen3Joan Jong Jing Yi3Xuehuang Weng4 Liming Jia1,2*
Liming Jia1,2* Jarkko Salojärvi3,5*
Jarkko Salojärvi3,5*- 1Key Laboratory for Silviculture and Conservation of the Ministry of Education, Beijing Forestry University, Beijing, China
- 2National Energy R&D Center for Non-Food Biamass, Beijing Forestry University, Beijing, China
- 3School of Biological Sciences, Nanyang Technological University, Singapore, Singapore
- 4Yuanhua Forestry Biological Technology Co., Ltd., Sanming, China
- 5Organismal and Evolutionary Biology Research Program, Faculty of Biological and Environmental Sciences, The Viikki Plant Science Centre, University of Helsinki, Helsinki, Finland
Sapindus is an important forest tree genus with utilization in biodiesel, biomedicine, and it harbors great potential for biochemical engineering applications. For advanced breeding of Sapindus, it is necessary to evaluate the genetic diversity and construct a rationally designed core germplasm collection. In this study, the genetic diversity and population structure of Sapindus were conducted with 18 expressed sequence tag-simple sequence repeat (EST-SSR) markers in order to establish a core germplasm collection from 161 Sapindus accessions. The population of Sapindus showed high genetic diversity and significant population structure. Interspecific genetic variation was significantly higher than intraspecific variation in the Sapindus mukorossi, Sapindus delavayi, and combined Sapindus rarak plus Sapindus rarak var. velutinus populations. S. mukorossi had abundant genetic variation and showed a specific pattern of geographical variation, whereas S. delavayi, S. rarak, and S. rarak var. velutinus showed less intraspecific variation. A core germplasm collection was created that contained 40% of genetic variation in the initial population, comprising 53 S. mukorossi and nine S. delavayi lineages, as well as single representatives of S. rarak and S. rarak var. velutinus. These results provide a germplasm basis and theoretical rationale for the efficient management, conservation, and utilization of Sapindus, as well as genetic resources for joint genomics research in the future.
Highlights
- Sapindus is biodiesel, biomedical, and multifunctional economic forest species.
- Interspecific genetic variation was significantly higher than intraspecific variation in the Sapindus populations.
- Sapindus mukorossi showed a specific pattern of geographical variation, whereas Sapindus delavayi, Sapindus rarak, and Sapindus rarak var. velutinus showed less intraspecific genetic variation.
- A core germplasm collection was created that contained 40% of the initial population; it comprised 53 individuals of Sapindus mukorossi, nine of Sapindus delavayi, one of Sapindus rarak, and one of Sapindus rarak var. velutinus.
Introduction
Sapindus is a widely distributed economic forest genus of Sapindaceae family; it is typically scattered as single plant or extremely small populations in temperate to tropical regions, with main biodiversity in Southeast Asia and America (Liu et al., 2017). Among Sapindus, Sapindus mukorossi Gaertn. (S. mukorossi), Sapindus delavayi (Franch.) Radlk. (S. delavayi), Sapindus rarak DC. (S. rarak), and Sapindus rarak var. velutinus (S. rarak var.) are concentrated in east and southeast Asia (Liu et al., 2017). Seed oils of Sapindus are suitable for the preparation of biodiesel under both American and European standards (D6751 and EN 14214, respectively) (Chakraborty and Baruah, 2013; Pelegrini et al., 2017; Caowen et al., 2019), owing to the high oil content (26.69–44.69%) and unsaturated fatty acid (mean: 86.21%) (Sun et al., 2017; Liu et al., 2021a). The pericarp of Sapindus also contains abundant triterpene saponins (4.14–27.04%) and sesquiterpenes (Xu et al., 2018; Liu et al., 2019). More than 70 of these triterpenoid saponin compounds have been identified (Xu et al., 2018) and shown to exhibit outstanding surface activity, antibacterial (Basu et al., 2015), elution (Mukhopadhyay et al., 2013; Mukherjee et al., 2015; Mukhopadhyay et al., 2016), pharmacological (Rodriguez-Hernández et al., 2015), and physiological properties (Singh and Singh, 2008). Saponins from the pericarp of Sapinuds are widely used in commercial soaps, shampoos, and body washes (Muntaha and Khan, 2015), seeds oils are utilized in biodiesel and premium lubricants, and seedlings are commonly used for landscaping in southern China. Sapindus is consequently recognized as a sustainable biodiesel, biomedical, biochemical, and multifunctional economic forestry species in China (Sun et al., 2016; Liu et al., 2017; Liu et al., 2021a) with annual production values exceeding 100M USD. However, with worldwide deforestation and the rapidly anthropogenic expanding, the habitat and populations of Sapindus have been severely damaged or vanished in recent centuries, and the genetic diversity of Sapindus faces unprecedented threats (Liu et al., 2017; Liu et al., 2021b). Hence, breeders have recently carried out several surveys and collections of Sapindus germplasm resources, and over 1,000 samples have been collected (Liu et al., 2017). However, due to inconsistencies in the timing, standards, and designation of germplasm collections, there is considerable homonymy, synonymy, and genetic redundancy within the resources. Therefore, a comprehensive evaluation of the genetic diversity in Sapindus and the construction of a rationally designed core germplasm collection are needed.
Germplasm resources form the foundation of forest genetic breeding, and the development of forest tree breeding and industry depends largely on the extent and diversity captured by these resources. However, redundancy in germplasm resources may lead to lower conservation and management efficiency. The construction of core germplasm collection is the optimal solution to genetic redundancy. Core germplasm collection is a subset of germplasm accessions that represents the minimum repeatability and maximum genetic diversity of one species (Frankel, 1984; Brown, 1989; Lv et al., 2020). They have been widely used for germplasm management, conservation, and application in crop, flower, and horticultural tree species. Most core germplasm collections represent only 5–20% of the total germplasm collected (Hintum et al., 2000; Lv et al., 2020), thereby reducing conservation and management costs and improving the efficiency of germplasm utilization. However, woody plant germplasm is predominantly derived from natural populations with brief history of domestication and long generation time, therefore the accessions have a high intrinsic genetic diversity and core germplasm collections typically represent 10–45% of the complete germplasm collections within these species (Belaj et al., 2012; Duan et al., 2017; Min et al., 2017; Preethi et al., 2020).
Molecular markers are one of the most powerful and inexpensive tools for analyzing genetic diversity and establishing core germplasm collections, compared to whole genome sequencing, resequencing, or transcriptome sequencing approaches. Microsatellite markers, also referred to as the simple sequence repeats (SSR) markers, have been widely applied in genetic breeding, variety identification, germplasm diversity evaluation and conservation (Powell et al., 1996). EST-SSR (expressed sequence tags microsatellite markers) markers not only have the beneficial characteristics of high intraspecific polymorphism, co-dominant nature, and high reproducibility, but also originate from genomic coding regions and thus directly reflect the diversity of the underlying genes (Adams et al., 1991; Wang et al., 2017; Parthiban et al., 2018). EST-SSRs have been commonly used to evaluate genetic diversity of Dendrobium officinale (Xie et al., 2020), Paeonia rockii (Guo et al., 2020), coconut (Preethi et al., 2020), and Stevia rebaudiana (Cosson et al., 2019) and to construct core germplasm collections of Rosa roxburghii (Min et al., 2017), crape myrtle (Ye et al., 2017), and olive (Dervishi et al., 2021). Previous studies have applied ISSR (inter-simple sequence repeat) and RAPD (random amplified polymorphic DNA) molecular markers to evaluate the genetic diversity of the S. mukorossi population (Mahar et al., 2011b; Diao et al., 2016), however, there have been no studies or reports on the construction of Sapindus core germplasm collection.
In this study, 18 EST-SSR markers were selected based on whole transcriptome sequencing and used to evaluate the genetic diversity and population structure of 161 Sapindus individuals. The aim was to obtain a representative core germplasm collection which would retain maximum amount of genetic diversity and population structure of the sampled Sapindus population. The core germplasm will enable more scientific and rational conservation, management, and utilization of the genetic resources in Sapindus.
Materials and Methods
Expressed Sequence Tag-Simple Sequence Repeat Marker Retrieval and Primer Design
Expressed sequence tag-simple sequence repeats were identified in transcript sequences from the Sapindus whole transcriptome sequencing project using MISA1 (Thiel et al., 2003), and Primer3 (Untergasser et al., 2012) was used to design EST-SSR primers. We screened all EST-SSR markers against eight Sapindus germplasm accessions representing different geographic origins and selected 18 pairs of highly polymorphic and stable EST-SSR markers (Table 1) (unpublished). The 18 EST-SSR primers were synthesized by Beijing Ruiboxingke Biotechnology Co. Ltd. and used in subsequent experiments.
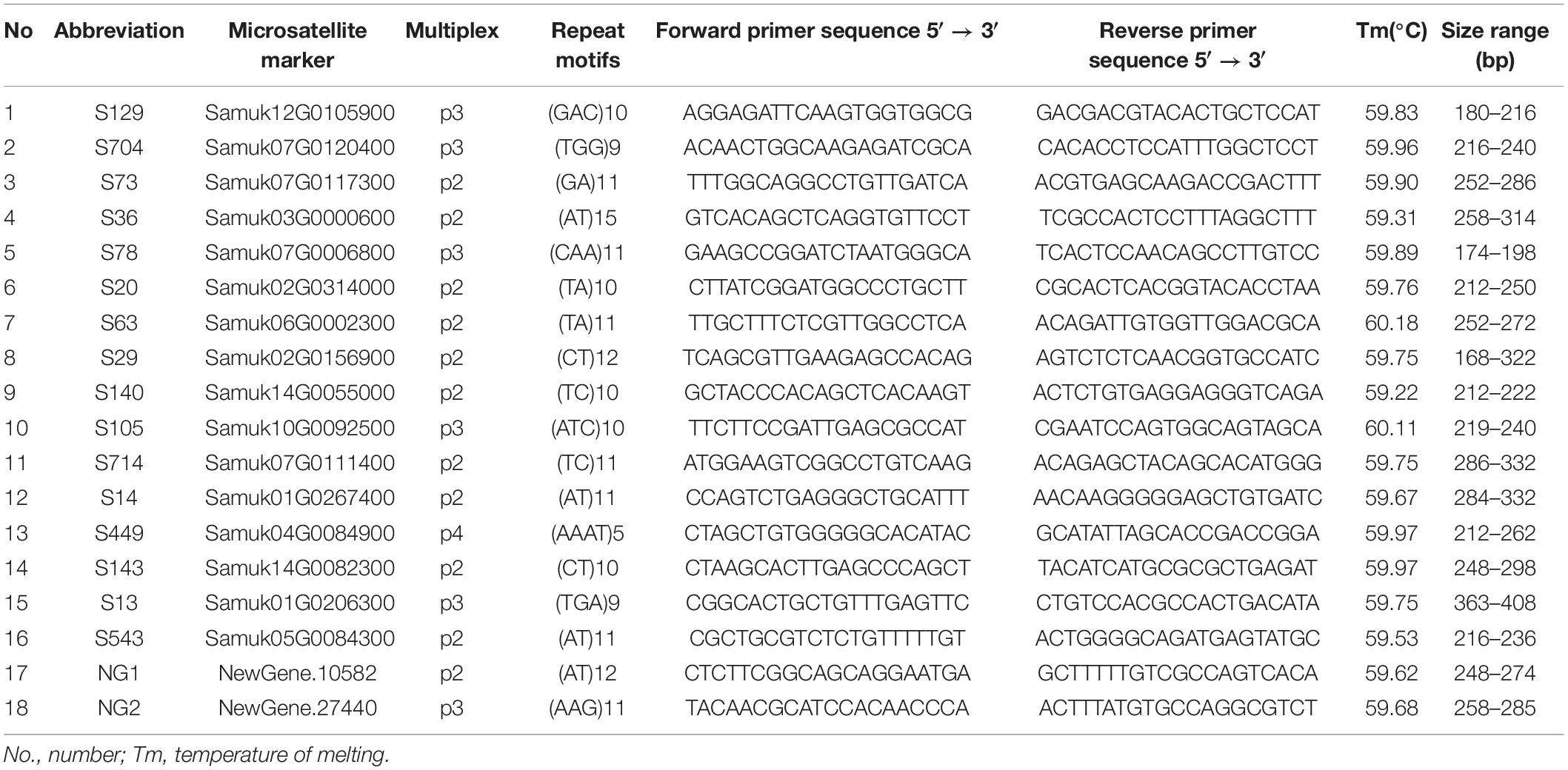
Table 1. Information of 18 EST-SSR primers used in this study.
Plant Materials
The Sapindus population analyzed in this study comprised 161 wild individuals, with 160 individuals from 16 provinces in China and one individual from Vietnam (Figure 1 and Supplementary Table 1). It included 117 S. mukorossi individuals, as well as 36 S. delavayi, four S. rarak, and four S. rarak var. velutinus representatives. The criteria for Sapindus germplasm collection were representative local, naturally superior plants with a diameter at breast height of at least 30 cm. These germplasms were conserved ex situ by grafting at a Sapindus national germplasm nursery in Jianning County, Fujian Province, China (27°06′ N, 117°25′ E), comprising 175 clones and 64 half-sib families by grafting and seeding respectively, with at least 15 plants for each clone. The average temperature in germplasm nursery is 17.4°C, with a maximum temperature of 36.8°C (July) and a minimum temperature of 4.3°C (January), and an average relative humidity of 83.9% (Wang et al., 2020). At present, the grafted clones are 6 years old.
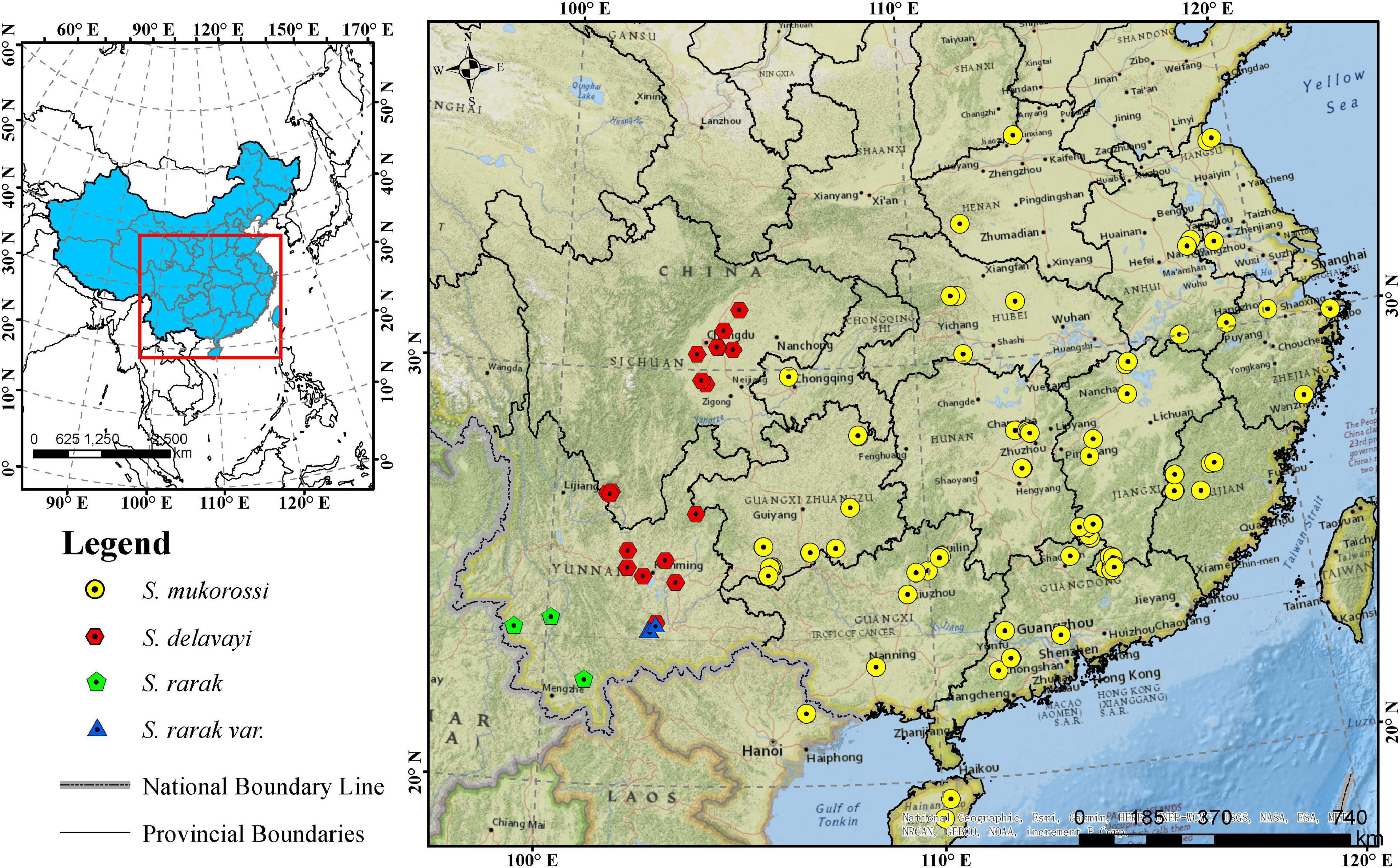
Figure 1. Geographic distribution of 161 Sapindus individuals. The map was generated using ArcMap 10.5 software.
DNA Extraction and Expressed Sequence Tag-Simple Sequence Repeat Genotyping
300 mg fresh leaf tissue from each Sapindus accession was used for DNA extraction with a Plant Genomic DNA Extraction Kit (Tiangen, DP320-03). The DNA samples concentration and quality were determined with a VERTEX 70 UV spectrophotometer (Bruker, Germany).
Eighteen EST-SSR markers were applied to genotype the 161 Sapindus tree samples. The forward primer of each marker was labeled at the 5′ end with a fluorescent dye (FAM, HEX, TAMRA, or ROX) according to the target fragment size of the marker. PCR (Polymerase Chain Reaction) was performed on a T100 thermal cycler (Biorad) in a 20-μL volume that contained 10.0 μL 2×Taq Plus PCR mix [Taq Plus DNA Polymerase (recombinant), 3 mM MgCl2, 0.4 mM dNTPs (dATP, dCTP, dGTP, and dTTP): 0.4 mM], 0.3 μL 10 μM forward primer, 0.3 μL 10 μM reverse primer, and 1.0 μL genomic DNA. The PCR protocol was 5 min denaturation at 94°C; 20 cycles of 30 s at 94°C, 30 s at 52/72/95/50°C (annealing temperature depended on the primer used, see Table 1), and 30 s at 72°C; and a final extension for 5 min at 72°C. Microsatellite alleles were called on an ABI 3730XL DNA analyzer, and the amplicons were statistically analyzed by GeneMarker 2.2.0 software (SoftGenetics, State College, PA, United States).
Data Analysis
For each microsatellite marker, the number of alleles (Na), number of effective alleles (Ne), observed heterozygosity (Ho), expected heterozygosity (He), unbiased expected heterozygosity (uHe), ibreeding coefficient (F), and Shannon’s information index (I) were calculated using GenAlEx v6.5 (Peakall and Smouse, 2006). The polymorphic information content (PIC) was calculated using PowerMarker V3.25 software (Liu and Muse, 2005). F-statistics calculations (FIS, FIT, and FST) and principal coordinate analysis (PCoA) were also performed in GenAlEx v6.5 combined with Microsoft Excel. A neighbor-joining (NJ) tree was generated based on pairwise genetic distances between individuals by using PowerMarker V3.25 (Liu and Muse, 2005), and plotted with iTOL.2
The population structure analysis was performed with Bayesian model-based admixture analyses in STRUCTURE v2.3.4 (Pritchard et al., 2000). We set the number of Markov chain Monte Carlo (MCMC) iterations after burn-in to 100,000 with a 100,000-run length, and the number of genetically homogeneous clusters (K value) ranged from 1 to 20 with 10 replicate runs for each analysis. The optimum K-value was determined by the highest ΔK method (Evanno et al., 2005) in Structure Harvester3 (Earl and VonHoldt, 2012). The structure plot was constructed in R 4.1.0 (R Core Team, 2013).
The optimal set of core germplasm was extracted by the Core Hunter 3 (De Beukelaer et al., 2018) which maximized the genetic variation and allelic richness using local search algorithms. Based on the previously reported distribution of core germplasm fractions in woody plants ranging from 10 to 45% (Belaj et al., 2012; Duan et al., 2017; Feng et al., 2018; Lv et al., 2020), we decided to test 10 sampling fractions (10, 15, 20, 25, 30, 35, 40, 45, and 50% and initial group) respectively by Core Hunter 3. Na, Ne, Ho, He, I, and uHe were calculated separately for each fraction using GenAlEx software, as described above. These indicators were t-tested between the core subset and the initial group using Microsoft Excel. The smallest core subset that did not differ significantly with the 100% population group (P ≤ 0.05) was then selected as the optimal core germplasm collection (Lv et al., 2020).
Results
Genetic Diversity of Sapindus
There were 236 alleles identified by the 18 EST-SSR markers. All 161 individuals could be uniquely genotyped using these 236 alleles, demonstrating the high discrimination capacity of these 18 EST-SSR markers. The markers showed considerable variation (Table 2), with number of alleles (Na) ranging from 6 to 25 (mean 13.1), number of effective alleles (Ne) from 2.625 to 11.503 (mean 5.711), observed heterozygosity (Ho) from 0.277 to 1.000 (mean 0.558), expected heterozygosity (He) from 0.619 to 0.913 (mean 0.798), unbiased expected heterozygosity (uHe) from 0.621 to 0.916 (mean 0.801), Shannon’s information index (I) from 1.202 to 2.683 (mean 1.937), and polymorphic information content (PIC) from 0.561 to 0.907 (mean 0.775). The marker with the highest number of alleles was S36 (25), and the marker with the lowest was S140 (6). The observed heterozygosity of all markers was lower than the expected heterozygosity, with the exception of S105. All markers exhibited high polymorphism (PIC > 0.5; Table 2). The S36 marker captured the most genetic diversity with the highest PIC value (0.907).
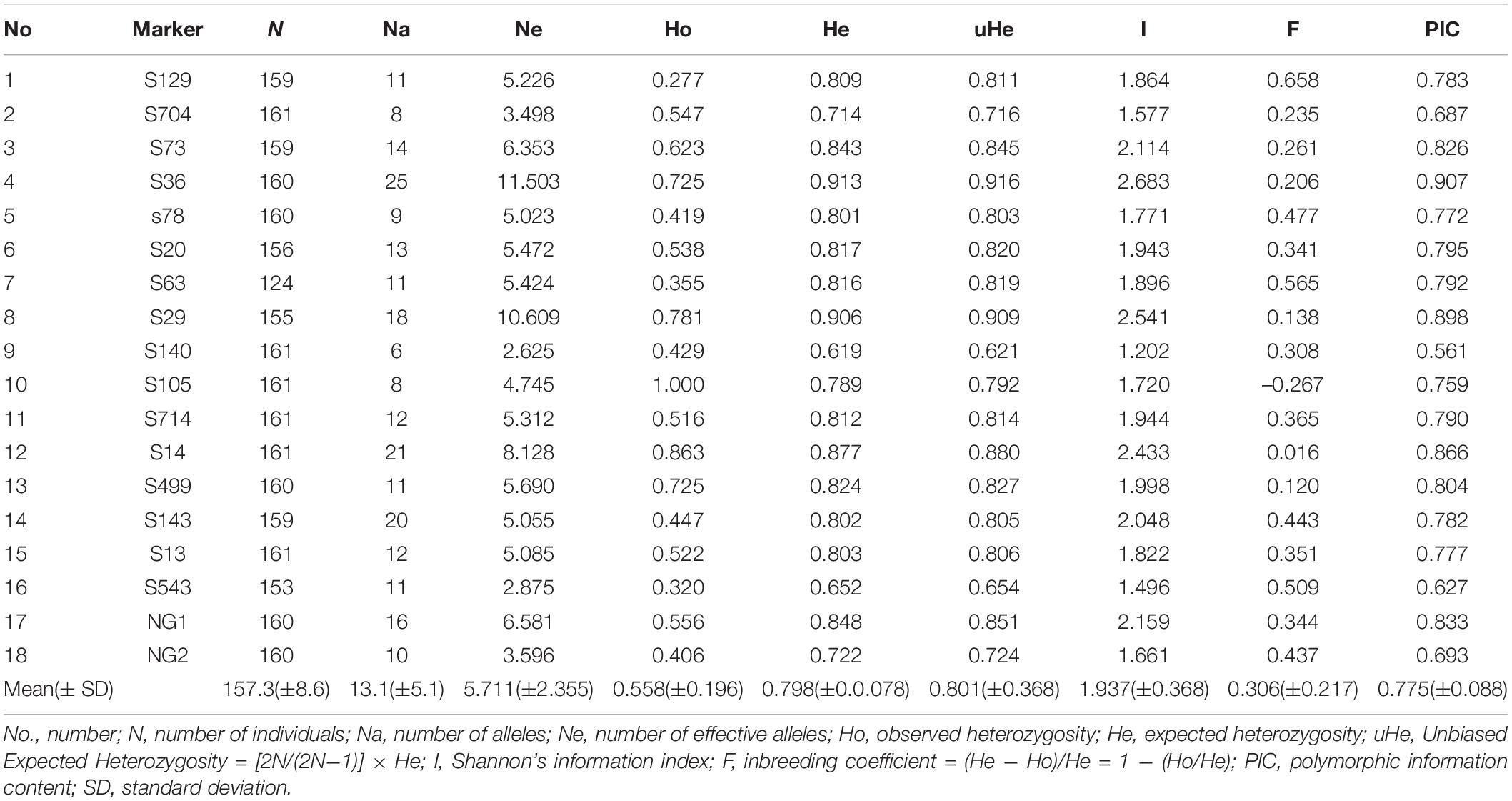
Table 2. Genetic diversity parameters for Sapindus individuals at the 18 microsatellite markers.
There were marked differences in genetic diversity among the four different Sapindus taxa (Table 3). S. mukorossi exhibited the highest Na, Ne, I, Ho, He, and uHe values and the lowest F value compared with S. delavayi, S. rarak, and S. rarak var. velutinus. By contrast, S. rarak var. velutinus showed lower levels of genetic diversity, probably because of its narrow distribution area and smaller number of individuals. The mean pairwise FST coefficient between the four species of Sapindus was 0.154 (Table 4); highest differentiation was between S. delavayi and S. rarak (0.183) and lowest for S. mukorossi versus S. delavayi (0.122).
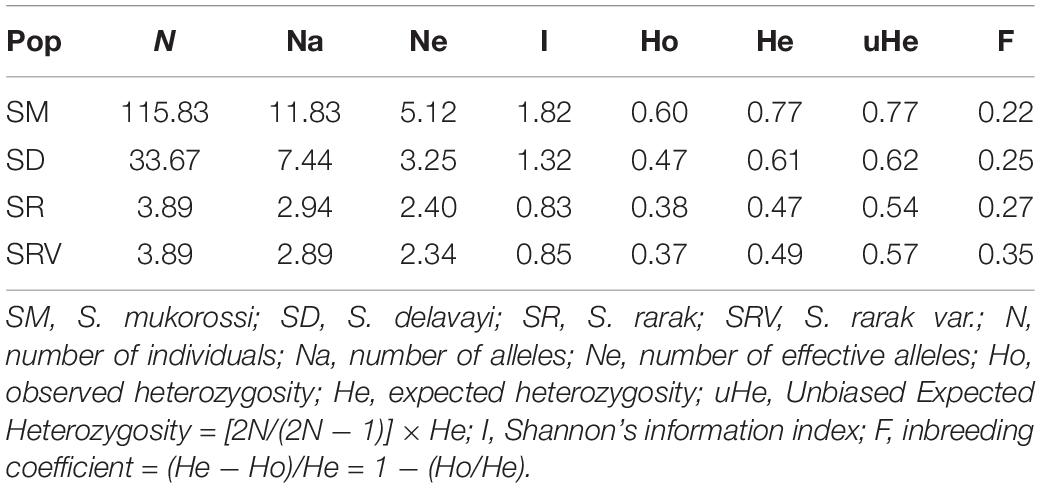
Table 3. Genetic diversity parameters for four different Sapindus species.

Table 4. The pairwise FST comparison among four different Sapindus population.
Genetic Structure of Sapindus
The first and second coordinates of the PCoA analysis accounted for 40.54 and 13.44% of the total genetic variation, respectively (Figure 2). The population of Sapindus was split into three clusters on the first principal coordinate axis (PCoA 1), which corresponded approximately to the S. delavayi group, the S. rarak and S. rarak var. velutinus group, and the S. mukorossi group. On the second principal coordinate axis (PCoA 2), individuals of S. mukorossi were roughly divided into two subgroups: the individuals from southern Guizhou province and the remaining S. mukorossi germplasm. Likewise, S. rarak and S. rarak var. velutinus were also divided into two subgroups along the second principal coordinate axis.
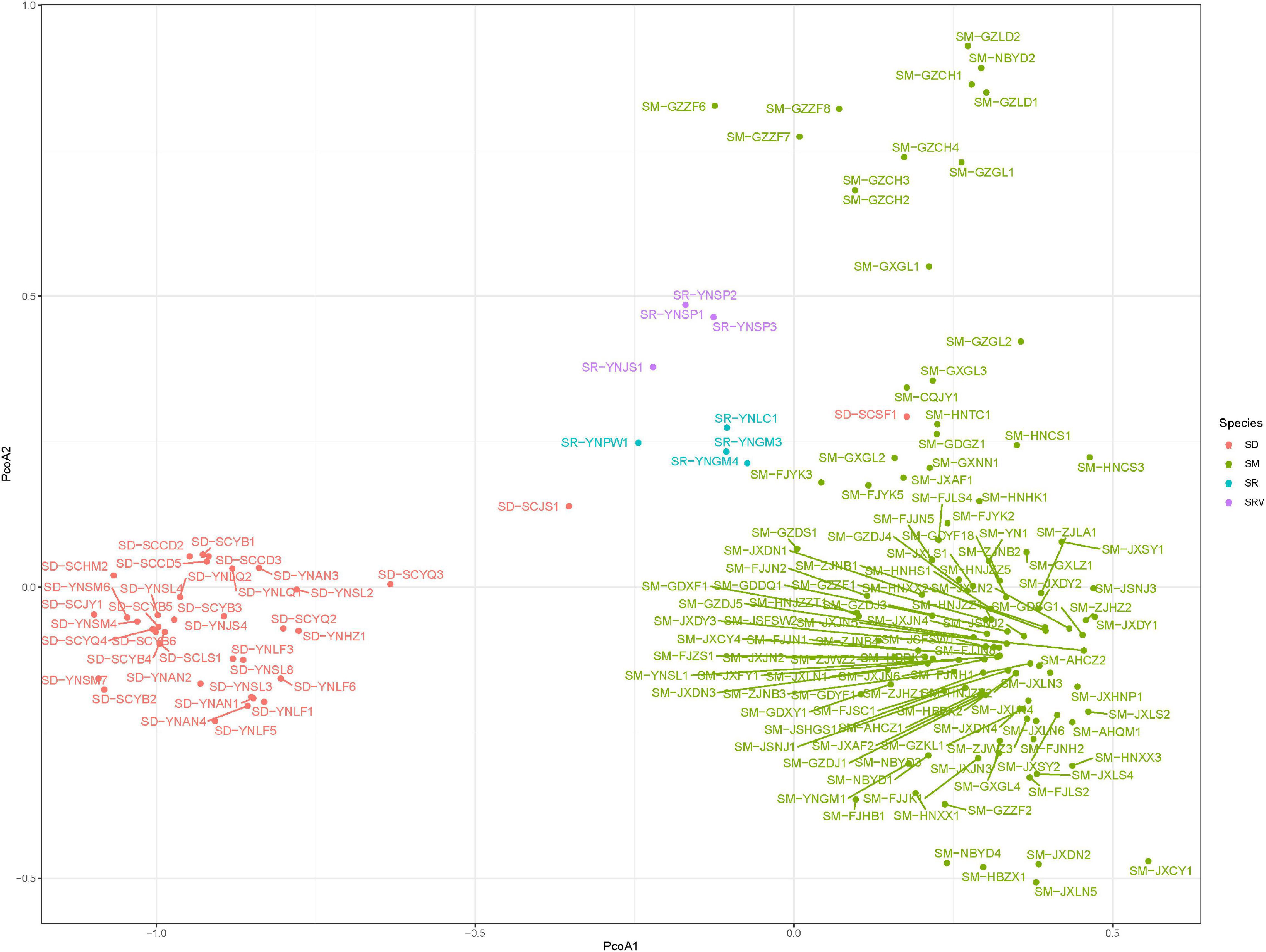
Figure 2. Principal coordinate analysis of 161 Sapindus individuals. SM, S. mukorossi; SD, S. delavayi; SR, S. rarak; SRV, S. rarak var. velutinus.
In contrast to the PCoA results, the STRUCTURE analysis indicated that the Sapindus population could be genetically divided into two distinct subgroups by the ΔK method (Figures 3B,C). Subgroup 1 contained all individuals of S. mukorossi, and subgroup 2 included all individuals of S. delavayi, S. rarak, and S. rarak var. velutinus. When K was equal to 3–6, individuals of subgroup 2 were consistently divided into S. rarak subgroup and S. rarak var. velutinus subgroup (Figure 3A). Furthermore, S. mukorossi individuals were divided into several subgroups (K = 3–6). Intriguingly, individuals from southern Guizhou province formed a separate subgroup (K = 3–6), suggesting that they originated from a distinct ancestral population.
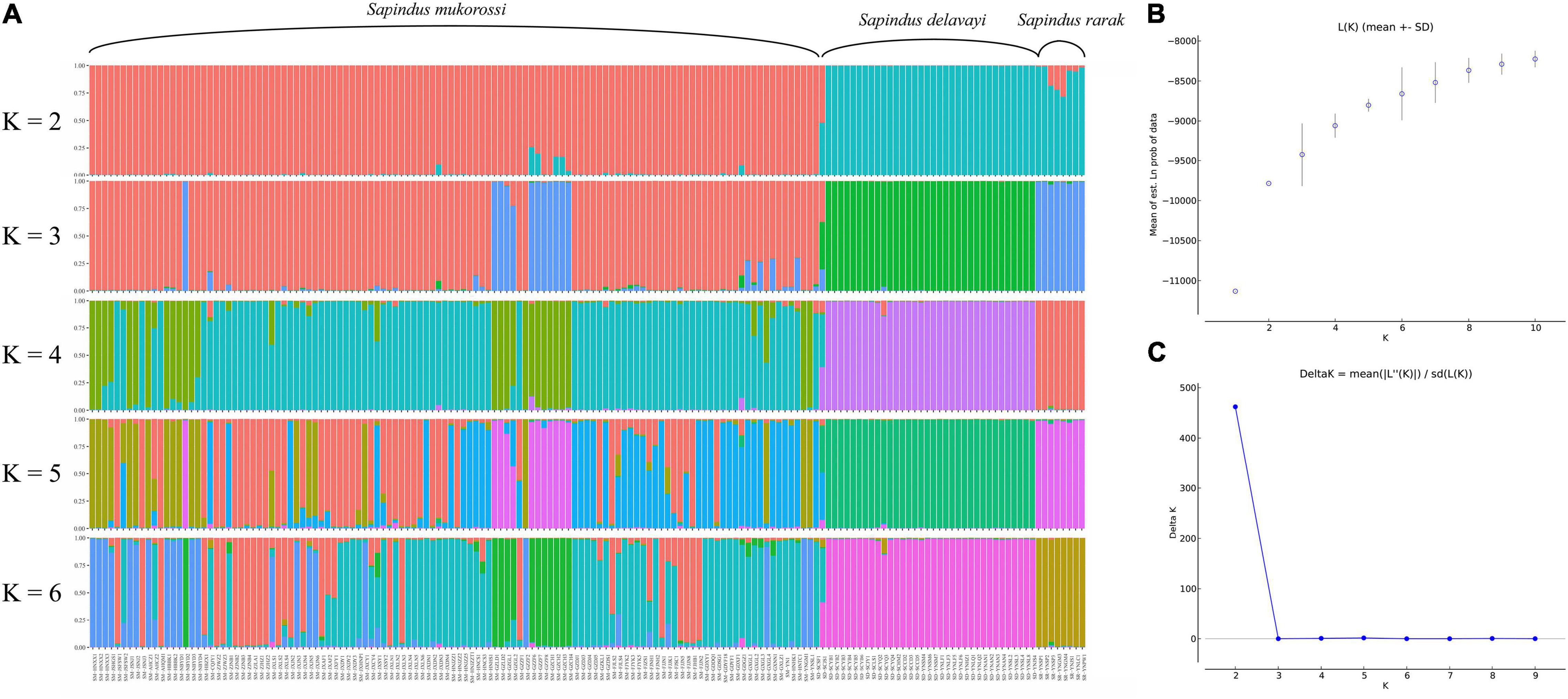
Figure 3. Structure analysis of 161 Sapindus individuals. (A) The population structure of Sapindus was determined using STRUCTURE 2.3.4 software (K = 2–6); (B) Estimated average likelihood L(K) distribution (mean ± SD) from 2 to 10 possible clusters (K); (C) Delta K distribution based on the rate of change in L(K) between continuous K values.
The neighbor-joining dendrogram based on genetic distances among individuals (Figure 4) grouped S. mukorossi, S. delavayi, and S. rarak into their own populations, except for one S. delavayi individual that was grouped together with the S. mukorossi individuals. Furthermore, S. mukorossi individuals could be divided into multiple subgroups which largely correlated with their geographic distribution. Interestingly, some S. mukorossi individuals from Guizhou province appeared to be more closely related to S. rarak.
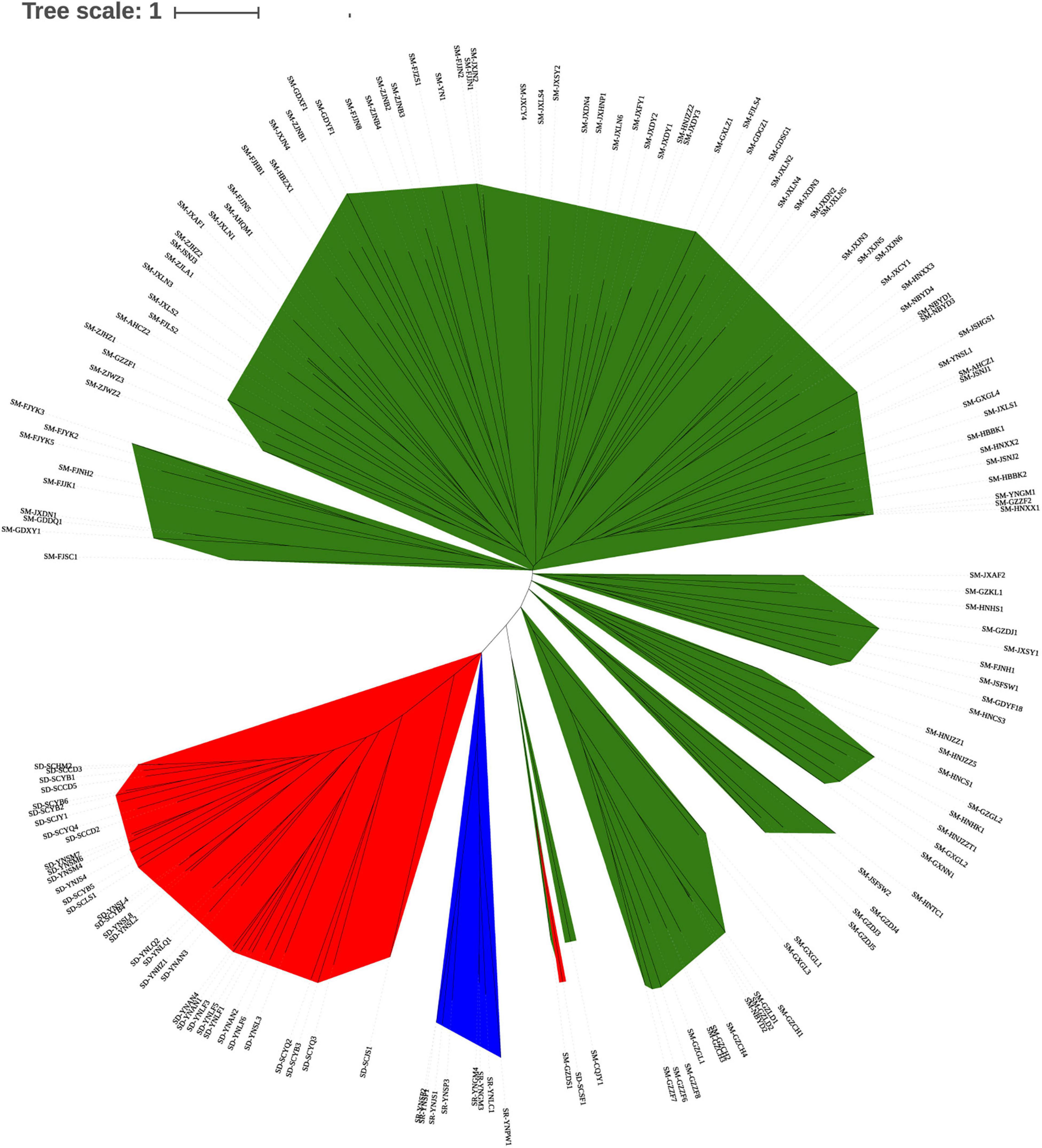
Figure 4. Dendrogram based on genetic distances among individuals in the Sapindus population. Green, S. mukorossi; yellow, S. rarak and S. rarak var. velutinus; red, S. delavayi.
Construction and Evaluation of a Sapindus Core Germplasm Collection
We produced nine candidate core germplasm collections of different sizes using Core Hunter. To determine the optimal core germplasm size, we compared the nine subgroups to the whole population according to six diversity parameters: Na, Ne, Ho, He, uHe, and I (Table 5). The core 10 and 15% subgroups differed significantly (P ≤ 0.05) and highly significantly (P ≤ 0.01) from the full population in Ne and I, respectively. The core 10–25% subgroups differed highly significantly (P ≤ 0.01) and significantly (P ≤ 0.05) in Na from the core 30 and 35% subgroups. The remaining parameters showed no significant differences between the subgroups and the full collection. Hence, the core 40% subgroup was selected as the optimal core germplasm collection. It contained 64 Sapindus individuals: 53 S. mukorossi, nine S. delavayi, one S. rarak, and one S. rarak var. velutinus (Supplementary Table 2), respectively. Relative to the full collection, the core 40% subgroup maintained 80.66% of Na, 97.65% of I, and 90.69% of Ho, and it had 101.90% higher Ne, 103.68% higher He, and 109.59% higher uHe.
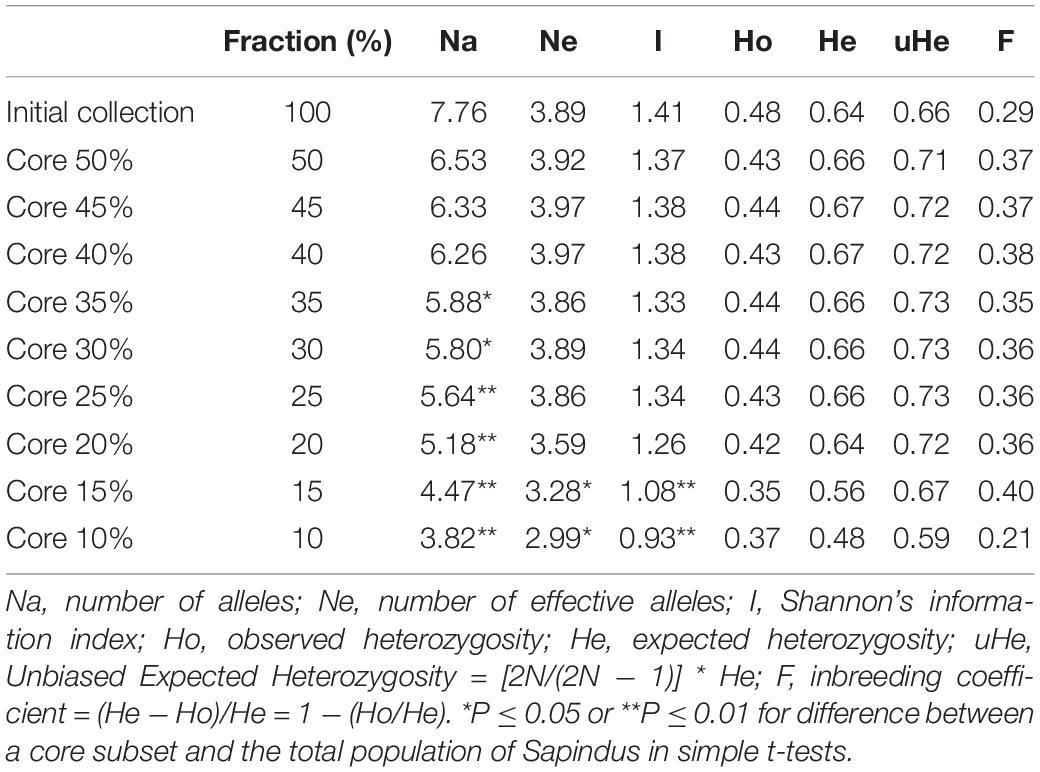
Table 5. Comparison of genetic diversity parameters of different fractions of core germplasm subgroups.
Discussion
Genetic Diversity and Population Structure of Sapindus Germplasm
The collection of natural germplasm resources and genetic diversity evaluation is important for the conservation, breeding, and utilization of germplasm resources (Glaszmann et al., 2010). Previous studies have documented abundant genetic diversity in Sapindus. Diao et al. (2016) and Jiang et al. (2016) found significant genetic divergence among germplasm accessions of S. mukorossi using ISSR markers. Sun et al. (2018) also found significant interspecific genetic differences between S. mukorossi and S. delavayi in China using ISSR markers, and they reported that S. mukorossi could be broadly divided into two subgroups. In this study, for the first time, we have pooled S. mukorossi, S. delavayi, S. rarak, and S. rarak var. velutinus germplasms from China to analyze their genetic diversity and population structure using EST-SSR markers. We also found substantial genetic diversity within the Sapindus germplasm, with S. mukorossi exhibiting the highest genetic variation (Na = 11.83, I = 1.82); S. rarak and S. rarak var. velutinus showed less variation (Na = 2.94 and 2.89 and I = 0.83 and 0.85, respectively) (Tables 2, 3). This may be due to the wider distribution and population size of S. mukorossi; S. rarak and S. rarak var. velutinus have a limited distribution in Yunnan Province, China, with S. rarak found only in Xishuangbanna Dai Autonomous Prefecture, Yunnan Province and S. rarak var. velutinus only in Shiping and Jianshui counties in Honghe Hani and Yi Autonomous Prefecture, Yunnan Province.
It is difficult to distinguish S. mukorossi, S. delavayi, S. rarak, and S. rarak var. velutinus in nature because of their high phenotypic similarity, and there has been a lack of molecular biological support for the species splits in the Sapindus taxonomy. A better understanding of Sapindus genetic structure is a first step toward addressing these issues. Here, we have identified varying levels of genetic divergence among S. mukorossi, S. delavayi, S. rarak, and S. rarak var. velutinus for the first time using SSR markers. Pairwise FST comparisons among the four Sapindus taxa showed high genetic differentiation (Table 4), and PCoA (Figure 2) and a neighbor-joining dendrogram (Figure 4) supported this (Figure 2). However, structure analysis divided the Sapindus germplasms into two subgroups, a S. mukorossi subgroup and subgroup containing the other taxa (Figure 3A), probably due to the higher representation of S. mukorossi among the samples. Structure analysis was developed for comparing populations within a single species and requires the assumption of Hardy-Weinberg equilibrium in the population, which may be violated by the populations of Sapindus studied here (Lv et al., 2020). Hence, PCoA analysis can be a more valid and efficient approach for genetic structure identification owing to its relaxed Hardy-Weinberg equilibrium hypothesis (Lv et al., 2020). We presented the results of structure analysis for K = 3–6 and found that S. mukorossi, S. delavayi, and the combination of S. rarak and S. rarak var. velutinus were divided into three distinct subgroups; S. mukorossi could be further divided into multiple subgroups (Figure 3A). Intriguingly, structure analysis suggested that S. rarak and S. rarak var. velutinus originated from the same ancestral population (Figure 3A), and PCoA analysis also indicated that S. rarak and S. rarak var. velutinus were closely related (Figure 2), supporting the theory that S. rarak var. velutinus is a variety of S. rarak. Moreover, we also found that individuals from southern Guizhou Province formed a distinct subgroup (Figure 3A), consistent with the PCoA analysis in which S. mukorossi individuals were divided into two subgroups along PCoA axis 2 (Figure 2). Previously, (Mahar et al., 2011a,b, 2013) used RAPD, DAMD, and ISSR molecular markers to analyze germplasm of S. mukorossi, Sapindus trifoliatus, and Sapindus emarginatus. They found higher variation in genetic diversity within populations than between populations. Here, we found that Sapindus was genetically diverse, with interspecific genetic variation significantly higher than intraspecific variation. S. mukorossi had higher levels of genetic variation and showed a pattern of geographic variation, whereas S. delavayi, S. rarak, and S. rarak var. velutinus showed low levels of intraspecific genetic variation. These results differ from those of Mahar et al., perhaps because their population originated in India, where the germplasm collection was smaller and unevenly distributed (Mahar et al., 2011a,b, 2013). It is also possible that interspecific incompatibility is lower between the Indian species. Overall, we believe that these results provide molecular biological support for the current consensus taxonomy of Sapindus in China. Furthermore, it suggests that special attention should be paid to Sapindus rarak var. velutinus with respect to its relationship with the other Sapindus species when carrying phylogenetic studies in future.
The Core Germplasm Collection of Sapindus
The construction of a core germplasm resource is an effective way to achieve efficient, scientific and rational conservation and utilization of genetic diversity (Xu et al., 2020), and the selection of an appropriate core population size is a crucial factor in establishing a core germplasm collection. Balakrishnan et al. (2000) and Zhang et al. (2010) suggested that the proportion of core germplasm should be determined by the size of the initial germplasm resource. Li et al. (2002) recommended sampling 5–40% of the core germplasm in crops, with 10% being optimal. However, woody plants are more genetically diverse, and the sampling percentage of core germplasm for woody plants is typically in the range of 10–45%. For instance, 14.71% (64/435) in apple (Malus domestica Borkh.) (Zhang et al., 2010), 17.96% (30/167) in Citrus reticulata (Garcia-Lor et al., 2017), 35% (247/707) in Eucalyptus cloeziana F. Muell (Lv et al., 2020), 35% (63/180) in Ginkgo biloba (Xuan et al., 2016), and 42.9% (300/700) in Chinese fir (Duan et al., 2017).
In this study, we selected a 40% (64/161) subgroup of the Sapindus core germplasm using Core Hunter. The population contained 53 S. mukorossi, nine S. delavayi, one S. rarak, and one S. rarak var. velutinus individuals (Supplementary Table 2). During the construction of core germplasm collections, allele retention is frequently considered as an evaluation criterion. For example, in Saccharum officinarum germplasm collection the criteria were to retain at least 70% of allele richness as well as other genetic diversity parameters (Balakrishnan et al., 2000). Compared with all Sapindus germplasm, the core germplasm collection had higher genetic diversity and maintained 80.66% of the allelic richness (Table 5), showing a balanced geographic composition (Supplementary Table 2). Previously, the core germplasm collection of lychee using 18 SSR markers resulted in 29.92% (38/127) individuals (Wang et al., 2012). The combined results indicate that the Sapindus core germplasm collection constructed in this study well represent the initial collection. The non-core germplasm of Sapindus population, also called reserve collection of Sapindus, is important for the conservation and utilization of Sapindus diversity, and it may harbor specific phenotypic, phenological and ecological characteristics to be of future use, thus it is important to conserve, exploit and understand the reserve collection as well. Although we have achieved our objectives of exploring genetic diversity and population structure in Sapindus and constructing a core germplasm collection using EST-SSR markers, our study still have some limitations. Compared with the millions of single nucleotide polymorphisms (SNP) and indel markers that can be obtained by whole genome resequencing or transcriptome sequencing, the number of EST-SSR markers in this study is relatively low, and the results obtained are not sufficient to fully elucidate the genetic structure of Sapindus. Hence, in future studies we hope to perform transcriptome sequencing or resequencing of this core germplasm collection to further explore interspecific and intraspecific genetic structure, evolutionary history, and regions under natural selection in Sapindus. Nevertheless, we believe the results of this study can help facilitate the efficient management, conservation, and utilization of Sapindus germplasm resources in the future.
Conclusion
In this study, we have revealed high genetic diversity and significant genetic structure in Sapindus germplasms using 18 EST-SSR markers. Interspecific genetic variation was significantly higher than intraspecific variation in S. mukorossi, S. delavayi, and S. rarak plus S. rarak var. velutinus populations. S. mukorossi had abundant genetic variation and showed a pattern of geographic variation, whereas S. delavayi, S. rarak, and S. rarak var. velutinus showed less intraspecific genetic variation. A core germplasm collection was defined as 40% of the initial population; it comprised 53 S. mukorossi, nine S. delavayi, one S. rarak, and one S. rarak var. velutinus. The results obtained here provide a germplasm basis and theoretical rationale for the efficient management, conservation, and utilization of Sapindus germplasm, as well as genetic resources for joint genomics research in the future.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
JL, SG, LJ, and JS conceived and designed the project. YX completed the transcriptome sequencing of Sapindus mukorossi. JL, SG, MW, and XW collected the samples. JL, SG, and MW performed molecular labwork and scored the markers. JL, JN, NR, and JY analyzed the data. JL wrote the manuscript with input from JS, and feedback from all the authors. LJ contributed to the special foundation for National Science and Technology Basic Research Program of China and National Natural Science Foundation of China. JL contributed to the China Scholars Council. JS contributed to the Academy of Finland. All authors read and approved final manuscript.
Funding
This work was supported by Special Foundation for National Science and Technology Basic Research Program of China (No. 2019FY100803), the National Natural Science Foundation of China (32071793), the China Scholarship Council, and the Academy of Finland (decisions 346100 and 319947).
Conflict of Interest
XW was employed by Yuanhua Forestry Biological Technology Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to thank Caowen Sun, Yulin Zheng, and Ying Cao for valuable cooperation in germplasms collection and maintenance. We also thank Yulin Zheng for technical assistance in DNA extraction.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.857993/full#supplementary-material
Footnotes
- ^ https://webblast.ipk-gatersleben.de/misa/
- ^ https://itol.embl.de/
- ^ http://taylor0.biology.ucla.edu/structureHarvester/
References
Adams, M. D., Kelley, J. M., Gocayne, J. D., Dubnick, M., Polymeropoulos, M. H., Xiao, H., et al. (1991). Complementary DNA sequencing: expressed sequence tags and human genome project. Science 252, 1651–1656. doi: 10.1126/science.2047873
Balakrishnan, R., Nair, N., and Sreenivasan, T. (2000). A method for establishing a core collection of Saccharum officinarum L. germplasm based on quantitative-morphological data. Genet. Resour. Crop Evol. 47, 1–9.
Basu, A., Basu, S., Bandyopadhyay, S., and Chowdhury, R. (2015). Optimization of evaporative extraction of natural emulsifier cum surfactant from Sapindus mukorossi—Characterization and cost analysis. Indus. Crops Prod. 77, 920–931. doi: 10.1016/j.indcrop.2015.10.006
Belaj, A., del Carmen Dominguez-García, M., Atienza, S. G., Urdíroz, N. M., De la Rosa, R., Satovic, Z., et al. (2012). Developing a core collection of olive (Olea europaea L.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genet. Genomes 8, 365–378. doi: 10.1007/s11295-011-0447-6
Brown, A. H. D. (1989). Core collections: a practical approach to genetic resources management. Genome 31, 818–824. doi: 10.1111/jnu.12259
Caowen, S., Liming, J., Benye, X., Jiming, L., and Lianchun, W. (2019). Genetic diversity and association analyses of fruit traits with microsatellite ISSRs in Sapindus. J. For. Res. 30, 193–203. doi: 10.1007/s11676-017-0580-7
Chakraborty, M., and Baruah, D. C. (2013). Production and characterization of biodiesel obtained from Sapindus mukorossi kernel oil. Energy 60, 159–167. doi: 10.1016/j.energy.2013.07.065
Cosson, P., Hastoy, C., Errazzu, L. E., Budeguer, C. J., Boutié, P., Rolin, D., et al. (2019). Genetic diversity and population structure of the sweet leaf herb, Stevia rebaudiana B., cultivated and landraces germplasm assessed by EST-SSRs genotyping and steviol glycosides phenotyping. BMC Plant Biol. 19:436. doi: 10.1186/s12870-019-2061-y
De Beukelaer, H., Davenport, G. F., and Fack, V. (2018). Core Hunter 3: flexible core subset selection. BMC Bioinformatics 19:203. doi: 10.1186/s12859-018-2209-z
Dervishi, A., Jakše, J., Ismaili, H., Javornik, B., and Štajner, N. (2021). Genetic structure and core collection of olive germplasm from albania revealed by microsatellite markers. Genes 12:256. doi: 10.3390/genes12020256
Diao, S., Shao, W., Chen, T., Jiang, J., and Duan, W. (2016). Genetic diversity of Sapindus mokorossi natural populations in China based on ISSR. For. Res. 29, 176–182.
Duan, H., Cao, S., Zheng, H., Hu, D., Lin, J., Cui, B., et al. (2017). Genetic characterization of Chinese fir from six provinces in southern China and construction of a core collection. Sci. Rep. 7:13814. doi: 10.1038/s41598-017-13219-0
Earl, D. A., and VonHoldt, B. M. (2012). STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 4, 359–361. doi: 10.1007/s12686-011-9548-7
Evanno, G., Regnaut, S., and Goudet, J. (2005). Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14, 2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x
Feng, Y., Yang, Z., Tan, J., Li, H., and Chen, X. (2018). Selection of first generation nucleus population of Pinus massoniana in Guangxi. J. Northeast For. Univ. 46, 20–24.
Frankel, O. H. (1984). “Genetic perspectives of germplasm conservation,” in Genetic Manipulation Impact on Man & Society, eds W. K. Arber, K. Llimensee, W. J. Peacock, and P. Stralinger (Cambridge: Cambridge University Press), 161–170.
Garcia-Lor, A., Luro, F., Ollitrault, P., and Navarro, L. (2017). Comparative analysis of core collection sampling methods for mandarin germplasm based on molecular and phenotypic data. Ann. Appl. Biol. 171, 327–339. doi: 10.1111/aab.12376
Glaszmann, J.-C., Kilian, B., Upadhyaya, H. D., and Varshney, R. K. (2010). Accessing genetic diversity for crop improvement. Curr. Opin. Plant Biol. 13, 167–173. doi: 10.1016/j.pbi.2010.01.004
Guo, X., Cheng, F., and Zhong, Y. (2020). Genetic diversity of Paeonia rockii (flare tree peony) germplasm accessions revealed by phenotypic traits, EST-SSR markers and chloroplast DNA sequences. Forests 11:672. doi: 10.3390/f11060672
Hintum, J. L., Brown, A., Spillane, C., and Hodgkin, T. (2000). Core collections of plant genetic resources. IPGRI Technical Bulletin No.3, Vol. 48. Rome: International Plant Genetic Resources Institute, 3.
Jiang, C., Lu, X., and Ye, X. (2016). Molecular genetic diversity analysis of energy plantsapindus mukorossi germplasm resources. Mol. Plant Breed. 14, 2888–2895.
Li, Z., Zhang, H., Zeng, Y., Yang, Z., Shen, S., Sun, C., et al. (2002). Studies on sampling schemes for the establishment of corecollection of rice landraces in Yunnan, China. Genet. Resour. Crop Evol. 49, 67–74.
Liu, J., Chen, Z., Sun, C., Wang, L., He, Q., Dai, T., et al. (2019). Variation in fruit and seed properties and comprehensive assessment of germplasm resources of the genus Sapindus. Sci. Silva Sin. 55, 44–54.
Liu, J., Liu, S., Xu, Y., Sun, C., Chen, Z., Wang, X., et al. (2021a). Screening of Sapindus germplasm resources in China based on agro-morphological traits. J. For. Res. 33, 203–216. doi: 10.1007/s11676-021-01350-8
Liu, J., Xu, Y., Sun, C., Wang, X., Zheng, Y., Shi, S., et al. (2021b). Distinct ecological adaptations and habitat responses to future climate change in three east and Southeast Asian Sapindus species. For. Ecol. Manage. 507:119982. doi: 10.1016/j.foreco.2021.119982
Liu, J., Sun, C., He, Q., Jia, L. M., Weng, X. H., and Yu, J. (2017). Research progress in Sapindus L. germplasm resources. World For. Res. 30, 15–21.
Liu, K., and Muse, S. V. (2005). PowerMarker: an integrated analysis environment for genetic marker analysis. Bioinformatics 21, 2128–2129. doi: 10.1093/bioinformatics/bti282
Lv, J., Li, C., Zhou, C., Chen, J., Li, F., Weng, Q., et al. (2020). Genetic diversity analysis of a breeding population of Eucalyptus cloeziana F. Muell.(Myrtaceae) and extraction of a core germplasm collection using microsatellite markers. Indus. Crops Prod. 145:112157. doi: 10.1016/j.indcrop.2020.112157
Mahar, K. S., Rana, T. S., and Anand, S. (2011a). Genetic variability and population structure in Sapindus emarginatus Vahl from India. Gene 485, 32–39. doi: 10.1016/j.gene.2011.05.036
Mahar, K. S., Rana, T. S., and Ranade, S. A. (2011b). Molecular analyses of genetic variability in soap nut (Sapindus mukorossi Gaertn.). Indus. Crops Prod. 34, 1111–1118. doi: 10.1016/j.indcrop.2011.03.029
Mahar, K. S., Rana, T. S., Ranade, S. A., Pande, V., and Palni, L. M. S. (2013). Estimation of genetic variability and population structure in Sapindus trifoliatus L., using DNA fingerprinting methods. Trees 27, 85–96. doi: 10.1007/s00468-012-0770-z
Min, L., Xiuqin, Y., Jing, B., Huaishan, Z., Daoping, W., and Huaming, A. (2017). Construction of core collection in wild Rosa roxburghii from Guizhou province using EST-SSR markers and fruits quality traits. Acta Hortic. Sin. 44:1486.
Mukherjee, S., Mukhopadhyay, S., Pariatamby, A., Hashim, M. A., Redzwan, G., and Gupta, B. S. (2015). Optimization of pulp fibre removal by flotation using colloidal gas aphrons generated from a natural surfactant. J. Taiwan Inst. Chem. Eng. 53, 15–21. doi: 10.1016/j.jtice.2015.02.037
Mukhopadhyay, S., Hashim, M. A., Sahu, J. N., Yusoff, I., and Gupta, B. S. (2013). Comparison of a plant based natural surfactant with SDS for washing of As(V) from Fe rich soil. J. Environ. Sci. 25, 2247–2256. doi: 10.1016/s1001-0742(12)60295-2
Mukhopadhyay, S., Mukherjee, S., Adnan, N. F., Hayyan, A., Hayyan, M., Hashim, M. A., et al. (2016). Ammonium-based deep eutectic solvents as novel soil washing agent for lead removal. Chem. Eng. J. 294, 316–322. doi: 10.1016/j.chemosphere.2021.130568
Muntaha, S. T., and Khan, M. N. (2015). Natural surfactant extracted from Sapindus mukurossi as an eco-friendly alternate to synthetic surfactant - a dye surfactant interaction study. J. Clean. Product. 93, 145–150. doi: 10.1016/j.jclepro.2015.01.023
Parthiban, S., Govindaraj, P., and Senthilkumar, S. (2018). Comparison of relative efficiency of genomic SSR and EST-SSR markers in estimating genetic diversity in sugarcane. 3 Biotech 8, 1–12. doi: 10.1007/s13205-018-1172-8
Peakall, R., and Smouse, P. E. (2006). GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 6, 288–295. doi: 10.1093/bioinformatics/bts460
Pelegrini, B. L., Sudati, E. A., Re, F., Moreira, A. L., Piloto Ferreira, I. C., Sampaio, A. R., et al. (2017). Thermal and rheological properties of soapberry Sapindus saponaria L. (Sapindaceae) oil biodiesel and its blends with petrodiesel. Fuel 199, 627–640. doi: 10.1016/j.fuel.2017.02.059
Powell, W., Machray, G. C., and Provan, J. (1996). Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1, 215–222. doi: 10.1016/s1360-1385(96)86898-0
Preethi, P., Rahman, S., Naganeeswaran, S., Sabana, A., Gangaraj, K., Jerard, B., et al. (2020). Development of EST-SSR markers for genetic diversity analysis in coconut (Cocos nucifera L.). Mol. Biol. Rep. 47, 9385–9397. doi: 10.1007/s11033-020-05981-8
Pritchard, J. K., Stephens, M., and Donnelly, P. (2000). Inference of population structure using multilocus genotype data. Genetics 155, 945–959. doi: 10.1093/genetics/155.2.945
R Core Team (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rodriguez-Hernández, D., Demuner, A. J., Barbosa, L. C. A., Csuk, R., and Heller, L. (2015). Hederagenin as a triterpene template for the development of new antitumor compounds. Eur. J. Med. Chem. 105, 57–62. doi: 10.1016/j.ejmech.2015.10.006
Singh, P. T. D., and Singh, M. M. (2008). Anti-Trichomonas activity of Sapindus saponins, a candidate for development as microbicidal contraceptive. J. Antimicrob. Chemother. 62, 526–534. doi: 10.1093/jac/dkn223
Sun, C., Jia, L., Xi, B., Wang, L., and Weng, X. (2017). Natural variation in fatty acid composition of Sapindus spp. seed oils. Indus. Crops Prod. 102, 97–104. doi: 10.1016/j.indcrop.2017.03.011
Sun, C., Jia, L., Ye, H. O., Gao, Y., and Weng, X. (2016). Geographic variation evaluating and correlation with fatty acid composition of economic characters of Sapindus spp. Fruits. J. Beijing For. Univ. 12, 73–83.
Sun, C., Wang, L., Liu, J., Zhao, G., Gao, S., Xi, B., et al. (2018). Genetic structure and biogeographic divergence among Sapindus species: an inter-simple sequence repeat-based study of germplasms in China. Indus. Crops Prod. 118, 1–10. doi: 10.1016/j.indcrop.2018.03.029
Thiel, T., Michalek, W., Varshney, R., and Graner, A. (2003). Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 106, 411–422. doi: 10.1007/s00122-002-1031-0
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B. C., Remm, M., et al. (2012). Primer3—new capabilities and interfaces. Nucleic Acids Res. 40:e115. doi: 10.1093/nar/gks596
Wang, M., Dzievit, M., Chen, Z., Morris, J. B., Norris, J., Barkley, N. A., et al. (2017). Genetic diversity and population structure of castor (Ricinus communis L.) germplasm within the US collection assessed with EST-SSR markers. Genome 60, 193–200. doi: 10.1139/gen-2016-0116
Wang, X., Liu, J., Rui, X., Xu, Y., and Jia, L. (2020). Biogeographic divergence in leaf traits of Sapindus mukorossi and Sapindus delavayi and its relation to climate. J. For. Res. 32, 1445–1456. doi: 10.1007/s11676-020-01206-7
Wang, Y., Fu, J., Pan, L., Lu, B., Lin, S., Yang, X., et al. (2012). “Construction of core collection of lychee by SSR marker,” in Proceedings of the IV International Symposium on Lychee, Longan and Other Sapindaceae Fruits, White River, 87–92. doi: 10.17660/actahortic.2014.1029.9
Xie, J., Zuo, J., Huang, Y., Li, C., and Chen, Y. (2020). The origin and germplasm collection for cultivated Dendrobium officinale K. Kimura & Migo individuals revealed by EST-SSR markers. Genet. Resour. Crop Evol. 67, 1209–1219. doi: 10.1007/s10722-020-00906-7
Xu, J., Wang, L., Wang, H., Mao, C., Kong, D., Chen, S., et al. (2020). Development of a core collection of six-rowed hulless barley from the qinghai-tibetan plateau. Plant Mol. Biol. Rep. 38, 305–313. doi: 10.1007/s11105-020-01196-0
Xu, Y., Jia, L., Chen, Z., and Gao, Y. (2018). Advances on triterpenoid saponin of Sapindus mukorossi. Chem. Bull. 081, 1078–1088.
Xuan, W., Xiao-jing, L., Shi-yan, X., Qian-qian, K., Yan-hui, Z., Li-min, S., et al. (2016). AFLP analysis of genetic diversity and a construction of the core collection of partial ancient ginkgo trees in China. Acta Hortic. Sin. 43:249.
Ye, Y., Feng, L., Ju, Y., Cai, M., Cheng, T., Wang, J., et al. (2017). Genetic evaluation and core collection construction of crape myrtle accessions using newly developed EST-SSR markers. PeerJ Preprints 5:e3141v1.
Keywords: genetic diversity, population structure, core germplasm, Sapindus mukorossi, Sapindus delavayi, Sapindus rarak, Sapindus rarak var
Citation: Liu J, Gao S, Xu Y, Wang M, Ngiam JJ, Rui Wen NC, Yi JJJ, Weng X, Jia L and Salojärvi J (2022) Genetic Diversity Analysis of Sapindus in China and Extraction of a Core Germplasm Collection Using EST-SSR Markers. Front. Plant Sci. 13:857993. doi: 10.3389/fpls.2022.857993
Received: 19 January 2022; Accepted: 04 May 2022;
Published: 24 May 2022.
Edited by:
Keiichi Okazaki, Niigata University, JapanReviewed by:
Robert Jarret, Plant Genetic Resources Conservation Unit, Agricultural Research Service (USDA), United StatesShengzuo Fang, Nanjing Forestry University, China
Copyright © 2022 Liu, Gao, Xu, Wang, Ngiam, Rui Wen, Yi, Weng, Jia and Salojärvi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Liming Jia, jlm@bjfu.edu.cn; Jarkko Salojärvi, jarkko@ntu.edu.sg
†These authors have contributed equally to this work