Genome-wide Identification and Expression Analysis of TALE Gene Family in Pomegranate (Punica granatum L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome and Transcriptome Data Sources
2.2. Identification and Sequence Analysis of PgTALE Gene Family Members
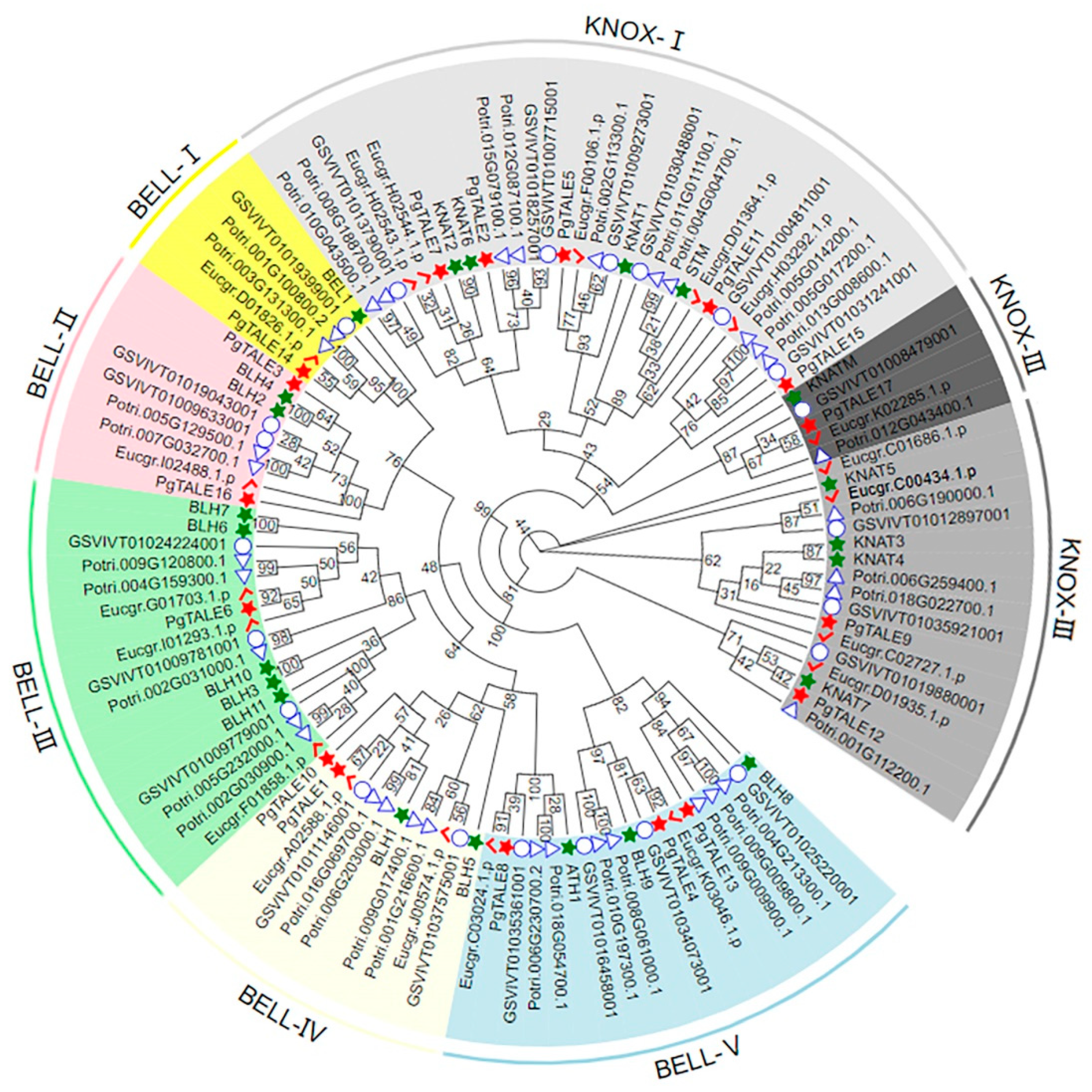
2.3. Construction of Phylogenetic tree of PgTALE Gene Family
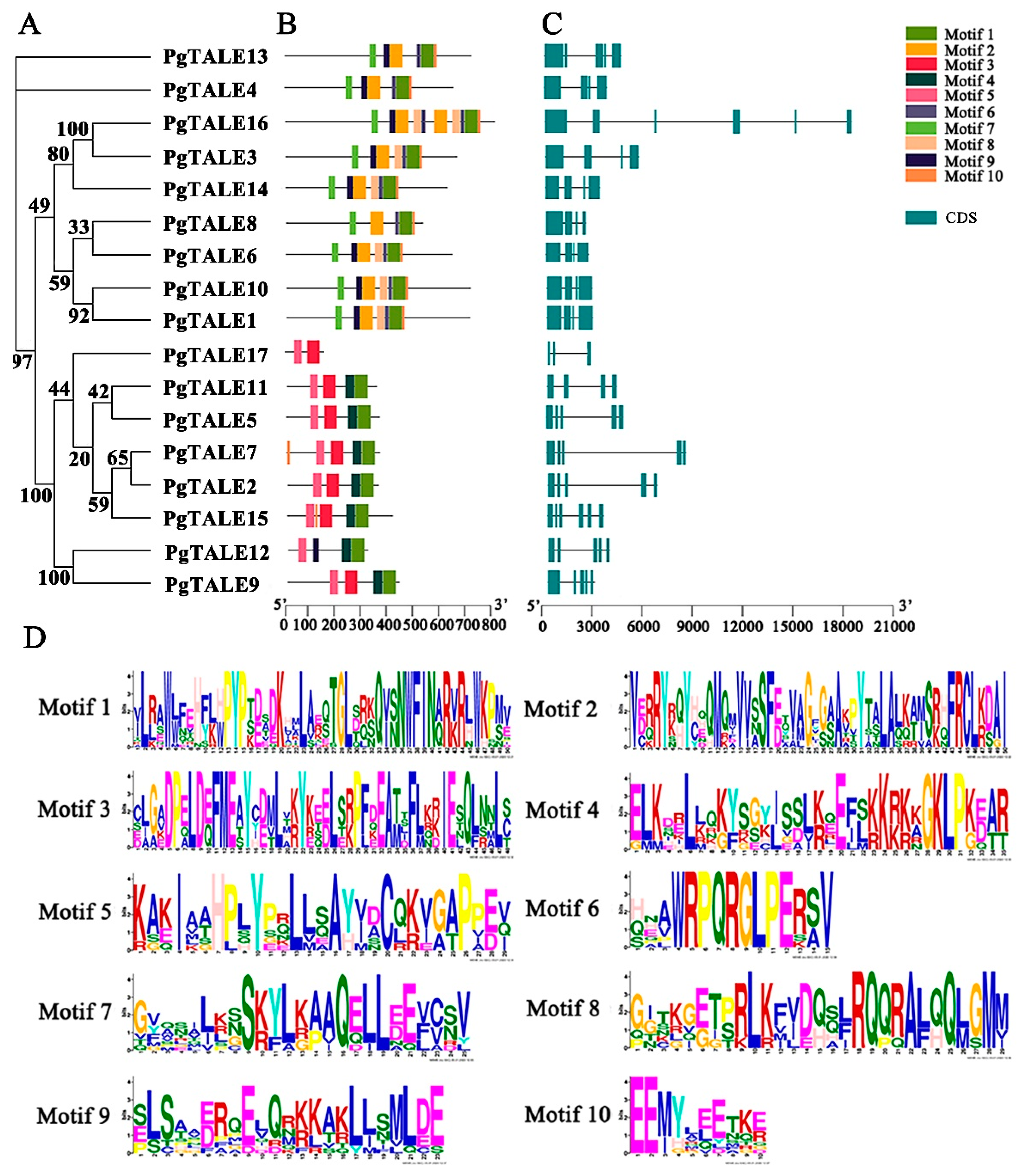
2.4. Analysis of PgTALE Conserved Motifs and Gene Structure
2.5. Analysis of PgTALE Protein Structure
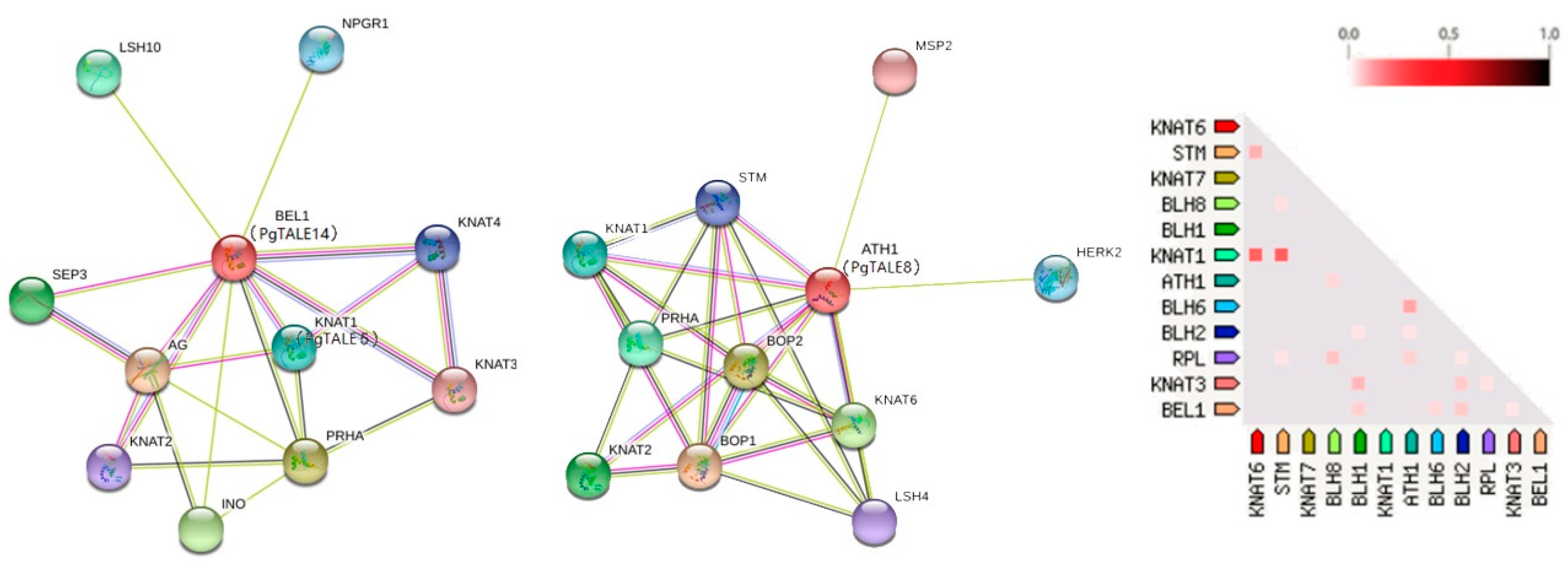
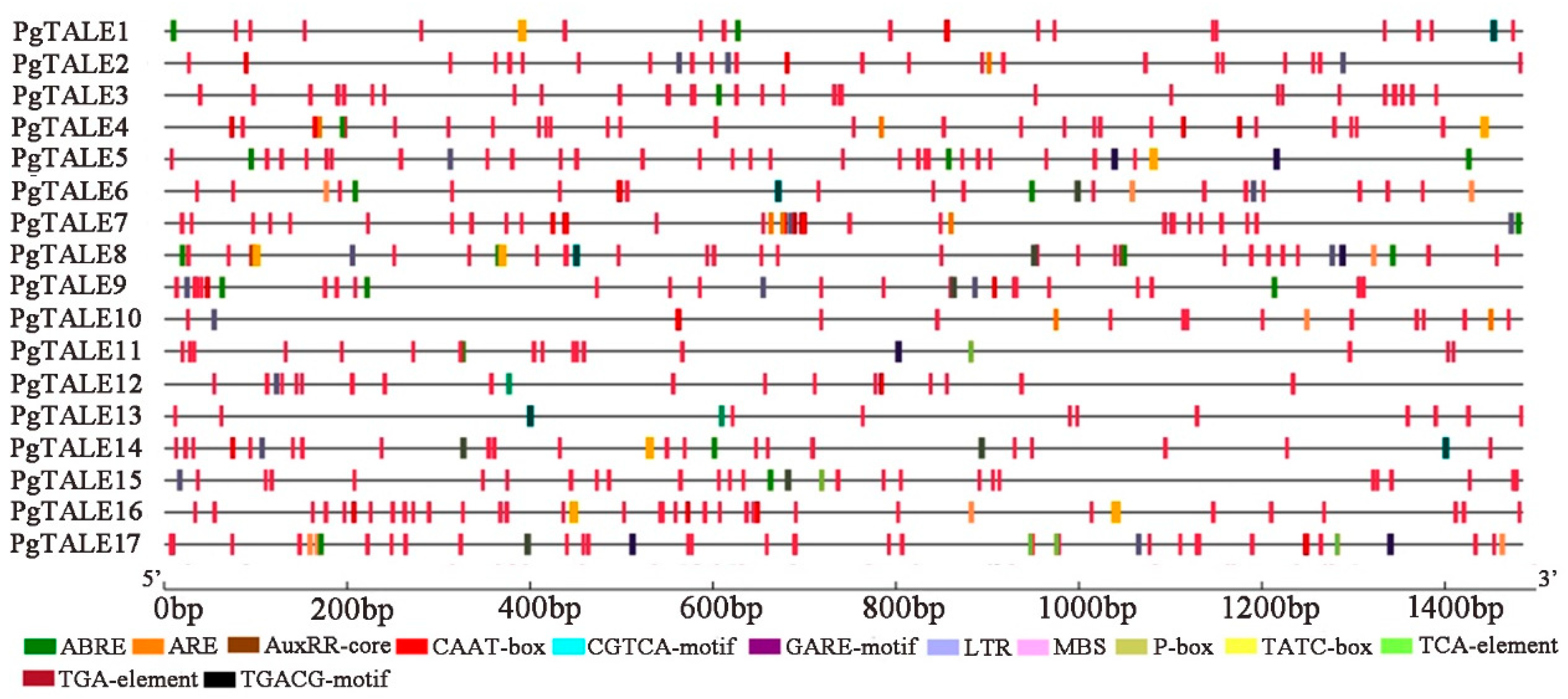
2.6. Analysis of Cis-elements and Protein-protein Interaction Network of PgTALE Gene Family
2.7. Expression Analysis of PgTALE Gene Family
3. Results
3.1. Identification and Sequence Analysis of PgTALE Gene Family Members
3.2. Phylogenetic Tree Analysis of PgTALE Gene Family
3.3. Analysis of Conserved Motifs and Gene Structures of PgTALE Gene Family
3.4. Protein Structure Analysis and Protein Interaction Networks of Pomegranate TALE Gene Family
3.5. Analysis of Cis-elements of PgTALE Gene Family
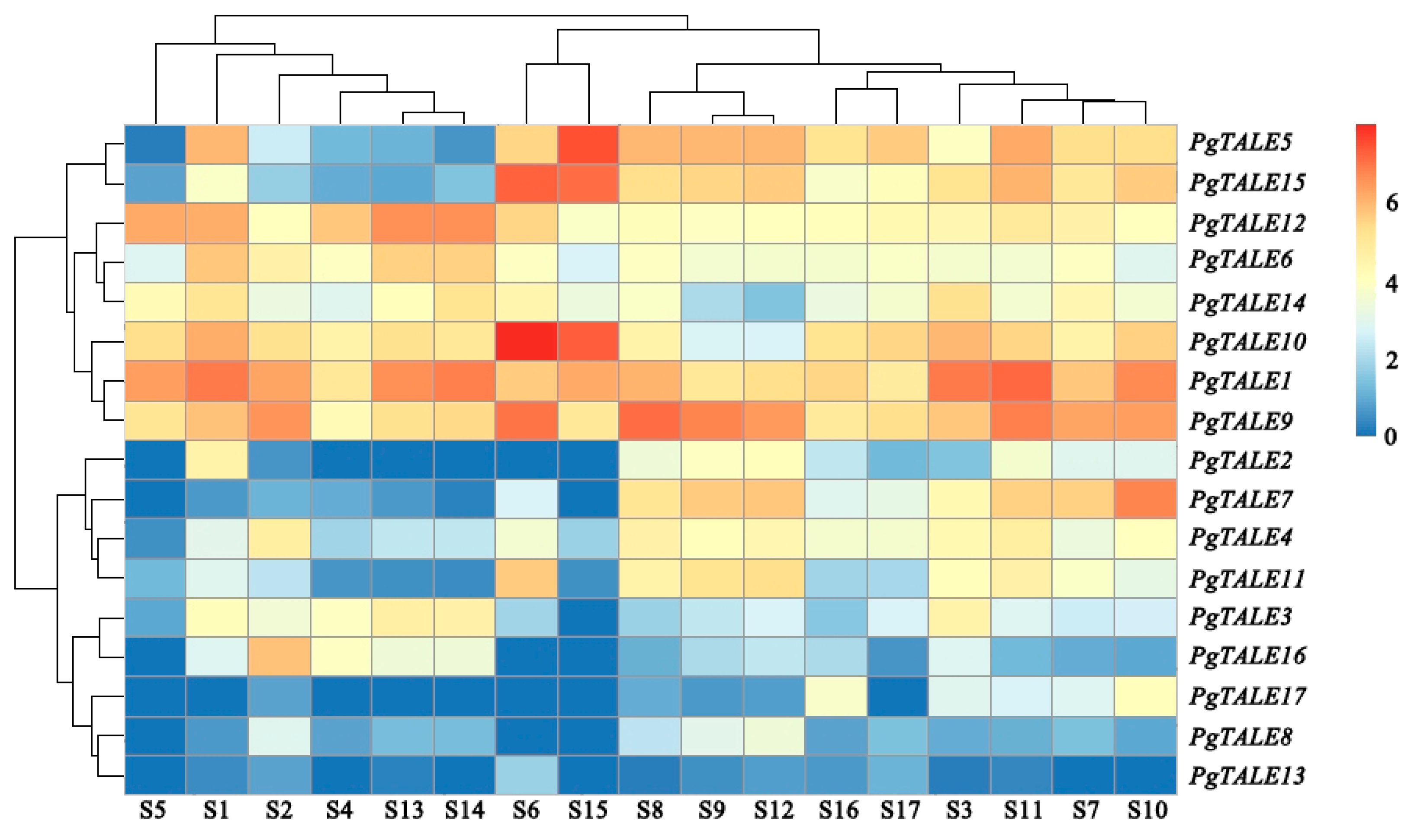
3.6. Expression Analysis of PgTALE Gene Family
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Abbreviations | Full Name |
| TALE | three-amino-acid-loop-extension |
| KNOX/KNAT | KNOTTED-like homeobox |
| BELL/BLH | BEL1-like |
| HB | homeobox |
| ZM-HOX | Zea mays homeobox |
| HAT1 | homeobox from Arabidopsis thaliana 1 |
| HAT2 | homeobox from A. thaliana 2 |
| ATHB8 | A. thaliana homeobox 8 |
| HD-ZIP | homeodomain-leucine zipper |
| PLINC | plant zinc finger |
| WOX | wuschel homeobox |
| PHD | plant homeodomain |
| NDX | nodulin homeobox genes |
| LD | luminidependens |
| OFP | OVATE family protein |
| ATH1 | A. thaliana homeobox 1 |
| STM | shoot meristemless |
| KNAT2 | KNOTTED-like from A. thaliana 2 |
| APG IV | Angiosperm Phylogeny Group IV |
| HOX | homeobox domain |
| SAM | shoot apical meristem |
References
- Di Giacomo, E.; Iannelli, M.A.; Frugis, G. TALE and shape: How to make a leaf different. Plants 2013, 2, 317–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billeter, M.; Qian, Y.Q.; Otting, G.; Müller, M.; Gehring, W.; Wüthrich, K. Determination of the nuclear magnetic resonance solution structure of an Antennapedia homeodomain-DNA complex. J. Mol. Biol. 1993, 234, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- Bharathan, G.; Janssen, B.J.; Kellogg, E.A. Did homeodomain proteins duplicate before the origin of angiosperms, fungi, and metazoa? Proc. Natl. Acad. Sci. USA 1997, 94, 13749–13753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, K.; Brocchieri, L.; Bürglin, T.R. A Comprehensive classification and evolutionary analysis of plant homeobox genes. Mol. Biol. Evol. 2009, 26, 2775–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bürglin, T.R.; Affolter, M. Homeodomain proteins: An update. Chromosoma 2016, 125, 497–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Tian, F.; Yang, D.; Meng, Y.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, N.; Pautot, V.R. Ring the BELL and tie the KNOX: Roles for TALEs in gynoecium development. Front. Plant Sci. 2014, 5, 93. [Google Scholar] [CrossRef]
- Ma, Q.; Wang, N.; Hao, P.; Sun, H.; Yu, S. Genome-wide identification and characterization of TALE superfamily genes in cotton reveals their functions in regulating secondary cell wall biosynthesis. BMC Plant Biol. 2019, 19, 432. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Rosin, F.M.; Prat, S.; Hannapel, D.J. Interacting transcription factors from the three-amino acid loop extension superclass regulate tuber formation. Plant Physiol. 2003, 132, 1391–1404. [Google Scholar] [CrossRef] [Green Version]
- Hay, A.; Tsiantis, M. KNOX genes: Versatile regulators of plant development and diversity. Development 2010, 137, 3153–3165. [Google Scholar] [CrossRef] [Green Version]
- Bertolino, E.; Reimund, B.; Wildtperinic, D.; Clerc, R.G. A novel homeobox protein which recognizes a TGT core and functionally interferes with a retinoid-responsive motif. J. Biol. Chem. 1995, 270, 31178–31188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahajan, A.; Bhogale, S.; Kang, I.H.; Hannapel, D.J.; Banerjee, A.K. The mRNA of a Knotted1-like transcription factor of potato is phloem mobile. Plant Mol. Biol. 2012, 79, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, K.; Ando, S.; Yip, H.K.; Tamada, Y.; Hiwatashi, Y.; Murata, T.; Deguchi, H.; Hasebe, M.; Bowman, J.L. KNOX2 genes regulate the haploid-to-diploid morphological transition in land plants. Science 2013, 339, 1067–1070. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Sharma, P.; Gonzalez, D.H.; Viola, I.L.; Hannapel, D.J. The impact of the long-distance transport of a BEL1-Like Messenger RNA on development. Plant Physiol. 2013, 161, 760–772. [Google Scholar] [CrossRef] [Green Version]
- Furumizu, C.; Alvarez, J.P.; Sakakibara, K.; Bowman, J.L. Antagonistic roles for KNOX1 and KNOX2 genes in patterning the land plant body plan following an ancient gene duplication. PLoS Genet. 2015, 11. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Estévez, M.; Bakkali, M.; Martinblazquez, R.; Garridoramos, M.A. Identification and characterization of TALE homeobox genes in the endangered fern Vandenboschia speciosa. Genes 2017, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- Belles-Boix, E.; Hamant, O.; Witiak, S.M.; Morin, H.; Traas, J.; Pautot, V. KNAT6: An Arabidopsis homeobox gene involved in meristem activity and organ separation. Plant Cell 2006, 18, 1900–1907. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Ishida, T.; Tasaka, M. Shoot apical meristem and cotyledon formation during Arabidopsis embryogenesis: Interaction among the CUP-SHAPED COTYLEDON and SHOOT MERISTEMLESS genes. Development 1999, 126, 1563. [Google Scholar]
- Shani, E.; Yanai, O.; Ori, N. The role of hormones in shoot apical meristem function. Curr. Opin. Plant Biol. 2006, 9, 484–489. [Google Scholar] [CrossRef]
- Cnops, G.; Neyt, P.; Raes, J.; Petrarulo, M.; Nelissen, H.; Malenica, N.; Luschnig, C.; Tietz, O.; Ditengou, F.A.; Palme, K. The TORNADO1 and TORNADO2 genes function in several patterning processes during early leaf development in Arabidopsis thaliana. Plant Cell 2006, 18, 852–866. [Google Scholar] [CrossRef] [Green Version]
- Kondhare, K.R.; Vetal, P.V.; Kalsi, H.S.; Banerjee, A.K. BEL1-like protein (StBEL5) regulates CYCLING DOF FACTOR1 (StCDF1) through tandem TGAC core motifs in potato. J. Plant Physiol. 2019, 241, 153014. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, A.M.; Etchells, J.P.; Canales, C.; Lagodienko, A.; Dickinson, H.G. VAAMANA-a BEL1-like homeodomain protein, interacts with KNOX proteins BP and STM and regulates inflorescence stem growth in Arabidopsis. Gene 2004, 328, 103–111. [Google Scholar] [CrossRef]
- Kim, D.; Cho, Y.; Ryu, H.; Kim, Y.; Kim, T.; Hwang, I. BLH1 and KNAT3 modulate ABA responses during germination and early seedling development in Arabidopsis. Plant J. 2013, 75, 755–766. [Google Scholar] [CrossRef]
- Smith, H.M.; Boschke, I.; Hake, S. Selective interaction of plant homeodomain proteins mediates high DNA-binding affinity. Proc. Natl. Acad. Sci. USA 2002, 99, 9579–9584. [Google Scholar] [CrossRef] [Green Version]
- Brambilla, V.; Battaglia, R.; Colombo, M.; Masiero, S.; Bencivenga, S.; Kater, M.M.; Colombo, L. Genetic and molecular interactions between BELL1 and MADS Box factors support ovule development in Arabidopsis. Plant Cell 2007, 19, 2544–2556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackbusch, J.; Richter, K.; Muller, J.; Salamini, F.; Uhrig, J.F. A central role of Arabidopsis thaliana ovate family proteins in networking and subcellular localization of 3-aa loop extension homeodomain proteins. Proc. Natl. Acad. Sci. USA 2005, 102, 4908–4912. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.E.; Groover, A.; Fontana, J.R.; Martienssen, R.A. Phyllotactic pattern and stem cell fate are determined by the Arabidopsis homeobox gene BELLRINGER. Development 2003, 130, 3941–3950. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Fan, Z.; Zhang, Q.; Wang, C.; Gao, Y.; Deng, Y.; Zhu, B.; Zhu, H.; Chen, J.; Shan, W. BEL1-LIKE HOMEODOMAIN 11 regulates chloroplast development and chlorophyll synthesis in tomato fruit. Plant J. 2018, 94, 1126–1140. [Google Scholar] [CrossRef] [Green Version]
- Rutjens, B.; Bao, D.; Van Eckstouten, E.; Brand, M.; Smeekens, S.; Proveniers, M. Shoot apical meristem function in Arabidopsis requires the combined activities of three BEL1-like homeodomain proteins. Plant J. 2009, 58, 641–654. [Google Scholar] [CrossRef]
- Li, E.; Bhargava, A.; Qiang, W.; Friedmann, M.; Forneris, N.; Savidge, R.; Johnson, L.; Mansfield, S.; Ellis, B.; Douglas, C. The Class II KNOX gene KNAT7 negatively regulates secondary wall formation in Arabidopsis and is functionally conserved in Populus. New Phytol. 2012, 194, 102–115. [Google Scholar] [CrossRef]
- Li, P.; Yu, S.; Shen, J.; Li, Q.; Li, D.; Li, D.; Zheng, C.; Shu, H. The transcriptional response of apple alcohol acyltransferase (MdAAT2) to salicylic acid and ethylene is mediated through two apple MYB TFs in transgenic tobacco. Plant Mol. Biol. 2014, 85, 627–638. [Google Scholar] [CrossRef]
- Magnani, E.; Hake, S. KNOX lost the OX: The Arabidopsis KNATM gene defines a novel class of KNOX transcriptional regulators missing the homeodomain. Plant Cell 2008, 20, 875–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamant, O.; Pautot, V. Plant development: A TALE story. C. R. Biol. 2010, 333, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; Zhou, J.; Mccarthy, R.L.; Ye, Z.H. A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell 2008, 20, 2763–2782. [Google Scholar] [CrossRef] [Green Version]
- Bhargava, A.; Mansfield, S.D.; Hall, H.; Douglas, C.J.; Ellis, B.E. MYB75 functions in regulation of secondary cell wall formation in the Arabidopsis inflorescence stem. Plant Physiol. 2010, 154, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- Li, E.; Wang, S.; Liu, Y.; Chen, J.; Douglas, C.J. OVATE FAMILY PROTEIN4 (OFP4) interaction with KNAT7 regulates secondary cell wall formation in Arabidopsis thaliana. Plant J. 2011, 67, 328–341. [Google Scholar] [CrossRef]
- Byng, J.W.; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Judd, W.S.; Mabberley, D.J.; Sennikov, A.N.; Soltis, D.E.; Soltis, P.S.; Stevens, P.F.; et al. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar]
- Yuan, Z.; Fang, Y.; Zhang, T.; Fei, Z.; Han, F.; Liu, C.; Liu, M.; Xiao, W.; Zhang, W.; Wu, S. The pomegranate (Punica granatum L.) genome provides insights into fruit quality and ovule developmental biology. Plant Biotechnol. J. 2018, 16, 1363–1374. [Google Scholar] [CrossRef] [Green Version]
- Qin, G.; Xu, C.; Ming, R.; Tang, H.; Guyot, R.; Kramer, E.M.; Hu, Y.; Yi, X.; Qi, Y.; Xu, X. The pomegranate (Punica granatum L.) genome and the genomics of punicalagin biosynthesis. Plant J. 2017, 91, 1108–1128. [Google Scholar] [CrossRef] [Green Version]
- Ophir, R.; Sherman, A.; Rubinstein, M.; Eshed, R.; Sharabi Schwager, M.; Harel-Beja, R.; Bar-Ya’Akov, I.; Holland, D. Single-Nucleotide Polymorphism markers from De-Novo assembly of the pomegranate transcriptome reveal germplasm genetic diversity. PLoS ONE 2014, 9, e88998. [Google Scholar] [CrossRef] [Green Version]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; De Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, 597–603. [Google Scholar] [CrossRef]
- Yu, C.; Lin, C.; Hwang, J. Predicting subcellular localization of proteins for Gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. A simple method to control overalignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML -NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 1–4453. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Bodén, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a Toolkit for Biologists integrating various HTS-data handling tools with a user-friendly interface. BioRxiv 2018. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Dhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Peer, Y.V.; Rouz, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, N.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.; Li, H.; Niu, J.; Xue, H.; Liu, B.; Wang, Q.; Luo, X.; Zhang, F.; Zhao, D.; et al. Transcriptomic analysis reveals candidate genes for female sterility in pomegranate flowers. Front Plant Sci. 2017, 8, 1430. [Google Scholar] [CrossRef] [Green Version]
- Ono, N.N.; Britton, M.T.; Fass, J.N.; Nicolet, C.M.; Lin, D.; Tian, L. Exploring the transcriptome landscape of pomegranate fruit peel for natural product biosynthetic gene and SSR marker discovery. J. Integr. Plant. Biol. 2011, 53, 800–813. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, C.; Huang, X.; Zhang, H.; Yuan, Z. Land-plant phylogenomic and pomegranate transcriptomic analyses reveal an evolutionary scenario of CYP75 genes subsequent to whole genome duplications. J. Plant Biol. 2019, 62, 48–60. [Google Scholar] [CrossRef]
- Chen, H.; Banerjee, A.K.; Hannapel, D.J. The tandem complex of BEL and KNOX partners is required for transcriptional repression of ga20ox1. Plant J. 2004, 38, 276–284. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, L.; He, P.; Chang, Y.; Li, L. Identification of genes encoding transcription factors of apple cold-resistant dwarfing rootstock in response to cold stress. Plant Physiol. J. 2017, 53, 1468–1478. [Google Scholar]
- Wen, B.; Song, W.; Sun, M.; Chen, M.; Mu, Q.; Zhang, X.; Wu, Q.; Chen, X.; Gao, D.; Wu, H. Identification and characterization of cherry (Cerasus pseudocerasus G. Don) genes responding to parthenocarpy induced by GA3 through transcriptome analysis. BMC Genet. 2019, 20, 65. [Google Scholar] [CrossRef] [PubMed]
- Qiu, R.; Zhang, T.; Yang, S.; Song, L.; Zhao, D. Genome-wide identification and bioinformatics analysis of TALE transcription factor family in Lotus japonicas. J. Plant Genet. Resour. 2019, 20, 466–475. [Google Scholar]
- Zhao, K.; Zhang, X.; Cheng, Z.; Yao, W.; Li, R.; Jiang, T.; Zhou, B. Comprehensive analysis of the three-amino-acid-loop-extension gene family and its tissue-differential expression in response to salt stress in poplar. Plant Physiol. Bioch. 2019, 136, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hundertmark, M.; Hincha, D.K. LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Liberman, L.M.; Sparks, E.E.; Morenorisueno, M.A.; Petricka, J.J.; Benfey, P.N. MYB36 regulates the transition from proliferation to differentiation in the Arabidopsis root. Proc. Natl. Acad. Sci. USA. 2015, 112, 12099–12104. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.; Robinsonbeers, K.; Ray, S.; Baker, S.C.; Lang, J.D.; Preuss, D.; Milligan, S.B.; Gasser, C.S. Arabidopsis floral homeotic gene BELL (BEL1) controls ovule development through negative regulation of AGAMOUS gene (AG). Proc. Natl. Acad. Sci. USA 1994, 91, 5761–5765. [Google Scholar] [CrossRef] [Green Version]
- Bencivenga, S.; Simonini, S.; Benkova, E.; Colombo, L. The transcription factors BEL1 and SPL are required for cytokinin and auxin signaling during ovule development in Arabidopsis. Plant Cell 2012, 24, 2886–2897. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Mena, C.; Sablowski, R. ARABIDOPSIS THALIANA HOMEOBOX GENE1 establishes the basal boundaries of shoot organs and controls stem growth. Plant Cell 2008, 20, 2059–2072. [Google Scholar] [CrossRef] [Green Version]
- Proveniers, M.; Rutjens, B.; Brand, M.; Smeekens, S. The Arabidopsis TALE homeobox gene ATH1 controls floral competency through positive regulation of FLC. Plant J. 2007, 52, 899–913. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Cultivars | Sample Type | Library | Platform | Reference | Note |
|---|---|---|---|---|---|---|
| SRR5279396 | ‘Dabenzi’ | root | Paired end | Illumina HiSeq 4000 | [39] | |
| SRR5279397 | ‘Dabenzi’ | leaf | Paired end | Illumina HiSeq 4000 | [39] | |
| SRR5279395 | ‘Dabenzi’ | flower | Paired end | Illumina HiSeq 4000 | [39] | |
| SRR5279391 | ‘Dabenzi’ | Inner seed coat (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [39] | |
| SRR5279388 | ‘Dabenzi’ | Outer seed coat (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [39] | |
| SRR5279394 | ‘Dabenzi’ | Pericarp (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [39] | |
| SRR5446598 | ‘Tunisia’ | flower (3.0–5.0 mm) | Paired end | Illumina HiSeq 2500 | [57] | Functional male flower |
| SRR5446595 | ‘Tunisia’ | flower (5.1–13.0 mm) | Paired end | Illumina HiSeq 2500 | [57] | Functional male flower |
| SRR5446592 | ‘Tunisia’ | flower (13.1–25.0 mm) | Paired end | Illumina HiSeq 2500 | [57] | Functional male flower |
| SRR5446607 | ‘Tunisia’ | flower (3.0–5.0 mm) | Paired end | Illumina HiSeq 2500 | [57] | Female sterility |
| SRR5446604 | ‘Tunisia’ | flower (5.1–13.0 mm) | Paired end | Illumina HiSeq 2500 | [57] | Female sterility |
| SRR5446601 | ‘Tunisia’ | flower (13.1–25.0 mm) | Paired end | Illumina HiSeq 2500 | [57] | Female sterility |
| SRR5678820 | ‘Tunisia’ | Inner seed coat (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [39] | |
| SRR5678819 | ‘Baiyushizi’ | Inner seed coat (50 days after pollination) | Paired end | Illumina HiSeq 4000 | [39] | |
| SRR080723 | ‘Wonderful’ | pericarp | Paired end | Illumina HiSeq 2000 | [58] | |
| SRR1055290 | ‘nana’ | Mixed samples of leaves, flowers, fruit and roots | Single end | 454 GS FLX Titanium | [40] | |
| SRR1054190 | ‘Black127’ | Mixed samples of root, leaf, flower and fruit | Single end | 454 GS FLX Titanium | [40] |
| Gene Name | Gene ID | Location | ExonNo. | CDS | AA | MW(Da) | pI | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|
| PgTALE1 | Pg002952.1 | scaffold11:2366550:2369265 | 4 | 2097 | 698 | 75,172.93 | 7.80 | −0.538 | Nuclear |
| PgTALE2 | Pg005682.1 | scaffold13:4182457:4188943 | 5 | 1035 | 344 | 38,810.15 | 5.25 | −0.795 | Nuclear |
| PgTALE3 | Pg009001.1 | scaffold17:1307284:1312800 | 4 | 1959 | 652 | 70,713.74 | 6.51 | −0.557 | Nuclear |
| PgTALE4 | Pg011533.1 | scaffold2:5274088:5277780 | 4 | 1932 | 643 | 70,086.66 | 6.33 | −0.566 | Nuclear |
| PgTALE5 | Pg014946.1 | scaffold26:1035323:1039874 | 5 | 1065 | 354 | 40,403.90 | 6.15 | −0.969 | Nuclear |
| PgTALE6 | Pg015766.1 | scaffold29:588196:590712 | 4 | 1905 | 634 | 70,031.46 | 5.87 | −0.717 | Nuclear |
| PgTALE7 | Pg022248.1 | scaffold49:1377908:1386181 | 5 | 1065 | 354 | 39,800.27 | 5.13 | −0.751 | Nuclear |
| PgTALE8 | Pg024529.1 | scaffold6:3685906:3688228 | 4 | 1563 | 520 | 58,385.62 | 6.10 | −0.449 | Nuclear |
| PgTALE9 | Pg024817.1 | scaffold6:2019598:2022376 | 5 | 1275 | 424 | 46,087.05 | 5.94 | −0.635 | Nuclear |
| PgTALE10 | Pg026506.1 | scaffold7:1735075:1737746 | 4 | 2103 | 700 | 76,325.21 | 7.82 | −0.647 | Nuclear |
| PgTALE11 | Pg027513.1 | scaffold73:612910:617019 | 4 | 1023 | 340 | 37,775.80 | 6.53 | −0.534 | Nuclear |
| PgTALE12 | Pg028434.1 | scaffold8:2877271:2880879 | 5 | 909 | 302 | 33,829.16 | 6.27 | −0.635 | Nuclear |
| PgTALE13 | Pg028770.1 | scaffold81:128287:132801 | 5 | 2136 | 711 | 78,483.78 | 6.54 | −0.513 | Nuclear |
| PgTALE14 | Pg029909.1 | scaffold9:754612:757823 | 4 | 1851 | 616 | 68,481.09 | 6.27 | −0.707 | Nuclear |
| PgTALE15 | Pg030082.1 | scaffold9:3730507:3733807 | 6 | 1203 | 400 | 45,378.59 | 5.30 | −0.730 | Nuclear |
| PgTALE16 | Pg030621.1 | scaffold96:200629:218877 | 6 | 2400 | 799 | 87,381.25 | 8.78 | −0.549 | Nuclear |
| PgTALE17 | Pg005241.1 | scaffold13:2266264:2268869 | 3 | 465 | 154 | 17,605.66 | 5.18 | −0.730 | Nuclear |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhao, Y.; Yan, M.; Zhao, H.; Zhang, X.; Yuan, Z. Genome-wide Identification and Expression Analysis of TALE Gene Family in Pomegranate (Punica granatum L.). Agronomy 2020, 10, 829. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060829
Wang Y, Zhao Y, Yan M, Zhao H, Zhang X, Yuan Z. Genome-wide Identification and Expression Analysis of TALE Gene Family in Pomegranate (Punica granatum L.). Agronomy. 2020; 10(6):829. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060829
Chicago/Turabian StyleWang, Yuying, Yujie Zhao, Ming Yan, Honglian Zhao, Xinhui Zhang, and Zhaohe Yuan. 2020. "Genome-wide Identification and Expression Analysis of TALE Gene Family in Pomegranate (Punica granatum L.)" Agronomy 10, no. 6: 829. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy10060829