
Identification of QTL for Tolerance to Flooding Stress at Seedling Stage of Soybean (Glycine max L. Merr.)

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotyping
2.3. DNA Extraction and Genotyping
2.4. Data Analysis
2.5. Linkage Mapping and QTL Analysis
2.6. Screening of Candidate Genes
3. Results
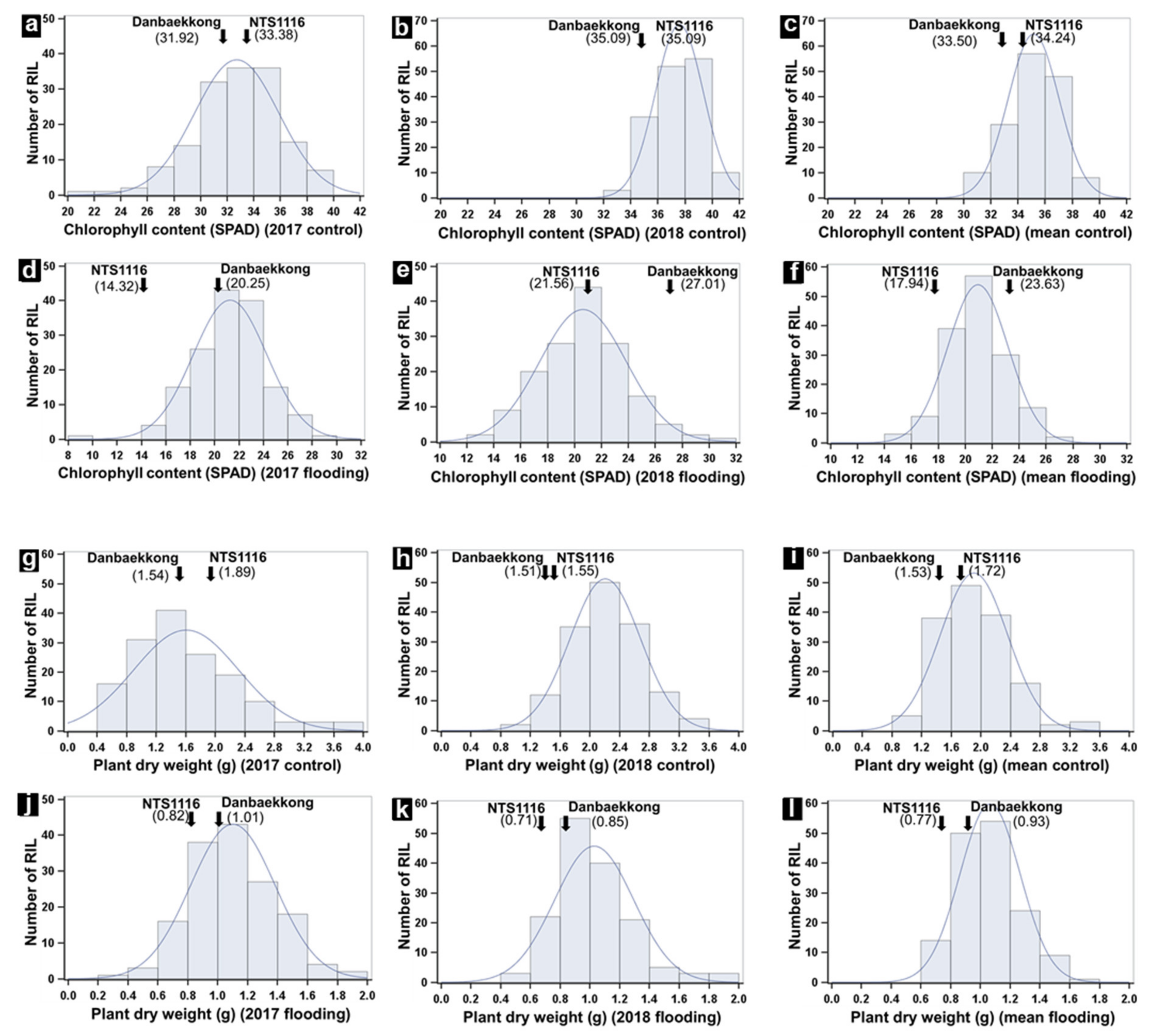
3.1. Phenotypic Variation and Correlation between Traits
3.2. Linkage Mapping
3.3. QTL Analysis under Flooding and Index (CCI, DWI, and FTI)
3.4. Screening of Candidate Genes in the QTL Hotspots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Najafi, E.; Devineni, N.; Khanbilvardi, R.M.; Kogan, F. Understanding the Changes in Global Crop Yields Through Changes in Climate and Technology. Earth’s Futur. 2018, 6, 410–427. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Fukao, T.; Gibbs, D.J.; Holdsworth, M.J.; Lee, S.C.; Licausi, F.; Perata, P.; Voesenek, L.A.C.J.; van Dongen, J.T. Making sense of low oxygen sensing. Trends Plant Sci. 2012, 17, 129–138. [Google Scholar] [CrossRef]
- Shanmugam, S.; Zhao, S.; Nandy, S.; Srivastava, V.; Khodakovskaya, M. Modification of soybean growth and abiotic stress tolerance by expression of truncated ERECTA protein from Arabidopsis thaliana. PLoS ONE 2020, 15, e0233383. [Google Scholar] [CrossRef]
- Qin, D.; Liu, X.-Y.; Miceli, C.; Zhang, Q.; Wang, P. Soybean plants expressing the Bacillus thuringiensis cry8-like gene show resistance to Holotrichia parallela. BMC Biotechnol. 2019, 19, 66. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, J.; Alagarasan, G.; Savitha, P.; Lydia, K.; Pothiraj, G.; Vijayakumar, E.; Sudhagar, R.; Singh, A.; Vedna, K.; Vanniarajan, C. Improved host-plant resistance to Phytophthora rot and powdery mildew in soybean (Glycine max (L.) Merr.). Sci. Rep. 2020, 10, 13928. [Google Scholar] [CrossRef] [PubMed]
- Valliyodan, B.; Van Toai, T.T.; Alves, J.D.; de Fátima, P.; Goulart, P.; Lee, J.D.; Fritschi, F.B.; Rahman, M.A.; Islam, R.; Shannon, J.G.; et al. Expression of Root-Related Transcription Factors Associated with Flooding Tolerance of Soybean (Glycine max). Int. J. Mol. Sci. 2014, 15, 17622–17643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.; Song, L.; Chen, H.; Valliyodan, B.; Cheng, P.; Ali, L.; Vuong, T.; Wu, C.; Orlowski, J.; Buckley, B.; et al. A major natural genetic variation associated with root system architecture and plasticity improves waterlogging tolerance and yield in soybean. Plant Cell Environ. 2018, 41, 2169–2182. [Google Scholar] [CrossRef]
- Hou, F.F.; Thseng, F.S. Studies on the flooding tolerance of soybean seed: Varietal differences. Euphytica 1991, 57, 169–173. [Google Scholar] [CrossRef]
- Wu, C.; Chen, P.; Hummer, W.; Zeng, A.; Klepadlo, M. Effect of Flood Stress on Soybean Seed Germination in the Field. Am. J. Plant Sci. 2017, 8, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Linkemer, G.; Board, J.E.; Musgrave, M.E. Waterlogging Effects on Growth and Yield Components in Late-Planted Soybean. Crop. Sci. 1998, 38, 1576–1584. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Vuong, T.D.; VanToai, T.; Lee, J.D.; Wu, X.; Mian, M.A.R.; Dorrance, A.E.; Shannon, J.G.; Nguyen, H.T. Mapping of Quantitative Trait Loci Associated with Resistance toPhytophthora sojaeand Flooding Tolerance in Soybean. Crop. Sci. 2012, 52, 2481–2493. [Google Scholar] [CrossRef]
- Githiri, S.M.; Watanabe, S.; Harada, K.; Takahashi, R. QTL analysis of flooding tolerance in soybean at an early vegetative growth stage. Plant Breed. 2006, 125, 613–618. [Google Scholar] [CrossRef]
- Kokubun, M. Genetic and cultural improvement of soybean for waterlogged conditions in Asia. Field Crop. Res. 2013, 152, 3–7. [Google Scholar] [CrossRef]
- Patil, P.K.; Singh, A.K.; Tripathi, N.; Yadav, D.; Hemantaranjan, A. Flooding: Abiotic Constraint Limiting Vegetable Productivity. Adv. Plants Agric. Res. 2014, 1, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Henshaw, T.L.; Gilbert, R.A.; Scholberg, J.M.S.; Sinclair, T.R. Soya Bean (Glycine max L. Merr.) Genotype Response to Early-season Flooding: I. Root and Nodule Development. J. Agron. Crop. Sci. 2007, 193, 177–188. [Google Scholar] [CrossRef]
- Maekawa, T.; Shimamura, S.; Shimada, S. Effects of Short-Term Waterlogging on Soybean Nodule Nitrogen Fixation at Different Soil Reductions and Temperatures. Plant Prod. Sci. 2011, 14, 349–358. [Google Scholar] [CrossRef]
- Arrese-Igor, C.; Royuela, M.; Aparicio-Tejo, P.M. Responses of Nodulated Legumes to Oxygen Deficiency. In Interacting Stresses on Plants in a Changing Climate; Jackson, M.B., Black, C.R., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; pp. 423–432. ISBN 978-3-642-78535-1. [Google Scholar]
- Oliveira, H.C.; Sodek, L. Effect of oxygen deficiency on nitrogen assimilation and amino acid metabolism of soybean root segments. Amino Acids 2013, 44, 743–755. [Google Scholar] [CrossRef]
- Oliveira, H.C.; Freschi, L.; Sodek, L. Nitrogen metabolism and translocation in soybean plants subjected to root oxygen deficiency. Plant Physiol. Biochem. 2013, 66, 141–149. [Google Scholar] [CrossRef]
- Limami, A.M.; Diab, H.; Lothier, J. Nitrogen metabolism in plants under low oxygen stress. Planta 2014, 239, 531–541. [Google Scholar] [CrossRef]
- Boru, G.; VanToai, T.; Alves, J.; Hua, D.; Knee, M. Responses of Soybean to Oxygen Deficiency and Elevated Root-zone Carbon Dioxide Concentration. Ann. Bot. 2003, 91, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Araki, H. Water uptake of soybean (Glycine max L. Merr.) during exposure to O2 deficiency and field level CO2 concentration in the root zone. Field Crop. Res. 2006, 96, 98–105. [Google Scholar] [CrossRef]
- Chen, W.; Yao, Q.; Patil, G.B.; Agarwal, G.; Deshmukh, R.K.; Lin, L.; Wang, B.; Wang, Y.; Prince, S.J.; Song, L.; et al. Identification and Comparative Analysis of Differential Gene Expression in Soybean Leaf Tissue under Drought and Flooding Stress Revealed by RNA-Seq. Front. Plant Sci. 2016, 7, 1044. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M. Improvement of Plant Waterlogging Tolerance. In Waterlogging Signalling and Tolerance in Plants; Mancuso, S., Shabala, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 267–285. ISBN 978-3-642-10304-9. [Google Scholar]
- VanToai, T.T.; St. Martin, S.K.; Chase, K.; Boru, G.; Schnipke, V.; Schmitthenner, A.F.; Lark, K.G. Identification of a QTL Associated with Tolerance of Soybean to Soil Waterlogging. Crop. Sci. 2001, 41, 1247–1252. [Google Scholar] [CrossRef] [Green Version]
- Reyna, N.; Cornelious, B.; Shannon, J.G.; Sneller, C.H. Evaluation of a QTL for Waterlogging Tolerance in Southern Soybean Germplasm. Crop. Sci. 2003, 43, 2077–2082. [Google Scholar] [CrossRef]
- Cornelious, B.; Chen, P.; Chen, Y.; De Leon, N.; Shannon, J.G.; Wang, D. Identification of QTLs Underlying Water-Logging Tolerance in Soybean. Mol. Breed. 2005, 16, 103–112. [Google Scholar] [CrossRef]
- Sayama, T.; Nakazaki, T.; Ishikawa, G.; Yagasaki, K.; Yamada, N.; Hirota, N.; Hirata, K.; Yoshikawa, T.; Saito, H.; Teraishi, M.; et al. QTL analysis of seed-flooding tolerance in soybean (Glycine max [L.] Merr.). Plant Sci. 2009, 176, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Salekdeh, G.H.; Reynolds, M.; Bennett, J.; Boyer, J. Conceptual framework for drought phenotyping during molecular breeding. Trends Plant Sci. 2009, 14, 488–496. [Google Scholar] [CrossRef]
- Berger, B.; Parent, B.; Tester, M. High-throughput shoot imaging to study drought responses. J. Exp. Bot. 2010, 61, 3519–3528. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.-W.; Ji, H.C.; Yamakawa, T. Comparison of Photosynthetic Response of Two Soybean Cultivars to Soil Flooding. J. Fac. Agric. Kyushu Univ. 2006, 51, 227–232. [Google Scholar] [CrossRef]
- Shimono, H.; Konno, T.; Sakai, H.; Sameshima, R. Interactive Effects of Elevated Atmospheric CO2 and Waterlogging on Vegetative Growth of Soybean (Glycine max (L.) Merr.). Plant Prod. Sci. 2012, 15, 238–245. [Google Scholar] [CrossRef]
- Hummer, W.S. Linkage Mapping for Soybean (Glycine max) Flood Tolerance. Master’s Thesis, University of Arkansas, Fayetteville, NC, USA, 2018. [Google Scholar]
- Dhungana, S.K.; Kim, H.-S.; Kang, B.-K.; Seo, J.-H.; Kim, H.-T.; Shin, S.-O.; Park, C.-H.; Kwak, D.-Y. Quantitative trait loci mapping for flooding tolerance at an early growth stage of soybean recombinant inbred line population. Plant Breed. 2020, 139, 626–638. [Google Scholar] [CrossRef]
- Soltani, A.; MafiMoghaddam, S.; Oladzad-Abbasabadi, A.; Walter, K.; Kearns, P.J.; Vasquez-Guzman, J.; Mamidi, S.; Lee, R.; Shade, A.L.; Jacobs, J.L.; et al. Genetic Analysis of Flooding Tolerance in an Andean Diversity Panel of Dry Bean (Phaseolus vulgaris L.). Front. Plant Sci. 2018, 9, 767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Chen, G.-Y. Conditional QTL mapping for waterlogging tolerance in two RILs populations of wheat. SpringerPlus 2013, 2, 245. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Mao, S.; Chen, G.; Liu, Y.; Li, W.; Wei, Y.; Liu, C.; Zheng, Y. QTLs for Waterlogging Tolerance at Germination and Seedling Stages in Population of Recombinant Inbred Lines Derived from a Cross Between Synthetic and Cultivated Wheat Genotypes. J. Integr. Agric. 2014, 13, 31–39. [Google Scholar] [CrossRef]
- Ballesteros, D.C.; Mason, R.E.; Addison, C.K.; Acuña, M.A.; Arguello, M.N.; Subramanian, N.; Miller, R.G.; Sater, H.; Gbur, E.E.; Miller, D.; et al. Tolerance of wheat to vegetative stage soil waterlogging is conditioned by both constitutive and adaptive QTL. Euphytica 2015, 201, 329–343. [Google Scholar] [CrossRef]
- Li, H.; Vaillancourt, R.; Mendham, N.; Zhou, M. Comparative mapping of quantitative trait loci associated with waterlogging tolerance in barley (Hordeum vulgare L.). BMC Genom. 2008, 9, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.Y.; Wu, P.; Hu, B.; Yi, K.K. Effects of genetic background and environment on QTLs and epistasis for rice (Oryza sativa L.) panicle number. Theor. Appl. Genet. 2001, 103, 104–111. [Google Scholar] [CrossRef]
- Xie, X.-W.; Xu, M.-R.; Zang, J.-P.; Sun, Y.; Zhu, L.-H.; Xu, J.-L.; Zhou, Y.-L.; Li, Z.-K. Genetic Background and Environmental Effects on QTLs for Sheath Blight Resistance Revealed by Reciprocal Introgression Lines in Rice. Acta Agron. Sin. 2008, 34, 1885–1893. [Google Scholar] [CrossRef]
- Koo, S.C.; Kim, H.T.; Kang, B.K.; Lee, Y.H.; Oh, K.W.; Baek, I.Y.; Yun, H.T.; Choi, M.S.; Kim, H.Y. Screening of Flooding Tolerance in Soybean Germplasm Collection. Korean J. Breed. Sci. 2014, 46, 129–135. [Google Scholar] [CrossRef]
- Lee, C.; Choi, M.-S.; Kim, H.-T.; Yun, H.-T.; Lee, B.; Chung, Y.-S.; Kim, R.W.; Choi, H.-K. Soybean [Glycine max (L.) Merrill]: Importance as A Crop and Pedigree Reconstruction of Korean Varieties. Plant Breed. Biotechnol. 2015, 3, 179–196. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Kim, K.H.; Van, K.; Kim, M.Y.; Lee, S.-H. Fine mapping of a resistance gene to bacterial leaf pustule in soybean. Theor. Appl. Genet. 2010, 120, 1443–1450. [Google Scholar] [CrossRef]
- Hong, E.H.; Kim, S.D.; Hwang, Y.H.; Lee, Y.H.; Moon, Y.H.; Park, E.H.; Park, R.K. A Semi-Dwarf, High Density-Adaptable and High Yielding Soybean Variety “Paldalkong. ” Res. Rep. Rural Dev. Adm. Crop. 1987, 29, 203–209. [Google Scholar]
- Lee, Y.-G.; Jeong, N.; Kim, J.H.; Lee, K.; Kim, K.H.; Pirani, A.; Ha, B.-K.; Kang, S.-T.; Park, B.-S.; Moon, J.-K.; et al. Development, validation and genetic analysis of a large soybean SNP genotyping array. Plant J. 2015, 81, 625–636. [Google Scholar] [CrossRef]
- Toker, C. Estimates of broad-sense heritability for seed yield and yield criteria in faba bean (Vicia faba L.). Hereditas 2004, 140, 222–225. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop. J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, H.; Zhang, L.; Meng, L. Users’ Manual of QTL IciMapping; The Quantitative Genetics Group, Institute of Crop Science, Chinese Academy of Agricultural Sciences (CAAS): Beijing, China; Genetic Resources Program, International Maize and Wheat Improvement Center (CIMMYT): Mexico City, Mexico, 2016. [Google Scholar]
- Wu, C.; Zeng, A.; Chen, P.; Florez-Palacios, L.; Hummer, W.; Mokua, J.; Klepadlo, M.; Yan, L.; Ma, Q.; Cheng, Y. An effective field screening method for flood tolerance in soybean. Plant Breed. 2017, 136, 710–719. [Google Scholar] [CrossRef]
- Wray, N.R.; Visscher, P.M. Estimating Trait Heritability. Nat. Educ. 2008, 1, 29. [Google Scholar]
- Djanaguiraman, M.; Prasad, P.V.; Boyle, D.L.; Schapaugh, W.T. High-Temperature Stress and Soybean Leaves: Leaf Anatomy and Photosynthesis. Crop. Sci. 2011, 51, 2125–2131. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Archer, M.A.; Wayne, R.K. Transgressive segregation, adaptation and speciation. Heredity 1999, 83, 363–372. [Google Scholar] [CrossRef]
- Wu, C.; Mozzoni, L.A.; Moseley, D.; Hummer, W.; Ye, H.; Chen, P.; Shannon, G.; Nguyen, H. Genome-wide association mapping of flooding tolerance in soybean. Mol. Breed. 2020, 40, 4. [Google Scholar] [CrossRef]
- Nguyen, H.T.M.; Neelakadan, A.K.; Quach, T.N.; Valliyodan, B.; Kumar, R.; Zhang, Z.; Nguyen, H.T. Molecular characterization of Glycine max squalene synthase genes in seed phytosterol biosynthesis. Plant Physiol. Biochem. 2013, 73, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Huang, Y.; Zhu, N.; Zhao, Y. The rice WUSCHEL-related homeobox genes are involved in reproductive organ development, hormone signaling and abiotic stress response. Gene 2014, 549, 266–274. [Google Scholar] [CrossRef]
- Ma, J.; Li, M.-Y.; Wang, F.; Tang, J.; Xiong, A.-S. Genome-wide analysis of Dof family transcription factors and their responses to abiotic stresses in Chinese cabbage. BMC Genom. 2015, 16, 33. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Chen, G.; Dong, T.; Wang, L.; Zhang, J.; Zhao, Z.; Hu, Z. SlDEAD31, a Putative DEAD-Box RNA Helicase Gene, Regulates Salt and Drought Tolerance and Stress-Related Genes in Tomato. PLoS ONE 2015, 10, e0133849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Wang, P.; Hou, L.; Zhao, S.; Zhao, C.; Xia, H.; Li, P.; Zhang, Y.; Bian, X.; Wang, X. Global Analysis of WRKY Genes and Their Response to Dehydration and Salt Stress in Soybean. Front. Plant Sci. 2016, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Li, R.; Dai, Y.; Yuan, L.; Sun, Q.; Zhang, S.; Wang, X. A B-box zinc finger protein, MdBBX10, enhanced salt and drought stresses tolerance in Arabidopsis. Plant Mol. Biol. 2019, 99, 437–447. [Google Scholar] [CrossRef]
- Guo, R.; Qiao, H.; Zhao, J.; Wang, X.; Tu, M.; Guo, C.; Wan, R.; Li, Z.; Wang, X. The Grape VlWRKY3 Gene Promotes Abiotic and Biotic Stress Tolerance in Transgenic Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 545. [Google Scholar] [CrossRef] [Green Version]
- Dossa, K.; Mmadi, M.A.; Zhou, R.; Zhang, T.; Su, R.; Zhang, Y.; Wang, L.; You, J.; Zhang, X. Depicting the Core Transcriptome Modulating Multiple Abiotic Stresses Responses in Sesame (Sesamum indicum L.). Int. J. Mol. Sci. 2019, 20, 3930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carther, K.; Ketehouli, T.; Ye, N.; Yang, Y.-H.; Wang, N.; Dong, Y.-Y.; Yao, N.; Liu, X.-M.; Liu, W.-C.; Li, X.-W.; et al. Comprehensive Genomic Analysis and Expression Profiling of Diacylglycerol Kinase (DGK) Gene Family in Soybean (Glycine max) under Abiotic Stresses. Int. J. Mol. Sci. 2019, 20, 1361. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.D.; Nguyen, K.-L.; Watanabe, Y.; Le, D.T.; Tran, L.-S.P. Expression analyses of soybean genes encoding methionine-R-sulfoxide reductase under various conditions suggest a possible role in the adaptation to stress. Appl. Biol. Chem. 2016, 59, 681–687. [Google Scholar] [CrossRef]
- Sun, X.; Sun, M.; Jia, B.; Qin, Z.; Yang, K.; Chen, C.; Yu, Q.; Zhu, Y. A Glycine soja methionine sulfoxide reductase B5a interacts with the Ca2+/CAM-binding kinase GsCBRLK and activates ROS signaling under carbonate alkaline stress. Plant J. 2016, 86, 514–529. [Google Scholar] [CrossRef] [Green Version]
- Rey, P.; Tarrago, L. Physiological Roles of Plant Methionine Sulfoxide Reductases in Redox Homeostasis and Signaling. Antioxidants 2018, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- Dhungana, S.K.; Kim, H.-S.; Kang, B.-K.; Seo, J.-H.; Kim, H.-T.; Oh, J.-H.; Shin, S.-O.; Baek, I.-Y. Analysis of differentially expressed genes in soybean leaf tissue of tolerant and susceptible cultivars under flooding stress revealed by RNA sequencing. J. Crop. Sci. Biotechnol. 2021, 24, 83–91. [Google Scholar] [CrossRef]
- Deshmukh, R.; Sonah, H.; Patil, G.; Chen, W.; Prince, S.; Mutava, R.; Vuong, T.; Valliyodan, B.; Nguyen, H.T. Integrating omic approaches for abiotic stress tolerance in soybean. Front. Plant Sci. 2014, 5, 244. [Google Scholar] [CrossRef] [PubMed]
- Sareen, S.; Tyagi, B.S.; Sarial, A.K.; Tiwari, V.; Sharma, I. Trait analysis, diversity, and genotype × environment interaction in some wheat landraces evaluated under drought and heat stress conditions. Chil. J. Agric. Res. 2014, 74, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated Responses to Oxygen and Sugar Deficiency Allow Rice Seedlings to Tolerate Flooding. Sci. Signal. 2009, 2, ra61. [Google Scholar] [CrossRef]
- Gonzali, S.; Loreti, E.; Cardarelli, F.; Novi, G.; Parlanti, S.; Pucciariello, C.; Bassolino, L.; Banti, V.; Licausi, F.; Perata, P. Universal stress protein HRU1 mediates ROS homeostasis under anoxia. Nat. Plants 2015, 1, 15151. [Google Scholar] [CrossRef] [PubMed]
- Mendiondo, G.M.; Gibbs, D.J.; Szurman-Zubrzycka, M.; Korn, A.; Marquez, J.; Szarejko, I.; Maluszynski, M.; King, J.; Axcell, B.; Smart, K.; et al. Enhanced waterlogging tolerance in barley by manipulation of expression of the N-end rule pathway E3 ligase PROTEOLYSIS6. Plant Biotechnol. J. 2016, 14, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Komatsu, S. Characterization of proteins in soybean roots under flooding and drought stresses. J. Proteom. 2015, 114, 161–181. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Treatment | Trait | Year | Parent | RIL | H2 1 | ||
|---|---|---|---|---|---|---|---|
| Danbaekkong (Mean ± SD) | NTS1116 (Mean ± SD) | Mean | Range | ||||
| Control | CC | 2017 | 31.92 ± 5.11 | 33.38 ± 5.26 | 32.55 | 21.00–39.58 | |
| 2018 | 35.09 ± 2.54 | 35.09 ± 2.22 | 37.60 | 33.04–41.27 | |||
| Mean 2 | 33.50 ± 2.24 | 34.24 ± 1.21 | 35.24 | 21.00–41.27 | |||
| DW (g) | 2017 | 1.54 ± 0.71 | 1.89 ± 1.64 | 1.60 | 0.49–3.80 | ||
| 2018 | 1.51 ± 0.52 | 1.55 ± 0.32 | 2.21 | 0.88–3.44 | |||
| Mean | 1.53 ± 0.02 | 1.72 ± 0.24 | 1.92 | 0.49–3.80 | |||
| Flooding | CC | 2017 | 20.25 ± 7.33 | 14.32 ± 2.42 | 20.67 | 9.80–29.38 | |
| 2018 | 27.01 ± 2.86 | 21.56 ± 2.97 | 20.70 | 12.64–30.04 | |||
| Mean | 23.63 ± 4.78 | 17.94 ± 5.12 | 20.73 | 9.80–30.04 | |||
| DW (g) | 2017 | 1.01 ± 0.59 | 0.82 ± 0.42 | 1.10 | 0.31–1.87 | ||
| 2018 | 0.85 ± 0.05 | 0.71 ± 0.16 | 1.03 | 0.48–1.90 | |||
| Mean | 0.93 ± 0.11 | 0.77 ± 0.09 | 1.06 | 0.31–1.90 | |||
| Combined 3 | CC (%) | 68.35 | |||||
| DW (%) | 68.42 | ||||||
| Trait | Year | Parent | RIL | ||
|---|---|---|---|---|---|
| Danbaekkong | NTS1116 | Mean | Range | ||
| CCI | 2017 | 0.63 | 0.43 | 0.66 | 0.29–0.97 |
| 2018 | 0.77 | 0.61 | 0.55 | 0.35–0.80 | |
| Mean 1 | 0.70 | 0.52 | 0.60 | 0.29–0.97 | |
| DWI | 2017 | 0.66 | 0.43 | 0.79 | 0.25–2.00 |
| 2018 | 0.56 | 0.46 | 0.47 | 0.27–0.89 | |
| Mean | 0.61 | 0.45 | 0.63 | 0.25–2.00 | |
| FTI | 2017 | 0.65 | 0.43 | 0.72 | 0.30–1.39 |
| 2018 | 0.67 | 0.54 | 0.51 | 0.35–0.75 | |
| Mean | 0.66 | 0.48 | 0.62 | 0.30–1.39 | |
| QTL Name 1 | Environment 2 | Chr (LG) 3 | Position (cM) | Marker Interval | Physical Position 4 (bp) | LOD 5 | PVE 6 (%) | Additive 7 |
|---|---|---|---|---|---|---|---|---|
| qSFT_3-38 | 2018DWI | 3 (N) | 36.8−40.9 | AX-90428828–AX-90340803 | 37,447,696–38,227,564 | 5.14 | 11.8 | −0.0509 |
| qSFT_3-64 | 2018DWI | 3 (N) | 62.8−71.3 | AX-90323988–AX-90489054 | 42,201,780–43,325,752 | 3.39 | 8.3 | 0.0314 |
| qSFT_4-17 | 2017CCF | 4 (C1) | 267.5−290.6 | AX-90368307–AX-90363550 | 6,322,051–41,712,698 | 3.51 | 8.2 | −0.8676 |
| qSFT_6-86 | 2017DWI | 6 (C2) | 200.5−215.2 | AX-90332662–AX-90454533 | 18,319,783–47,839,535 | 4.55 | 10.4 | −0.1081 |
| 2017FTI | 200.5−215.2 | AX-90332662–AX-90454533 | 18,319,783–47,839,535 | 4.50 | 10.2 | −0.064 | ||
| qSFT_7-3 | 2017FTI | 7 (M) | 77.9−115.2 | AX-90495740–AX-90333525 | 418,570–7,905,015 | 3.98 | 30.7 | −0.1081 |
| 2017DWI | 77.9−115.2 | AX-90495740–AX-90333525 | 418,570–7,905,015 | 4.10 | 25.6 | −0.1656 | ||
| MeanDWI | 114.8−115.2 | AX-90362660–AX-90333525 | 7,854,359–7,905,015 | 3.89 | 8.9 | −0.0546 | ||
| qSFT_7-14 | 2017FTI | 116.3−134.5 | AX-90316460–AX-90415397 | 7,984,980–15,641,186 | 4.22 | 12.8 | −0.0701 | |
| MeanFTI | 7 (M) | 121.2−121.9 | AX-90474615–AX-90329820 | 9,874,757–10,187,432 | 4.63 | 10.4 | −0.0351 | |
| qSFT_13-53 | 2018CCF | 13 (F) | 66.1−72.6 | AX-90380240–AX-90439239 | 26,719,782–30,966,459 | 3.64 | 8.1 | −0.9293 |
| qSFT_15-67 | 2018CCF | 15 (E) | 125.3−140.9 | AX-90306554–AX-90488715 | 6,048,170–8,976,050 | 4.70 | 10.6 | −1.1020 |
| MeanCCI | 129.5−131.3 | AX-90436179–AX-90424048 | 7,732,934–8,240,500 | 3.63 | 8.6 | −0.0223 | ||
| qSFT_16-40 | 2018CCI | 16 (J) | 43.8−76.3 | AX-90320765–AX-90450566 | 26,236,346–35,344,890 | 3.85 | 9.7 | −0.0274 |
| qSFT_16-62 | 2018CCF | 16 (J) | 83.7−87.2 | AX-90329869–AX-90316894 | 36,148,023–36,625,433 | 3.78 | 8.9 | −0.9824 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhungana, S.K.; Kim, H.-S.; Kang, B.-K.; Seo, J.-H.; Kim, H.-T.; Shin, S.-O.; Oh, J.-H.; Baek, I.-Y. Identification of QTL for Tolerance to Flooding Stress at Seedling Stage of Soybean (Glycine max L. Merr.). Agronomy 2021, 11, 908. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050908
Dhungana SK, Kim H-S, Kang B-K, Seo J-H, Kim H-T, Shin S-O, Oh J-H, Baek I-Y. Identification of QTL for Tolerance to Flooding Stress at Seedling Stage of Soybean (Glycine max L. Merr.). Agronomy. 2021; 11(5):908. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050908
Chicago/Turabian StyleDhungana, Sanjeev Kumar, Hong-Sik Kim, Beom-Kyu Kang, Jeong-Hyun Seo, Hyun-Tae Kim, Sang-Ouk Shin, Jae-Hyeon Oh, and In-Youl Baek. 2021. "Identification of QTL for Tolerance to Flooding Stress at Seedling Stage of Soybean (Glycine max L. Merr.)" Agronomy 11, no. 5: 908. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11050908