Relationships between Nutrient Uptake and Nitrogen Fixation with Aflatoxin Contamination in Peanut under Terminal Drought


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Materials
2.2. Crop Management
2.3. Irrigation
2.4. Aspergillus flavus Inoculation
2.5. Data Collection
2.5.1. Soil and Plant Water Status
2.5.2. Plant Nutrient Analysis
2.5.3. Nodule Dry Weight (NDW) and Nitrogen Fixation (NF)
2.5.4. Aspergillus flavus and Aflatoxin Measurements
2.5.5. Statistical Analysis
3. Results
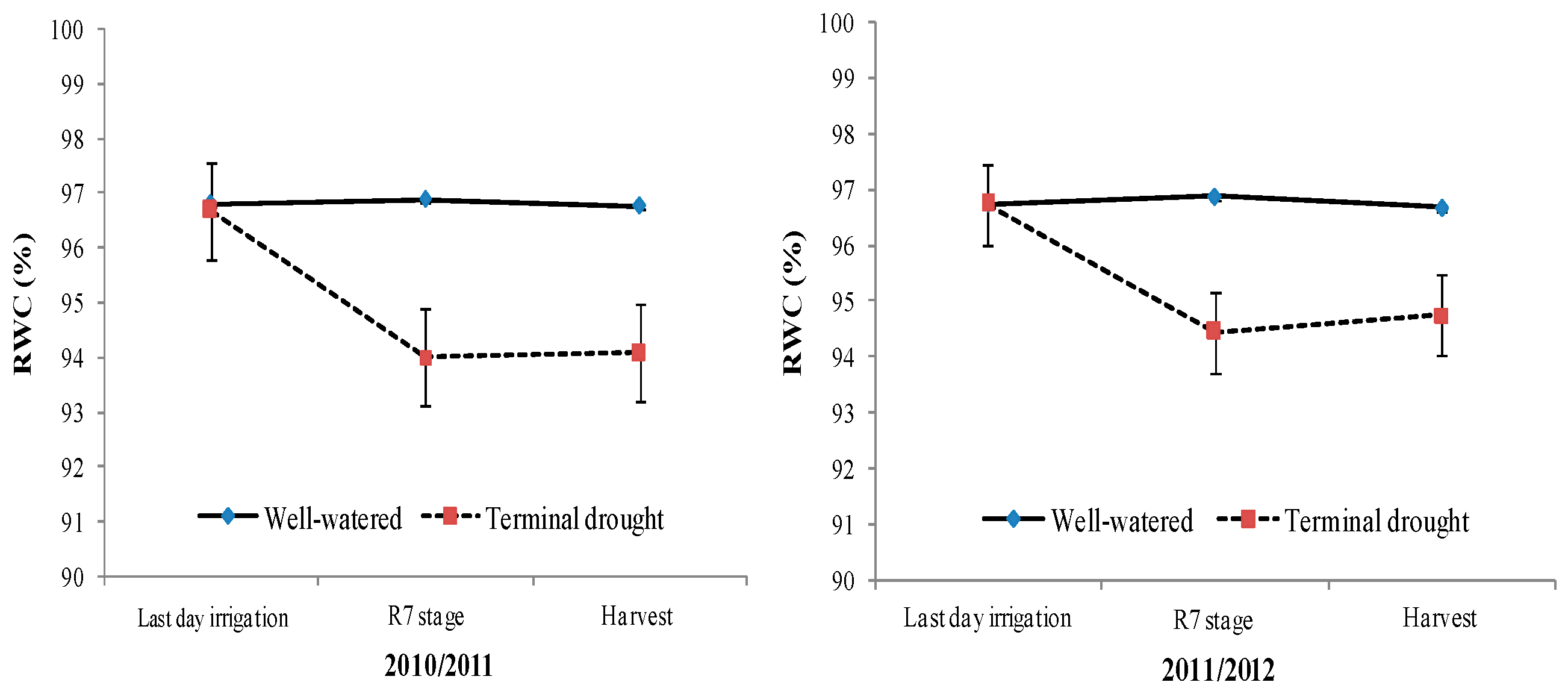
3.1. Soil and Plant Water Stress
3.2. Relationships between Nutrient Uptake with A. flavus Seed Infection and Aflatoxin Contamination
3.3. Relationships between N2-Fixation (NF) and Nodule Dry Weight (NDW) with A. flavus Seed Infection and Aflatoxin Contamination
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Girdthai, T.; Jogloy, S.; Vorasoot, N.; Akkasaeng, C.; Wongkaew, S.; Holbrook, C.C.; Patanothai, A. Associations between physiological traits for drought tolerance. Plant Breed. 2010, 129, 693–699. [Google Scholar] [CrossRef]
- Arunyanark, A.; Jogloy, S.; Akkasaeng, C.; Vorasoot, N.; Nageswara Rao, R.C.; Wright, G.C.; Patanothai, A. Association between aflatoxin contamination and drought tolerance traits in peanut. Field Crops Res. 2009, 114, 14–22. [Google Scholar] [CrossRef]
- Holbrook, C.C.; Kvien, C.K.; Rucker, K.S.; Wilson, D.W.; Hook, J.E. Preharvest aflatoxin contamination in drought tolerant and intolerant peanut genotypes. Peanut Sci. 2000, 27, 45–48. [Google Scholar] [CrossRef]
- Holbrook, C.C.; Guo, B.Z.; Wilson, D.M.; Timper, P. The U.S. breeding program to develop peanut with drought tolerance and reduced aflatoxin contamination. Peanut Sci. 2009, 36, 50–53. [Google Scholar] [CrossRef]
- Kolay, A.K. Soil moisture stress and plant growth. In Water and Crop Growth; Atlantic: New Delhi, India, 2008; pp. 43–48. [Google Scholar]
- Fageria, N.K.; Baligar, V.C.; Clark, R.B. Micronutrients in crop production. Adv. Agron. 2002, 77, 185–268. [Google Scholar] [CrossRef]
- Gunes, A.; Cicek, N.; Inal, A.; Alpaslan, M.; Eraslan, F.; Guneri, E.; Guzelordu, T. Genotypic response of chickpea (Cicer arietinum L.) cultivars to drought stress implemented at pre-and post- anthesis stages and its relations with nutrient uptake and efficiency. Plant Soil Environ. 2006, 52, 368–376. [Google Scholar] [CrossRef]
- Pimratch, S.; Jogloy, S.; Vorasoot, N.; Toomsan, B.; Kesmala, T.; Patanothai, A.; Holbrook, C.C. Effect of drought stress on traits related to N2-fixation in eleven peanut (Arachis hypogaea L.) genotypes differing in degrees of resistance to drought. Asian J. Plant Sci. 2008, 7, 334–342. [Google Scholar] [CrossRef]
- Htoon, W.; Jogloy, S.; Toomsan, B.; Sanitchon, J. Response to early drought for traits related to nitrogen fixation and their correlation to yield and drought tolerance traits in peanut (Arachis hypogaea L.). Asian J. Plant Sci. 2009, 8, 138–145. [Google Scholar] [CrossRef]
- Puangbut, D.; Jogloy, S.; Vorasoot, N.; Akkasaeng, C.; Patanothai, A. Association of transpiration efficiency with N2 fixation of peanut under early season drought. Int. J. Plant Prod. 2011, 5, 381–394. [Google Scholar] [CrossRef]
- Arunyanark, A.; Pimratch, S.; Jogloy, S.; Wongkaew, S.; Vorasoot, N.; Akkasaeng, C.; Kesmala, T.; Patanothai, A.; Holbrook, C.C. Association between aflatoxin contamination and N2 fixation in peanut under drought conditions. Int. J. Plant Prod. 2012, 6, 161–172. [Google Scholar] [CrossRef]
- Htoon, W.; Keawpradit, W.; Jogloy, S.; Vorasoot, N.; Toomsan, B.; Akkasaeng, C.; Puppala, N.; Patanothai, A. Responses of peanut (Arachis hypogaea L.) genotypes to N2-fixation under terminal drought and their contributions to peanut yield. SABRAO J. Breed. Genet. 2013, 45, 296–310. [Google Scholar]
- Doorenbos, J.; Pruitt, W.O. Calculation of crop water requirements. In Guideline for Predicting Crop Water Requirements; Doorenbos, J., Pruitt, W.O., Eds.; FAO Irrigation and Drainage Paper No. 24; FAO: Rome, Italy, 1992; pp. 1–65. [Google Scholar]
- Singh, S.; Russell, M.B. Water use by maize/pigeonpea intercrop on a deep vertisol. In Proceedings of the International Workshop on Pigeonpeas Held in ICRISAT Center Patancheru, Andhra Pradesh, India, 15–19 December 1980; ICRISAT: Andhra Pradesh, India, 1980; pp. 271–282. [Google Scholar]
- Kramer, P.J. Drought stress and the origin of adaptation. In Adaptation of Plant to Water and High Temperature Stress; Turner, N.C., Kramer, P.J., Eds.; John Wiley & Sons: New York, NY, USA, 1980; pp. 7–20. [Google Scholar]
- Clavel, D.; Diouf, O.; Khalfaoui, J.L.; Braconnier, S. Genotypes variations in fluorescence parameters among closely related groundnut (Arachis hypogaea L.) lines and their potential for drought screening programs. Crops Sci. 2006, 34, 92–97. [Google Scholar] [CrossRef]
- Kaewpradit, W.; Toomsan, B.; Cadisch, G.; Vityakon, P.; Limpinuntana, V.; Saenjan, P.; Jogloy, S.; Patanothai, A. Mixing groundnut residues and rice straw to improve rice yield and N use efficiency. Field Crops. Res. 2009, 110, 130–138. [Google Scholar] [CrossRef]
- Nautiyal, P.C.; Nageswara Rao, R.C.; Joshi, Y.C. Moisture-deficit-induced changes in leaf-water content, leaf carbon exchange rate and biomass production in groundnut cultivars differing in specific leaf area. Field Crops Res. 2002, 64, 67–79. [Google Scholar] [CrossRef]
- McMillian, W.W.; Widstrom, N.W.; Wilson, D.M. Aflatoxin contamination of preharvest corn in Georgia: A six-year study of insect damage and visible Aspergillus flavus. J. Environ. Qual. 1985, 14, 200–202. [Google Scholar] [CrossRef]
- Htoon, W.; Jogloy, S.; Vorasoot, N.; Toomsan, B.; Keawpradit, W.; Akkasaeng, C.; Puppala, N.; Patanothai, A. Nutrient uptakes and their contributions to yield in peanut genotypes with different levels of terminal drought resistance. Turk. J. Agric. For. 2014, 39, 781–791. [Google Scholar] [CrossRef]
- Koolachart, R.; Jogloy, S.; Vorasoot, N.; Wongkaew, S.; Holbrook, C.C.; Jongrungklang, N.; Kesmala, T.; Patanothai, A. Association of aflatoxin contamination and root responses of peanut genotypes with different levels of terminal drought resistance. SABRAO J. Breed. Genet. accepted.
- Samarah, N.; Mullen, R.; Cianzio, S. Size distribution and mineral nutrients of soybean seeds in response to drought stress. J. Plant Nutr. 2004, 27, 815–835. [Google Scholar] [CrossRef]
- Uppala, S.S. Factors affecting pre-harvest aflatoxin contamination of peanut (Arachis hypogaea L.). Ph.D. Thesis, Auburn University, Albama, AL, USA, 6 August 2011. [Google Scholar]
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient use efficiency in plants. Commun. Soil Sci. Plant Anal. 2007, 32, 921–950. [Google Scholar] [CrossRef]
- Tafteh, A.; Sepaskhah, A.R. Yield and nitrogen leaching in maize field under different nitrogen rates and partial root drying irrigation. Int. J. Plant Prod. 2012, 6, 93–114. [Google Scholar] [CrossRef]
- Ahmadi, S.H.; Andersen, M.N.; Laerke, P.E.; Plauborg, F.; Sepaskhah, A.R.; Jensen, C.R.; Hansen, S. Interaction effect of different irrigation strategies and soil textures on the nitrogen uptake of field grown potatoes. Int. J. Plant Prod. 2011, 5, 263–274. [Google Scholar]


{kind=link}
| Treatments | Soil Depth (cm) | Soil Moisture Content (%) | |||
|---|---|---|---|---|---|
| Sowing | Last Day of Irrigation | R7 Stage | Harvest | ||
| Well-watered | 0–30 | 10.20 | 10.28 | 10.27 | 10.29 |
| Terminal drought | 0–30 | 10.28 | 10.21 | 6.14 | 6.15 |
| Well-watered | 0–60 | 10.27 | 9.78 | 9.94 | 10.19 |
| Terminal drought | 0–60 | 10.24 | 9.75 | 6.02 | 6.44 |
| Nutrient Uptake (g plant−1) | A. flavus Seed Infection (%) | Aflatoxin Contamination (ppb) | ||||
|---|---|---|---|---|---|---|
| Well-Watered | Terminal Drought | DTI | Well-Watered | Terminal Drought | DTI | |
| N | −0.02 | −0.50 ** | −0.78 ** | −0.30 | −0.66 ** | −0.82 ** |
| P | −0.05 | −0.33 * | −0.66 ** | −0.15 | −0.43 ** | −0.60 ** |
| K | 0.08 | −0.51 ** | −0.72 ** | −0.18 | −0.60 ** | −0.68 ** |
| Ca | −0.45 ** | −0.52 ** | −0.65 ** | −0.21 | −0.75 ** | −0.73 ** |
| Mg | 0.11 | −0.32 * | −0.75 ** | −0.22 | −0.50 ** | −0.72 ** |
| NF and Related Trait (g plant−1) | A. flavus Seed Infection (%) | Aflatoxin Contamination (ppb) | ||||
|---|---|---|---|---|---|---|
| Well-Watered | Terminal Drought | DTI | Well-Watered | Terminal Drought | DTI | |
| NF | −0.41 ** | −0.50 ** | −0.65 ** | −0.58 ** | −0.62 ** | −0.38 * |
| NDW | −0.58 ** | −0.62 ** | −0.71 ** | −0.33 * | −0.35 * | −0.33 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Htoon, W.; Kaewpradit, W.; Vorasoot, N.; Toomsan, B.; Akkasaeng, C.; Puppala, N.; Wongkaew, S.; Jogloy, S. Relationships between Nutrient Uptake and Nitrogen Fixation with Aflatoxin Contamination in Peanut under Terminal Drought. Agronomy 2019, 9, 419. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9080419
Htoon W, Kaewpradit W, Vorasoot N, Toomsan B, Akkasaeng C, Puppala N, Wongkaew S, Jogloy S. Relationships between Nutrient Uptake and Nitrogen Fixation with Aflatoxin Contamination in Peanut under Terminal Drought. Agronomy. 2019; 9(8):419. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9080419
Chicago/Turabian StyleHtoon, Wunna, Wanwipa Kaewpradit, Nimitr Vorasoot, Banyong Toomsan, Chutipong Akkasaeng, Naveen Puppala, Sopone Wongkaew, and Sanun Jogloy. 2019. "Relationships between Nutrient Uptake and Nitrogen Fixation with Aflatoxin Contamination in Peanut under Terminal Drought" Agronomy 9, no. 8: 419. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9080419