Circulation of Indigenous Bovine Respiratory Syncytial Virus Strains in Turkish Cattle: The First Isolation and Molecular Characterization

 ,
,  ,
,  , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Samples
2.2. Virus Isolation
2.3. Antigen-ELISA for BRD-Associated Viruses
2.4. Nucleic Acid Extraction and Amplification
2.5. Sequencing and Phylogenetic Analysis
3. Results
3.1. The Identification of Lung Tissue Samples Infected with BRSV
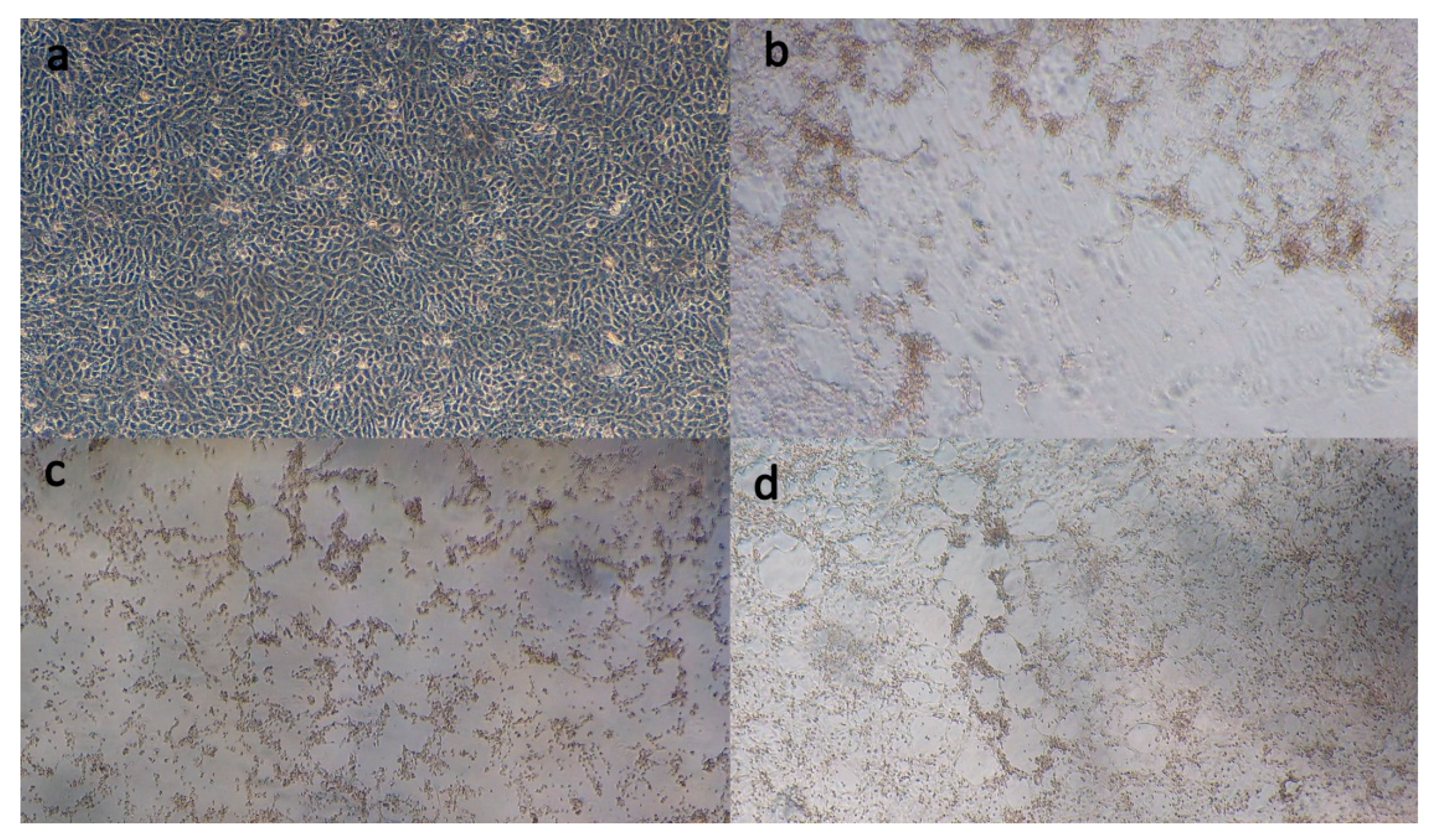
3.2. Virus Isolation
3.3. RT-Nested PCR
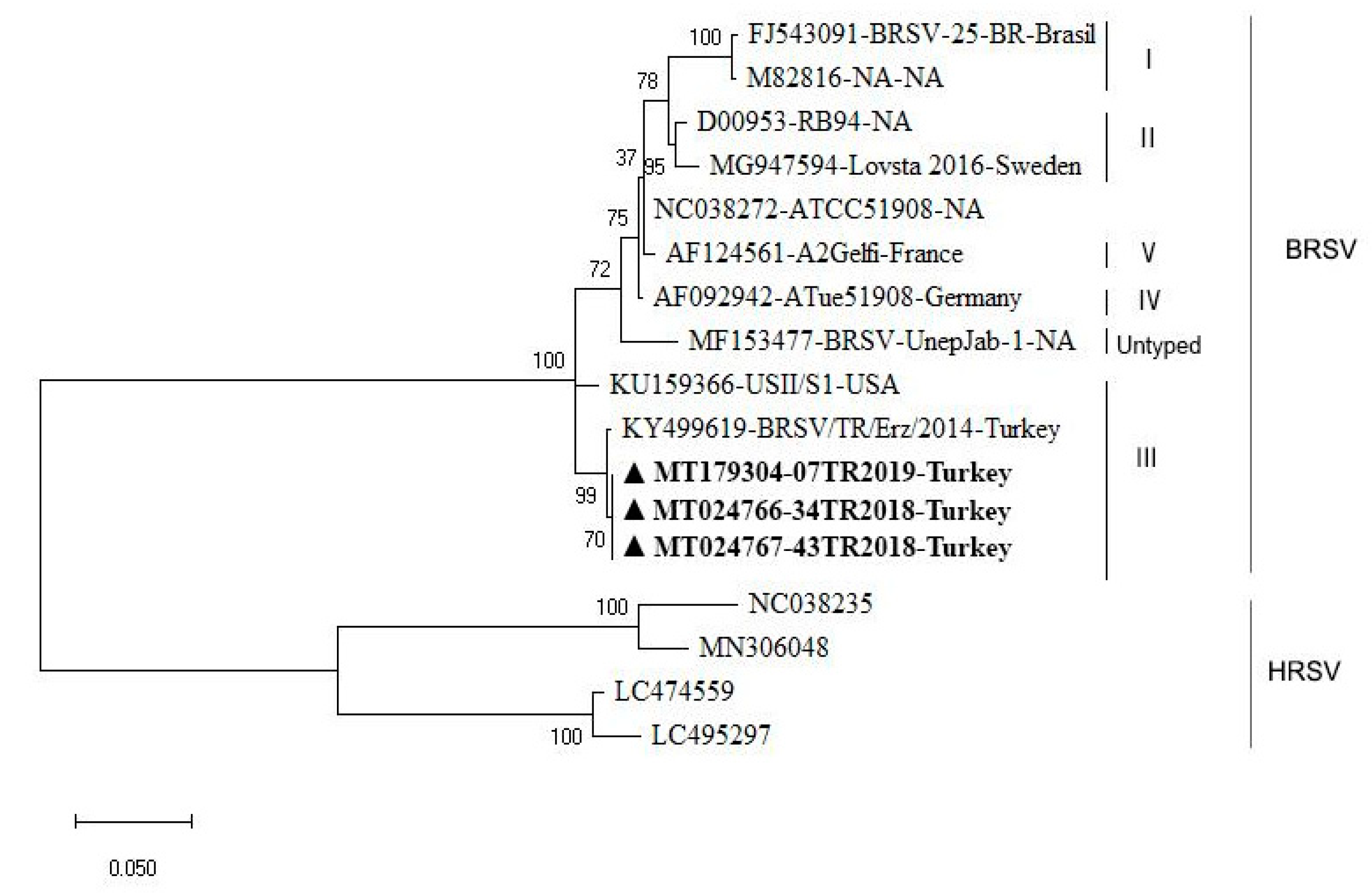
3.4. Sequencing and Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Ethical Statement
Data Availability
References
- Fulton, R.W. Bovine respiratory disease research (1983–2009). Anim. Health. Res. Rev. 2009, 10, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Headley, S.A.; Okano, W.; Balbo, L.C.; Marcasso, R.A.; Oliveira, T.E.; Alfieri, A.F.; NegriFilho, L.C.; Michelazzo, M.Z.; Rodrigues, S.C.; Baptista, A.L.; et al. Molecular survey of infectious agents associated with bovine respiratory disease in a beef cattle feedlot in southern Brazil. J. Vet. Diagn. Invest. 2018, 30, 249–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albayrak, H.; Yazici, Z.; Ozan, E.; Tamer, C.; Abd El Wahed, A.; Wehner, S.; Ulrich, K.; Weidmann, M. Characterisation of the First Bovine Parainfluenza Virus 3 Isolate Detected in Cattle in Turkey. Vet. Sci. 2019, 6, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacioglu, I.K.; Coskun, N.; Duran, S.; Sevinc, S.; Alkan, F. Phylogenetic Analysis of Bovine Respiratory Syncytial Virus from Calves with Respiratory Disorders. Kafkas Univ. Vet. Fak. Derg. 2019, 25, 251–256. [Google Scholar] [CrossRef]
- Timurkan, M.O.; Aydin, H.; Sait, A. Identification and Molecular Characterisation of Bovine Parainfluenza Virus-3 and Bovine Respiratory Syncytial Virus-First Report from Turkey. J. Vet. Res. 2019, 63, 167–173. [Google Scholar] [CrossRef] [Green Version]
- International Committee on Taxonomy of Viruses. 2019 Release. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/negative-sense-rna-viruses/mononegavirales/w/pneumoviridae/738/genus-orthopneumovirus/ (accessed on 3 September 2020).
- Bertolotti, L.; Giammarioli, M.; Rosati, S. Genetic characterization of bovine respiratory syncytial virus strains isolated in Italy: Evidence for the circulation of new divergent clades. J. Vet. Diagn. Invest. 2018, 30, 300–304. [Google Scholar] [CrossRef] [Green Version]
- Taylor, G.; Wyld, S.; Valarcher, J.F.; Guzman, E.; Thom, M.; Widdison, S.; Buchholz, U.J. Recombinant bovine respiratory syncytial virus with deletion of the SH gene induces increased apoptosis and pro-inflammatory cytokines in vitro, and is attenuated and induces protective immunity in calves. J. Gen. Virol. 2014, 95, 1244–1254. [Google Scholar] [CrossRef]
- Valarcher, J.F.; Schelcher, H.B.; Bourhy, H. Evolution of Bovine Respiratory Syncytial Virus. J. Virol. 2000, 74, 10714–10728. [Google Scholar] [CrossRef] [Green Version]
- Krešić, N.; Bedeković, T.; Brnić, D.; Šimić, I.; Lojkić, I.; Turk, N. Genetic analysis of bovine respiratory syncytial virus in Croatia. Comp. Immunol. Microbiol. Infect. Dis. 2018, 58, 52–57. [Google Scholar] [CrossRef]
- Leme, R.A.; Agnol, A.M.D.; Balbo, L.C.; Pereria, F.L.; Possatti, F.; Alfieri, A.F.; Alfieri, A.A. Molecular characterization of Brazilian wild-type strains of Bovine respiratory syncytial virus reveals genetic diversity and putative subgroup of the virus. Vet. Q. 2020, 40, 83–96. [Google Scholar] [CrossRef]
- Sarmiento-Silva, R.E.; Nakamura-Lopez, Y.; Vaughan, G. Epidemiology, Molecular Epidemiology and Evolution of Bovine Respiratory Syncytial Virus. Viruses 2012, 4, 3452–3467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppe, I.B.A.L.; Medeiros, A.S.R.; Arns, C.W.; Samara, S.I. Bovine respiratory syncytial virus seroprevalence and risk factors in non-vaccinated dairy cattle herds in Brazil. BMC Vet. Res. 2018, 14, 208. [Google Scholar] [CrossRef] [PubMed]
- Urban-Chmiel, R.; Wernicki, A.; Puchalski, A.; Dec, M.; Stęgierska, D.; Grooms, D.L.; Barbu, N.I. Detection of bovine respiratory syncytial virus infections in young dairy and beef cattle in Poland. Vet. Q. 2014, 35, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Baptista, A.L.; Rezende, A.L.; Fonseca, P.D.A.; Massi, R.P.; Nogueira, G.M.; Magalhães, L.Q.; Headley, S.A.; Menezes, G.L.; Alfieri, A.A.; Saut, J.P.E. Bovine respiratory disease complex associated mortality and morbidity rates in feedlot cattle from southeastern Brazil. J. Infect. Dev. Ctries. 2017, 11, 791–799. [Google Scholar] [CrossRef] [Green Version]
- Yazici, Z.; Gumusova, S.; Tamer, C.; Muftuoglu, B.; Ozan, E.; Arslan, S.; Bas, O.; Elhag, A.E.; Albayrak, H. The first serological report for genotype C bovine parainfluenza 3 virus in ruminant species of mid-northern Turkey: Traces from the past. Trop. Biomed. 2019, 36, 803–809, Scopus EID: 2-s2.0-85073422601. [Google Scholar]
- Boxus, M.; Letellier, C.; Kerkhofs, P. Real Time RT-PCR for the detection and quantitation of bovine respiratory syncytial virus. J. Virol. Methods. 2005, 125, 125–130. [Google Scholar] [CrossRef]
- Vilcek, S.; Elvander, M.; Ballagi-Pordány, A.; Belák, S. Development of nested PCR assays for detection of bovine respiratory syncytial virus in clinical samples. J. Clin. Microbiol. 1994, 32, 2225–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kurcubic, V.; Dokovic, R.; Ilic, Z.; Petrovic, M. Etiopathogenesis and economic significance of bovine respiratory disease complex (BRD). Acta Agric. Serb. 2018, 45, 85–100. [Google Scholar] [CrossRef]
- Albayrak, H.; Tamer, C.; Ozan, E.; Muftuoglu, B.; Kadi, H.; Dogan, F.; Cagirgan, A.A.; Elhag, A.E.; Akman, A.; Kurucay, H.N.; et al. Molecular identification and phylogeny of bovine herpesvirus-1 (BoHV-1) from cattle associated with respiratory disorders and death in Turkey. Med. Weter. 2020, 76, 358–361. [Google Scholar] [CrossRef]
- Alkan, F.; Ozkul, A.; Bilge-Dagalp, S.; Yesilbag, K.; Oguzoglu, C.; Akca, Y.; Burgu, I. Virological and serological studies on the role of PI-3 virus, BRSV, BVDV, and BHV- 1 on respiratory infections of cattle. I. The detection of etiological agents by direct immunofluorescence technique. DTW. Dtsch. Tierarztl. Wochenschr. 2000, 107, 193–195. [Google Scholar] [PubMed]
- Yesilbag, K.; Gungor, B. Seroprevalence of bovine respiratory viruses in North-Western Turkey. Trop. Anim. Health Prod. 2008, 40, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.S.; Domingues, H.G.; Coswig, L.T.; D’Arce, R.C.F.; Carvalho, R.F.; Arns, C.W. Detection of bovive respiratory syncytial virus in experimentally infected balb/c mice. Vet. Res. 2004, 35, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santangelo, P.; Nitin, N.; LaConte, L.; Woolums, A.; Bao, G. Live-cell characterization and analysis of a clinical isolate of bovine respiratory syncytial virus, using molecular beacons. J. Virol. 2006, 80, 682–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bortolin Affonso, I.; de Souza, A.; Cavalheiro Martini, M.; Bianchi dos Santos, M.; Rosado Spilki, F.; Weis Arns, C.; Issa Samara, S. Detection of an untyped strain of bovine respiratory syncytial virus in a dairy herd. Semin. Cienc. Agrar. 2014, 35, 2539–2549. [Google Scholar] [CrossRef]
- Valentova, V. The antigenic and genetic variability of bovine respiratory syncytial virus with emphasis on the G protein. Vet. Med. 2003, 48, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Dean, A.S.; Fournié, G.; Kulo, A.E.; Boukaya, G.A.; Schelling, E.; Bonfoh, B. Potential risk of regional disease spread in West Africa through cross-border cattle trade. PLoS ONE 2013, 8, e75570. [Google Scholar] [CrossRef] [Green Version]
- Domingues, H.G.; Spilki, F.R.; Arns, C.W. Molecular detection and phylogenetic analysis of bovine respiratory syncytial virus (BRSV) in swabs and lung tissues of adult cattle. Pesq. Vet. Bras. 2011, 31, 961–966. [Google Scholar] [CrossRef] [Green Version]
- Valarcher, J.F.; Furze, J.; Wyld, S.G.; Cook, R.; Zimmer, G.; Herrler, G.; Taylor, G. Bovine respiratory syncytial virus lacking the virokinin or with a mutation in furin cleavage site RA(R/K)R109 induces less pulmonary inflammation without impeding the induction of protective immunity in calves. J. Gen. Virol. 2006, 87, 1659–1667. [Google Scholar] [CrossRef]
- Zimmer, G.; Rohn, M.; McGregor, G.P.; Schemann, M.; Conzelmann, K.K.; Herrler, G. Virokinin, a bioactive peptide of the tachykinin family, is released from the fusion protein of bovine respiratory syncytial virus. J. Biol. Chem. 2003, 278, 46854–46861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okur-Gumusova, S.; Tamer, C.; Ozan, E.; Cavunt, A.; Kadi, H.; Muftuoglu, B.; Eisa Elhag, A.; Yazici, Z.; Albayrak, H. An investigation of the seroprevalence of crimean-congo hemorrhagic fever and lumpy skin disease in domesticated water buffaloes in Northern Turkey. Trop. Biomed. 2020, 37, 165–173, Scopus EID: 2-s2.0-85086915081. [Google Scholar]



{kind=link}
{kind=link}
| Primers and Probe | Sequences (5′-3′) | Product Size(bp) | Ref. |
|---|---|---|---|
| F primer | GCAATGCTGCAGGACTAGGTATAAT | 124 | Boxus et al. [17] |
| R primer | ACACTGTAATTGATGACCCCATTCT | ||
| Probe | FAM-ACCAAGACTTGTATGATGCTGCCAAAGCA-TAMRA | ||
| B1 | AATCAACATGCAGTGCAGTTAG | 711 | Vilcek et al. [18] |
| B2A | TTTGGTCATTCGTTATAGGCAT | ||
| B3 | GTGCAGTTAGTAGAGGTTATCTTAGT | 481 | |
| B4A | TAGTTCTTTAGATCAAGTACTTTGCT |
| GenBank Number | Strain Name and Country | Amino Acid Similarities (%) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NC038272 | FJ543091 | M82816 | D00953 | MG947594 | KU159366 | KY499619 | MT179304* | MT024766* | MT024767* | AF092942 | AF124561 | MF153477 | |||
| Nucleotide Similarities (%) | NC038272 | ATCC51908 NA | 96.05 | 96.05 | 97.65 | 98.44 | 93.60 | 92.77 | 93.60 | 93.60 | 93.60 | 98.44 | 100.00 | 95.24 | |
| FJ543091 | BRSV-25-BR Brasil | 96.14 | 98.44 | 95.24 | 96.05 | 89.38 | 88.51 | 89.38 | 89.38 | 89.38 | 94.42 | 96.05 | 91.09 | ||
| M82816 | NA NA | 96.14 | 99.49 | 95.24 | 96.05 | 90.24 | 89.38 | 90.24 | 90.24 | 90.24 | 94.42 | 96.05 | 91.93 | ||
| D00953 | RB 93 NA | 98.20 | 96.40 | 96.40 | 97.65 | 91.93 | 90.24 | 91.09 | 91.09 | 91.09 | 96.05 | 97.65 | 92.77 | ||
| MG947594 | Lovsta 2016 Sweden | 97.69 | 95.89 | 95.89 | 98.46 | 91.93 | 91.09 | 91.93 | 91.93 | 91.93 | 96.85 | 98.44 | 93.60 | ||
| KU159366 | USII/S1 USA | 96.14 | 93.06 | 93.57 | 94.60 | 94.09 | 97.65 | 98.44 | 98.44 | 98.44 | 93.60 | 93.60 | 93.60 | ||
| KY499619 | BRS/TR/Erz/2014 Turkey | 95.63 | 93.06 | 93.57 | 94.09 | 93.57 | 97.43 | 99.22 | 99.22 | 99.22 | 92.77 | 92.77 | 92.77 | ||
| MT179304 * | 07TR2019 Turkey | 95.63 | 93.57 | 94.09 | 94.09 | 93.57 | 97.43 | 99.49 | 100.00 | 100.00 | 93.60 | 93.60 | 93.60 | ||
| MT024766 * | 34TR2018 Turkey | 95.63 | 93.57 | 94.09 | 94.09 | 93.57 | 97.43 | 99.49 | 100.00 | 100.00 | 93.60 | 93.60 | 93.60 | ||
| MT024767 * | 43TR2018 Turkey | 95.63 | 93.57 | 94.09 | 94.09 | 93.57 | 97.43 | 99.49 | 100.00 | 100.00 | 93.60 | 93.60 | 93.60 | ||
| AF092942 | ATue51908 Germany | 99.49 | 95.89 | 95.89 | 97.94 | 97.43 | 96.14 | 95.63 | 95.63 | 95.63 | 95.63 | 98.44 | 95.24 | ||
| AF124561 | A2Gelfi France | 99.49 | 95.63 | 95.63 | 97.69 | 97.17 | 95.63 | 95.12 | 95.12 | 95.12 | 95.12 | 98.97 | 95.24 | ||
| MF153477 | BRSV-UnepJab-1 NA | 96.66 | 93.06 | 93.06 | 95.12 | 94.60 | 94.86 | 94.34 | 94.34 | 94.34 | 94.34 | 96.66 | 96.14 | ||
| Sub- Groups | GenBank Number | Strain Name | Amino Acid Positions of BRSV F Gene Sequence | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 67 | 70 | 71 | 75 | 80 | 91 | 100 | 101 | 102 | 104 | 105 | 113 | 114 | 115 | 118 | 124 | 148 | 168 | 173 | 176 | |||
| I | FJ543091 | BRSV-25-BR | N | N | G | K | K | V | E | P | T | S | S | E | S | I | T | K | I | N | S | K |
| M82816 | NA | . | . | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . | K | . | . | |
| II | D00953 | RB 94 | . | . | S | N | . | . | . | . | A | . | . | . | L | . | K | . | . | K | . | . |
| MG947594 | Lovsta 2016 | . | . | S | . | . | . | . | L | A | . | . | . | L | . | . | . | . | K | . | . | |
| III | KU159366 | USII/S1 | D | K | S | . | . | T | V | . | A | F | N | . | L | M | A | R | . | K | . | . |
| KY499619 | BRS/TR/Erz/2014 | D | K | S | . | . | T | V | . | A | F | N | . | L | M | . | R | . | K | T | E | |
| MT179304 * | 07TR2019 | D | K | S | . | . | T | V | . | A | F | N | . | L | M | . | R | . | K | T | . | |
| MT024766 * | 34TR2018 | D | K | S | . | . | T | V | . | A | F | N | . | L | M | . | R | . | K | T | . | |
| MT024767 * | 43TR2018 | D | K | S | . | . | T | V | . | A | F | N | . | L | M | . | R | . | K | T | . | |
| IV | AF092942 | ATue51908 | . | K | S | . | . | . | . | . | A | F | . | . | L | . | . | . | V | K | . | . |
| V | AF124561 | A2Gelfi | . | . | S | . | . | . | . | . | A | F | . | . | L | . | . | . | . | K | . | . |
| NC038272 | ATCC51908 | . | . | S | . | . | . | . | . | A | F | . | . | L | . | . | . | . | K | . | . | |
| Untyped | MF153477 | BRSV-UnepJab-1 | . | K | S | . | Q | I | . | . | A | F | . | G | L | T | . | R | . | K | . | . |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yazici, Z.; Ozan, E.; Tamer, C.; Muftuoglu, B.; Barry, G.; Kurucay, H.N.; Elhag, A.E.; Cagirgan, A.A.; Gumusova, S.; Albayrak, H. Circulation of Indigenous Bovine Respiratory Syncytial Virus Strains in Turkish Cattle: The First Isolation and Molecular Characterization. Animals 2020, 10, 1700. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10091700
Yazici Z, Ozan E, Tamer C, Muftuoglu B, Barry G, Kurucay HN, Elhag AE, Cagirgan AA, Gumusova S, Albayrak H. Circulation of Indigenous Bovine Respiratory Syncytial Virus Strains in Turkish Cattle: The First Isolation and Molecular Characterization. Animals. 2020; 10(9):1700. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10091700
Chicago/Turabian StyleYazici, Zafer, Emre Ozan, Cuneyt Tamer, Bahadir Muftuoglu, Gerald Barry, Hanne Nur Kurucay, Ahmed Eisa Elhag, Abdurrahman Anil Cagirgan, Semra Gumusova, and Harun Albayrak. 2020. "Circulation of Indigenous Bovine Respiratory Syncytial Virus Strains in Turkish Cattle: The First Isolation and Molecular Characterization" Animals 10, no. 9: 1700. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10091700