Effects of Tissue Preservation on Carbon and Nitrogen Stable Isotope Signatures in Syngnathid Fishes and Prey

 ,
, 
 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Live Prey
2.2. Fishes
2.3. Experimental Preservation Procedures
2.4. Isotope Ratio Mass Spectrometry
2.5. Data Analysis
2.6. Bioethical Approval
3. Results
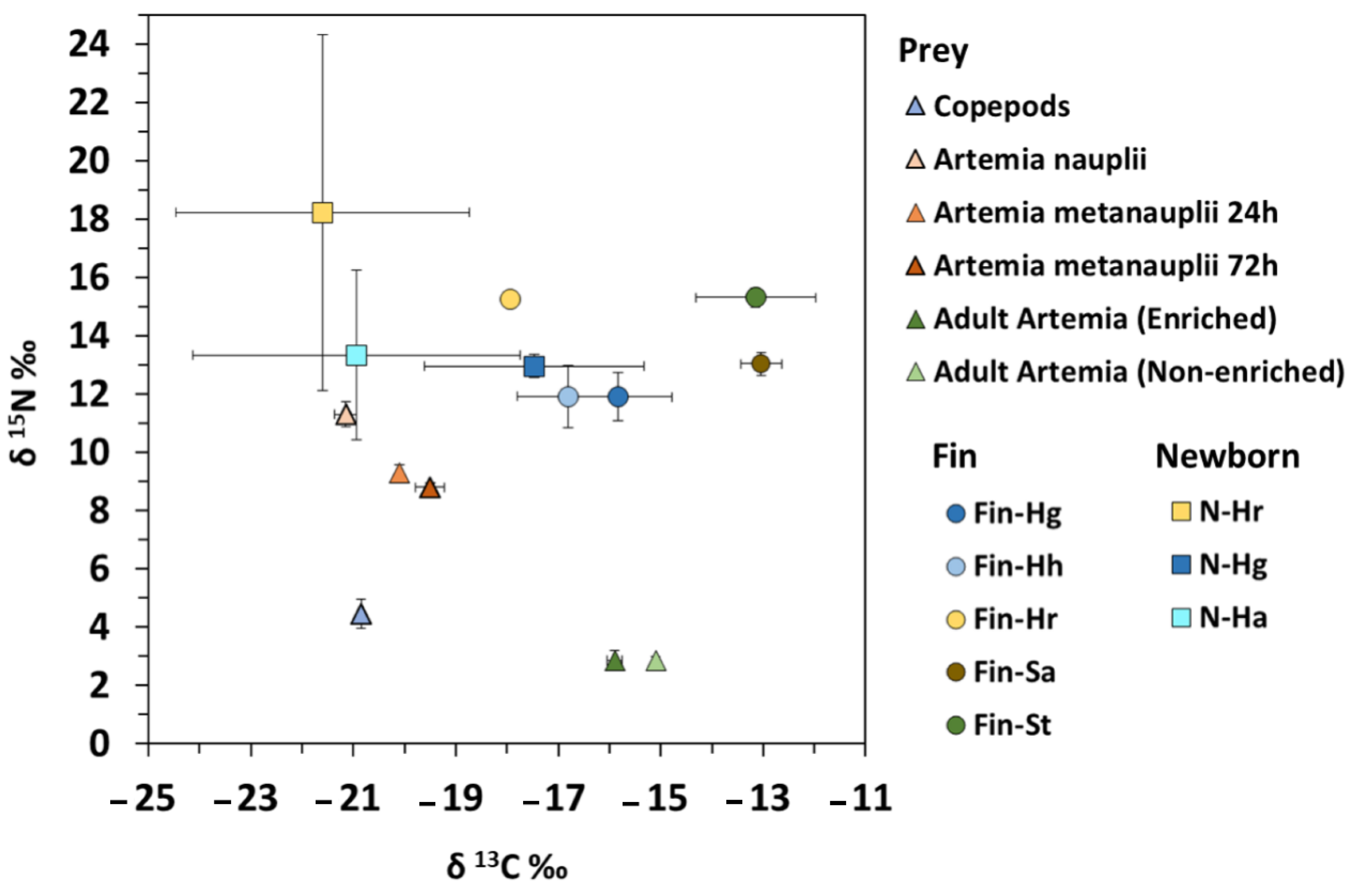
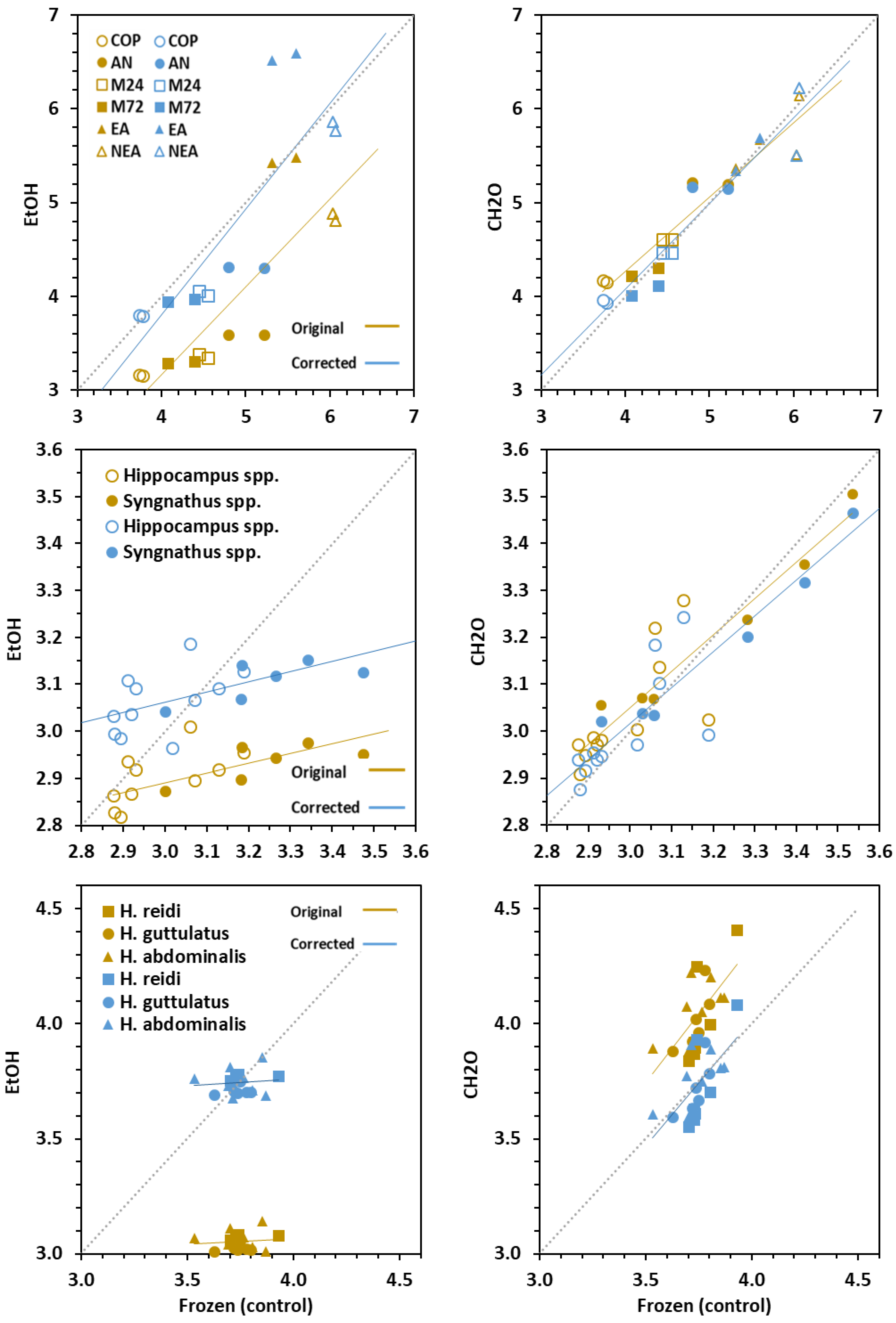
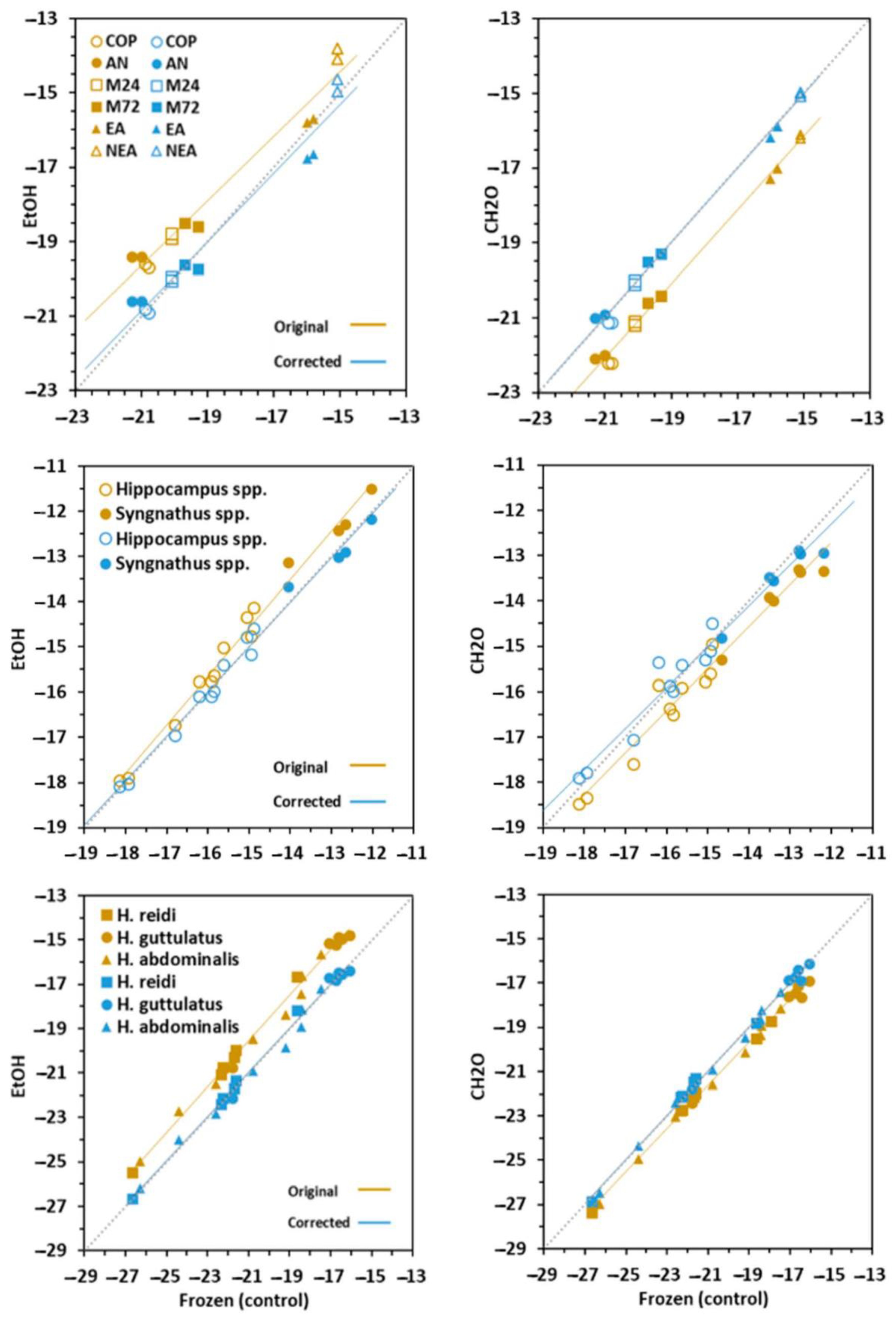
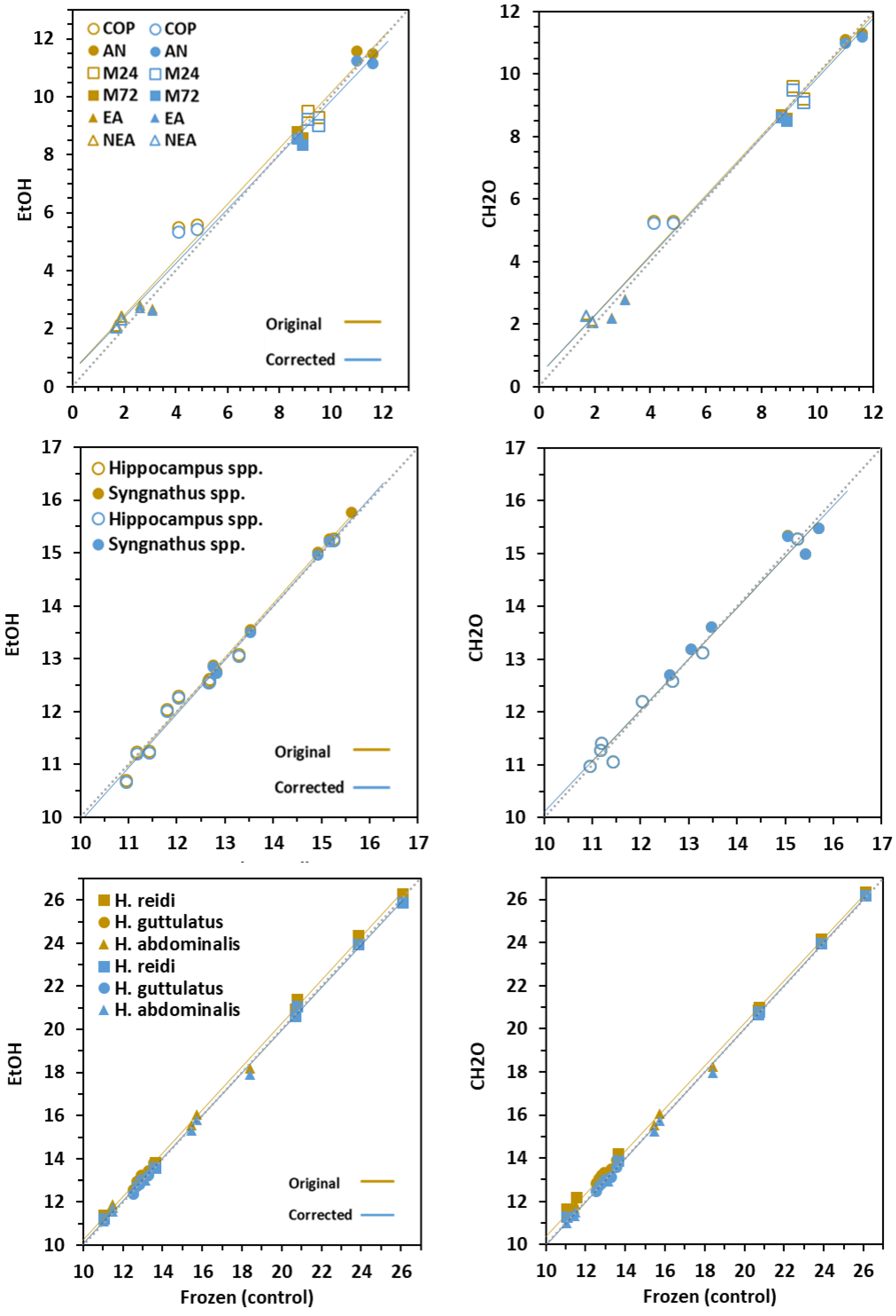
3.1. Effect on Carbon and Nitrogen Ratios
3.2. Effect on δ13C Signatures
3.3. Effect on δ15N Signatures
3.4. Conversion Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Peterson, B.J.; Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Rasmussen, J.B. Variation in δ15N and δ13C trophic fractionation: Implications for aquatic food web studies. Limnol. Oceanogr. 2001, 46, 2061–2066. [Google Scholar] [CrossRef]
- Gannes, L.Z.; O’Brien, D.M.; Martínez del Rio, C. Stable isotopes in animal ecology: Assumptions, caveats, and a call for more laboratory experiments. Ecology 1997, 78, 1271–1276. [Google Scholar] [CrossRef]
- Boecklen, W.J.; Yarnes, C.T.; Cook, B.A.; James, A.A. On the use of stable isotopes in trophic ecology. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 411–440. [Google Scholar] [CrossRef] [Green Version]
- Herzka, S.Z.; Holt, G.J. Changes in isotopic composition of red drum (Sciaenops ocellatus) larvae in response to dietary shifts: Potential applications to settlement studies. Can. J. Fish. Aquat. Sci. 2000, 57, 137–147. [Google Scholar] [CrossRef]
- Le Vay, L.; Gamboa-Delgado, J. Naturally-occurring stable isotopes as direct measures of larval feeding efficiency, nutrient incorporation and turnover. Aquaculture 2011, 315, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Hansson, S.; Hobbie, J.E.; Elmgren, R.; Larsson, U.; Fry, B.; Johansson, S. The stable nitrogen isotope ratio as a marker of food-web interactions and fish migration. Ecology 1997, 78, 2249–2257. [Google Scholar] [CrossRef]
- Hobson, K.A. Tracing origins and migration of wildlife using stable isotopes: A review. Oecologia 1999, 120, 314–326. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Acta 1981, 45, 341–351. [Google Scholar] [CrossRef]
- Pinnegar, J.K.; Polunin, N.V.C. Differential fractionation of δ13C and δ15N among fish tissues: Implications for the study of trophic interactions. Funct. Ecol. 1999, 13, 225–231. [Google Scholar] [CrossRef]
- Wolf, N.; Carleton, S.A.; Martínez del Rio, C. Ten years of experimental animal isotopic ecology. Funct. Ecol. 2009, 23, 17–26. [Google Scholar] [CrossRef]
- Minagawa, M.; Wada, E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochim. Cosmochim. Acta 1984, 48, 1135–1140. [Google Scholar] [CrossRef]
- Jardine, T.D.; Gray, M.A.; McWilliam, S.M.; Cunjak, R.A. Stable isotope variability in tissues of temperate stream fishes. Trans. Am. Fish. Soc. 2005, 134, 1103–1110. [Google Scholar] [CrossRef]
- Caut, S.; Angulo, E.; Courchamp, F. Variation in discrimination factors (∆15N and ∆13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 2009, 46, 443–453. [Google Scholar] [CrossRef]
- Jardine, T.D.; Hunt, R.J.; Pusey, J.B.; Bunn, S.E. A non-lethal sampling method for stable carbon and nitrogen isotope studies of tropical fishes. Mar. Freshw. Res. 2011, 62, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Blanco, A.; Deudero, S.; Box, A. Muscle and scale isotopic offset of three fish species in the Mediterranean Sea: Dentex dentex, Argyrosomus regius and Xyrichtys novacula. Rapid Commun. Mass Spectrom. 2009, 23, 2321–2328. [Google Scholar] [CrossRef]
- Cano-Rocabayera, O.; Maceda-Veiga, A.; de Sostoa, A. Fish fins and scales as non-lethally sampled tissues for stable isotope analysis in five fish species of north—Eastern Spain. Environ. Biol. Fishes 2015, 98, 925–932. [Google Scholar] [CrossRef]
- Winter, E.R.; Nyqvist, M.; Britton, J.R. Non-lethal sampling for stable isotope analysis of pike Esox lucius: How mucus, scale and fin tissue compare to muscle. J. Fish Biol. 2019, 95, 956–958. [Google Scholar] [CrossRef]
- Sanderson, B.L.; Tran, C.D.; Coe, H.J.; Pelekis, V.; Steel, E.A.; Reichert, W.L. Nonlethal sampling of fish caudal fins yields valuable stable isotope data for threatened and endangered fishes. Trans. Am. Fish. Soc. 2009, 138, 1166–1177. [Google Scholar] [CrossRef]
- Valladares, S.; Planas, M. Non-lethal dorsal fin sampling for stable isotope analysis in seahorses. Aquat. Ecol. 2012, 46, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Planas, M.; Chamorro, A.; Paltrinieri, A.; Campos, S.; Jiménez, A.; Nedelec, K.; Hernández-Urcera, J. Effect of diet on breeders and inheritance in syngnathids: Application of isotopic experimentally derived data to field studies. Mar. Ecol. Prog. Ser. 2020, 650, 107–123. [Google Scholar] [CrossRef]
- Planas, M.; Olivotto, I.; González, M.J.; Laurà, R.; Zarantoniello, M. A multidisciplinary experimental study of the effects of breeders diet on newborn seahorses (Hippocampus guttulatus). Front. Mar. Sci. 2020, 7, 638. [Google Scholar] [CrossRef]
- Bosley, K.L.; Wainright, S.C. Effects of preservatives and acidification on the stable isotope ratios (15N:14N, 13C:12C) of two species of marine animals. Can. J. Fish. Aquat. Sci. 1999, 56, 2181–2185. [Google Scholar] [CrossRef]
- Kelly, B.; Dempson, J.B.; Power, M. The effects of preservation on fish tissue stable isotope signatures. J. Fish Biol. 2006, 69, 1595–1611. [Google Scholar] [CrossRef]
- Vizza, C.; Sanderson, B.L.; Burrows, D.G.; Coe, H.J. The effects of ethanol preservation on fish fin stable isotopes: Does variation in C:N ratio and body size matter? Trans. Am. Fish. Soc. 2013, 142, 1469–1476. [Google Scholar] [CrossRef]
- Stallings, C.D.; Nelson, J.A.; Rozar, K.L.; Adams, C.S.; Wall, K.R.; Switzer, T.S.; Winner, B.L.; Hollander, D.J. Effects of preservation methods of muscle tissue from upper-trophic level reef fishes on stable isotope values (δ(13)C and δ(15)N). PeerJ 2015, 3, e874. [Google Scholar] [CrossRef] [Green Version]
- Kishe-Machumu, M.A.; van Rijssel, J.C.; Poste, A.; Hecky, R.E.; Witte, F. Stable isotope evidence from formalin–ethanol-preserved specimens indicates dietary shifts and increasing diet overlap in Lake Victoria cichlids. Hydrobiologia 2017, 791, 155–173. [Google Scholar] [CrossRef]
- Focken, U.; Becker, K. Metabolic fractionation of stable carbon isotopes: Implications of different proximate compositions for studies of the aquatic food webs using δ13C data. Oecologia 1998, 115, 337–343. [Google Scholar] [CrossRef]
- Logan, J.M.; Jardine, T.D.; Miller, T.J.; Bunn, S.E.; Cunjak, R.A.; Lutcavage, M.E. Lipid corrections in carbon and nitrogen stable isotope analyses: Comparison of chemical extraction and modelling methods. J. Anim. Ecol. 2008, 77, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Logan, J.M.; Lutcavage, M.E. A comparison of carbon and nitrogen stable isotope ratios of fish tissues following lipid extractions with non-polar and traditional chloroform/methanol solvent systems. Rapid Commun. Mass Spectrom. 2008, 22, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Svensson, E.; Schouten, S.; Hopmans, E.C.; Middelburg, J.J.; Sinninghe Damsté, J.S. Factors controlling the stable nitrogen isotopic composition (δ15N) of lipids in marine animals. PLoS ONE 2016, 11, e0146321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiljunen, M.; Grey, J.; Sinisalo, T.; Harrod, C.; Immonen, H.; Jones, R. A revised model for lipid-normalizing δ13C values from aquatic organisms, with implications for isotope mixing models. J. Appl. Ecol. 2006, 43, 1213–1222. [Google Scholar] [CrossRef]
- Edwards, M.S.; Turner, T.F.; Sharp, Z.D. Short and long-term effects of fixation and preservation on stable isotope values (δ13C, δ15N, δ34S) of fluid-preserved museum specimens. Copeia 2002, 4, 1106–1112. [Google Scholar] [CrossRef]
- Ahnesjö, I.; Craig, J.F. The biology of Syngnathidae: Pipefishes, seadragons and seahorses. J. Fish Biol. 2011, 78, 1597–1602. [Google Scholar] [CrossRef]
- Vincent, A.C.J.; Foster, S.J.; Koldewey, H.J. Conservation and management of seahorses and other Syngnathidae. J. Fish Biol. 2011, 78, 1681–1724. [Google Scholar] [CrossRef]
- Planas, M.; Silva, C.; Quintas, P.; Chamorro, A.; Piñero, S. Ongrowing and enhancement of n-3 HUFA profile in adult Artemia: Short vs. long-time enrichment. J. Appl. Phycol. 2017, 29, 1409–1420. [Google Scholar] [CrossRef]
- Planas, M.; Chamorro, A.; Quintas, P.; Vilar, A. Establishment and maintenance of threatened long-snouted seahorse, Hippocampus guttulatus, broodstock in captivity. Aquaculture 2008, 283, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Randazzo, B.; Rolla, L.; Ofelio, C.; Planas, M.; Gioacchini, G.; Vargas, A.; Giorgini, E.; Olivotto, I. The influence of diet on the early development of two seahorse species (H. guttulatus and H. reidi): Traditional and innovative approaches. Aquaculture 2018, 490, 75–90. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2014. Available online: http://www.R-project.org (accessed on 23 February 2018).
- Howell, D.C. Statistical Methods for Psychology; Wadsworth Publishing Company: Belmont, CA, USA, 2013; p. 770. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobson, K.A.; Gibbs, H.L.; Gloutney, M.L. Preservation of blood and tissue samples for stable-carbon and stable-nitrogen isotope analysis. Can. J. Zool. 1997, 75, 1720–1723. [Google Scholar] [CrossRef]
- Kelly, M.H.; Hagar, W.G.; Jardine, T.D.; Cunjak, R.A. Nonlethal sampling of sunfish and slimy sculpin for stable isotope analysis: How scale and fin tissue compare with muscle tissue. N. Am. J. Fish Manag. 2006, 26, 921–925. [Google Scholar] [CrossRef]
- Sarakinos, H.C.; Johnson, M.L.; Zanden, M.J.V. A synthesis of tissue-preservation effects on carbon and nitrogen stable isotope signatures. Can. J. Zool. 2002, 80, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Reich, K.J.; Bjorndal, K.A.; Martinez Del Rio, C. Effects of growth and tissue type on the kinetics of 13C and 15N incorporation in a rapidly growing ectotherm. Oecologia 2008, 155, 651–663. [Google Scholar] [CrossRef]
- Britton, J.R.; Busst, G.M.A. Stable isotope discrimination factors of omnivorous fishes: Influence of tissue type, temperature, diet composition and formulated feeds. Hydrobiologia 2018, 808, 219–234. [Google Scholar] [CrossRef] [Green Version]
- Tieszen, L.L.; Boutton, T.W.; Tesdahl, K.G.; Slade, N.A. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet. Oecologia 1983, 57, 32–37. [Google Scholar] [CrossRef]
- Vizzini, S.; Mazzola, A. Stable isotopes and trophic positions of littoral fishes from a Mediterranean marine protected area. Environ. Biol. Fishes 2009, 48, 13–25. [Google Scholar] [CrossRef]
- McCarthy, I.D.; Waldron, S. Identifying migratory Salmo trutta using carbon and nitrogen stable isotope ratios. Rapid Commun. Mass Spectrom. 2000, 14, 1325–1331. [Google Scholar] [CrossRef]
- Correia, M.; Campoy, A.; Madeira, C.; Andrade, J.P. Is filament clipping an effective tool for tissue sampling in Hippocampus guttulatus? Environ. Biol. Fish 2018, 101, 1517–1523. [Google Scholar] [CrossRef]
- Gristina, M.; Bertrandino, S.; Cardone, F.; Mentino, D.; Corriero, G.; Scillitani, G. Skin filament recovery after clipping in Hippocampus guttulatus: Behavioural and histological aspects. Aquat. Biol. 2017, 26, 149–157. [Google Scholar] [CrossRef]
- Woodall, L.C.; Jones, R.; Zimmerman, B.; Guillaume, S.; Stubbington, T.; Shaw, P.; Koldewey, H.J. Partial fin-clipping as an effective tool for tissue sampling seahorses, Hippocampus spp. J. Mar. Biol. Assoc. 2012, 92, 1427–1432. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Mechanism of carbon isotope fractionation associated with lipid synthesis. Science 1977, 197, 261–263. [Google Scholar] [CrossRef]
- Post, D.M.; Craig, A.; Layman, D.; Arrington, D.A.; Takimoto, G.; Quatrocchi, J.; Montaña, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Sotiropoulos, M.A.; Tonn, W.M.; Wassenaar, L.I. Effects of lipid extraction on stable carbon and nitrogen isotope analyses of fish tissues: Potential consequences for food web studies. Ecol. Freshw. Fish 2004, 13, 155–160. [Google Scholar] [CrossRef]
- Sweeting, C.J.; Polunin, N.V.C.; Jennings, S. Effects of chemical lipid extraction and arithmetic lipid correction on stable isotope ratios of fish tissues. Rapid Commun. Mass Spectrom. 2006, 20, 595–601. [Google Scholar] [CrossRef]
- Perga, M.E.; Gerdeaux, D. Are fish what they eat’ all year round? Oecologia 2005, 144, 598–606. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ANCOVA | Group Comparisons (p) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Effect | SS | d.f. | MS | F | Adj. p | FR–ET | FR–FO | ET–FO | |
| Prey | |||||||||
| C:N | Treatment | 6.90 | 2 | 3.45 | 41.43 | <0.001 | <0.001 | 0.403 | <0.001 |
| Prey | 18.83 | 5 | 3.77 | 45.19 | <0.001 | ||||
| Residuals | 2.33 | 28 | 0.08 | ||||||
| δ13C | Treatment | 28.39 | 2 | 14.19 | 199.00 | <0.001 | <0.001 | <0.001 | <0.001 |
| Prey | 186.41 | 5 | 37.28 | 522.80 | <0.001 | ||||
| Residuals | 2.00 | 28 | 0.07 | ||||||
| δ15N | Treatment | 0.50 | 2 | 0.24 | 2.75 | 0.081 | 0.027 | 0.311 | 0.201 |
| Prey | 435.30 | 5 | 87.05 | 990.40 | <0.001 | ||||
| Residuals | 2.50 | 28 | 0.09 | ||||||
| Fins | |||||||||
| C:N | Treatment | 0.570 | 2 | 0.28 | 13.01 | <0.001 | <0.001 | 0.384 | <0.001 |
| Genus | 0.310 | 1 | 0.31 | 14.15 | <0.001 | ||||
| Residuals | 1.206 | 55 | 0.02 | ||||||
| δ13C | Treatment | 9.26 | 2 | 4.63 | 3.70 | 0.031 | 0.192 | 0.365 | 0.037 |
| Genus | 127.92 | 1 | 127.92 | 102.11 | <0.001 | ||||
| Residuals | 68.90 | 55 | 1.25 | ||||||
| δ15N | Treatment | 0.38 | 2 | 0.19 | 0.10 | 0.901 | 0.712 | 0.718 | 0.998 |
| Genus | 42.74 | 1 | 42.74 | 23.67 | <0.001 | ||||
| Residuals | 92.09 | 55 | 1.81 | ||||||
| Newborns | |||||||||
| C:N | Treatment | 7.37 | 2 | 3.68 | 332.51 | <0.001 | <0.001 | <0.001 | <0.001 |
| Species | 0.01 | 2 | 0.01 | 0.69 | 0.506 | ||||
| Residuals | 0.443 | 40 | 0.010 | ||||||
| δ13C | Treatment | 36.75 | 2 | 18.38 | 4.74 | 0.014 | 0.040 | 0.391 | 0.005 |
| Species | 118.48 | 2 | 59.24 | 15.29 | <0.001 | ||||
| Residuals | 154.99 | 40 | 3.87 | ||||||
| δ15N | Treatment | 1.00 | 2 | 0.49 | 0.05 | 0.955 | 0.801 | 0.787 | 0.986 |
| Species | 613.90 | 2 | 306.96 | 28.38 | <0.001 | ||||
| Residuals | 427.30 | 40 | 10.68 | ||||||
| Conversion Model (Linear Regression) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | Model | S.E. | S.E. | |||||||
| y-x | n | F | p | y = ax + b | a | b | Adj R2 | β | ||
| Prey | ||||||||||
| C:N | FR-ET | 12 | 736 | <0.017 | y = 1.201x | I | 0.044 | 0.894 | 0.993 | |
| FR-FO | 12 | 104 | <0.001 | y = 1.154x − 0.854 | I | 0.113 | 0.562 | 0.903 | 0.955 | |
| δ13C | FR-ET | 12 | 19,070 | <0.001 | y = 1.061 | S,I | 0.008 | 0.908 | 0.999 | |
| FR-FO | 12 | 2009 | <0.001 | y = 1.006x + 1.222 | S,I | 0.022 | 0.449 | 0.995 | 0.998 | |
| δ15N | FR-ET | 12 | 2138 | <0.001 | y = 0.971x | I | 0.021 | 0.903 | 0.997 | |
| FR-FO | 12 | 2519 | <0.001 | y = 0.990x | I | 0.020 | 0.904 | 0.998 | ||
| Dorsal fins | ||||||||||
| C:N | FR-ET | 17 | 7510 | <0.001 | y = 1.059x | I | 0.012 | 0.935 | 0.999 | |
| FR-FO | 17 | 23,903 | <0.001 | y = 0.989x | I | 0.006 | 0.937 | 0.908 | ||
| δ13C | FR-ET | 14 | 865.0 | <0.001 | y = 0.918x − 1.617 | S,I | 0.031 | 0.466 | 0.985 | 0.993 |
| FR-FO | 16 | 30,169 | <0.001 | y = 0.969x | I | 0.006 | 0.999 | 0.999 | ||
| δ15N | FR-ET | 15 | 107,259 | <0.001 | y = 0.997x | I | 0.003 | 0.928 | 0.999 | |
| FR-FO | 16 | 50,677 | <0.001 | y = 1.000x | 0.004 | 0.923 | 0.999 | |||
| Newborn | ||||||||||
| C:N | FR-ET | 20 | 32,140 | <0.001 | y = 1.226x | I | 0.007 | 0.999 | 0.999 | |
| FR-FO | 21 | 22,370 | <0.001 | y = 0.349x + 2.338 | I | 0.096 | 0.384 | 0.384 | 0.999 | |
| δ13C | FR-ET | 20 | 2045 | <0.001 | y = 0.961x − 2.178 | I | 0.021 | 0.407 | 0.991 | 0.996 |
| FR-FO | 21 | 4622 | <0.001 | y = 1.024x + 1.166 | I | 0.015 | 0.317 | 0.996 | 0.998 | |
| δ15N | FR-ET | 20 | 146,531 | <0.001 | y = 0.984x | 0.003 | 0.999 | 0.999 | ||
| FR-FO | 21 | 12,356 | <0.001 | y = 0.982x − 0.528 | I | 0.003 | 0.315 | 0.999 | 0.999 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Planas, M.; Paltrinieri, A.; Carneiro, M.D.D.; Hernández-Urcera, J. Effects of Tissue Preservation on Carbon and Nitrogen Stable Isotope Signatures in Syngnathid Fishes and Prey. Animals 2020, 10, 2301. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10122301
Planas M, Paltrinieri A, Carneiro MDD, Hernández-Urcera J. Effects of Tissue Preservation on Carbon and Nitrogen Stable Isotope Signatures in Syngnathid Fishes and Prey. Animals. 2020; 10(12):2301. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10122301
Chicago/Turabian StylePlanas, Miquel, Alex Paltrinieri, Mario Davi Dias Carneiro, and Jorge Hernández-Urcera. 2020. "Effects of Tissue Preservation on Carbon and Nitrogen Stable Isotope Signatures in Syngnathid Fishes and Prey" Animals 10, no. 12: 2301. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10122301