
Effects of Quercitrin on PRV-Induced Secretion of Reactive Oxygen Species and Prediction of lncRNA Regulatory Targets in 3D4/2 Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagent, Cells and Viruses
2.2. Assessment of Cell Viability
2.3. Titrating TCID50 of PRV
2.4. Detection of Intracellular ROS by Flow Cytometry
2.5. Sequencing and Analysis RNA
2.6. Verification of Differential Expression RNA
2.7. Calculation and Statistical Analysis
3. Results
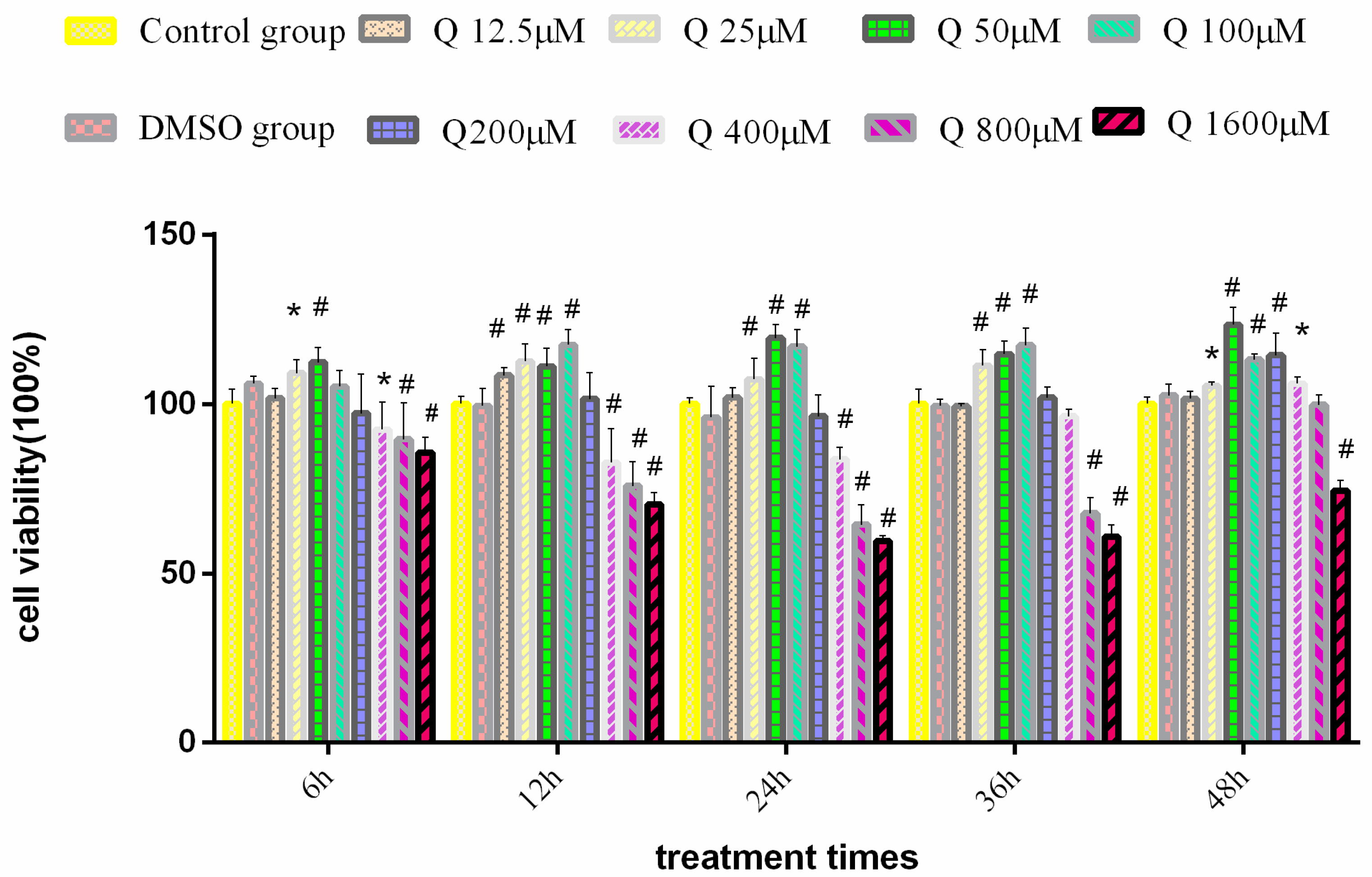
3.1. Effect of Quercitrin on the 3D4/2 Cells’ Viability
3.2. The TCID50 of PRV
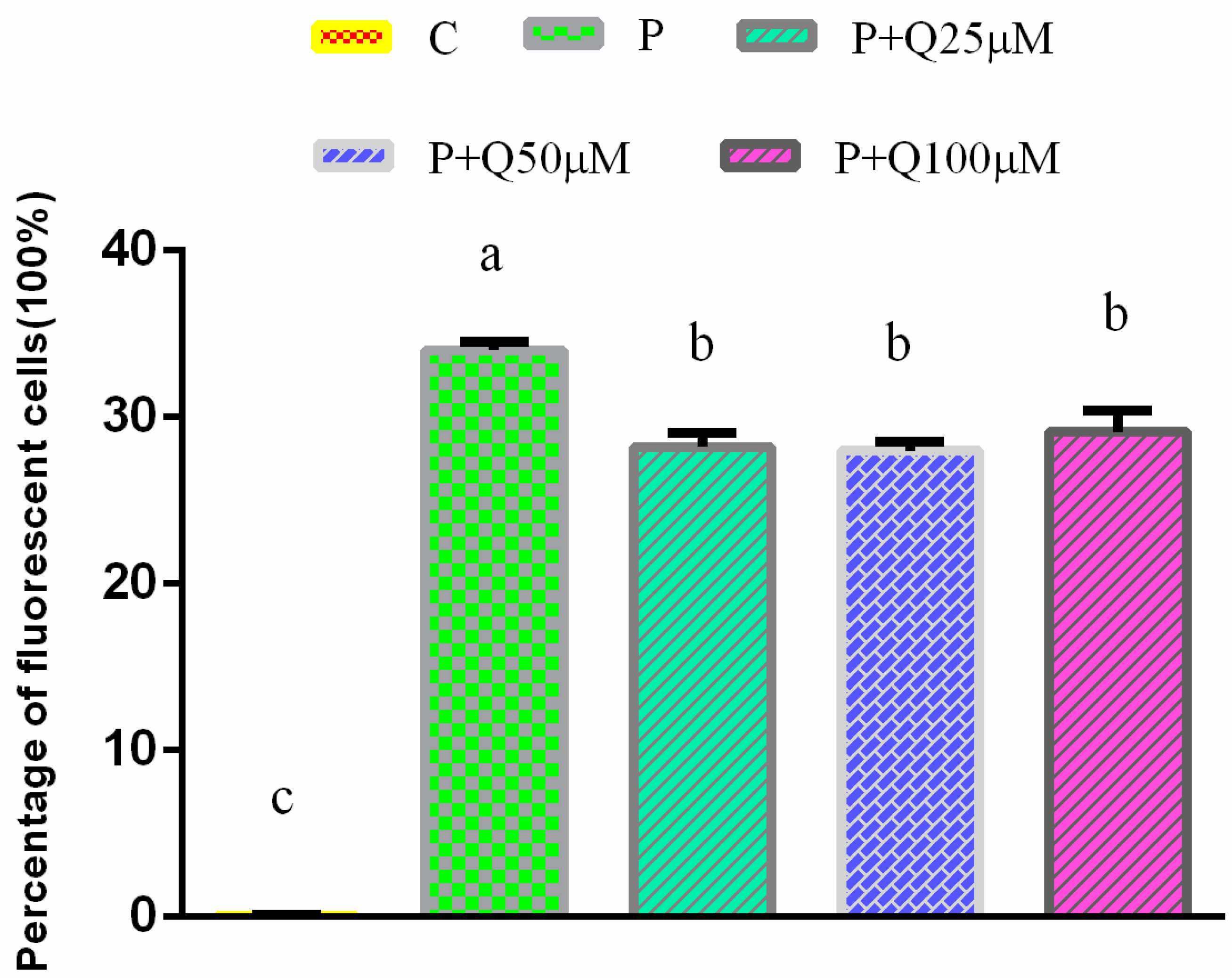
3.3. Effect of PRV Infection on ROS Production in 3D4/2 Cells
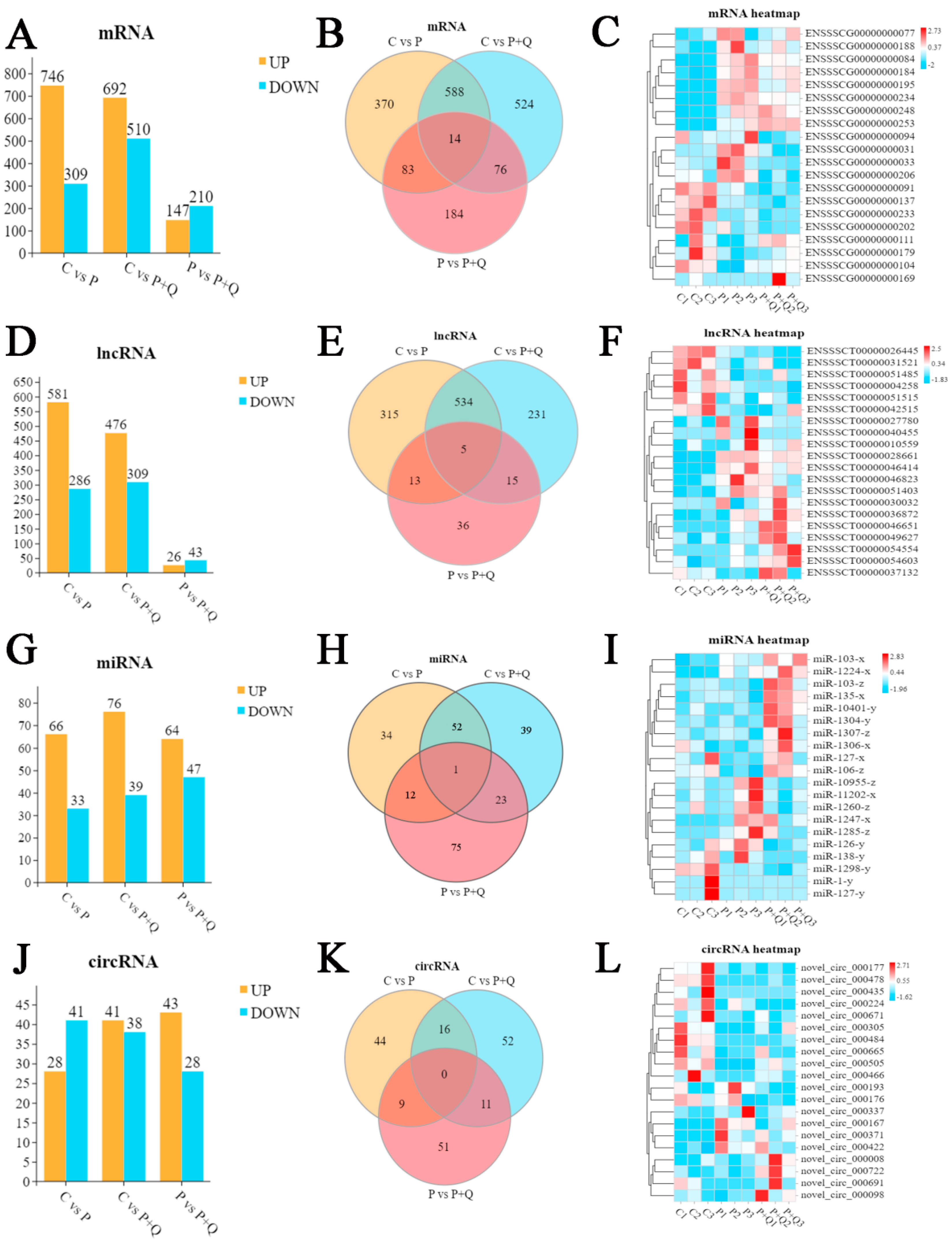
3.4. The Differential Expression of mRNA, lncRNA, miRNA and circRNA
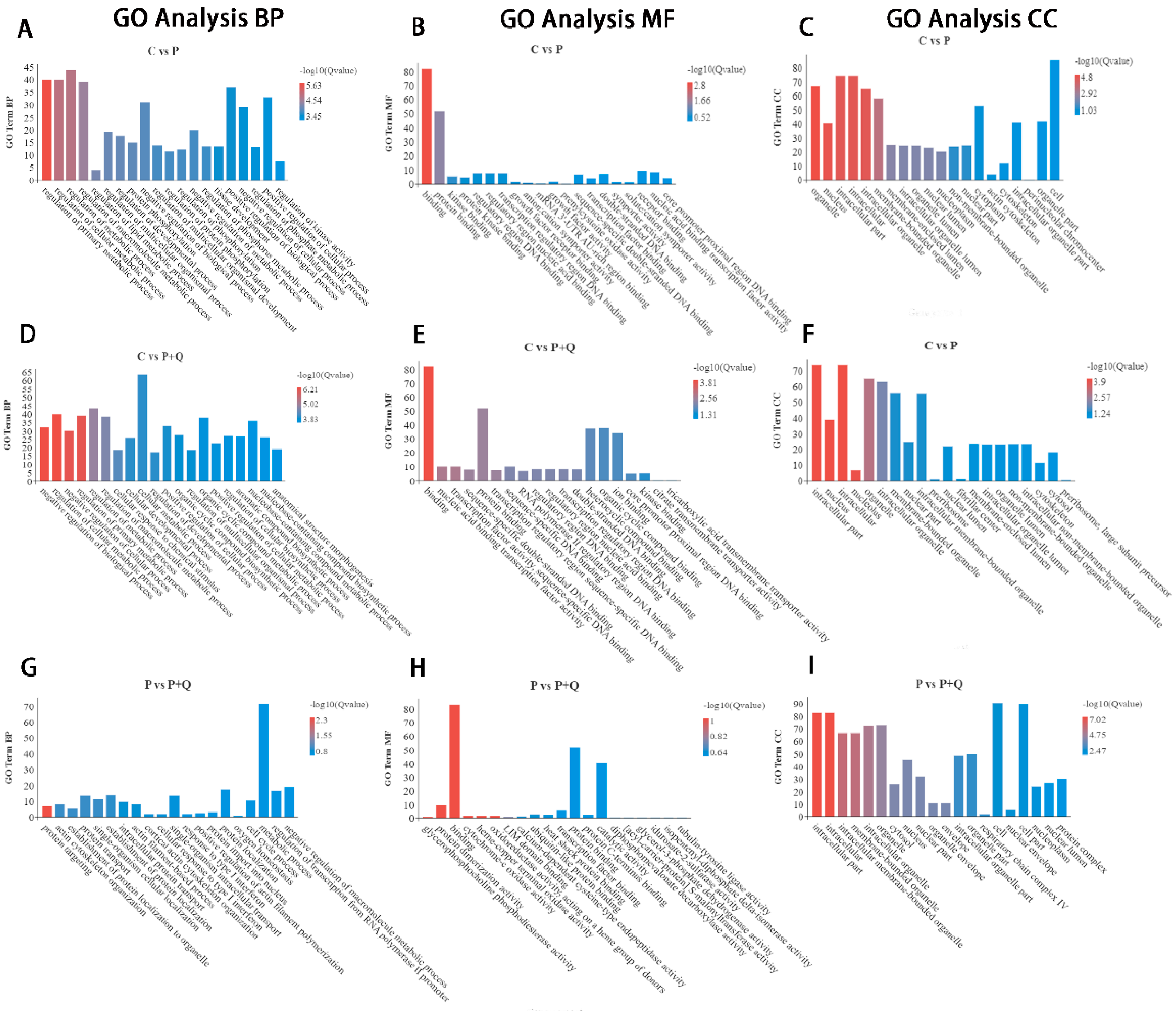
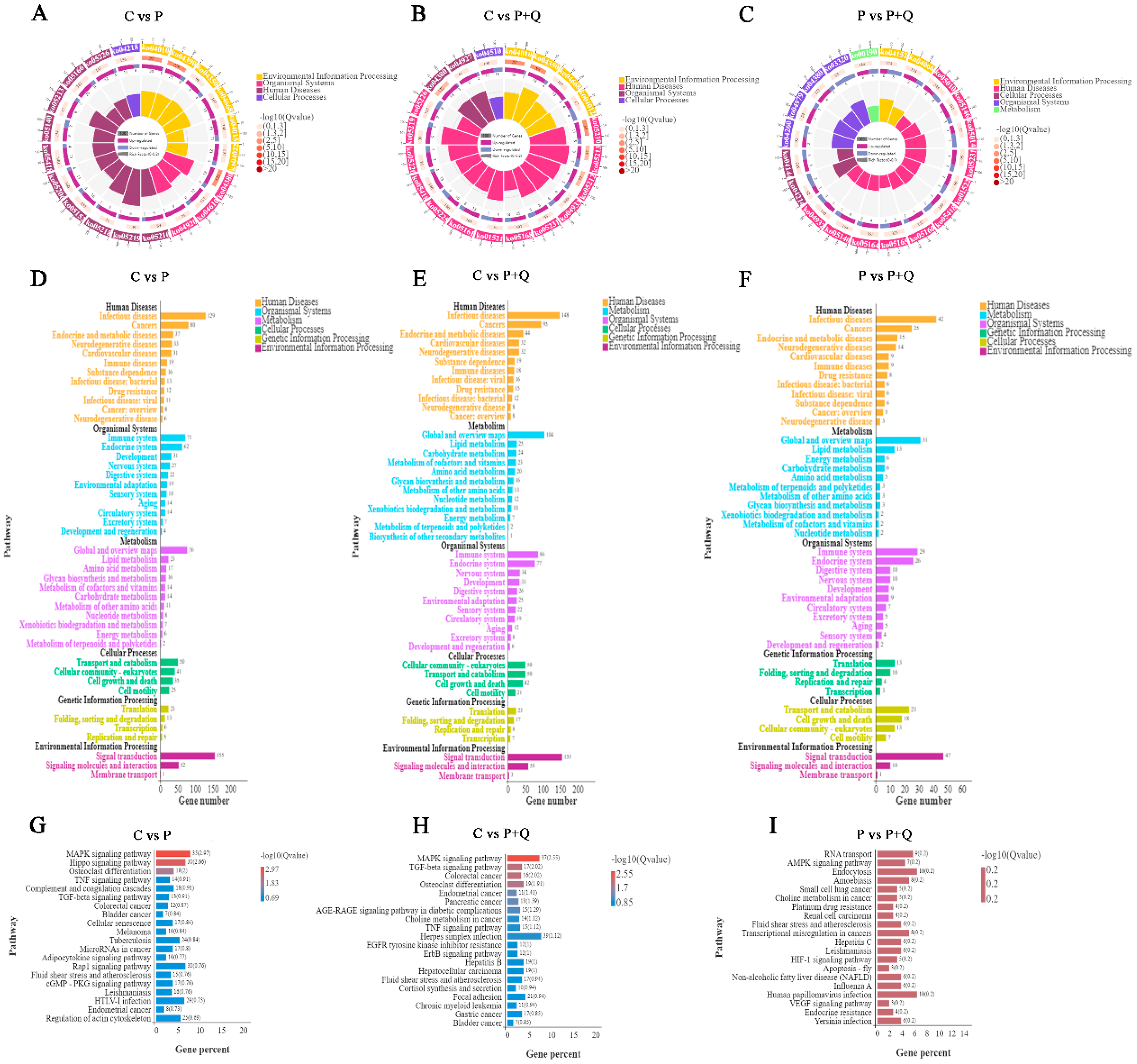
3.5. GO Analyses of DE mRNAs
3.6. KEGG Analyses of DE mRNAs
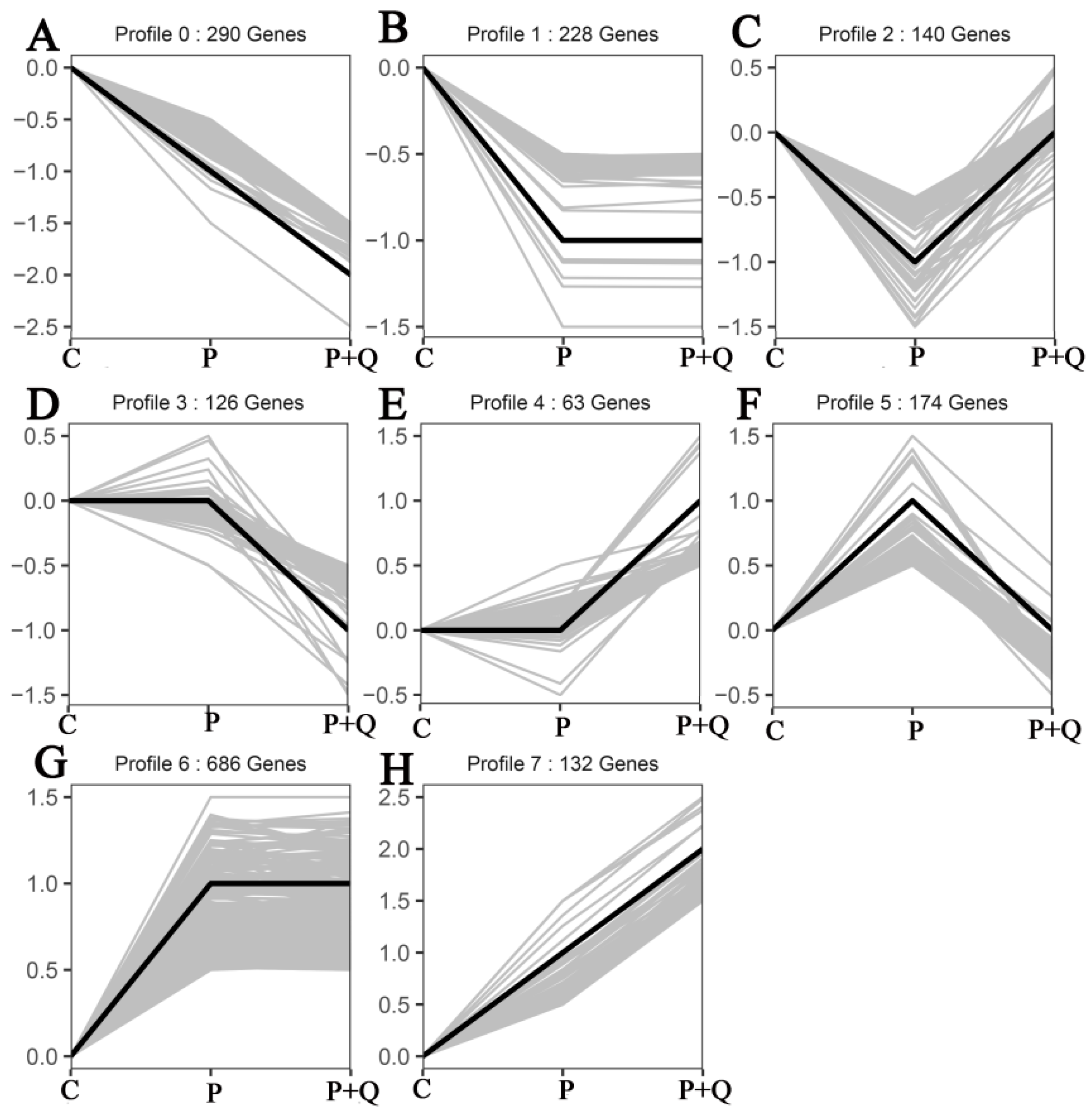
3.7. Trend Analysis of mRNAs
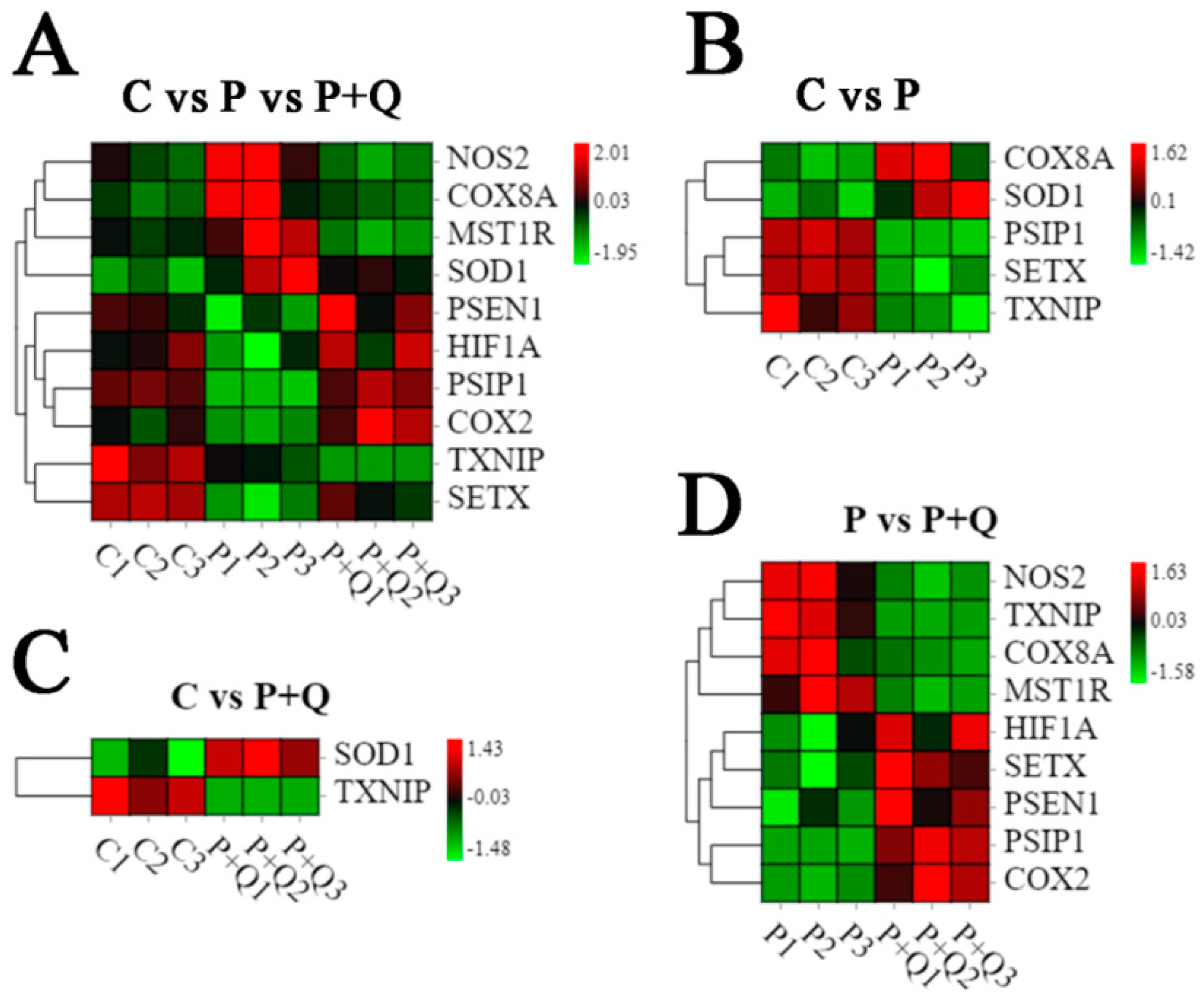
3.8. Analysis of DE mRNAs Related to Oxidative Stress and Antioxidant Stress
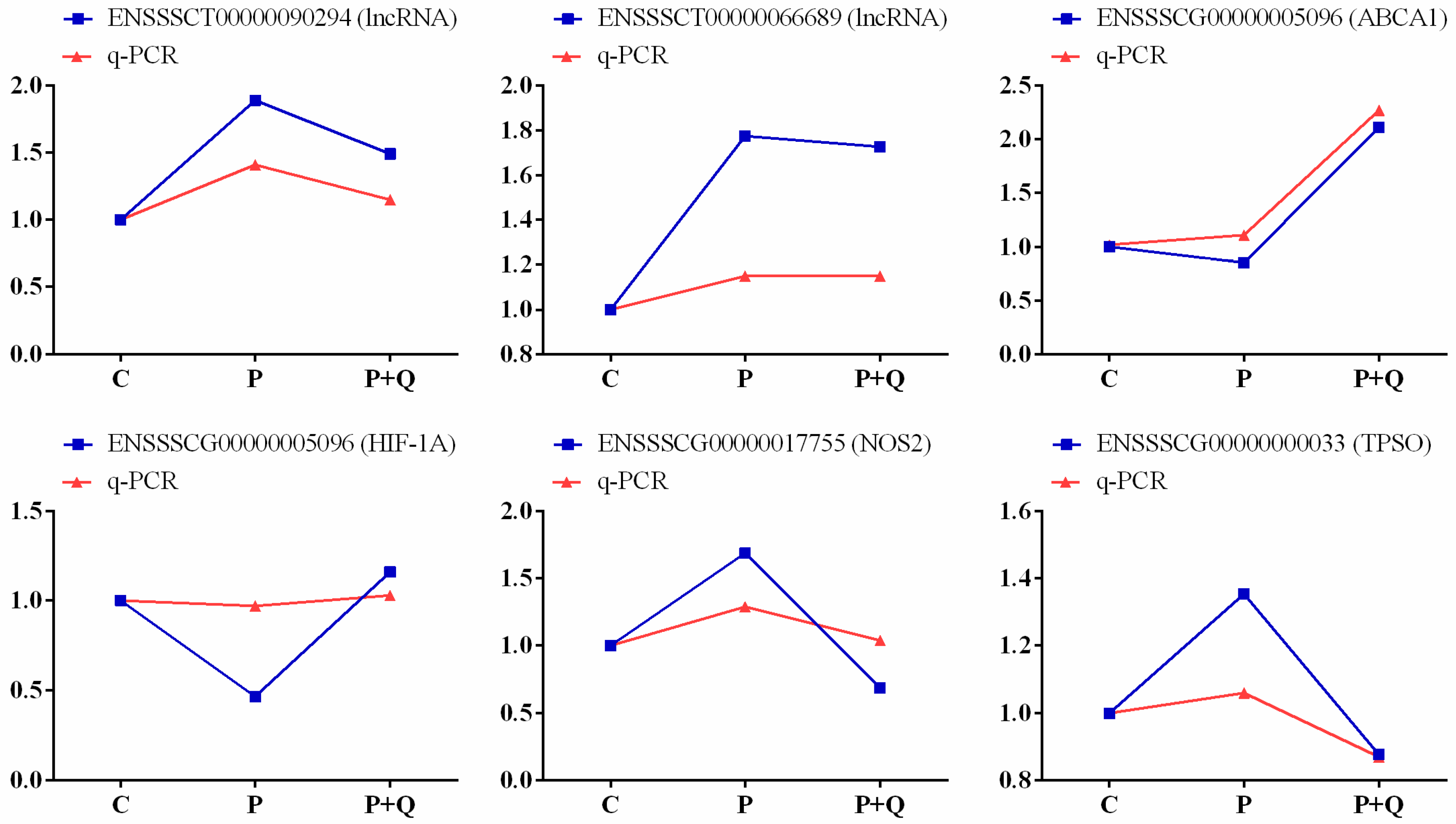
3.9. Validation of lncRNA and mRNA
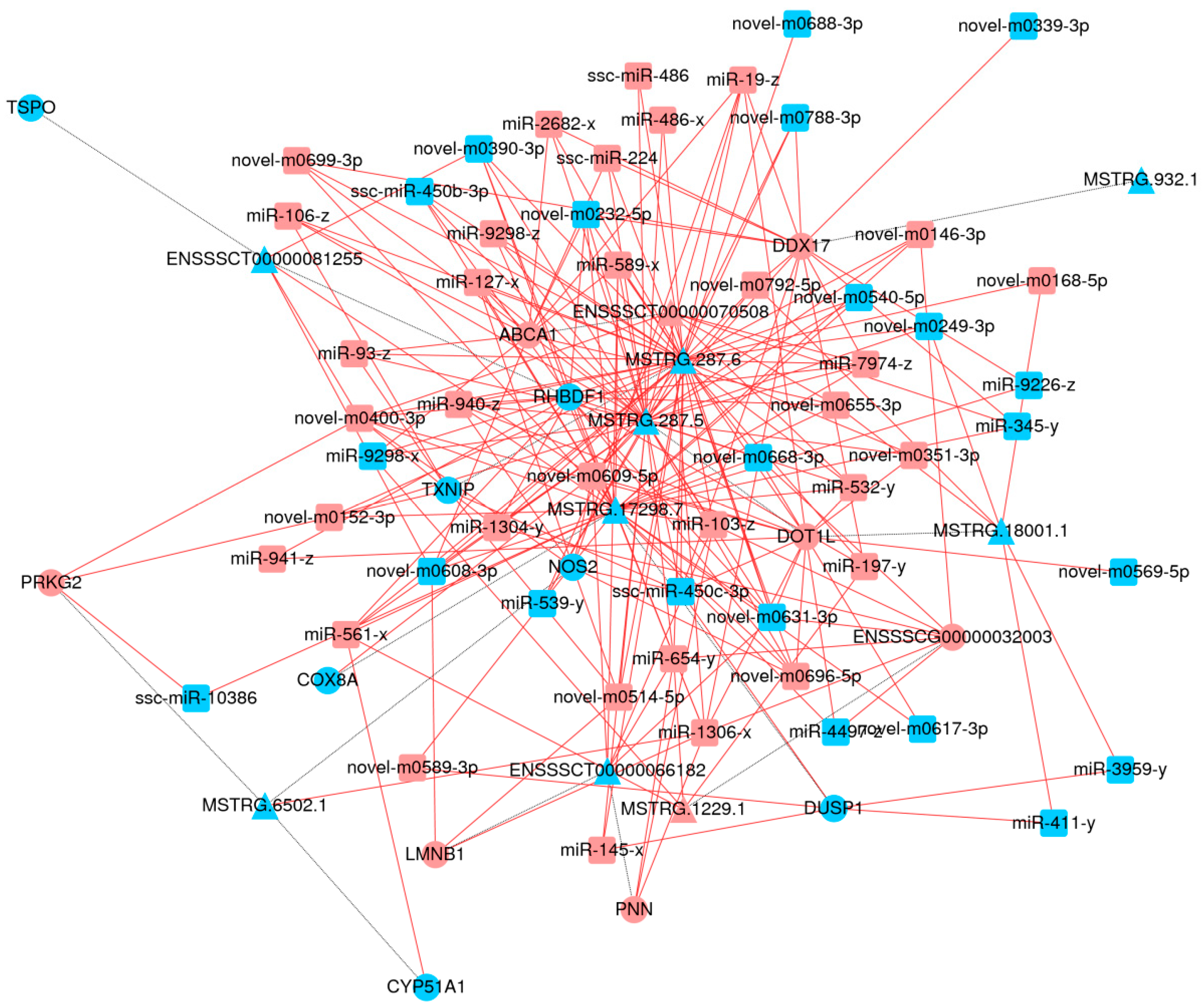
3.10. The Interaction between lncRNAs, mRNAs and miRNAs in PRV + Quercitrin Group and PRV Group
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mettenleiter, T.C. Aujeszky’s disease (pseudorabies) virus: The virus and molecular pathogenesis-State of the art, June 1999. Vet. Res. 2000, 31, 99–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, G.; Lu, J.; Zhang, W.; Gao, G.F. Pseudorabies virus: A neglected zoonotic pathogen in humans? Emerg. Microbes Infect. 2019, 8, 150–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Auclert, L.Z.; Zhai, X.; Wong, G.; Zhang, C.; Zhu, H.; Su, S. Interspecies transmission, genetic diversity, and evolutionary dynamics of Pseudorabies virus. J. Infect. Dis. 2018, 219, 1705–1715. [Google Scholar] [CrossRef]
- Liu, M.; Chen, F.; Liu, T.; Chen, F.; Liu, S.; Yang, J. The role of oxidative stress in influenza virus infection. Microbes Infect. 2017, 19, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Lee, C. Therapeutic modulation of virus-induced oxidative stress via the Nrf2-dependent antioxidative pathway. Oxid. Med. Cell Longev. 2018, 2018, 6208067. [Google Scholar] [CrossRef]
- Lai, I.H.; Chang, C.D.; Shih, W.L. Apoptosis induction by Pseudorabies virus via oxidative stress and subsequent DNA damage signaling. Intervirology 2019, 62, 116–123. [Google Scholar] [CrossRef]
- Zhai, N.; Liu, K.; Li, H.; Liu, Z.; Wang, H.; Korolchuk, V.I.; Chen, X. PCV2 replication promoted by oxidative stress is dependent on the regulation of autophagy on apoptosis. Vet. Res. 2019, 50, 19. [Google Scholar] [CrossRef] [Green Version]
- Bauer, M.E.; Fuente, M.D.L. The role of oxidative and inflammatory stress and persistent viral infections in immunosenescence. Mech. Ageing Dev. 2016, 158, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Ohl, K.; Tenbrock, K. Reactive oxygen species as regulators of MDSC-mediated immune suppression. Front. Immunol. 2018, 9, 2499. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Mishra, K.P.; Ganju, L. Broad-spectrum antiviral properties of andrographolide. Arch. Virol. 2017, 162, 611–623. [Google Scholar] [CrossRef]
- Song, M.; Chen, Y.; Du, H.; Zhang, S.; Wang, Y.; Zeng, L.; Liu, J. Raw Rehmannia radix polysaccharide can effectively release peroxidative injury induced by duck hepatitis a virus. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 8–21. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.F.; Jiang, L.H.; Zhao, W.D.; Xi-Nan, M.; Huang, S.Q.; Yang, J.; Chen, H.L. Immunomodulatory and antioxidant effects of total flavonoids of Spatholobus suberectus Dunn on PCV2 infected mice. Sci Rep. 2017, 7, 8676. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.J.; Tao, J.Y.; Yang, J.; Zeng, Y.; Wei, Y.Y.; Hu, T.J. Effects of flavonoids from Polygonum hydropiper on levels of reactive oxygen species and inflammatory factors in RAW264.7 cells induced by lipopolysaccharide. Prog. Vet. Med. 2017, 38, 1–6. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.M.M.; Bast, A. Health effects of quercitrin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Formica, J.V.; Regelson, W. Review of the biology of quercitrin and related bioflavonoids. Food Chem. Toxicol. 1995, 33, 1061–1080. [Google Scholar] [CrossRef]
- Goliomytis, M.; Tsoureki, D.; Simitzis, P.E.; Charismiadou, M.A.; Hager-Theodorides, A.L.; Deligeorgis, S.G. The effects of quercitrin dietary supplementation on broiler growth performance, meat quality, and oxidative stability. Poult. Sci. 2014, 93, 1957–1962. [Google Scholar] [CrossRef]
- Nambooppha, B.; Photichai, K.; Wongsawan, K.; Chuammitri, P. Quercitrin manipulates the expression of genes involved in the reactive oxygen species (ROS) process in chicken heterophils. J. Vet. Med. Sci. 2018, 80, 1204–1211. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zhou, Y.; Li, H. LncRNA, miRNA and lncRNA-miRNA interaction in viral infection. Virus Res. 2018, 257, 25–32. [Google Scholar] [CrossRef]
- Wang, P.; Xu, J.; Wang, Y.; Cao, X. An interferon-independent lncRNA promotes viral replication by modulating cellular metabolism. Science 2017, 358, 1051–1055. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Zhang, K.; Huang, W.; Tang, W.; Li, H.; Dong, W.; Zhou, J. Identification of functional lncRNAs in pseudorabies virus type II infected cells. Vet. Microbiol. 2020, 242, 108564. [Google Scholar] [CrossRef]
- Gong, W.; Li, J.; Zhu, G.; Wang, Y.; Zheng, G.; Kan, Q. Chlorogenic acid relieved oxidative stress injury in retinal ganglion cells through IncRNA-TUG1/Nrf2. Cell Cycle 2019, 18, 1549–1559. [Google Scholar] [CrossRef]
- Yoshihara, E. TXNIP/TBP-2: A Master Regulator for Glucose Homeostasis. Antioxidants 2020, 9, 765. [Google Scholar] [CrossRef]
- Alhawiti, N.M.; Al Mahri, S.; Aziz, M.A.; Malik, S.S.; Mohammad, S. TXNIP in Metabolic regulation: Physiological role and therapeutic outlook. Curr. Drug Targets 2017, 18, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ning, J.; Cao, W.; Wang, S.; Du, T.; Jiang, J.; Zhang, B. Research progress of TXNIP as a tumor suppressor gene participating in the metabolic reprogramming and oxidative stress of cancer cells in various cancers. Front. Oncol. 2020, 10, 568574. [Google Scholar] [CrossRef]
- Cho, M.J.; Yoon, S.J.; Kim, W.; Park, J.; Lee, J.; Park, J.G.; Min, J.K. Oxidative stress-mediated TXNIP loss causes RPE dysfunction. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef]
- Rao, K.M. Molecular mechanisms regulating iNOS expression in various cell types. J. Toxicol. Environ. Health B Crit. Rev. 2000, 3, 27–58. [Google Scholar] [CrossRef]
- Gao, Q.; Liu, Y.; Wu, Y.; Zhao, Q.; Wang, L.; Gao, S.; Yuan, Z. IL-17 intensifies IFN-gamma-induced NOS2 upregulation in RAW 264.7 cells by further activating STAT1 and NF-kappaB. Int. J. Mol. Med. 2016, 37, 347–358. [Google Scholar] [CrossRef]
- Thomas, D.D.; Wink, D.A. NOS2 as an Emergent player in progression of cancer. Antioxid. Redox Signal. 2017, 26, 963–965. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ROS Probe | PRV | Quercitrin |
|---|---|---|---|
| Negative control | − | − | − |
| Control(C) | + | − | − |
| P | + | + | − |
| P + Q 25 μM | + | + | + |
| P + Q 50 μM | + | + | + |
| P + Q 100 μM | + | + | + |
| Target Gene | Accession | Sequence (5′–3′) | Amplicon Size (bp) | Genbank |
|---|---|---|---|---|
| lncRNA1 F | TGAATGCGGTTAGTCCTTGGTTCC | 148 | ENSSSCT00000090294 | |
| lncRNA1 F | GACGACGACACAGGCAGAGTAAG | |||
| lncRNA2 R | GAGATTCGTGGCTGCTGTGAGTAG | 108 | ENSSSCT00000066689 | |
| lncRNA2 R | GAAGCGTGGGCGAGGAAGAAC | |||
| ABCA1 F | 100152112 | CGCCTCCTTCGTGTTCAAGATCC | 82 | ENSSSCG00000005423 |
| ABCA1 R | ACTGCCATTGATGCCGATGAAGAG | |||
| HIF1A F | 396696 | CATTTCCATCTCCTCCCCACGTA | 170 | ENSSSCG00000005096 |
| HIF1A R | ACTCAAAGCGACAGATAACACA | |||
| NOS2 F | 396859 | TACCCCACCAGACGAGCTTC | 122 | ENSSSCG00000017755 |
| NOS2 R | CTATCTCCTTTGTTACCGCTTCC | |||
| TPSO F | 396592 | CTCACGCAATGTCCTCGGAA | 134 | ENSSSCG00000000033 |
| TPSO R | TCATGTAGGAGCCATACCCCAT | |||
| β-actin F | 414396 | GATGAGATTGGCATGGCTTT | 101 | ENSSSCG00000007585 |
| β-actin R | CACCTTCACCGTTCCAGTTT |
| PRV Dilution | Number of CPE Holes Present | Number of No CPE Holes Appear | Accumulative Total | Percentage of CPE Holes Present (%) | |
|---|---|---|---|---|---|
| No CPE | CPE | ||||
| 10−1 | 8 | 0 | 0 | 42 | 100 |
| 10−2 | 8 | 0 | 0 | 34 | 100 |
| 10−3 | 8 | 0 | 0 | 26 | 100 |
| 10−4 | 8 | 0 | 0 | 18 | 100 |
| 10−5 | 8 | 0 | 0 | 10 | 100 |
| 10−6 | 1 | 7 | 7 | 2 | 22.2 |
| 10−7 | 1 | 7 | 14 | 1 | 6.7 |
| 10−8 | 0 | 8 | 22 | 0 | 0 |
| 10−9 | 0 | 8 | 30 | 0 | 0 |
| 10−10 | 0 | 8 | 38 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Xie, X.; Chen, Q.; Yi, S.; Chen, J.; Xiao, Q.; Yu, M.; Wei, Y.; Hu, T. Effects of Quercitrin on PRV-Induced Secretion of Reactive Oxygen Species and Prediction of lncRNA Regulatory Targets in 3D4/2 Cells. Antioxidants 2022, 11, 631. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040631
Wang Q, Xie X, Chen Q, Yi S, Chen J, Xiao Q, Yu M, Wei Y, Hu T. Effects of Quercitrin on PRV-Induced Secretion of Reactive Oxygen Species and Prediction of lncRNA Regulatory Targets in 3D4/2 Cells. Antioxidants. 2022; 11(4):631. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040631
Chicago/Turabian StyleWang, Qiuhua, Xiaodong Xie, Qi Chen, Shouli Yi, Jiaji Chen, Qi Xiao, Meiling Yu, Yingyi Wei, and Tingjun Hu. 2022. "Effects of Quercitrin on PRV-Induced Secretion of Reactive Oxygen Species and Prediction of lncRNA Regulatory Targets in 3D4/2 Cells" Antioxidants 11, no. 4: 631. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11040631