Genetic Diversity and Identification of Vietnamese Paphiopedilum Species Using DNA Sequences

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Collecting of Sequence Data
2.4. Data Analysis
3. Results and Discussion
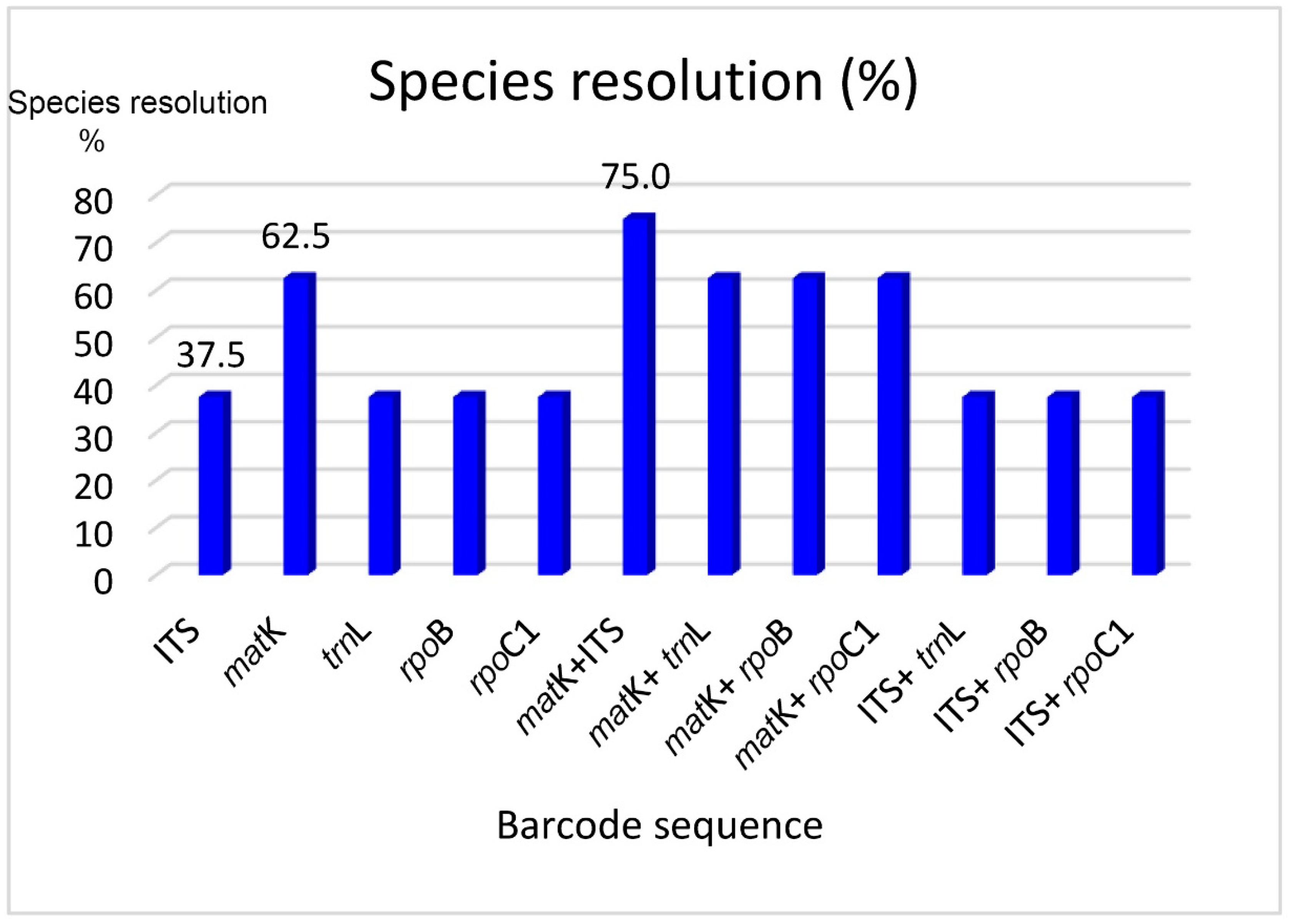
3.1. Identification Loci in Vietnamese Endemic Paphiopedilum Species
3.2. Effects of Different Bioinformatic Tools on the Identification of the Vietnamese Paphiopedilum Population
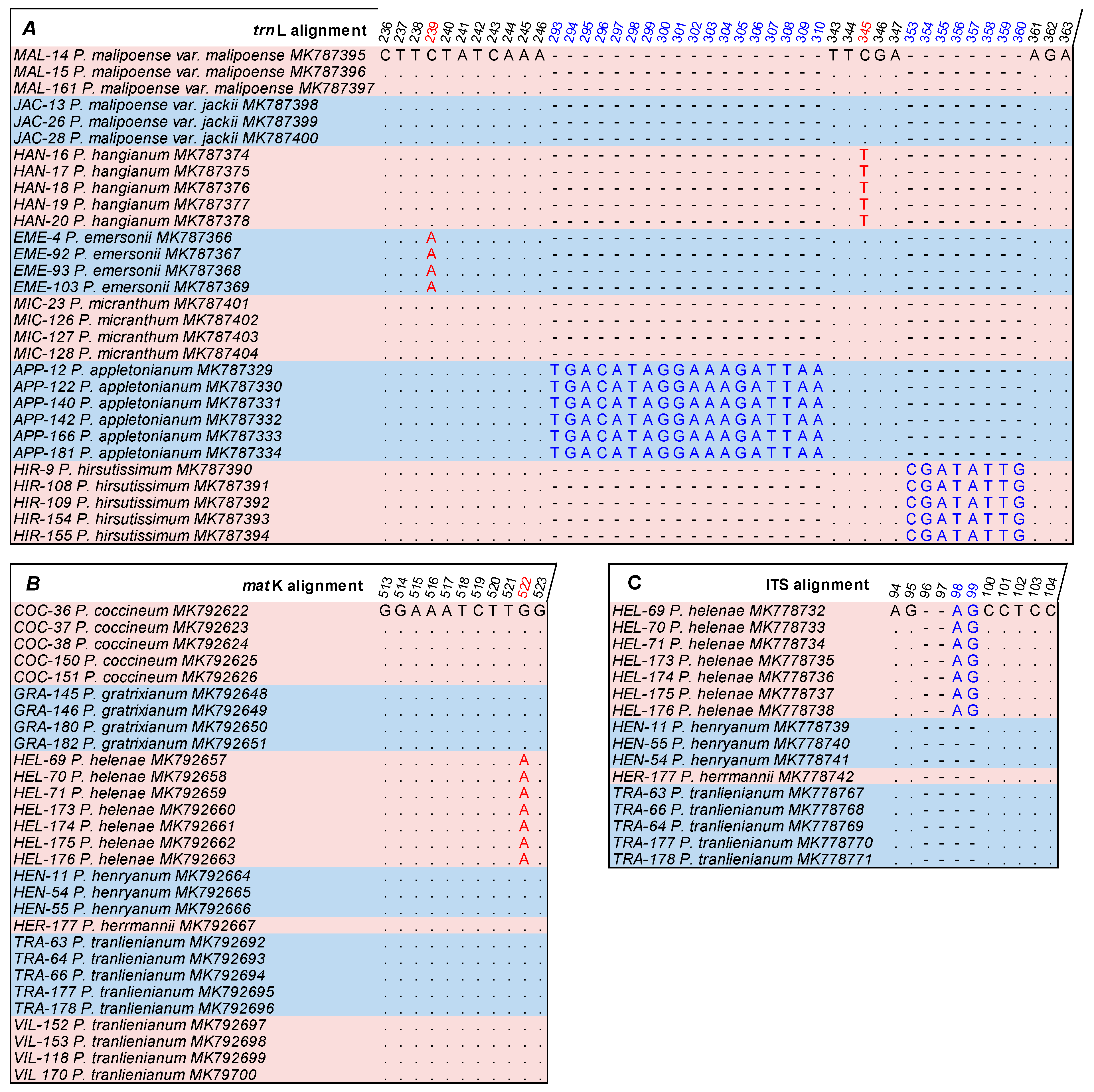
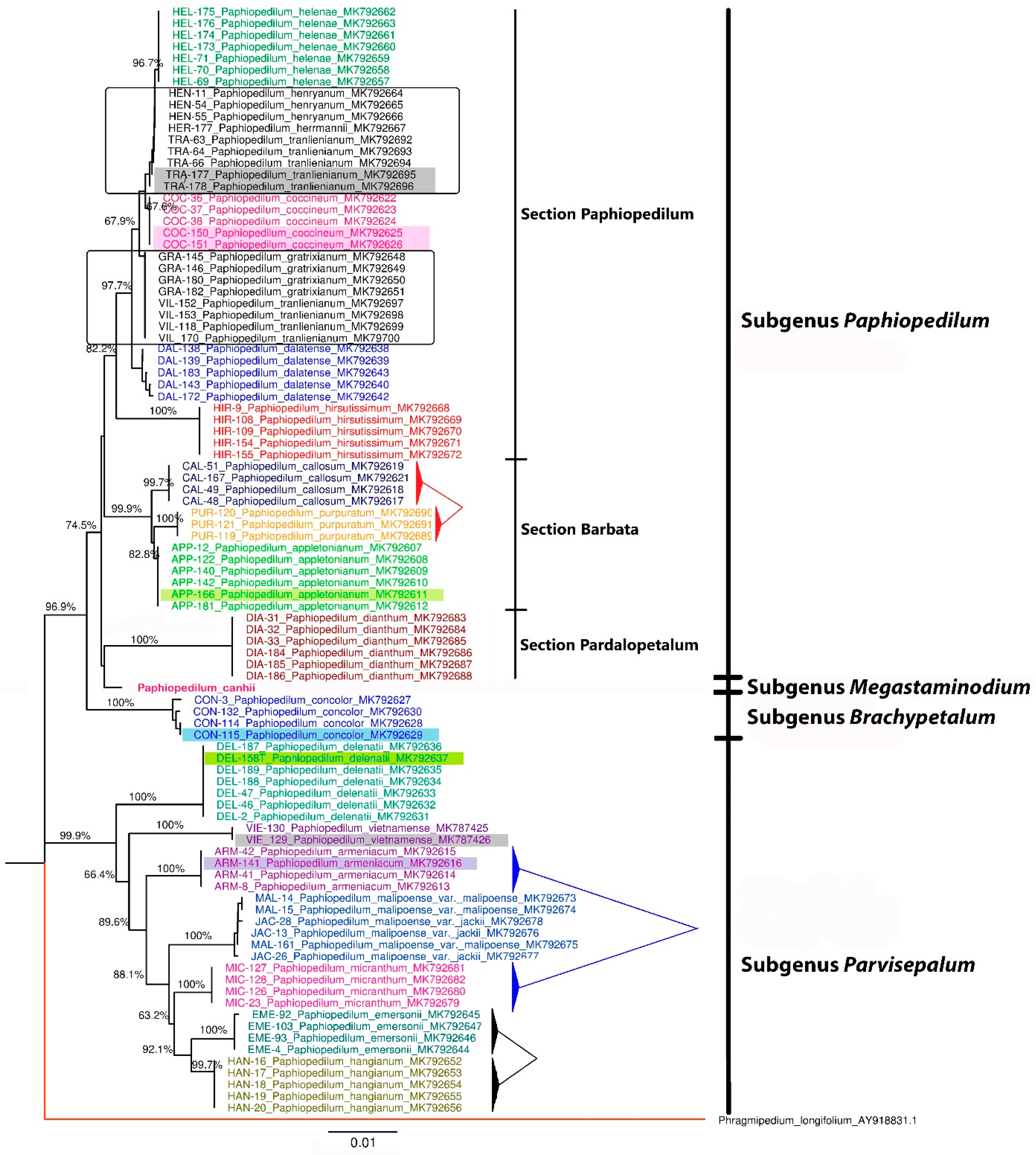
3.3. Genetic Diversity and Identification of the Paphiopedilum Population of Vietnam
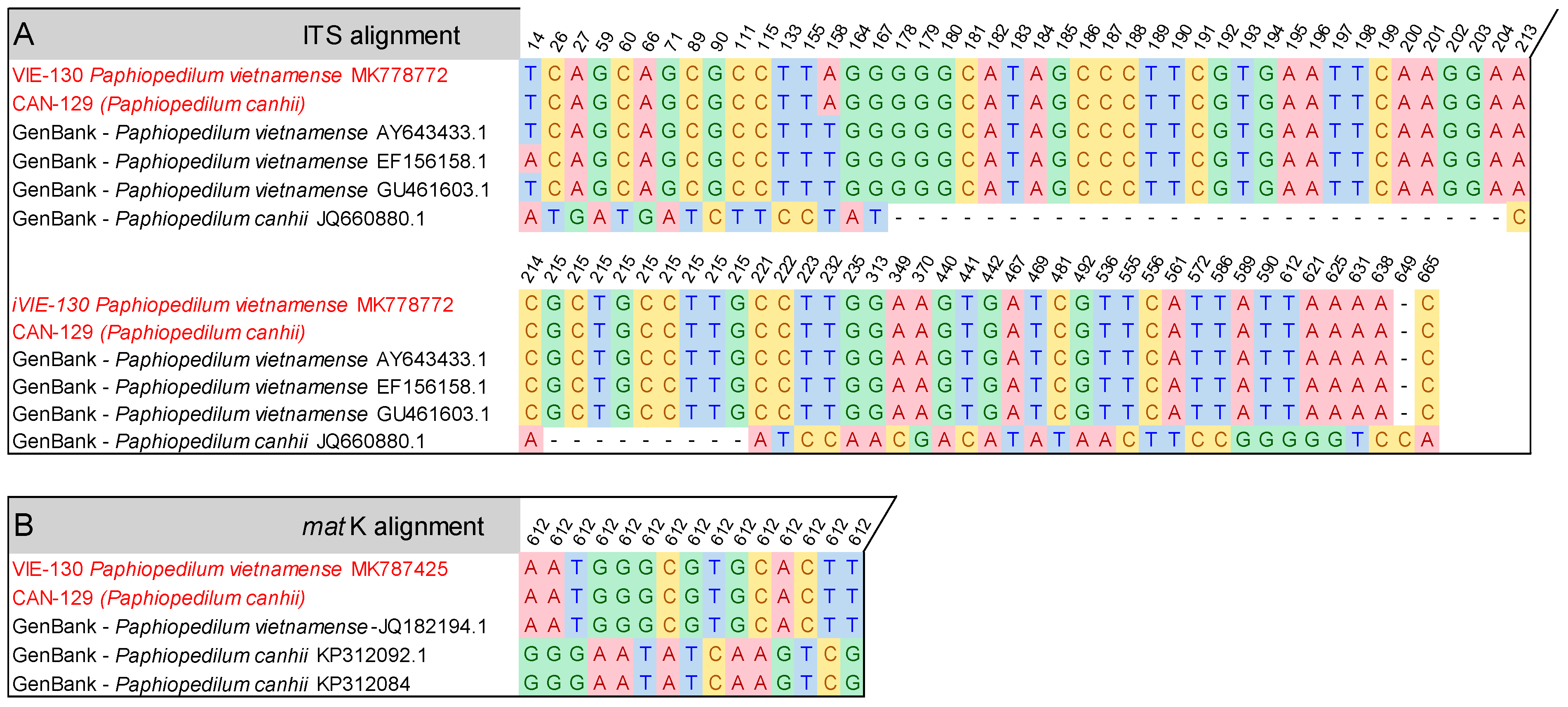
3.4. Application in the Identification of Trading Paphiopedilum Samples
3.5. The Support of Molecular Characters for Morphological Features
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Averyanov, L.; Cribb, P.; Phan, K.L.; Nguyen, T.H. Slipper Orchids of Vietnam; Bird Life, Royal Botanic Gardens KEW; World Bank: Ho Chi Minh City, Vietnam, 2004; p. 308. [Google Scholar]
- Braem, G.; Gruss, O. Paphiopedilum subgenus Megastaminodium Braem & Gruss, a new subgenus to accommodate Paphiopedilum canhii. Orchid Dig. 2012, 76, 32–35. [Google Scholar]
- Averyanov, L.V.; Gruss, O.; Canh, C.X.; Loc, P.K.; Dang, B.; Hiep, N.T. Paphiopedilum canhii—A new species from Northern Vietnam. Orchids 2010, 79, 288–290. [Google Scholar]
- Following Vietnam Paphiopedilum (Period 1). Available online: https://www.thiennhien.net/2012/01/26/theo-dau-lan-hai-viet-nam-ky-1/ (accessed on 6 December 2019).
- IUCN. The IUCN Red List of Threatened Species; Version 2019-2; IUCN: Gland, Switzerland, 2019.
- Vu, H.-T.; Bui, M.-H.; Vu, Q.; Nguyen, T.-D.; Tran, H.; Khuat, H.-T.; Le, L.; Tran, H.-D. Identification of Vietnamese Paphiopedilum Species Using Vegetative Morphology. Asian J. Plant Sci. under processing.
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrado, G.; Rao, R. Special Issue: Plant Genetics and Biotechnology in Biodiversity. Diversity 2018, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Mondini, L.; Noorani, A.; Pagnotta, M. Assessing Plant Genetic Diversity by Molecular Tools. Diversity 2009, 1, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Vandergast, A.; Inman, R.; Barr, K.; Nussear, K.; Esque, T.; Hathaway, S.; Wood, D.; Medica, P.; Breinholt, J.; Malone, C.; et al. Evolutionary Hotspots in the Mojave Desert. Diversity 2013, 5, 293–319. [Google Scholar] [CrossRef] [Green Version]
- Lahaye, R.; van der Bank, M.; Bogarin, D.; Warner, J.; Pupulin, F.; Gigot, G.; Maurin, O.; Duthoit, S.; Barraclough, T.G.; Savolainen, V. DNA barcoding the floras of biodiversity hotspots. Proc. Natl. Acad. Sci. USA 2008, 105, 2923–2928. [Google Scholar] [CrossRef] [Green Version]
- Lima, R.A.F.d.; Oliveira, A.A.d.; Colletta, G.D.; Flores, T.B.; Coelho, R.L.G.; Dias, P.; Frey, G.P.; Iribar, A.; Rodrigues, R.R.; Souza, V.C.; et al. Can plant DNA barcoding be implemented in species-rich tropical regions? A perspective from São Paulo State, Brazil. Genet. Mol. Biol. 2018, 41, 661–670. [Google Scholar] [CrossRef]
- Ghorbani, A.; Saeedi, Y.; de Boer, H.J. Unidentifiable by morphology: DNA barcoding of plant material in local markets in Iran. PLoS ONE 2017, 12, e0175722. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Liu, Y.; Wang, X.; Wei, X.; Han, J. DNA Mini-Barcoding: A Derived Barcoding Method for Herbal Molecular Identification. Front. Plant Sci. 2019, 10, 987. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.-W.; Liao, B.-S.; Luo, L.; Ren, Y.-Y. Species identification of poisonous medicinal plant using DNA barcoding. Chin. J. Natl. Med. 2019, 17, 585–590. [Google Scholar] [CrossRef]
- Asahina, H.; Shinozaki, J.; Masuda, K.; Morimitsu, Y.; Satake, M. Identification of medicinal Dendrobium species by phylogenetic analyses using matK and rbcL sequences. J. Natl. Med. 2010, 64, 133–138. [Google Scholar] [CrossRef]
- Xu, S.; Li, D.; Li, J.; Xiang, X.; Jin, W.; Huang, W.; Jin, X.; Huang, L. Evaluation of the DNA Barcodes in Dendrobium (Orchidaceae) from Mainland Asia. PLoS ONE 2015, 10, e0115168. [Google Scholar] [CrossRef]
- Yukawa, T.; Kinoshita1, A.; Tanaka, N. Molecular Identification Resolves Taxonomic Confusion in Grammatophyllum speciosum Complex (Orchidaceae). Bull. Natl. Mus. Nat. Sci. Ser. B 2013, 39, 137–145. [Google Scholar]
- Singh, H.K.; Parveen, I.; Raghuvanshi, S.; Babbar, S.B. The loci recommended as universal barcodes for plants on the basis of floristic studies may not work with congeneric species as exemplified by DNA barcoding of Dendrobium species. BMC Res. Notes 2012, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-T.; Gupta, S.K.; Wang, A.Z.-M.; Lo, S.-F.; Kuo, C.; Ko, Y.-J.; Chen, C.; Hsieh, C.-C.; Tsay, H.-S. Internal Transcribed Spacer Sequence Based Identification and Phylogenic Relationship of Herba Dendrobii. J. Food Drug Anal. 2012, 20, 143–151. [Google Scholar]
- Xiang, X.G.; Hu, H.; Wang, W.; Jin, X.H. DNA barcoding of the recently evolved genus Holcoglossum (Orchidaceae: Aeridinae): A test of DNA barcode candidates. Mol. Ecol. Resour. 2011, 11, 1012–1021. [Google Scholar] [CrossRef]
- Veldman, S.; Kim, S.J.; van Andel, T.R.; Bello Font, M.; Bone, R.E.; Bytebier, B.; Chuba, D.; Gravendeel, B.; Martos, F.; Mpatwa, G.; et al. Trade in Zambian Edible Orchids-DNA Barcoding Reveals the Use of Unexpected Orchid Taxa for Chikanda. Genes 2018, 9, 595. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.D.; Khuat, H.T.; La, T.N.; Nguyen, T.T.T.; Pham, B.H.; Nguyen, T.K.; Tran, H.D.; Do, M.T.; Tran, D.K. Identification of Vietnamese Native Dendrobium Species Based on Ribosomal DNA Internal Transcribed Spacer Sequence. Adv. Stud. Biol. 2018, 10, 1–12. [Google Scholar]
- Tsai, C.C.; Huang, S.C.; Huang, P.L.; Chou, C.H. Phylogeny of the genus Phalaenopsis (Orchidaceae) with emphasis on the subgenus Phalaenopsis based on the sequences of the internal transcribed spacers 1 and 2 of rDNA. J. Hortic. Sci. Biotechnol. 2003, 78, 879–887. [Google Scholar] [CrossRef]
- Yu, N.; Wei, Y.; Zhang, X.; Zhu, N.; Wang, Y.; Zhu, Y.; Zhang, H.; Li, F.; Yang, L.; Sun, J.; et al. Barcode ITS2: A useful tool for identifying Trachelospermum jasminoides and a good monitor for medicine market. Sci. Rep. 2017, 7, 5037. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Song, J.Y.; Ma, X.Y.; Liu, C.; Li, Y.; Xu, H.X.; Han, J.P.; Duan, L.S.; Chen, S.L. Identification of Dendrobium species by a candidate DNA barcode sequence: The chloroplast psbA-trnH intergenic region. Plant. Med. 2009, 75, 667–669. [Google Scholar] [CrossRef]
- Rajaram, M.C.; Yong, C.; Azlan, G.J.; Go, R. DNA Barcoding of Endangered Paphiopedilum species (Orchidaceae) of Peninsular Malaysia. Phytotaxa 2019, 387, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Chase, M.W.; Cowan, R.S.; Hollingsworth, P.M.; van den Berg, C.; Madriñán, S.; Petersen, G.; Seberg, O.; Jørgsensen, T.; Cameron, K.M.; Carine, M.; et al. A proposal for a standardised protocol to barcode all land plants. Taxon 2007, 56, 295–299. [Google Scholar] [CrossRef]
- Gigot, G.; Van Alphen-Stahl, J.; Bogarin, D.; Warner, J.; Chase, M.W.; Savolainen, V. Finding a suitable DNA barcode for Mesoamerican orchids. Lankesteriana Int. J. Orchidol. 2007, 7, 200–203. [Google Scholar] [CrossRef] [Green Version]
- Siripiyasing, P. DNA barcoding of the Cymbidium species (Orchidaceae) in Thailand. Afr. J. Agric. Res. 2012, 7, 393–404. [Google Scholar]
- Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; Erickson, D.L.; Fazekas, A.J.; et al. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar]
- Chattopadhyay, P.; Banerjee, G.; Banerjee, N. Distinguishing Orchid Species by DNA Barcoding: Increasing the Resolution of Population Studies in Plant Biology. Omics 2017, 21, 711–720. [Google Scholar] [CrossRef]
- Feng, S.; Jiang, Y.; Wang, S.; Jiang, M.; Chen, Z.; Ying, Q.; Wang, H. Molecular Identification of Dendrobium Species (Orchidaceae) Based on the DNA Barcode ITS2 Region and Its Application for Phylogenetic Study. Int. J. Mol. Sci. 2015, 16, 21975–21988. [Google Scholar] [CrossRef]
- Xu, Q.; Zhang, G.-Q.; Liu, Z.-J.; Luo, Y.-B. Two new species of Dendrobium (Orchidaceae: Epidendroideae) from China: Evidence from morphology and DNA. Phytotaxa 2014, 174, 15. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.-Y.; Lin, B.-Y.; Chang, C.-D.; Liao, S.-C.; Liu, Y.-C.; Wu, W.; Chang, C.-C. Evaluation of chloroplast DNA markers for distinguishing Phalaenopsis species. Sci. Hortic. 2015, 192, 302–310. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, H.T.; Son, S.-W.; Kim, J.-H. Molecular identification of endangered Korean lady’s slipper orchids (Cypripedium, Orchidaceae) and related taxa. Botany 2015, 93, 603–610. [Google Scholar] [CrossRef]
- Khew, G.S.-W.; Chia, T.F. Parentage determination of Vanda Miss Joaquim (Orchidaceae) through two chloroplast genes rbcL and matK. AoB Plants 2011, 2011, plr018. [Google Scholar] [CrossRef]
- Senthilkumar, S.; Xavier, T.F.; Gomathi, G. DNA Barcoding of Vanda Species from the Regions of Shevaroy and Kolli Hills using rbcL gene. Biotechnol. Res. 2017, 3, 126–128. [Google Scholar]
- Ginibun, F.C.; Saad, M.R.M.; Hong, T.L.; Othman, R.Y.; Khalid, N.; Bhassu, S. Chloroplast DNA Barcoding of Spathoglottis Species for Genetic Conservation. Acta Hortuc. 2010, 878, 453–460. [Google Scholar] [CrossRef]
- Parveen, I.; Singh, H.K.; Raghuvanshi, S.; Pradhan, U.C.; Babbar, S.B. DNA barcoding of endangered Indian Paphiopedilum species. Mol. Ecol. Resour. 2012, 12, 82–90. [Google Scholar] [CrossRef]
- Parveen, I.; Singh, H.K.; Malik, S.; Raghuvanshi, S.; Babbar, S.B. Evaluating five different loci (rbcL, rpoB, rpoC1, matK, and ITS) for DNA barcoding of Indian orchids. Genome 2017, 60, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.-Y.; Luo, Y.-B.; Liu, Z.-J.; Wang, X.-Q. Evolution and Biogeography of the Slipper Orchids: Eocene Vicariance of the Conduplicate Genera in the Old and New World Tropics. PLoS ONE 2012, 7, e38788. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.-Y.; Luo, Y.-B.; Liu, Z.-J.; Wang, X.-Q. Reticulate evolution and sea-level fluctuations together drove species diversification of slipper orchids (Paphiopedilum) in South-East Asia. Mol. Ecol. 2015, 24, 2838–2855. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.Y.; Huang, L.Q.; Liu, Z.J.; Wang, X.Q. Promise and Challenge of DNA Barcoding in Venus Slipper (Paphiopedilum). PLoS ONE 2016, 11, e0146880. [Google Scholar] [CrossRef] [PubMed]
- Trung, K.H.; Khanh, T.D.; Ham, L.H.; Duong, T.D.; Khoa, T. Molecular Phylogeny of the Endangered Vietnamese Paphiopedilum Species Based on the Internal Transcribed Spacer of the Nuclear Ribosomal DNA. Adv. Stud. Biol. 2013, 5, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Vu, H.-T.; Huynh, P.; Tran, H.-D.; Le, L. In Silico Study on Molecular Sequences for Identification of Paphiopedilum Species. Evolut. Bioinform. 2018, 14, 117693431877454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, T.H.T.; Le, L.; Nguyen, T.K.; Tran, D.D.; Tran, H.D. Review on molecular markers for identification of Orchids. Vietnam Sci. Technol. 2017, 59, 62–75. [Google Scholar] [CrossRef]
- Cribb, P. The Genus Paphiopedilum, 2nd ed.; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 1998. [Google Scholar]
- Dang, V.K.; Nguyen, T.N.; Vu, H.-T. Screening, designing, pilot evaluation and application of some potential primers for molecular discrimination of Paphiopedilum species in Vietnam. Vietnam. J. Agric. Rural Devel. 2017, 113–118. Available online: http://www.tapchikhoahocnongnghiep.vn/uploads/news/2017_12/16a.pdf (accessed on 1 December 2019).
- Geospiza, I. FinchTV version 1.4.0: DNA sequence chromatogram trace viewer. 2004. Available online: http://www.geospiza.com (accessed on 11 January 2019).
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2009, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony and Other Methods; Version 4.0.; Sinauer Associates, Oxford University: Oxford, UK, 2003. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree, version 1.4.3 [Computer program]. 2009. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 11 January 2019).
- Dong, W.; Liu, H.; Xu, C.; Zuo, Y.; Chen, Z.; Zhou, S. A chloroplast genomic strategy for designing taxon specific DNA mini-barcodes: A case study on ginsengs. BMC Genet. 2014, 15, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bafeel, S.; Arif, I.; Bakir, M.; Khan, H.; Farhan, A.; Al-Homaidan, A.; Ahamed, A.; Thomas, J. Comparative evaluation of PCR success with universal primers of maturase K (matK) and ribulose-1, 5-bisphosphate carboxylase oxygenase large subunit (rbcL) for barcoding of some arid plants. Plant Omics J. 2011, 4, 195–198. [Google Scholar]
- Kress, W.J.; Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Perez, R.; Sanjur, O.; Bermingham, E. Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama. Proc. Natl. Acad. Sci. USA 2009, 106, 18621–18626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.I.; Chang, F.C.; Chung, M.C. Chromosome pairing affinities in interspecific hybrids reflect phylogenetic distances among lady’s slipper orchids (Paphiopedilum). Ann. Bot. 2011, 108, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, X.-G.; Li, D.-Z.; Jin, W.-T.; Zhou, H.; Jianwu, L.; Jin, X.-H. Phylogenetic placement of the enigmatic orchid genera Thaia and Tangtsinia: Evidence from molecular and morphological characters. Taxon 2012, 61, 45–54. [Google Scholar] [CrossRef]
- Kolaczkowski, B.; Thornton, J.W. Performance of maximum parsimony and likelihood phylogenetics when evolution is heterogeneous. Nature 2004, 431, 980. [Google Scholar] [CrossRef]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Mar, J.C.; Harlow, T.J.; Ragan, M.A. Bayesian and maximum likelihood phylogenetic analyses of protein sequence data under relative branch-length differences and model violation. BMC Evol. Biol. 2005, 5, 8. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Annealing Temperature (°C) | Primer Name | Primer Sequence | Expected Product Length (bp) | Reference |
|---|---|---|---|---|---|
| ITS | 58 | IT1–F | 5′-AGTCGTAACAAGGTTTCC-3′ | 900 | [24] |
| IT2–R | 5′-GTAAGTTTCTTCTCCTCC-3′ | ||||
| matK | 55 | F56–mo | 5′-CCTATCCATCTGGAAATCTTAG-3′ | 1200 | [51] |
| R1326–mo | 5′-GTTCTAGCACAAGAAAGTCG-3′ | ||||
| trnL | 62 | trnL–F | 5′-GGTAGAGCTACGACTTGATT-3′ | 600 | |
| trnL–R | 5′-CGGTATTGACATGTAAAATGGGACT-3′ | ||||
| rpoB | 53 | 2F | 5′-ATGCAACGTCAAGCAGTTCC-3′ | 600 | [32] |
| 4R | 5′-GATCCCAGCATCACAATTCC-3′ | ||||
| rpoC1 | 53 | 1F | 5′-GTGGATACACTTCTTGATAATGG-3′ | 600 | |
| 3R | 5′-TGAGAAAACATAAGTAAAGGGC-3′ | ||||
| trnH-psbA | 53 | psbA3′f | 5′-CGCGCATGGTGGTTCACAATCC-3′ | 900 | |
| trnHf | 5′-GTTATGCATGAACGTAATGCTC-3′ |
| No. | Species | Specimen Voucher | ACO | LEAFY | ITS | matK | trnL | rpoB | rpoC1 | trnH-psbA | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ACO F1/R1 | ACO F2/R2 | LFY F1/R1 | LFY F2/R2 | |||||||||
| 1 | Paphiopedilum delenatii | DEL-2 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/− |
| DEL-46 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||
| DEL-47 | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||||||
| DEL-187 | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||||||
| DEL-188 | +/+ | +/+ | +/+ | +/+ | +/+ | −/ | ||||||
| 2 | Paphiopedilum x dalatense | DAL-138 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ |
| DAL-139 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ | ||
| DAL-143 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||
| 3 | Paphiopedilum gratrixianum | GRA-145 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ |
| GRA-146 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||
| GRA-180 | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||||||
| GRA-182 | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||||||
| 4 | Paphiopedilum hangianum | HAN-16 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/− |
| HAN-17 | +/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||
| HAN-18 | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | |||||
| 5 | Paphiopedilum helenea | HEL-69 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | −/ |
| HEL-70 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||
| HEL-71 | +/+ | +/+ | +/+ | +/+ | +/+ | +/− | ||||||
| 6 | Paphiopedilum x herrmannii | HER-177 | −/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | −/ |
| 7 | Paphiopedilum tranlienianum | TRA-63 | +/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ |
| TRA-64 | +/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ | ||
| TRA-66 | +/ | +/+ | +/+ | +/+ | +/+ | +/+ | +/+ | |||||
| 8 | Paphiopedilum vietnamense | VIE-130 | +/ | −/ | −/ | −/ | +/+ | +/+ | +/+ | +/+ | +/+ | −/ |
| Number of successful amplification | 5/16 | 0/14 | 0/14 | 0/14 | 23/23 | 23/23 | 23/23 | 23/23 | 23/23 | 19/23 | ||
| Rate of successful amplification | 31.25% | 0% | 0% | 0% | 100% | 100% | 100% | 100% | 100% | 82.61% | ||
| Number of successful sequencing | 23/23 | 23/23 | 23/23 | 23/23 | 23/23 | 6/19 | ||||||
| Rate of successful sequencing | 100% | 100% | 100% | 100% | 100% | 31.58% | ||||||
| Sequence Locus | Alignment Length (bp) (L) | Parsimony Site (P) | Singleton Site (S) | Variable Site (V = P + S) | Variable Rate (%) (V/L) | Indel Fragment |
|---|---|---|---|---|---|---|
| ITS | 725 | 166 | 71 | 237 | 32.7 | 20 |
| matK | 1132 | 74 | 39 | 113 | 10.0 | 1 |
| trnL | 466 | 25 | 9 | 34 | 7.3 | 3 |
| rpoB | 483 | 7 | 5 | 12 | 2.5 | 0 |
| rpoC1 | 460 | 5 | 3 | 8 | 1.7 | 0 |
| ITS | matK | trnL | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | SPECIES | NJ | ML1 | ML2 | MP | BA | NJ | ML1 | ML2 | MP | BA | NJ | ML1 | ML2 | MP | BA |
| 1 | Paphiopedilum appletonianum | + | + | + | − | + | − | − | − | − | − | − | − | − | − | − |
| 2 | Paphiopedilum armeniacum | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 3 | Paphiopedilum callosum | + | + | + | + | + | − | − | − | − | − | − | − | − | − | − |
| 4 | Paphiopedilum coccineum | + | + | + | + | + | − | − | − | − | − | − | − | − | − | − |
| 5 | Paphiopedilum concolor | + | + | + | + | + | + | + | + | + | + | − | − | − | − | − |
| 6 | Paphiopedilum dalatense | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| 7 | Paphiopedilum delenatii | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 8 | Paphiopedilum dianthum | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 9 | Paphiopedilum emersonii | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + |
| 10 | Paphiopedilum gratrixianum | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| 11 | Paphiopedilum hangianum | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + |
| 12 | Paphiopedilum helenea | − | − | − | − | − | + | + | + | − | + | − | − | − | − | − |
| 13 | Paphiopedilum henryanum | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| 14 | Paphiopedilum herrmannii | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| 15 | Paphiopedilum hirsutissimum | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 16 | Paphiopedilum malipoense | + | + | + | + | + | + | + | + | + | + | − | − | − | − | − |
| 17 | Paphiopedilum micranthum | + | + | + | + | + | + | + | + | + | + | − | − | − | − | − |
| 18 | Paphiopedilum purpuratum | + | + | + | + | + | + | + | + | + | + | − | − | − | − | − |
| 19 | Paphiopedilum tranlienianum | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| 20 | Paphiopedilum vietnamense | + | + | + | + | + | + | + | + | + | + | − | − | − | − | − |
| 21 | Paphiopedilum villosum | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Number of monophyletic species | 14 | 14 | 14 | 13 | 14 | 12 | 12 | 12 | 11 | 12 | 6 | 6 | 6 | 4 | 6 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, H.-T.; Vu, Q.-L.; Nguyen, T.-D.; Tran, N.; Nguyen, T.-C.; Luu, P.-N.; Tran, D.-D.; Nguyen, T.-K.; Le, L. Genetic Diversity and Identification of Vietnamese Paphiopedilum Species Using DNA Sequences. Biology 2020, 9, 9. https://0-doi-org.brum.beds.ac.uk/10.3390/biology9010009
Vu H-T, Vu Q-L, Nguyen T-D, Tran N, Nguyen T-C, Luu P-N, Tran D-D, Nguyen T-K, Le L. Genetic Diversity and Identification of Vietnamese Paphiopedilum Species Using DNA Sequences. Biology. 2020; 9(1):9. https://0-doi-org.brum.beds.ac.uk/10.3390/biology9010009
Chicago/Turabian StyleVu, Huyen-Trang, Quoc-Luan Vu, Thanh-Diem Nguyen, Ngan Tran, Thanh-Cong Nguyen, Phuong-Nam Luu, Duy-Duong Tran, Truong-Khoa Nguyen, and Ly Le. 2020. "Genetic Diversity and Identification of Vietnamese Paphiopedilum Species Using DNA Sequences" Biology 9, no. 1: 9. https://0-doi-org.brum.beds.ac.uk/10.3390/biology9010009