
Antimicrobial, Immunomodulatory and Anti-Inflammatory Potential of Liposomal Thymoquinone: Implications in the Treatment of Bacterial Pneumonia in Immunocompromised Mice

 ,
,  , , ,
, , ,  , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Acinetobacter baumannii
2.3. Antimicrobial Susceptibility Testing
2.4. Bactericidal Activity of Free TQ or Lip-TQ on the Biofilm Formation and on the Preformed Biofilm by A. baumannii
2.5. Formulation of TQ-Loaded Lipid Nanoparticles
2.6. Mice
2.7. Induction of Leukopenia in Mice
2.8. Standardization of A. baumannii Infection in Leukopenic Mice
2.9. Infection of Leukopenic Mice with A. baumannii
2.10. Treatment of A. baumannii-Infected Mice with TQ Formulations
2.11. Determination of Bacterial Load in the Lung Tissues
2.12. Determination of Inflammation Markers
2.13. Evaluation of the Status of Oxidative Stress in the Lungs
2.14. Histological Study of the Lung Tissue
2.15. Statistical Analyses
3. Results
3.1. TQ Shows Potent Activity against A. baumannii
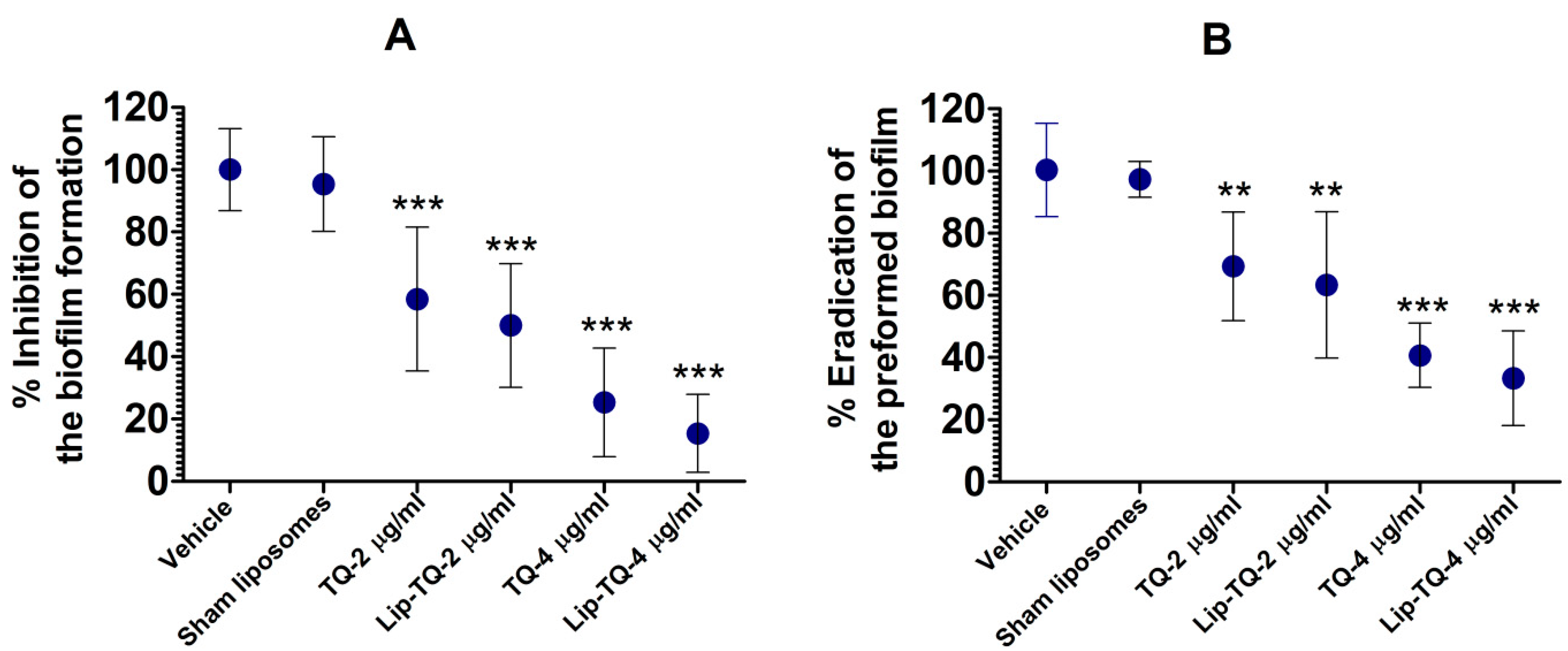
3.2. TQ Effectively Inhibited the Biofilm Formation and Eradicated the Preformed Biofilm
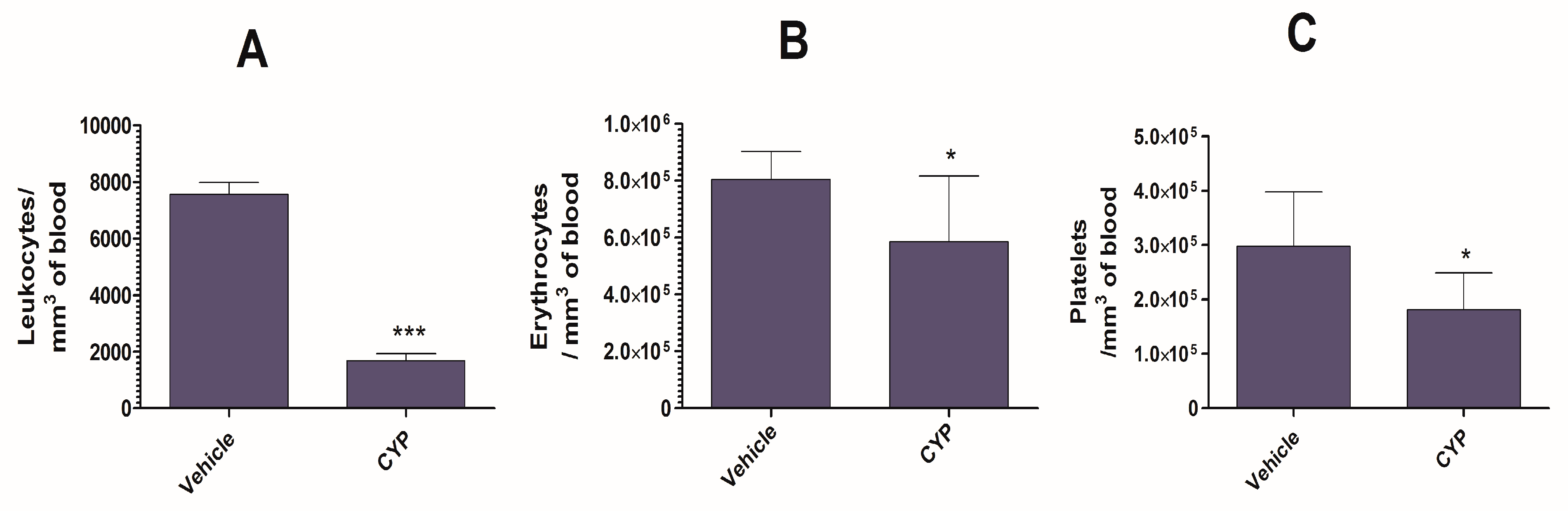
3.3. CYP Administration Induces Acute Leukopenia in Mice
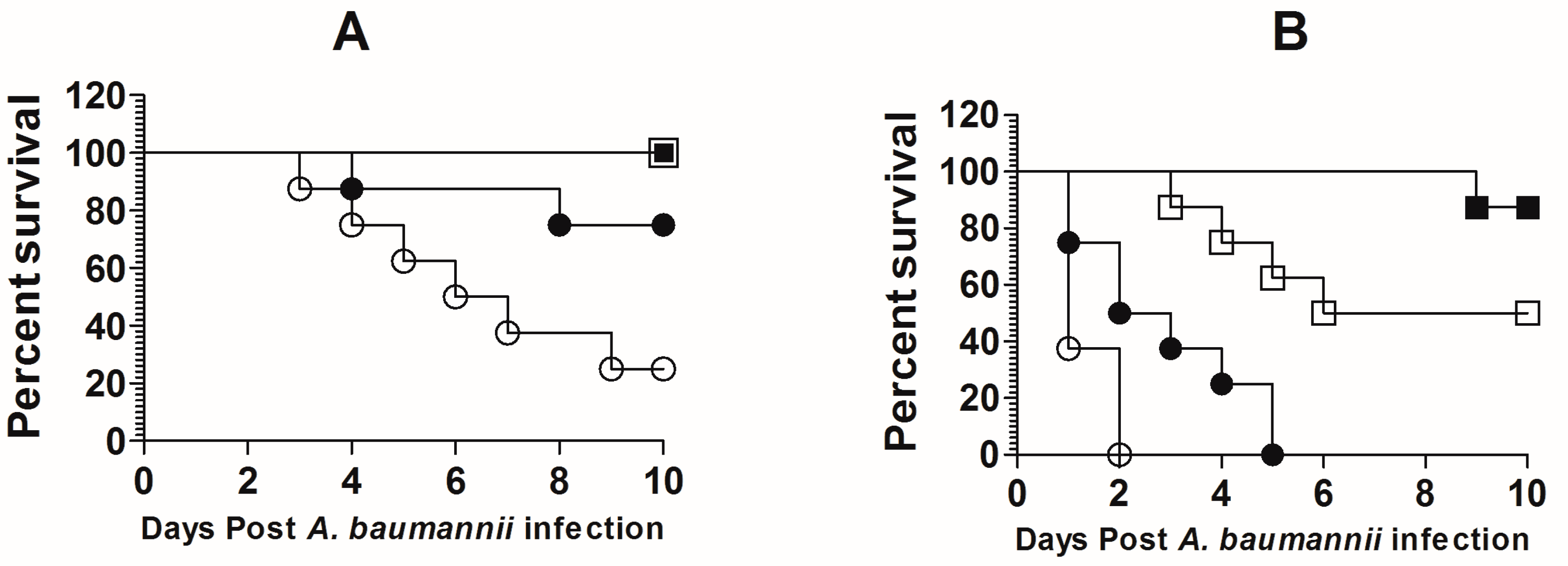
3.4. Standardization of the Dose of A. baumannii in Immunocompetent and Immunocompromised Mice
3.5. Treatment with Lip-TQ, Not with Free TQ, Was Effective against A. baumannii in Leukopenic Mice
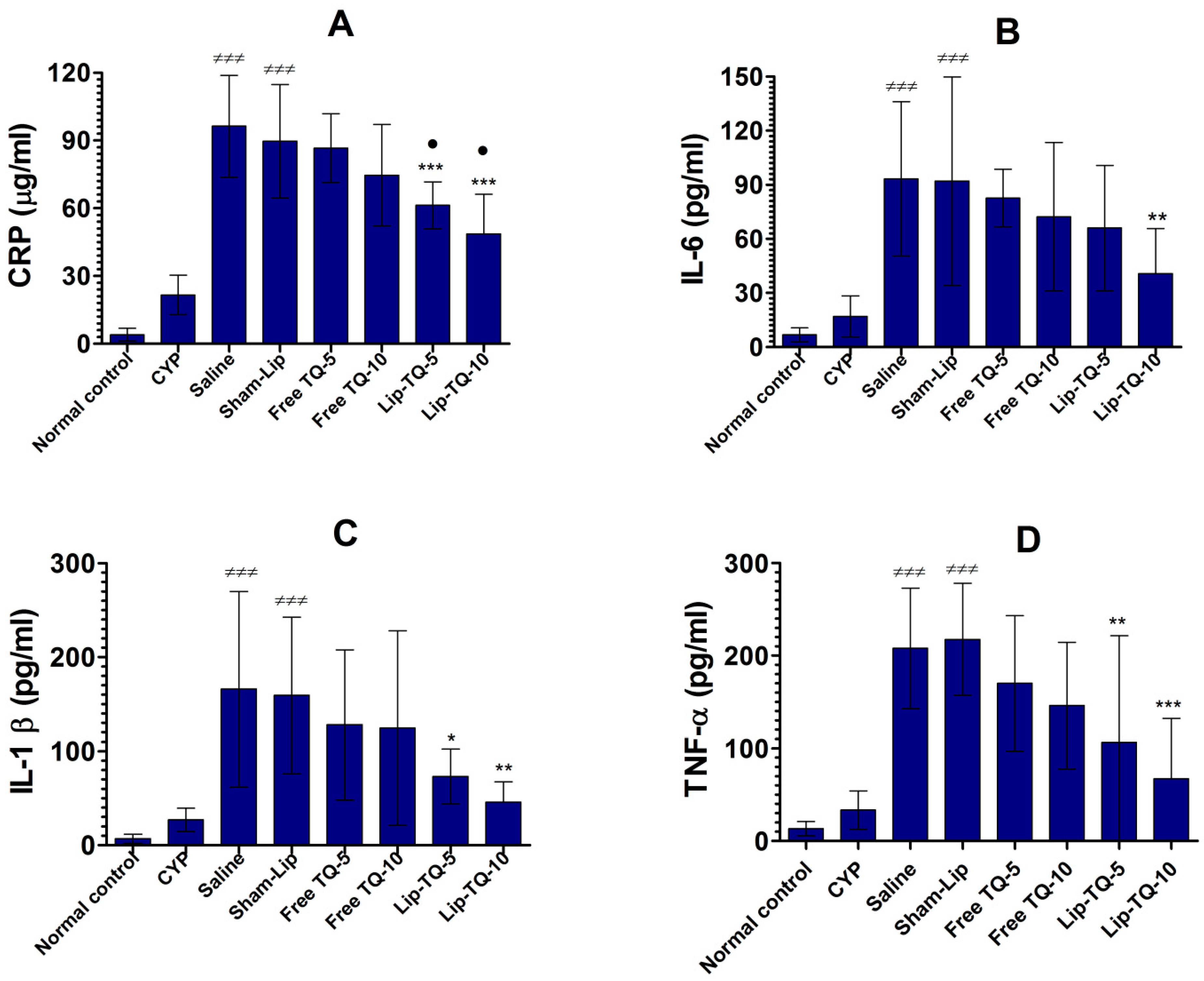
3.6. Lip-TQ Treatment Alleviated the Levels of CRP, IL-6, IL-1β, and TNF-α in the Systemic Circulation of A. baumannii-Infected Mice
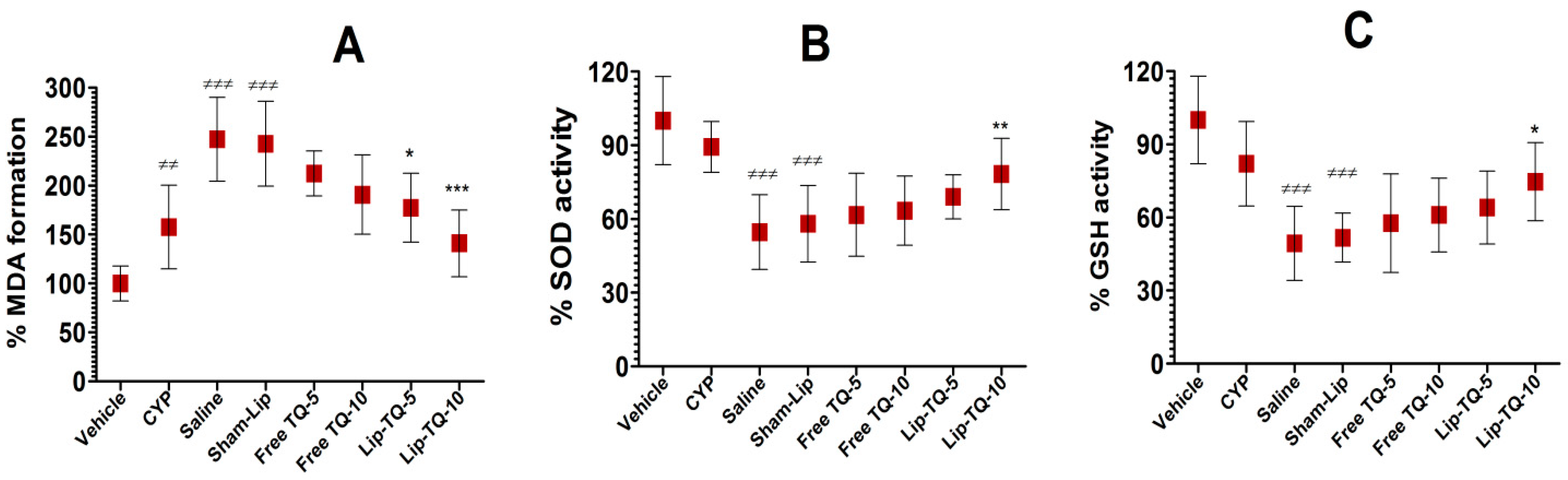
3.7. Lip-TQ Treatment Improved the Status of Oxidative Stress in A. baumannii-Infected Mice
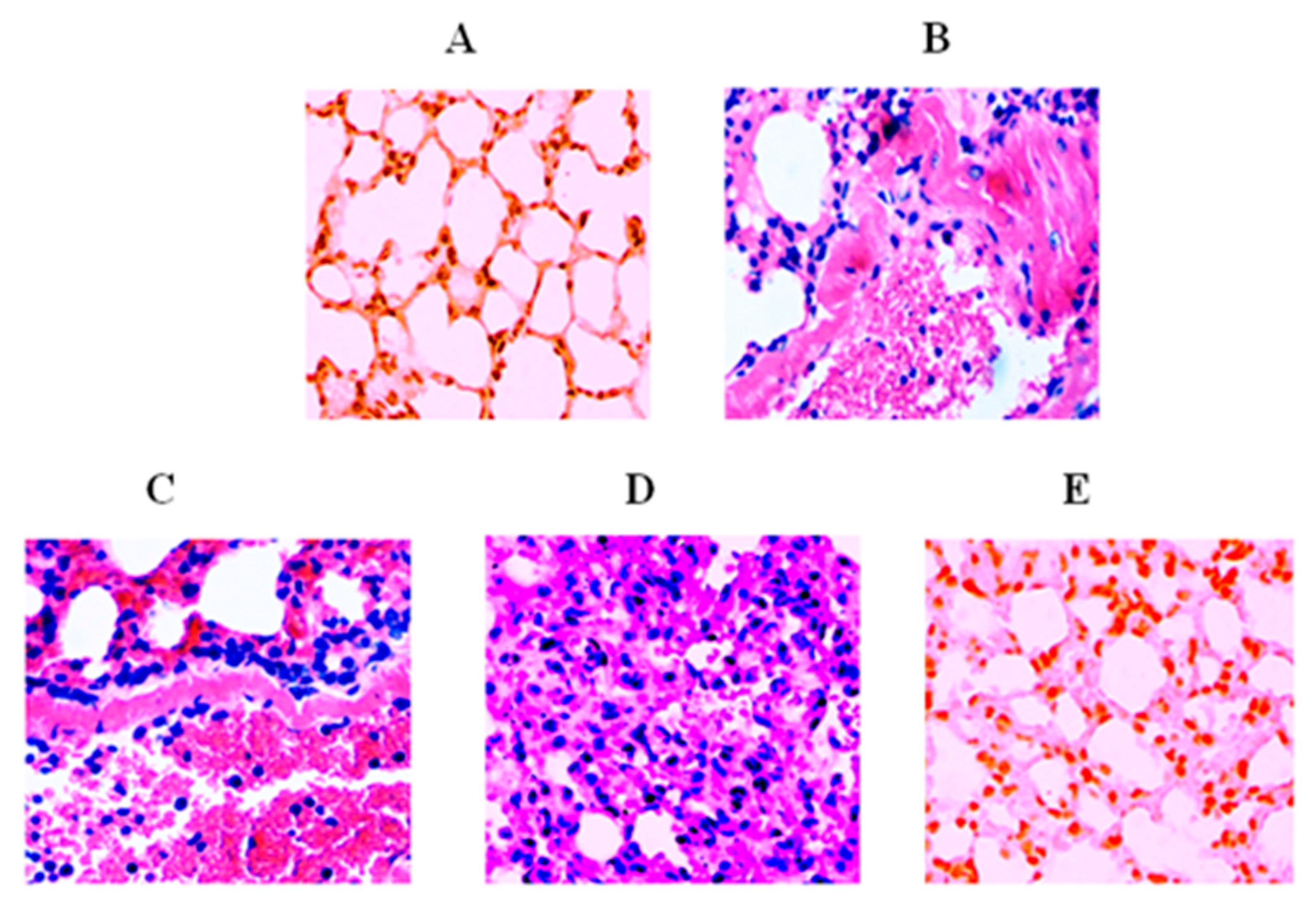
3.8. Lip-TQ Reversed Infection-Induced Pathological Changes in the Lung Tissues
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vázquez-López, R.; Solano-Gálvez, S.G.; Juárez Vignon-Whaley, J.J.; Abello Vaamonde, J.A.; Padró Alonzo, L.A.; Rivera Reséndiz, A.; Muleiro Álvarez, M.; Vega López, E.N.; Franyuti-Kelly, G.; Álvarez-Hernández, D.A.; et al. Acinetobacter baumannii Resistance: A Real Challenge for Clinicians. Antibiotics 2020, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Fukuta, Y.; Muder, R.R.; Agha, M.E.; Clarke, L.G.; Wagener, M.M.; Hensler, A.M.; Doi, Y. Risk factors for acquisition of multidrug-resistant Acinetobacter baumannii among cancer patients. Am. J. Infect. Control. 2013, 41, 1249–1252. [Google Scholar] [CrossRef] [PubMed]
- Chen, W. Host Innate Immune Responses to Acinetobacter baumannii Infection. Front. Cell Infect. Microbiol. 2020, 10, 486. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.R.; Du, W.J.; Xie, J.F.; Xu, J.Y.; Huang, Y.Z.; Qiu, H.B.; Yang, Y. Role of immunodeficiency in Acinetobacter baumannii associated pneumonia in mice. Chin. Med. J. Engl. 2020, 133, 2161–2169. [Google Scholar] [CrossRef]
- Grguric-Smith, L.M.; Lee, H.H.; Gandhi, J.A.; Brennan, M.B.; DeLeon-Rodriguez, C.M.; Coelho, C.; Han, G.; Martinez, L.R. Neutropenia exacerbates infection by Acinetobacter baumannii clinical isolates in a murine wound model. Front. Microbiol. 2015, 6, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, X.Z.; Zhang, X.; Chen, Y.; Tomofusa, T. Multiple resistance in Acinetobacter baumannii ATCC 19606 and cloning of genes responsible for the resistance. Chin. J. Antibiot. 2006, 31, 688–691. [Google Scholar]
- Cai, Y.; Chai, D.; Wang, R.; Liang, B.; Bai, N. Colistin resistance of Acinetobacter baumannii: Clinical reports, mechanisms and antimicrobial strategies. Antimicrob. Chemother. 2012, 67, 1607–1615. [Google Scholar] [CrossRef]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef]
- Colquhoun, J.M.; Rather, P.N. Insights into Mechanisms of Biofilm Formation in Acinetobacter baumannii and Implications for Uropathogenesis. Front. Cell Infect. Microbiol. 2020, 10, 253. [Google Scholar] [CrossRef]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Ullah, F.; Ovais, M.; Ahmed, J.; Devkota, H.P. Synergistic interactions of phytochemicals with antimicrobial agents: Potential strategy to counteract drug resistance. Chem. Biol. Interact. 2019, 308, 294–303. [Google Scholar] [CrossRef]
- Khan, A.; Azam, M.; Allemailem, K.S.; Alrumaihi, F.; Almatroudi, A.; Alhumaydhi, F.A.; Ahmad, H.I.; Khan, M.U.; Khan, M.A. Coadministration of Ginger Extract and Fluconazole Shows a Synergistic Effect in the Treatment of Drug-Resistant Vulvovaginal Candidiasis. Infect. Drug Resist. 2021, 14, 1585–1599. [Google Scholar] [CrossRef]
- Khan, M.A. Antimicrobial actions of thymoquinone. In Molecular and Therapeutic Actions of Thymoquinone; Younus, H., Ed.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 57–64. [Google Scholar]
- Khan, M.A.; Younus, H. Thymoquinone Shows the Diverse Therapeutic Actions by Modulating Multiple Cell Signaling Pathways: Single Drug for Multiple Targets. Curr. Pharm. Biotechnol. 2018, 19, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Odeh, F.; Ismail, S.I.; Abu-Dahab, R.; Mahmoud, I.S.; Al Bawab, A. Thymoquinone in liposomes: A study of loading efficiency and biological activity towards breast cancer. Drug Deliv. 2012, 19, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Sixth Informational Supplement M100-S26; CLSI: Wayne, PA, USA, 2016. [Google Scholar]
- Allemailem, K.S.; Alnuqaydan, A.M.; Almatroudi, A.; Alrumaihi, F.; Aljaghwani, A.; Khalilullah, H.; Younus, H.; Khan, A.; Khan, M.A. Safety and Therapeutic Efficacy of Thymoquinone-Loaded Liposomes against Drug-Sensitive and Drug-Resistant Acinetobacter baumannii. Pharmaceutics 2021, 13, 677. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Khan, A.; Azam, M.; Allemailem, K.S.; Alrumaihi, F.; Almatroudi, A.; Alhumaydhi, F.; Azam, F.; Khan, S.H.; Zofair, S.F.F.; et al. Liposomal Ellagic Acid Alleviates Cyclophosphamide-Induced Toxicity and Eliminates the Systemic Cryptococcus neoformans Infection in Leukopenic Mice. Pharmaceutics 2021, 13, 882. [Google Scholar] [CrossRef]
- Alrumaihi, F.; Almatroudi, A.; Allemailem, K.S.; Rahmani, A.H.; Khan, A.; Khan, M.A. Therapeutic Effect of Bilsaan, Sambucus nigra Stem Exudate, on the OVA-Induced Allergic Asthma in Mice. Oxid. Med. Cell Longev. 2020, 2020, 3620192. [Google Scholar] [CrossRef]
- Breslow, J.M.; Meissler, J.J., Jr.; Hartzell, R.R.; Spence, P.B.; Truant, A.; Gaughan, J.; Eisenstein, T.K. Innate immune responses to systemic Acinetobacter baumannii infection in mice: Neutrophils, but not interleukin-17, mediate host resistance. Infect. Immun. 2011, 79, 3317–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Patiño, M.G.; García-Contreras, R.; Licona-Limón, P. The Immune Response against Acinetobacter baumannii, an Emerging Pathogen in Nosocomial Infections. Front. Immunol. 2017, 8, 441. [Google Scholar] [CrossRef] [Green Version]
- Miao, X.; Liu, H.; Zheng, Y.; Guo, D.; Shi, C.; Xu, Y.; Xia, X. Inhibitory Effect of Thymoquinone on Listeria monocytogenes ATCC 19115 Biofilm Formation and Virulence Attributes Critical for Human Infection. Front. Cell. Infect. Microbiol. 2019, 9, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randhawa, M.A.; Gondal, M.A.; Al-Zahrani, A.H.; Rashid, S.G.; Ali, A. Synthesis, morphology and antifungal activity of nano-particulated amphotericin-B, ketoconazole, and thymoquinone against Candida albicans yeasts and Candida biofilm. J. Environ. Sci. Health Part A Tox. Hazard. Subst. Environ. Eng. 2015, 50, 119–124. [Google Scholar] [CrossRef]
- Dera, A.A.; Ahmad, I.; Rajagopalan, P.; Shahrani, M.A.; Saif, A.; Alshahrani, M.Y.; Alraey, Y.; Alamri, A.M.; Alasmari, S.; Makkawi, M.; et al. Synergistic efficacies of thymoquinone and standard antibiotics against multi-drug resistant isolates. Saudi Med. J. 2021, 42, 196–204. [Google Scholar] [CrossRef]
- Fan, Q.; Yuan, Y.; Jia, H.; Zeng, X.; Wang, Z.; Hu, Z.; Gao, Z.; Yue, T. Antimicrobial and anti-biofilm activity of thymoquinone against Shigella flexneri. Appl. Microbiol. Biotechnol. 2021, 105, 4709–4718. [Google Scholar] [CrossRef]
- Almshawit, H.; Macreadie, I. Fungicidal effect of thymoquinone involves generation of oxidative stress in Candida glabrata. Microbiol. Res. 2017, 195, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Harris, G.; KuoLee, R.; Xu, H.H.; Chen, W. Acute intraperitoneal infection with a hypervirulent Acinetobacter baumannii isolate in mice. Sci. Rep. 2019, 9, 6538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, M.J.; Jo, S.G.; Kim, D.J.; Park, J.H. NLRP3 inflammasome mediates interleukin-1β production in immune cells in response to Acinetobacter baumannii and contributes to pulmonary inflammation in mice. Immunology 2017, 150, 495–505. [Google Scholar] [CrossRef]
- Al-Gabri, N.A.; Qaid, M.M.; El-Shaer, N.H.; Ali, M.H.; Abudabos, A.M. Thymoquinone ameliorates pulmonary vascular damage induced by Escherichia coli-derived lipopolysaccharide via cytokine downregulation in rats. Environ. Sci. Pollut. Res. Int. 2019, 26, 18465–18469. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, T. IL-6: From its discovery to clinical applications. Int. Immunol. 2010, 22, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Agustí, A.; Edwards, L.D.; Rennard, S.I.; MacNee, W.; Tal-Singer, R.; Miller, B.E.; Vestbo, J.; Lomas, D.A.; Calverley, P.M.; Wouters, E.; et al. Evaluation of COPD Longitudinally to Identify Predictive Surrogate Endpoints (ECLIPSE) Investigators. Persistent systemic inflammation is associated with poor clinical outcomes in COPD: A novel phenotype. PLoS ONE 2012, 7, e37483. [Google Scholar] [CrossRef]
- Shaterzadeh-Yazdi, H.; Noorbakhsh, M.F.; Hayati, F.; Samarghandian, S.; Farkhondeh, T. Immunomodulatory and Anti-inflammatory Effects of Thymoquinone. Cardiovasc. Hematol. Disord. Drug Targets 2018, 18, 52–60. [Google Scholar] [CrossRef]
- Qiu, H.; KuoLee, R.; Harris, G.; Chen, W. High susceptibility to respiratory Acinetobacter baumannii infection in A/J mice is associated with a delay in early pulmonary recruitment of neutrophils. Microbes Infect. 2009, 11, 946–955. [Google Scholar] [CrossRef]
- Laskar, A.A.; Khan, M.A.; Rahmani, A.H.; Fatima, S.; Younus, H. Thymoquinone, an active constituent of Nigella sativa seeds, binds with bilirubin and protects mice from hyperbilirubinemia and cyclophosphamide-induced hepatotoxicity. Biochimie 2016, 127, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Smani, Y.; Docobo-Pérez, F.; McConnell, M.J.; Pachón, J. Acinetobacter baumannii-induced lung cell death: Role of inflammation, oxidative stress and cytosolic calcium. Microb. Pathog. 2011, 50, 224–232. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allemailem, K.S.; Almatroudi, A.; Alrumaihi, F.; Aljaghwani, A.; Alnuqaydan, A.M.; Khalilullah, H.; Younus, H.; El-Kady, A.M.; Aldakheel, F.M.; Khan, A.A.; et al. Antimicrobial, Immunomodulatory and Anti-Inflammatory Potential of Liposomal Thymoquinone: Implications in the Treatment of Bacterial Pneumonia in Immunocompromised Mice. Biomedicines 2021, 9, 1673. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111673
Allemailem KS, Almatroudi A, Alrumaihi F, Aljaghwani A, Alnuqaydan AM, Khalilullah H, Younus H, El-Kady AM, Aldakheel FM, Khan AA, et al. Antimicrobial, Immunomodulatory and Anti-Inflammatory Potential of Liposomal Thymoquinone: Implications in the Treatment of Bacterial Pneumonia in Immunocompromised Mice. Biomedicines. 2021; 9(11):1673. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111673
Chicago/Turabian StyleAllemailem, Khaled S., Ahmad Almatroudi, Faris Alrumaihi, Aseel Aljaghwani, Abdullah M. Alnuqaydan, Habibullah Khalilullah, Hina Younus, Asmaa M. El-Kady, Fahad M. Aldakheel, Amjad Ali Khan, and et al. 2021. "Antimicrobial, Immunomodulatory and Anti-Inflammatory Potential of Liposomal Thymoquinone: Implications in the Treatment of Bacterial Pneumonia in Immunocompromised Mice" Biomedicines 9, no. 11: 1673. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111673