Working Memory, Cognitive Load and Cardiorespiratory Fitness: Testing the CRUNCH Model with Near-Infrared Spectroscopy

 , ,
, ,
Abstract
:1. Introduction
2. Method
2.1. Participants
2.2. Cognitive Assessment
2.3. CRF Assessment
2.4. Instrumentation
fNIRS Data Analysis
2.5. Procedure
2.6. Statistical Analysis
3. Results
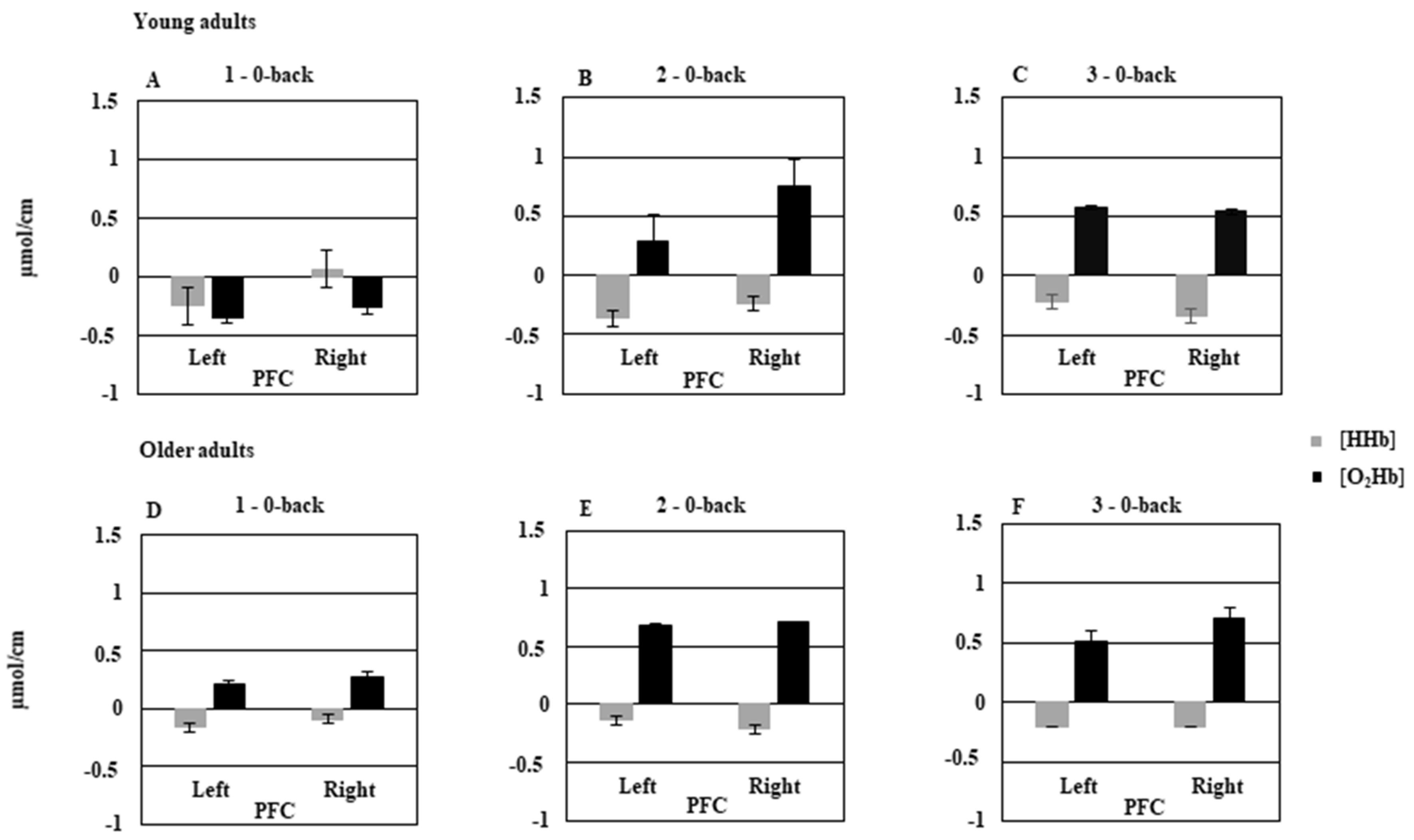
3.1. Young vs. Older Adults
3.1.1. Behavioral Data
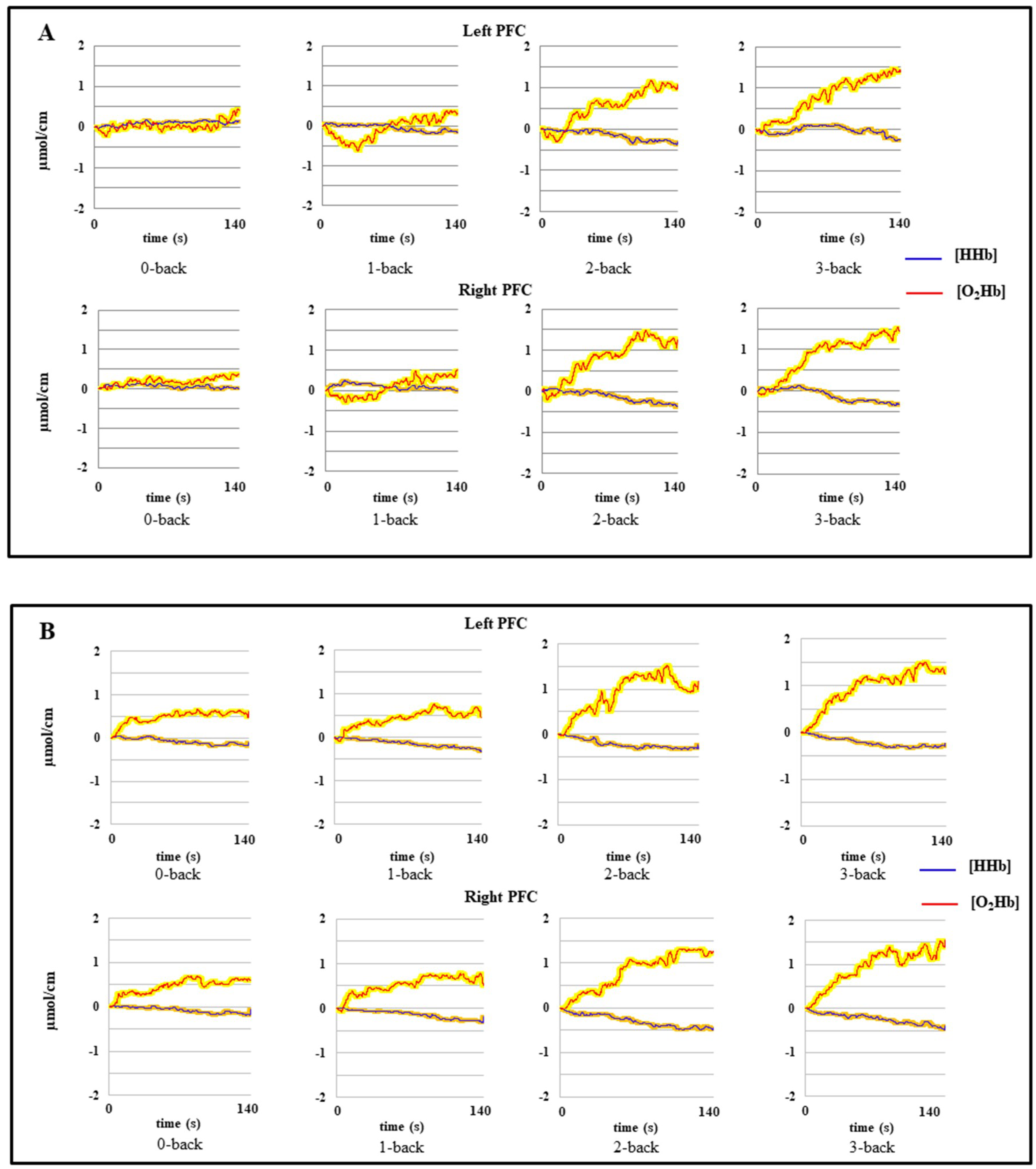
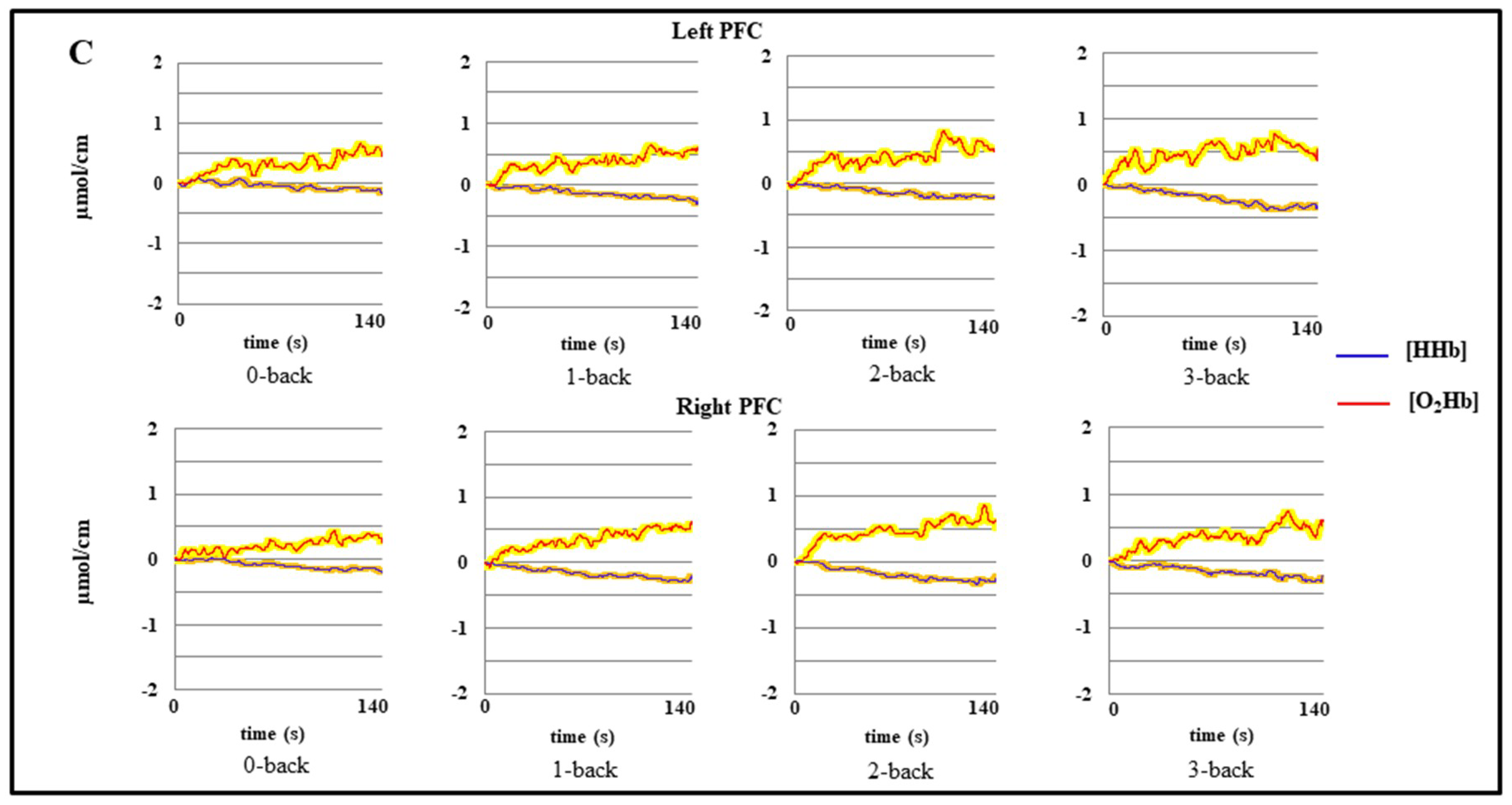
3.1.2. fNIRS Data
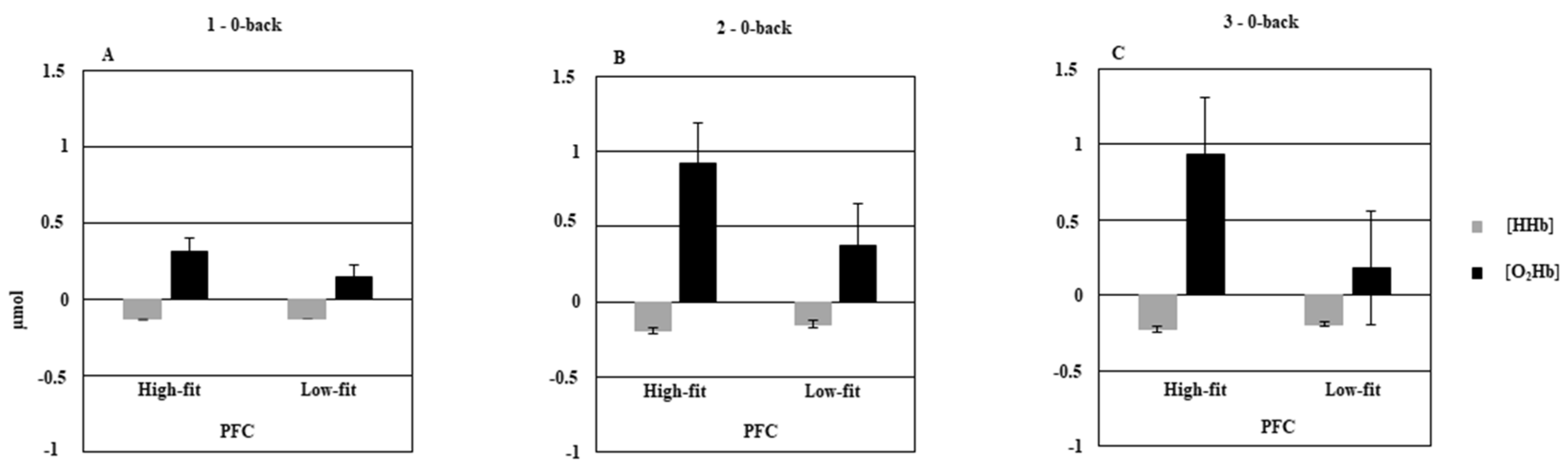
3.2. Low-Fit vs. High-Fit Older Adults
3.2.1. Behavioral Data
3.2.2. fNIRS Data
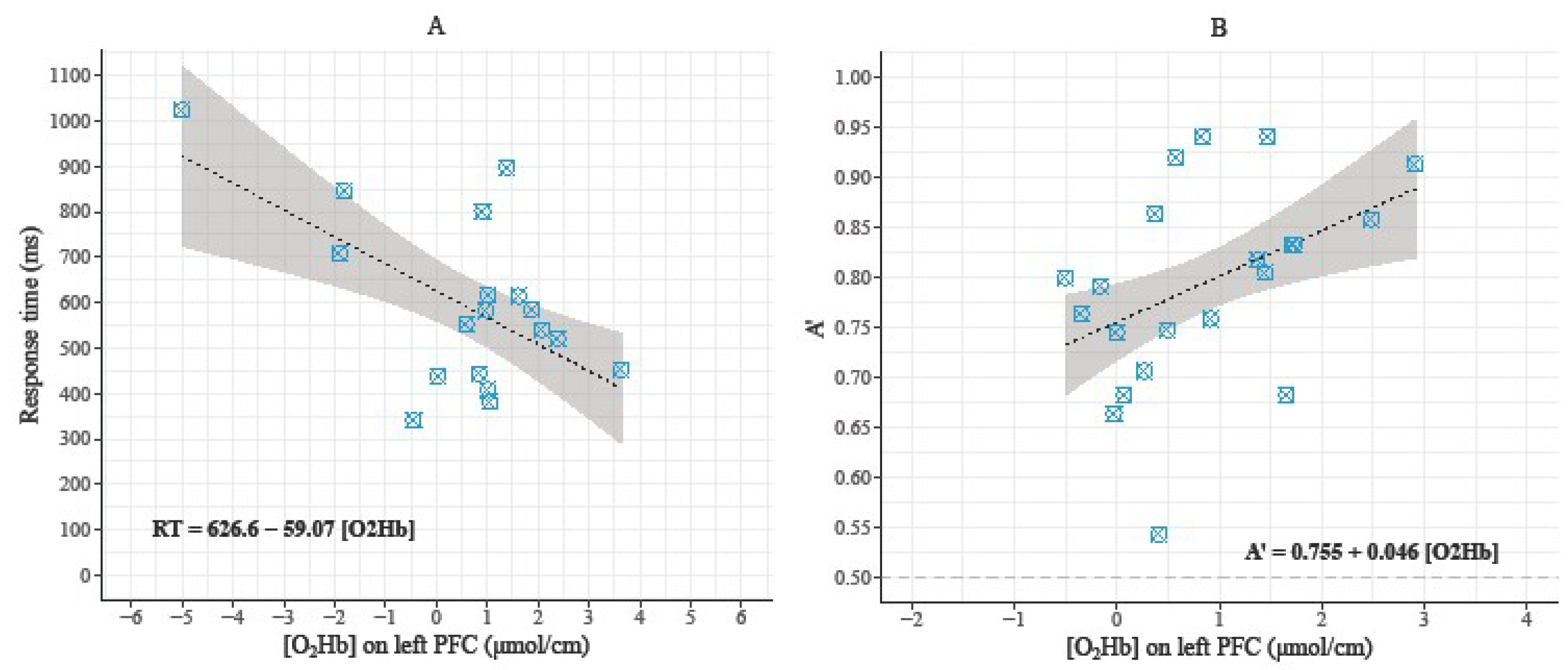
3.3. Relationships between Behavioral Data and Hemodynamic Parameters during the 3-Back Condition
4. Discussion
4.1. Effect of Age on Behavioral Data and Prefrontal Hemodynamic Activity
4.2. Effect of CRF Level on Behavioral Data and Prefrontal Hemodynamic Activity
4.3. Relationships between Behavioral Data and Prefrontal Hemodynamic Activity
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kausler, D.H. Learning and Memory in Normal Aging; Academic Press: San Diego, CA, USA, 1994. [Google Scholar]
- Phillips, L.H.; Henry, J.D. Adult aging and executive functioning. In Executive Functions and The Frontal Lobes: A Lifespan Perspective; Anderson, V., Jacobs, R., Anderson, P.J., Eds.; Taylor & Francis: New York, NY, USA, 2008; pp. 57–79. [Google Scholar]
- Turner, G.R.; Spreng, R.N. Executive functions and neurocognitive aging: dissociable patterns of brain activity. Neurobiol. Aging 2012, 33, 826-e1–826-e13. [Google Scholar] [CrossRef]
- Verhaeghen, P.; Cerella, J. Aging, executive control, and attention: A review of meta-analyses. Neurosci. Biobehav. Rev. 2002, 26, 849–857. [Google Scholar] [CrossRef]
- Grady, C.L. Cognitive neuroscience of aging. Ann. N. Y. Acad. Sci. 2008, 1124, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Park, D.C.; Reuter-Lorenz, P. The adaptive brain: Aging and neurocognitive scaffolding. Annu. Rev. Psychol. 2009, 60, 173–196. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; Anderson, N.D.; Locantore, J.K.; McIntosh, A.R. Aging gracefully: Compensatory brain activity in high-performing older adults. Neuroimage 2002, 17, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.; Daselaar, S.M.; Dolcos, F.; Prince, S.E.; Budde, M.; Nyberg, L. Task-independent and task-specific age effects on brain activity during working memory, visual attention and episodic retrieval. Cereb. Cortex 2004, 14, 364–375. [Google Scholar] [CrossRef]
- Grady, C. The cognitive neuroscience of ageing. Nat. Rev. Neurosci. 2012, 13, 491–505. [Google Scholar] [CrossRef]
- Cabeza, R. Hemispheric asymmetry reduction in older adults: The HAROLD model. Psychol. Aging 2002, 17, 85–100. [Google Scholar] [CrossRef]
- Bäckman, L.; Almkvist, O.; Andersson, J.; Nordberg, A.; Windblad, B.; Rineck, R.; Lågström, B. Brain activation in young and older adults during implicit and explicit retrieval. J. Cogn. Neurosci. 1997, 9, 378–391. [Google Scholar] [CrossRef]
- Grady, C.L.; McIntosh, A.R.; Horwitz, B.; Rapoport, S.I. Age-related changes in the neural correlates of degraded and nondegraded face processing. Cogn. Neuropsychol. 2000, 217, 165–186. [Google Scholar] [CrossRef]
- Grady, C.L.; Bernstein, L.J.; Beig, S.; Siegenthaler, A.L. The effects of encoding strategy on age-related changes in the functional neuroanatomy of face memory. Psychol. Aging 2002, 17, 7–23. [Google Scholar] [CrossRef]
- Logan, J.M.; Sanders, A.L.; Snyder, A.Z.; Morris, J.C.; Buckner, R.L. Under-recruitment and nonselective recruitment: Dissociable neural mechanisms associated with aging. Neuron 2002, 33, 827–840. [Google Scholar] [CrossRef]
- Madden, D.J.; Turkington, T.G.; Provenzale, J.M.; Denny, L.L.; Hawk, T.C.; Gottlob, L.R.; Coleman, R.E. Adult age differences in functional neuroanatomy of verbal recognition memory. Hum. Brain Mapp. 1999, 7, 115–135. [Google Scholar] [CrossRef]
- Nielson, K.A.; Langenecker, S.A.; Garavan, H.P. Differences in the functional neuroanatomy of inhibitory control across the adult lifespan. Psychol. Aging 2002, 17, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Reuter-Lorenz, P.; Jonides, J.; Smith, E.S.; Hartley, A.; Miller, A.; Marshuetz, C.; Koeppe, R.A. Age differences in the frontal lateralization of verbal and spatial working memory revealed by PET. J. Cogn. Neurosci. 2000, 12, 174–187. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, G.T.; Carrillo, M.C.; Dorman, J.; Dirksen, C.; Desond, J.; Turner, D.A.; Bennett, D.A.; Wilson, R.S.; Glover, G.; Gabrieli, D.E. Aging effects on memory encoding in the frontal lobes. Psychol. Aging 2002, 17, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-C.; Lindenberger, U. Cross-level unification: A computational exploration of the link between deterioration of neurotransmitter systems dedifferentiation of cognitive abilities in old age. In Cognitive Neuroscience of Memory, Nilsson, L.G., Markowitsch, H.J., Eds.; Hogrefe & Huber: Seattle, WA, USA, 1999; pp. 103–146. [Google Scholar]
- Cabeza, R.; Nyberg, L. Imaging Cognition II: An empirical review of 275 PET and fMRI studies. J. Cogn. Neurosci. 2000, 12, 1–47. [Google Scholar] [CrossRef]
- Nolde, S.F.; Johnson, M.K.; Raye, C.L. The role of prefrontal cortex during tests of episodic memory. Trends Cogn. Sci. 1998, 2, 399–406. [Google Scholar] [CrossRef]
- Cabeza, R.; Albert, M.; Belleville, S.; Craik, F.I.M.; Duarte, A.; Grady, C.L.; Lindenberger, U.; Nyberg, L.; Park, D.C.; Reuter-Lorenz, P.A.; et al. Maintenance, reserve and compensation: the cognitive neuroscience of healthy ageing. Nat. Rev. Neurosci. 2018, 19, 701–710. [Google Scholar] [CrossRef]
- Reuter-Lorenz, P.A.; Cappell, K.A. Neurocognitive aging and the compensation hypothesis. Curr. Dir. Psychol. Sci. 2008, 17, 177–182. [Google Scholar] [CrossRef]
- Reuter-Lorenz, P.; Park, D.C. Human neuroscience and the aging mind: A new look at old problems. J. Gerontol. 2010, 65, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Mattay, V.S.; Fera, F.; Tessitore, A.; Hariri, A.R.; Berman, K.F.; Das, S.; Meyer-Lindenberg, A.; Goldberg, T.E.; Callicott, J.H.; Weinberger, D.R. Neurophysiological correlates of age-related changes in working memory capacity. Neurosci. Lett. 2006, 392, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Garces, N.; Gordon, B.; Brumback-Peltz, C.; Shin, E.; Lee, Y.; Sutton, B.P.; Maclin, E.L.; Gratton, G.; Fabiani, M. Span, CRUNCH, and beyond: working memory capacity and the aging brain. J. Cogn. Neurosci. 2010, 22, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Angel, L.; Fay, S.; Bouazzaoui, B.; Isingrini, M. Two hemispheres for better memory in old age: role of executive functioning. J. Cogn. Neurosci. 2011, 23, 3767–3777. [Google Scholar] [CrossRef] [PubMed]
- Vermeij, A.; van Beek, A.H.; Olde Rikkert, M.G.; Claassen, J.A.; Kessels, R.P. Effects of aging on cerebral oxygenation during working-memory performance: a functional near-infrared spectroscopy study. PLoS ONE 2012, 7, e46210. [Google Scholar] [CrossRef]
- Vermeij, A.; van Beek, A.H.; Reijs, B.L.; Claassen, J.A.; Kessels, R.P. An exploratory study of the effects of spatial working-memory load on prefrontal activation in low- and high-performing elderly. Front. Aging Neurosci. 2014, 6, 303. [Google Scholar] [CrossRef] [PubMed]
- Bherer, L.; Erickson, K.I.; Liu-Ambrose, T. A review of the effects of physical activity and exercise on cognitive and brain functions in older adults. J. Aging Res. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Deweerdt, S. Prevention: Activity is the best medicine. Nature 2011, 475, S16–S17. [Google Scholar] [CrossRef] [PubMed]
- Hayes, S.M.; Hayes, J.P.; Cadden, M.; Verfaellie, M. A review of cardiorespiratory fitness-related neuroplasticity in the aging brain. Front. Aging Neurosci. 2013, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Hillman, C.H.; Erickson, K.I.; Kramer, A.F. Be smart, exercise your heart: Exercise effects on brain and cognition. Nat. Rev. Neurosci. 2008, 9, 58–65. [Google Scholar] [CrossRef]
- Abou-Dest, A.; Albinet, C.T.; Boucard, G.; Audiffren, M. Swimming as a positive moderator of cognitive aging: A cross-sectional study with a multitask approach. J. Aging Res. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Albinet, C.T.; Boucard, G.; Bouquet, C.A.; Audiffren, M. Increased heart rate variability and executive performance after aerobic training in the elderly. Eur. J. Appl. Physiol. 2010, 109, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Albinet, C.T.; Mandrick, K.; Bernard, P.L.; Perrey, S.; Blain, H. Improved cerebral oxygenation response and executive performance as a function of cardiorespiratory fitness in older women: a fNIRS study. Front. Aging Neurosci. 2014, 6, 272. [Google Scholar] [CrossRef] [PubMed]
- Colcombe, S.J.; Kramer, A.F. Fitness effects on the cognitive function of older adults: A meta-analytic study. Psychol. Sci. 2003, 14, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.F.; Hahn, S.; Cohen, N.J.; Banich, M.T.; McAuley, E.; Harrison, C.R.; Chason, J.; Vakil, E.; Bardell, L.; Boileau, R.A.; et al. Ageing, fitness and neurocognitive function. Nature 1999, 400, 418–419. [Google Scholar] [CrossRef] [PubMed]
- Prakash, R.S.; Voss, M.W.; Erickson, K.I.; Lewis, J.M.; Chaddock, L.; Malkowski, E.; Alves, H.; Kim, J.; Szabo, A.; White, S.M.; et al. Cardiorespiratory fitness and attentional control in the aging brain. Front. Hum. Neurosci. 2011, 4, 229. [Google Scholar] [CrossRef] [PubMed]
- Van Praag, H.; Kempermann, G.; Gage, F.H. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat. Neurosci. 1999, 2, 266–270. [Google Scholar] [CrossRef]
- Voss, M.W.; Vivar, C.; Kramer, A.F.; van Praag, H. Bridging animal and human models of exercise-induced brain plasticity. Trends Cogn. Sci. 2013, 17, 525–544. [Google Scholar] [CrossRef]
- Colcombe, S.J.; Kramer, A.F.; Erickson, K.I.; Scalf, P.; McAuley, E.; Cohen, N.J.; Webb, A.; Jerome, G.J.; Marquez, D.X.; Elavsky, S. Cardiovascular fitness, cortical plasticity, and aging. Proc. Natl. Acad. Sci. USA 2004, 101, 3316–3321. [Google Scholar] [CrossRef]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef]
- Voelcker-Rehage, C.; Niemann, C. Structural and functional brain changes related to different types of physical activity across the life span. Neurosci. Biobehav. Rev. 2013, 37, 2268–2295. [Google Scholar] [CrossRef] [PubMed]
- Voss, M.W.; Prakash, R.S.; Erickson, K.I.; Basak, C.; Chaddock, L.; Kim, J.S.; Alves, H.; Heo, S.; Szabo, A.N.; White, S.M.; et al. Plasticity of brain networks in a randomized intervention trial of exercise training in older adults. Front. Aging Neurosci. 2010, 2, pii:32. [Google Scholar] [CrossRef] [PubMed]
- Hyodo, K.; Dan, I.; Kyutoku, Y.; Suwabe, K.; Byun, K.; Ochi, G.; Kato, M.; Soya, H. The association between aerobic fitness and cognitive function in older men mediated by frontal lateralization. Neuroimage 2016, 125, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Voelcker-Rehage, C.; Godde, B.; Staudinger, U.M. Physical and motor fitness are both related to cognition in old age. Eur. J. Neurosci. 2010, 31, 167–176. [Google Scholar] [CrossRef]
- Nagel, I.E.; Preuschhof, C.; Li, S.C.; Nyberg, L.; Bäckman, L.; Lindenberger, U.; Heekeren, H.R. Load modulation of BOLD response and connectivity predicts working memory performance in younger and older adults. J. Cogn. Neurosci. 2011, 23, 2030–2045. [Google Scholar] [CrossRef] [PubMed]
- Albinet, C.T.; Abou-Dest, A.; André, N.; Audiffren, M. Executive functions improvement following a 5-month aquaerobics program in older adults: Role of cardiac vagal control in inhibition performance. Biol. Psychol. 2016, 115, 69–77. [Google Scholar] [CrossRef]
- Gothe, N.P.; Kramer, A.F.; McAuley, E. The effects of an 8-week Hatha yoga intervention on executive function in older adults. J. Gerontol. 2014, 69, 1109–1116. [Google Scholar] [CrossRef]
- Dupuy, O.; Gauthier, C.J.; Fraser, S.A.; Desjardins-Crèpeau, L.; Desjardins, M.; Mekary, S.; Lesage, F.; Hoge, R.D.; Pouliot, P.; Bherer, L. Higher levels of cardiovascular fitness are associated with better executive function and prefrontal oxygenation in younger and older women. Front. Hum. Neurosci. 2015, 9, 66. [Google Scholar] [CrossRef]
- Agbangla, N.F.; Audiffren, M.; Albinet, C.T. Use of near-infrared spectroscopy in the investigation of brain activation during cognitive aging: A systematic review of an emerging area of research. Ageing Res. Rev. 2017, 38, 52–66. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Hébert, R.; Bravo, G.; Girouard, D. Validation de l’adaptation française du modified mini-mental state (3MS). Rev. Geriatr. 1992, 17, 443–450. [Google Scholar]
- Bourque, P.; Blanchard, L.; Vézina, J. Étude psychométrique de l’Échelle de dépression gériatrique. Can. J. Aging 1990, 9, 348–355. [Google Scholar] [CrossRef]
- Boucard, G.K.; Albinet, C.T.; Bugaiska, A.; Bouquet, C.A.; Clarys, D.; Audiffren, M. Impact of physical activity on executive functions in aging: A selective effect on inhibition among old adults. J. Sport Exerc. Psychol. 2012, 34, 808–827. [Google Scholar] [CrossRef] [PubMed]
- Albinet, C.T.; Boucard, G.; Bouquet, C.A.; Audiffren, M. Processing speed and executive functions in cognitive aging: How to disentangle their mutual relationship? Brain Cogn. 2012, 79, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fishburn, F.A.; Norr, M.E.; Medvedev, A.V.; Vaidya, C.J. Sensitivity of fNIRS to cognitive state and load. Front. Hum. Neurosci. 2014, 8, 76. [Google Scholar] [CrossRef]
- Herff, C.; Heger, D.; Fortmann, O.; Hennrich, J.; Putze, F.; Schultz, T. Mental workload during N-back task-quantified in the prefrontal cortex using fNIRS. Front. Hum. Neurosci. 2014, 7, 935. [Google Scholar] [CrossRef]
- Molteni, E.; Contini, D.; Caffini, M.; Baselli, G.; Spinelli, L.; Cubeddu, R.; Cerutti, S.; Bianchi, A.M.; Torricelli, A. Load-dependent brain activation assessed by time-domain functional near-infrared spectroscopy during a working memory task with graded levels of difficulty. J. Biomed. Opt. 2012, 17, 056005. [Google Scholar] [CrossRef] [PubMed]
- Jonides, J.; Schumacher, E.H.; Smith, E.E.; Lauber, E.J.; Awh, E.; Minoshima, S.; Koeppe, R.A. Verbal working memory load affects regional brain activation as measured by PET. J. Cogn. Neurosci. 1997, 9, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Léger, L.A.; Lambert, J. A maximal multistage 20-m shuttle run test to predict VO2 max. Eur. J. Appl. Physiol. 1982, 49, 1–12. [Google Scholar] [CrossRef]
- Jackson, A.S.; Blair, S.N.; Mahar, M.T.; Wier, L.T.; Ross, R.M.; Stuteville, J.E. Prediction of functional aerobic capacity without exercise testing. Med. Sci. Sports Exerc. 1990, 22, 863–870. [Google Scholar] [CrossRef]
- Shvartz, E.; Reibold, R.C. Aerobic fitness norms for males and females aged 6 to 75 years: A review. Aviat. Space Environ. Med. 1990, 61, 3–11. [Google Scholar] [PubMed]
- Shinba, T.; Nagano, M.; Kariya, N.; Ogawa, K.; Shinozaki, T.; Shimosato, S.; Hoshi, Y. Near-infrared spectroscopy analysis of frontal lobe dysfunction in schizophrenia. Biol. Psychiatry 2004, 55, 154–164. [Google Scholar] [CrossRef]
- Duncan, A.; Meek, J.H.; Clemence, M.; Elwell, C.E.; Fallon, P.; Tyszczuk, L.; Cope, M.; Delpy, D.T. Measurement of cranial optical path length as a function of age using phase resolved near infrared spectroscopy. Pediatr. Res. 1996, 39, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Dan, H.; Sakamoto, K.; Takeo, K.; Shimizu, K.; Kohno, S.; Oda, I.; Isobe, S.; Suzuki, T.; Kohyama, K.; et al. Three-dimensional probabilistic anatomical cranio-cerebral correlation via the international 10-20 system oriented for transcranial functional brain mapping. Neuroimage 2004, 21, 99–111. [Google Scholar] [CrossRef]
- Scholkmann, F.; Spichtig, S.; Muehlemann, T.; Wolf, M. How to detect and reduce movement artifacts in near-infrared imaging using moving standard deviation and spline interpolation. Physiol. Meas. 2010, 31, 649–662. [Google Scholar] [CrossRef]
- Watanabe, H.; Homae, F.; Nakano, T.; Taga, G. Functional activation in diverse regions of the developing brain of human infants. Neuroimage 2008, 43, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, Y.; Tsou, B.H.; Billock, V.A.; Tanosaki, M.; Iguchi, Y.; Shimada, M.; Shinba, T.; Yamada, Y.; Oda, I. Spatiotemporal characteristics of hemodynamic changes in the human lateral prefrontal cortex during working memory tasks. Neuroimage 2003, 20, 1493–1504. [Google Scholar] [CrossRef]
- Stanislaw, H.; Todorov, N. Calculation of signal detection theory measures. Behav. Res. Methods Instrum. Comput. 1999, 31, 137–149. [Google Scholar] [CrossRef]
- Delignières, D.; Famose, J.P.; Genty, J. Validation d’une échelle de catégories pour la perception de la difficulté. Rev. S.T.A.P.S 1994, 34, 77–88. [Google Scholar]
- R: A Language and Environment for Statistical Computing; Version 3.5.2.; R Foundation for Statistical Computing; The R Foundation Conference Committee: Vienna, Austria, 2018.
- Mendenhall, W.; Sincich, T. A Second Course in Statistics: Regression Analysis, 7th ed.; Pearson: Boston, MA, USA, 2012. [Google Scholar]
- Artuso, C.; Cavallini, E.; Bottiroli, S.; Palladino, P. Updating working memory: Memory load matters with aging. Aging Clin. Exp. Res. 2017, 29, 371–377. [Google Scholar] [CrossRef]
- De Beni, R.; Palladino, P. Decline in working memory updating through ageing: Intrusion error analyses. Memory 2004, 12, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Sorel, O.; Pennequin, V. Aging of the planning process: The role of executive functioning. Brain Cogn. 2008, 66, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Van Der Linden, M.; Brédart, S.; Beerten, A. Age-related differences in updating working memory. Br. J. Psychol. 1994, 85, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, M.; Manenti, R.; Ferrari, C.; Cotelli, M. Better together: Left and right hemisphere engagement to reduce age-related memory loss. Behav. Brain Res. 2015, 293, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Audiffren, M.; André, N.; Albinet, C.T. Positive effects of chronic physical exercise on cognitive functions in aging people: Assessment and prospects. Rev. Neuropsychol. 2011, 3, 207–225. [Google Scholar] [CrossRef]
- Brown, A.D.; McMorris, C.A.; Longman, R.S.; Leigh, R.; Hill, M.D.; Friedenreich, C.M.; Poulin, M.J. Effects of cardiorespiratory fitness and cerebral blood flow on cognitive outcomes in older women. Neurobiol. Aging 2010, 31, 2047–2057. [Google Scholar] [CrossRef] [PubMed]
- Dustman, R.E.; Ruhling, R.O.; Russell, E.M.; Shearer, D.E.; Bonekat, H.W.; Shigeoka, J.W.; Wood, J.S.; Bradford, D.C. Aerobic exercise training and improved neuropsychological function of older individuals. Neurobiol. Aging 1984, 5, 35–42. [Google Scholar] [CrossRef]
- Perrey, S. Non-invasive NIR spectroscopy of human brain function during exercise. Methods 2008, 45, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Tachtsidis, I.; Scholkmann, F. False positives and false negatives in functional near-infrared spectroscopy: Issues, challenges, and the way forward. Neurophotonics 2016, 3, 031405. [Google Scholar] [CrossRef]
- Friston, K.J. Functional and effective connectivity: A review. Brain Connect. 2011, 1, 13–36. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young Adults (N = 19) | Older Adults (N = 37) | Older High-Fit (N = 21) | Older Low-Fit (N = 16) | p Value | |
|---|---|---|---|---|---|
| Age (years) | 19.7 ± 1 | 68.95 ± 4.74 | 67.90 ± 4.86 | 70.31 ± 4.33 | p = 0.12 |
| Gender (M/F) | 17/2 | 15/22 | 8/13 | 7/9 | p = 0.79 |
| Education (years) | 14.00 ± 0.00 | 13.35 ± 3.85 | 14.52 ± 3.57 | 11.81 ± 3.75 | p = 0.031 * |
| VO2max (mL/Kg/min) | 54.83 ± 7.21 | 22.31 ± 7.88 | 26.10 ± 6.73 | 17.40 ± 6.58 | p = 0.0004 * |
| MMSE | 29.24 ± 0.95 | 29.29 ± 1.01 | 29.19 ± 0.91 | p = 0.76 | |
| GDS | 5.78 ± 4.25 | 5.95 ± 4.86 | 5.56 ± 3.42 | p = 0.78 |
| Young Adults (N = 19) | Older Adults (N = 37) | Older High-Fit (N = 21) | Older Low-Fit (N = 16) | ||
|---|---|---|---|---|---|
| RT (ms) | 0-back | 366.77 ± 43.46 | 509.41 ± 72.79 | 499.84 ± 67.38 | 521.00 ± 79.95 |
| 1-back | 404.39 ± 73.68 | 661.39 ± 161.77 * | 655.22 ± 172.32 | 669.50 ± 151.95 | |
| 2-back | 517.46 ± 152.98 | 970.15 ± 316.46 * | 959.12 ± 304.53 | 984.63 ± 341.04 | |
| 3-back | 592.72 ± 186.32 | 1110.80 ± 370.47 * | 1114.56 ± 412.97 | 1105.86 ± 319.30 | |
| A’ | 0-back | 0.99 ± 0.01 | 0.99 ± 0.01 | 0.99 ± 0.01 | 0.99 ± 0.01 |
| 1-back | 0.98 ± 0.02 | 0.96 ± 0.04 | 0.97 ± 0.03 | 0.95 ± 0.05 | |
| 2-back | 0.93 ± 0.06 | 0.90 ± 0.09 | 0.90 ± 0.07 | 0.89 ± 0.12 | |
| 3-back | 0.85 ± 0.06 | 0.75 ± 0.16 * | 0.79 ± 0.1 | 0.69 ± 0.20 ** | |
| Perceived difficulty | 0-back | 2.24 ± 1.17 | 2.51 ± 1.26 | 2.33 ± 1.35 | 2.75 ± 1.13 |
| 1-back | 4.42 ± 2.17 | 5.24 ± 1.94 | 5.24 ± 2.21 | 5.25 ± 1.57 | |
| 2-back | 7.74 ± 1.63 | 8.51 ± 2.27 | 8.95 ± 2.16 | 7.94 ± 2.53 | |
| 3-back | 10.74 ± 1.99 | 11.49 ± 1.79 | 11.86 ± 1.62 | 11.00 ± 1.93 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agbangla, N.F.; Audiffren, M.; Pylouster, J.; Albinet, C.T. Working Memory, Cognitive Load and Cardiorespiratory Fitness: Testing the CRUNCH Model with Near-Infrared Spectroscopy. Brain Sci. 2019, 9, 38. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9020038
Agbangla NF, Audiffren M, Pylouster J, Albinet CT. Working Memory, Cognitive Load and Cardiorespiratory Fitness: Testing the CRUNCH Model with Near-Infrared Spectroscopy. Brain Sciences. 2019; 9(2):38. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9020038
Chicago/Turabian StyleAgbangla, Nounagnon Frutueux, Michel Audiffren, Jean Pylouster, and Cédric T. Albinet. 2019. "Working Memory, Cognitive Load and Cardiorespiratory Fitness: Testing the CRUNCH Model with Near-Infrared Spectroscopy" Brain Sciences 9, no. 2: 38. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9020038