Western-Type Helicobacter pylori CagA are the Most Frequent Type in Mongolian Patients


Abstract
:1. Introduction
2. Results
2.1. Patient Demographics
2.2. CagA Genotypes
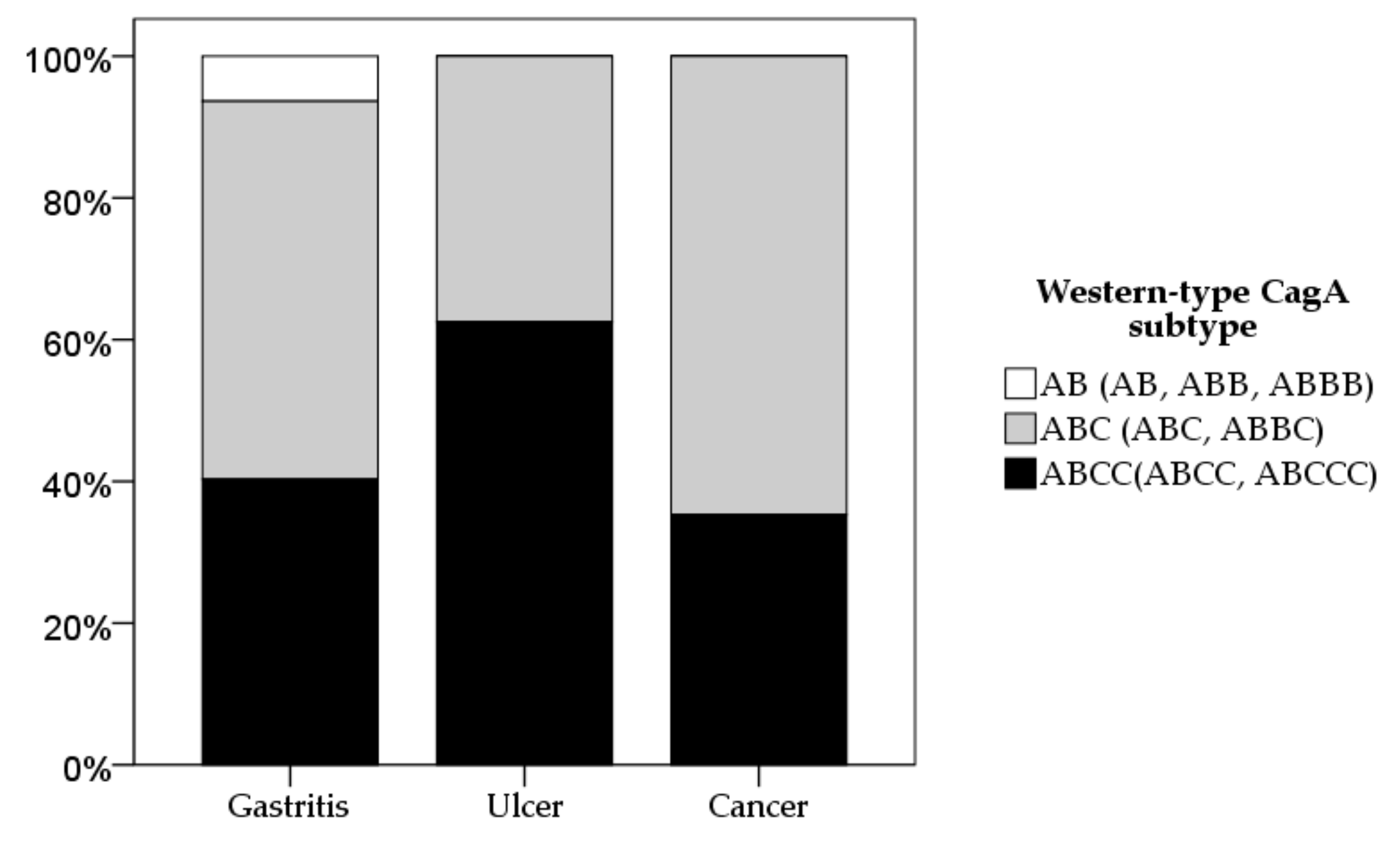
2.3. CagA and vacA Genotyping Based on Diseases
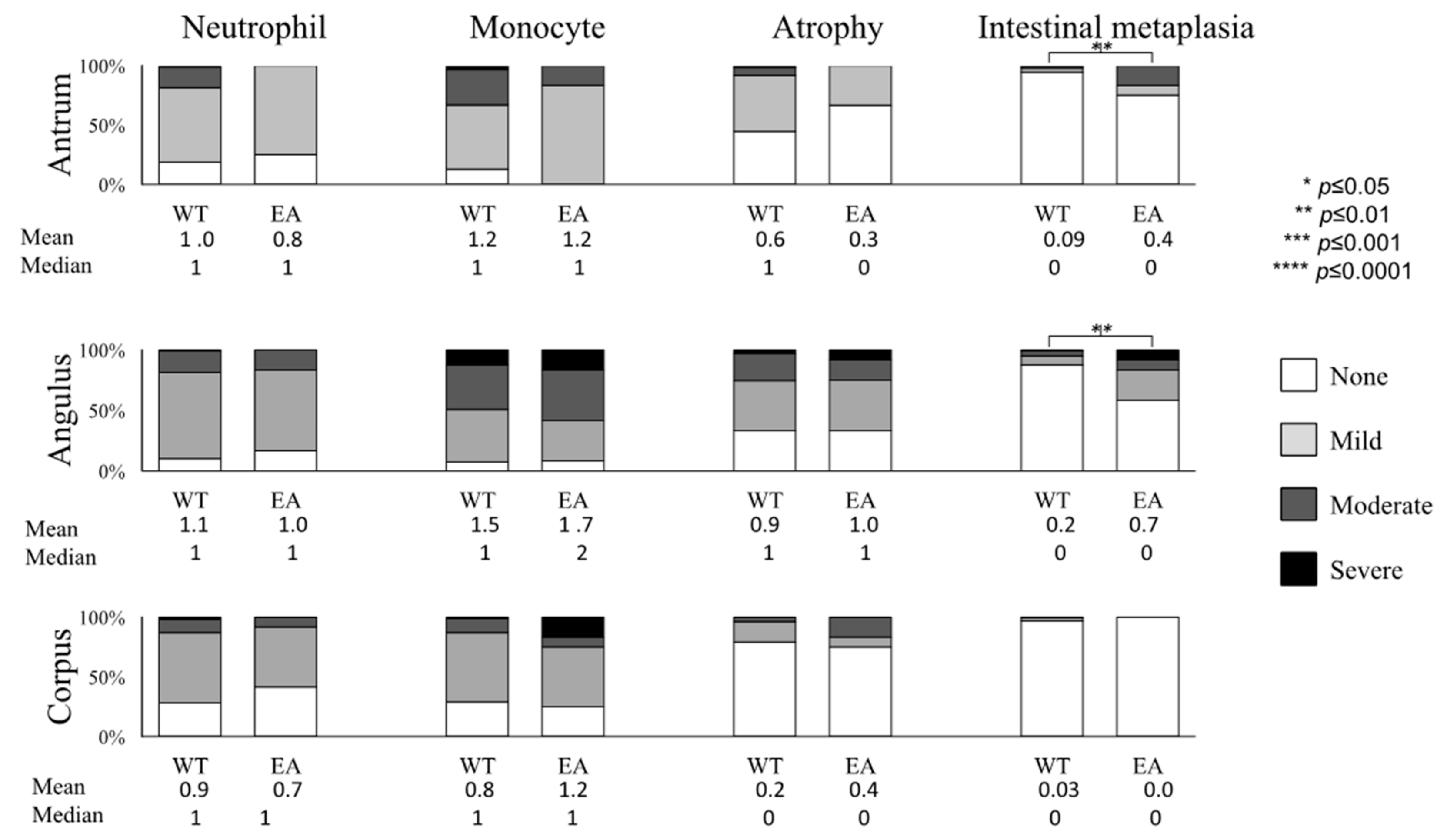
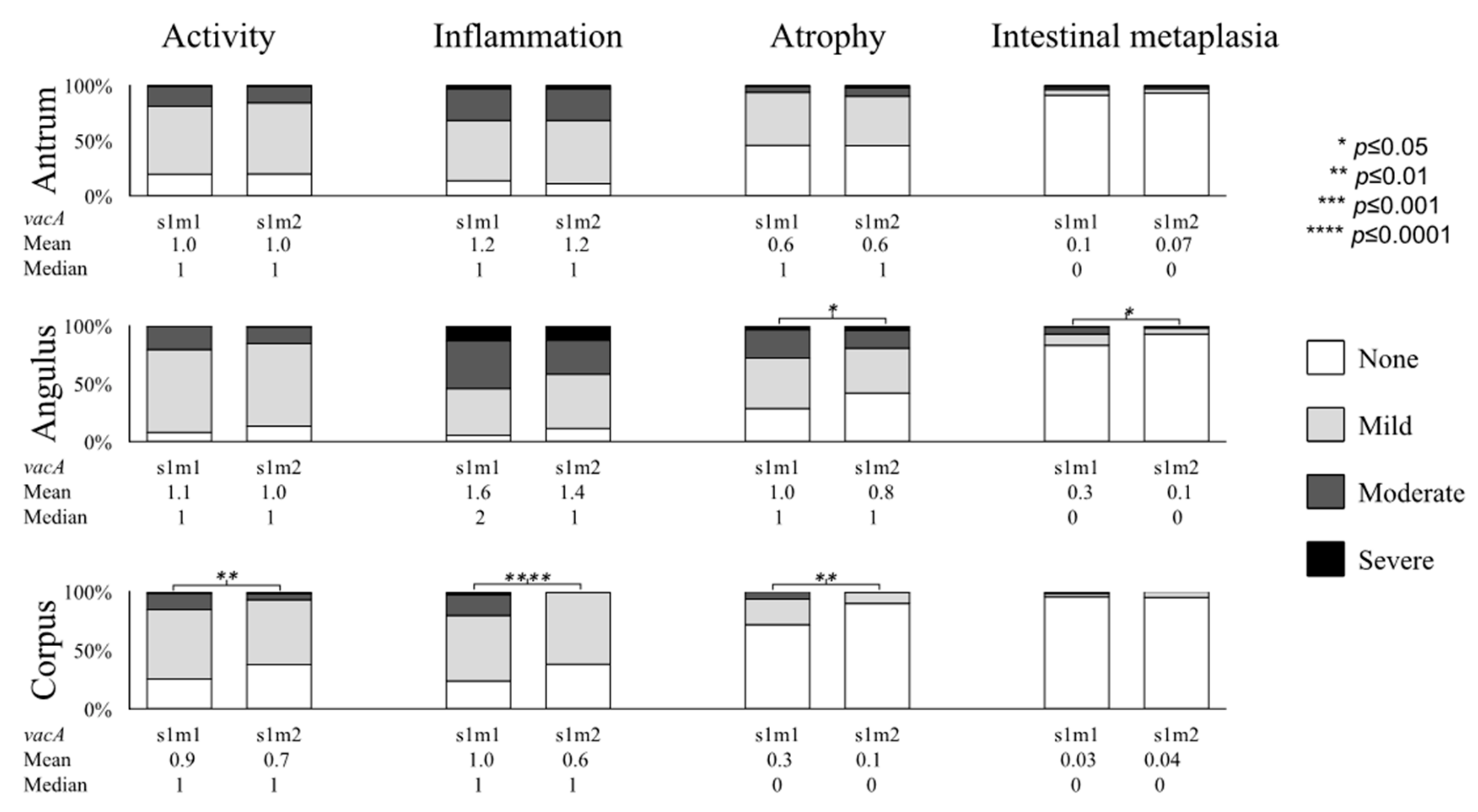
2.4. Gastric Mucosal Status with Respect to H. pylori Virulence Factors
3. Discussion
4. Materials and Methods
4.1. Study Population and Sampling
4.2. Histologic Diagnosis for Gastritis and Gastric Cancer
4.3. H. pylori Isolation and Sequencing
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Plummer, M.; Franceschi, S.; Vignat, J.; Forman, D.; de Martel, C. Global burden of gastric cancer attributable to Helicobacter pylori. Int. J. Cancer 2015, 136, 487–490. [Google Scholar]
- Yamaoka, Y. Mechanisms of disease: Helicobacter pylori virulence factors. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 629. [Google Scholar] [CrossRef]
- Backert, S.; Blaser, M.J. The role of CagA in the gastric biology of Helicobacter Pylori. Cancer Res. 2016, 76, 4028–4031. [Google Scholar] [CrossRef]
- Hatakeyama, M. Oncogenic mechanisms of the Helicobacter pylori CagA protein. Nat. Rev. Cancer 2004, 4, 688. [Google Scholar] [CrossRef]
- Hatakeyama, M. Anthropological and clinical implications for the structural diversity of the Helicobacter pylori CagA oncoprotein. Cancer Sci. 2011, 102, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Backert, S.; Clyne, M.; Tegtmeyer, N. Molecular mechanisms of gastric epithelial cell adhesion and injection of CagA by Helicobacter pylori. Cell Commun. Signal. 2011, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y.; Kodama, T.; Kashima, K.; Graham, D.Y.; Sepulveda, A.R. Variants of the 3′ region of the cagA gene in Helicobacter pylori isolates from patients with different H. pylori-associated diseases. J. Clin. Microbiol. 1998, 36, 2258–2263. [Google Scholar] [PubMed]
- Atherton, J.C.; Cao, P.; Peek, R.M.; Tummuru, M.K.; Blaser, M.J.; Cover, T.L. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori association of specific vacA types with cytotoxin production and peptic ulceration. J. Biol. Chem. 1995, 270, 17771–17777. [Google Scholar] [CrossRef]
- Thi Huyen Trang, T.; Thanh Binh, T.; Yamaoka, Y. Relationship between vacA types and development of gastroduodenal diseases. Toxins 2016, 8, 182. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Yamaoka, Y. The association of vacA genotype and Helicobacter pylori-related disease in Latin American and African populations. Clin. Microbiol. Infect. 2009, 15, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Khasag, O.; Boldbaatar, G.; Tegshee, T.; Duger, D.; Dashdorj, A.; Uchida, T.; Matsuhisa, T.; Yamaoka, Y. The prevalence of Helicobacter pylori infection and other risk factors among Mongolian dyspeptic patients who have a high incidence and mortality rate of gastric cancer. Gut Pathog. 2018, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Matsuhisa, T.; Yamaoka, Y.; Uchida, T.; Duger, D.; Adiyasuren, B.; Khasag, O.; Tegshee, T.; Tsogt-Ochir, B. Gastric mucosa in Mongolian and Japanese patients with gastric cancer and Helicobacter pylori infection. World J. Gastroenterol. 2015, 21, 8408–8417. [Google Scholar] [CrossRef]
- Gantuya, B.; Bolor, D.; Oyuntsetseg, K.; Erdene-Ochir, Y.; Sanduijav, R.; Davaadorj, D.; Tserentogtokh, T.; Azzaya, D.; Uchida, T.; Matsuhisa, T. New observations regarding Helicobacter pylori and gastric cancer in Mongolia. Helicobacter 2018, 23, e12491. [Google Scholar] [CrossRef]
- Dixon, M.F.; Genta, R.M.; Yardley, J.H.; Correa, P. Classification and grading of gastritis: The updated Sydney system. Am. J. Surg. Pathol. 1996, 20, 1161–1181. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, M. Structure and function of Helicobacter pylori CagA, the first-identified bacterial protein involved in human cancer. Proc. Jpn. Acad. Ser. B 2017, 93, 196–219. [Google Scholar] [CrossRef]
- Matsunari, O.; Shiota, S.; Suzuki, R.; Watada, M.; Kinjo, N.; Murakami, K.; Fujioka, T.; Kinjo, F.; Yamaoka, Y. Association between Helicobacter pylori virulence factors and gastroduodenal diseases in Okinawa, Japan. J. Clin. Microbiol. 2012, 50, 876–883. [Google Scholar] [CrossRef]
- Yamaoka, Y.; El–Zimaity, H.M.; Gutierrez, O.; Figura, N.; Kim, J.K.; Kodama, T.; Kashima, K.; Graham, D.Y. Relationship between the cagA 3′repeat region of Helicobacter pylori, gastric histology, and susceptibility to low pH. Gastroenterology 1999, 117, 342–349. [Google Scholar] [CrossRef]
- Batista, S.A.; Rocha, G.A.; Rocha, A.M.; Saraiva, I.E.; Cabral, M.M.; Oliveira, R.C.; Queiroz, D.M. Higher number of Helicobacter pylori CagA EPIYA C phosphorylation sites increases the risk of gastric cancer, but not duodenal ulcer. BMC Microbiol. 2011, 11, 61. [Google Scholar] [CrossRef]
- Blaser, M.J.; Perez-Perez, G.I.; Kleanthous, H.; Cover, T.L.; Peek, R.M.; Chyou, P.; Stemmermann, G.N.; Nomura, A. Infection with Helicobacter pylori strains possessing cagA is associated with an increased risk of developing adenocarcinoma of the stomach. Cancer Res. 1995, 55, 2111–2115. [Google Scholar]
- Parsonnet, J.; Friedman, G.; Orentreich, N.; Vogelman, H. Risk for gastric cancer in people with CagA positive or CagA negative Helicobacter pylori infection. Gut 1997, 40, 297–301. [Google Scholar] [CrossRef]
- Huang, J.Q.; Zheng, G.F.; Sumanac, K.; Irvine, E.J.; Hunt, R.H. Meta-analysis of the relationship between cagA seropositivity and gastric cancer. Gastroenterology 2003, 125, 1636–1644. [Google Scholar] [CrossRef]
- Binh, T.T.; Tuan, V.P.; Dung, H.D.Q.; Tung, P.H.; Tri, T.D.; Thuan, N.P.M.; Van Khien, V.; Hoan, P.Q.; Suzuki, R.; Uchida, T. Advanced non-cardia gastric cancer and Helicobacter pylori infection in Vietnam. Gut Pathog. 2017, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y.; Graham, D.Y. Helicobacter pylori virulence and cancer pathogenesis. Future Oncol. 2014, 10, 1487–1500. [Google Scholar] [CrossRef]
- Inoue, M.; Tsugane, S. Epidemiology of gastric cancer in Japan. Postgrad. Med. J. 2005, 81, 419–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, P. Human gastric carcinogenesis: A multistep and multifactorial process--First American Cancer Society Award Lecture on Cancer Epidemiology and Prevention. Cancer Res. 1992, 52, 6735–6740. [Google Scholar]
- Gantuya, B.; Oyuntsetseg, K.; Bolor, D.; Erdene-Ochir, Y.; Sanduijav, R.; Davaadorj, D.; Tserentogtokh, T.; Uchida, T.; Yamaoka, Y. Evaluation of serum markers for gastric cancer and its precursor diseases among high incidence and mortality rate of gastric cancer area. Gastric Cancer 2015, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sharaf, R.N.; Shergill, A.K.; Odze, R.D.; Krinsky, M.L.; Fukami, N.; Jain, R.; Appalaneni, V.; Anderson, M.A.; Ben-Menachem, T.; Chandrasekhara, V. Endoscopic mucosal tissue sampling. Gastrointest. Endosc. 2013, 78, 216–224. [Google Scholar] [CrossRef]
- Lauren, P. The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma: An attempt at a histo-clinical classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef]
- Association, J.G.C. Japanese classification of gastric carcinoma–2nd English edition. Gastric Cancer 1998, 1, 10–24. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CagA Types | Gastric Cancer n = 17 (%) | Gastritis n = 341 (%) | Peptic Ulcer n = 10 (%) | Total n = 368 (%) | |
|---|---|---|---|---|---|
| Negative (−) | 0 (0) | 62 (18.2) | 0 (0) | 62 (16.8) | |
| Western | a AB | 0 (0) | 17 (5.0) | 0 (0) | 17 (4.7) |
| b ABC | 11 (64.7) | 142 (41.6) | 3 (30) | 156 (42.4) | |
| c ABC* | 6 (35.3) | 109 (32.0) | 5 (50) | 120 (32.6) | |
| East-Asian | d ABD | 0 (0) | 10 (2.9) | 2(20) | 12 (3.2) |
| Hybrid | ABDC | 0 (0) | 1 (0.3) | 0 (0) | 1 (0.3) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tserentogtokh, T.; Gantuya, B.; Subsomwong, P.; Oyuntsetseg, K.; Bolor, D.; Erdene-Ochir, Y.; Azzaya, D.; Davaadorj, D.; Uchida, T.; Matsuhisa, T.; et al. Western-Type Helicobacter pylori CagA are the Most Frequent Type in Mongolian Patients. Cancers 2019, 11, 725. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11050725
Tserentogtokh T, Gantuya B, Subsomwong P, Oyuntsetseg K, Bolor D, Erdene-Ochir Y, Azzaya D, Davaadorj D, Uchida T, Matsuhisa T, et al. Western-Type Helicobacter pylori CagA are the Most Frequent Type in Mongolian Patients. Cancers. 2019; 11(5):725. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11050725
Chicago/Turabian StyleTserentogtokh, Tegshee, Boldbaatar Gantuya, Phawinee Subsomwong, Khasag Oyuntsetseg, Dashdorj Bolor, Yansan Erdene-Ochir, Dashdorj Azzaya, Duger Davaadorj, Tomohisa Uchida, Takeshi Matsuhisa, and et al. 2019. "Western-Type Helicobacter pylori CagA are the Most Frequent Type in Mongolian Patients" Cancers 11, no. 5: 725. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11050725