Analysis of Promoter-Associated Chromatin Interactions Reveals Biologically Relevant Candidate Target Genes at Endometrial Cancer Risk Loci

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. H3K27Ac HiChIP Analysis Identifies Promoter-Associated Chromatin Loops in Endometrial Cell Lines
2.2. HiChIP Promoter Loops Are Enriched for Endometrial Cancer Heritability
2.3. HiChIP Promoter Looping Reveals 103 Candidate Target Genes at Endometrial Cancer Risk Loci
2.4. HiChIP Target Genes Are Enriched for Potential Targets of a Mirna Encoded by the HiChIP Target Gene Mir196a1
2.5. HiChIP Target Genes Are Differentially Expressed in Endometrial Tumors
2.6. HiChIP Target Gene Expression Associates with CVs
2.7. Protein-Protein Interaction Network of HiChIP Target Genes Reveals Enrichment for Endometrial Cancer Driver Genes
2.8. HiChIP Target Genes and Interacting Proteins Are Over-Represented in Relevant Biological Pathways
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Fixation
4.3. HiChIP Library Generation
4.4. HiChIP Bioinformatic Analyses
4.5. Stratified LD Score Regression Analysis
4.6. Identification and Analysis of HiChIP Target Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Mara, T.A.; Glubb, D.M.; Amant, F.; Annibali, D.; Ashton, K.; Attia, J.; Auer, P.L.; Beckmann, M.W.; Black, A.; Bolla, M.K.; et al. Identification of nine new susceptibility loci for endometrial cancer. Nat. Commun. 2018, 9, 3166. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.H.; Thompson, D.J.; O’Mara, T.A.; Painter, J.N.; Glubb, D.M.; Flach, S.; Lewis, A.; French, J.D.; Freeman-Mills, L.; Church, D.; et al. Five endometrial cancer risk loci identified through genome-wide association analysis. Nat. Genet. 2016, 48, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Spurdle, A.B.; Thompson, D.J.; Ahmed, S.; Ferguson, K.; Healey, C.S.; O’Mara, T.; Walker, L.C.; Montgomery, S.B.; Dermitzakis, E.T.; Fahey, P.; et al. Genome-wide association study identifies a common variant associated with risk of endometrial cancer. Nat. Genet. 2011, 43, 451–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Mara, T.A.; Glubb, D.M.; Kho, P.F.; Thompson, D.J.; Spurdle, A. Genome-wide association studies of endometrial cancer: Latest developments and future directions. Cancer Epidemiol. Prev. Biomark. 2019, 28, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.J.; O’Mara, T.A.; Glubb, D.M.; Painter, J.N.; Cheng, T.; Folkerd, E.; Doody, D.; Dennis, J.; Webb, P.M.; ANECS Group; et al. CYP19A1 fine-mapping and Mendelian randomization: Estradiol is causal for endometrial cancer. Endocr. Relat. Cancer 2016, 23, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Key, T.J.; Pike, M.C. The dose-effect relationship between ‘unopposed’ oestrogens and endometrial mitotic rate: Its central role in explaining and predicting endometrial cancer risk. Br. J. Cancer 1988, 57, 205–212. [Google Scholar] [CrossRef]
- Antunes, C.M.; Strolley, P.D.; Rosenshein, N.B.; Davies, J.L.; Tonascia, J.A.; Brown, C.; Burnett, L.; Rutledge, A.; Pokempner, M.; Garcia, R. Endometrial cancer and estrogen use. Report of a large case-control study. N. Engl. J. Med. 1979, 300, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Glubb, D.M.; Johnatty, S.E.; Quinn, M.C.J.; O’Mara, T.A.; Tyrer, J.P.; Gao, B.; Fasching, P.A.; Beckmann, M.W.; Lambrechts, D.; Vergote, I.; et al. Analyses of germline variants associated with ovarian cancer survival identify functional candidates at the 1q22 and 19p12 outcome loci. Oncotarget 2017. [Google Scholar] [CrossRef] [PubMed]
- Glubb, D.M.; Maranian, M.J.; Michailidou, K.; Pooley, K.A.; Meyer, K.B.; Kar, S.; Carlebur, S.; O’Reilly, M.; Betts, J.A.; Hillman, K.M.; et al. Fine-scale mapping of the 5q11.2 breast cancer locus reveals at least three independent risk variants regulating MAP3K1. Am. J. Hum. Genet. 2015, 96, 5–20. [Google Scholar] [CrossRef]
- Michailidou, K.; Lindstrom, S.; Dennis, J.; Beesley, J.; Hui, S.; Kar, S.; Lemacon, A.; Soucy, P.; Glubb, D.; Rostamianfar, A.; et al. Association analysis identifies 65 new breast cancer risk loci. Nature 2017, 551, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, M.D.; Chen-Plotkin, A.S. The post-GWAS era: From association to function. Am. J. Hum. Genet. 2018, 102, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Mumbach, M.R.; Satpathy, A.T.; Boyle, E.A.; Dai, C.; Gowen, B.G.; Cho, S.W.; Nguyen, M.L.; Rubin, A.J.; Granja, J.M.; Kazane, K.R.; et al. Enhancer connectome in primary human cells identifies target genes of disease-associated DNA elements. Nat. Genet. 2017, 49, 1602–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, J.O.; Oudelaar, A.M.; Higgs, D.R.; Hughes, J.R. How best to identify chromosomal interactions: A comparison of approaches. Nat. Methods 2017, 14, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Mumbach, M.R.; Rubin, A.J.; Flynn, R.A.; Dai, C.; Khavari, P.A.; Greenleaf, W.J.; Chang, H.Y. HiChIP: Efficient and sensitive analysis of protein-directed genome architecture. Nat. Methods 2016, 13, 919–922. [Google Scholar] [CrossRef]
- Phelan, C.M.; Kuchenbaecker, K.B.; Tyrer, J.P.; Kar, S.P.; Lawrenson, K.; Winham, S.J.; Dennis, J.; Pirie, A.; Riggan, M.J.; Chornokur, G.; et al. Identification of 12 new susceptibility loci for different histotypes of epithelial ovarian cancer. Nat. Genet. 2017, 49, 680–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadaev, T.; Saunders, E.J.; Newcombe, P.J.; Anokian, E.; Leongamornlert, D.A.; Brook, M.N.; Cieza-Borrella, C.; Mijuskovic, M.; Wakerell, S.; Olama, A.A.A.; et al. Fine-mapping of prostate cancer susceptibility loci in a large meta-analysis identifies candidate causal variants. Nat. Commun. 2018, 9, 2256. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Finucane, H.K.; Schumacher, F.R.; Schmit, S.L.; Tyrer, J.P.; Han, Y.; Michailidou, K.; Lesseur, C.; Kuchenbaecker, K.B.; Dennis, J.; et al. Shared heritability and functional enrichment across six solid cancers. Nat. Commun. 2019, 10, 431. [Google Scholar] [CrossRef] [Green Version]
- Jeng, M.Y.; Mumbach, M.R.; Granja, J.M.; Satpathy, A.T.; Chang, H.Y.; Chang, A.L.S. Enhancer connectome nominates target genes of inherited risk variants from inflammatory skin disorders. J. Investig. Dermatol. 2019, 139, 605–614. [Google Scholar] [CrossRef]
- Kandoth, C.; Schultz, N.; Cherniack, A.D.; Akbani, R.; Liu, Y.; Shen, H.; Robertson, A.G.; Pashtan, I.; Shen, R.; Benz, C.C.; et al. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Bordoloi, D.; Banik, K.; Shabnam, B.; Padmavathi, G.; Monisha, J.; Arfuso, F.; Dharmarajan, A.; Mao, X.; Lim, L.H.K.; Wang, L.; et al. TIPE family of proteins and its implications in different chronic diseases. Int. J. Mol. Sci. 2018, 19, 2974. [Google Scholar] [CrossRef]
- Oberst, M.D.; Chen, L.Y.; Kiyomiya, K.; Williams, C.A.; Lee, M.S.; Johnson, M.D.; Dickson, R.B.; Lin, C.Y. HAI-1 regulates activation and expression of matriptase, a membrane-bound serine protease. Am. J. Physiol. Cell Physiol. 2005, 289, C462–C470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Võsa, U.; Claringbould, A.; Westra, H.-J.; Bonder, M.J.; Deelen, P.; Zeng, B.; Kirsten, H.; Saha, A.; Kreuzhuber, R.; Kasela, S.; et al. Unraveling the polygenic architecture of complex traits using blood eQTL meta-analysis. BioRxiv 2018. [Google Scholar] [CrossRef]
- Lim, Y.W.; Chen-Harris, H.; Mayba, O.; Lianoglou, S.; Wuster, A.; Bhangale, T.; Khan, Z.; Mariathasan, S.; Daemen, A.; Reeder, J.; et al. Germline genetic polymorphisms influence tumor gene expression and immune cell infiltration. Proc. Natl. Acad. Sci. USA 2018, 115, E11701–E11710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, M.H.; Tokheim, C.; Porta-Pardo, E.; Sengupta, S.; Bertrand, D.; Weerasinghe, A.; Colaprico, A.; Wendl, M.C.; Kim, J.; Reardon, B.; et al. Comprehensive characterization of cancer driver genes and mutations. Cell 2018, 173, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Gibson, W.J.; Hoivik, E.A.; Halle, M.K.; Taylor-Weiner, A.; Cherniack, A.D.; Berg, A.; Holst, F.; Zack, T.I.; Werner, H.M.; Staby, K.M.; et al. The genomic landscape and evolution of endometrial carcinoma progression and abdominopelvic metastasis. Nat. Genet. 2016, 48, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.E.; O’Mara, T.A.; Glubb, D.M. Enhancing the promise of drug repositioning through genetics. Front. Pharmacol. 2017, 8, 896. [Google Scholar] [CrossRef]
- Liu, T.; Li, J.; Liu, Y.; Qu, Y.; Li, A.; Li, C.; Zhang, Q.; Wu, W.; Li, J.; Liu, Y.; et al. SNX11 identified as an essential host factor for SFTS virus infection by CRISPR knockout screening. Virol. Sin. 2019. [Google Scholar] [CrossRef]
- Joyal, J.S.; Nim, S.; Zhu, T.; Sitaras, N.; Rivera, J.C.; Shao, Z.; Sapieha, P.; Hamel, D.; Sanchez, M.; Zaniolo, K.; et al. Subcellular localization of coagulation factor II receptor-like 1 in neurons governs angiogenesis. Nat. Med. 2014, 20, 1165–1173. [Google Scholar] [CrossRef]
- Barrow, J.R.; Capecchi, M.R. Targeted disruption of the Hoxb-2 locus in mice interferes with expression of Hoxb-1 and Hoxb-4. Development 1996, 122, 3817–3828. [Google Scholar]
- Nodale, C.; Ceccarelli, S.; Giuliano, M.; Cammarota, M.; D’Amici, S.; Vescarelli, E.; Maffucci, D.; Bellati, F.; Panici, P.B.; Romano, F.; et al. Gene expression profile of patients with Mayer-Rokitansky-Küster-Hauser syndrome: New insights into the potential role of developmental pathways. PLoS ONE 2014, 9, e91010. [Google Scholar] [CrossRef]
- Lindblad, O.; Chougule, R.A.; Moharram, S.A.; Kabir, N.N.; Sun, J.; Kazi, J.U.; Ronnstrand, L. The role of HOXB2 and HOXB3 in acute myeloid leukemia. Biochem. Biophys. Res. Commun. 2015, 467, 742–747. [Google Scholar] [CrossRef] [Green Version]
- Boimel, P.J.; Cruz, C.; Segall, J.E. A functional in vivo screen for regulators of tumor progression identifies HOXB2 as a regulator of tumor growth in breast cancer. Genomics 2011, 98, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Berger, A.; Ivanova, E.; Gareau, C.; Scherrer, A.; Mazroui, R.; Strub, K. Direct binding of the Alu binding protein dimer SRP9/14 to 40S ribosomal subunits promotes stress granule formation and is regulated by Alu RNA. Nucleic Acids Res. 2014, 42, 11203–11217. [Google Scholar] [CrossRef]
- Donnelly, N.; Gorman, A.M.; Gupta, S.; Samali, A. The eIF2α kinases: Their structures and functions. Cell. Mol. Life Sci. 2013, 70, 3493–3511. [Google Scholar] [CrossRef]
- El-Naggar, A.M.; Sorensen, P.H. Translational control of aberrant stress responses as a hallmark of cancer. J. Pathol. 2018, 244, 650–666. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Keller, J.R.; Ortiz, M.; Tessarollo, L.; Rachel, R.A.; Nakamura, T.; Jenkins, N.A.; Copeland, N.G. Bcl11a is essential for normal lymphoid development. Nat. Immunol. 2003, 4, 525–532. [Google Scholar] [CrossRef]
- Khaled, W.T.; Choon Lee, S.; Stingl, J.; Chen, X.; Raza Ali, H.; Rueda, O.M.; Hadi, F.; Wang, J.; Yu, Y.; Chin, S.F.; et al. BCL11A is a triple-negative breast cancer gene with critical functions in stem and progenitor cells. Nat. Commun. 2015, 6, 5987. [Google Scholar] [CrossRef] [Green Version]
- Lazarus, K.A.; Hadi, F.; Zambon, E.; Bach, K.; Santolla, M.F.; Watson, J.K.; Correia, L.L.; Das, M.; Ugur, R.; Pensa, S.; et al. BCL11A interacts with SOX2 to control the expression of epigenetic regulators in lung squamous carcinoma. Nat. Commun. 2018, 9, 3327. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, S.-J.; Cho, H.J.; Kim, J.-T.; Yoon, H.R.; Lee, K.H.; Kim, B.Y.; Lee, Y.; Lee, H.G. Epsilon-globin HBE1 enhances radiotherapy resistance by down-regulating BCL11A in colorectal cancer cells. Cancers 2019, 11, 498. [Google Scholar] [CrossRef]
- Huang, H.T.; Chen, S.M.; Pan, L.B.; Yao, J.; Ma, H.T. Loss of function of SWI/SNF chromatin remodeling genes leads to genome instability of human lung cancer. Oncol. Rep. 2015, 33, 283–291. [Google Scholar] [CrossRef]
- Darda, L.; Hakami, F.; Morgan, R.; Murdoch, C.; Lambert, D.W.; Hunter, K.D. The role of HOXB9 and miR-196a in head and neck squamous cell carcinoma. PLoS ONE 2015, 10, e0122285. [Google Scholar] [CrossRef]
- Milevskiy, M.J.G.; Gujral, U.; Del Lama Marques, C.; Stone, A.; Northwood, K.; Burke, L.J.; Gee, J.M.W.; Nephew, K.; Clark, S.; Brown, M.A. MicroRNA-196a is regulated by ER and is a prognostic biomarker in ER+ breast cancer. Br. J. Cancer 2019, 120, 621–632. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Jin-Cheng, G.; Jue, Z.; Quan-Fu, M.; Bin, Y.; Xu-Feng, W. Protein-coding genes, long non-coding RNAs combined with microRNAs as a novel clinical multi-dimension transcriptome signature to predict prognosis in ovarian cancer. Oncotarget 2017, 8, 72847–72859. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Wang, J.; Zhao, L. The effect and mechanism of miR196a in HepG2 cell. Biomed. Pharmacother. 2015, 72, 1–5. [Google Scholar] [CrossRef]
- Xu, H.; Li, G.; Yue, Z.; Li, C. HCV core protein-induced upregulation of microRNA-196a promotes aberrant proliferation in hepatocellular carcinoma by targeting FOXO1. Mol. Med. Rep. 2016, 13, 5223–5229. [Google Scholar] [CrossRef]
- Chen, H.; Fan, Y.; Xu, W.; Chen, J.; Meng, Y.; Fang, D.; Wang, J. Exploration of miR-1202 and miR-196a in human endometrial cancer based on high throughout gene screening analysis. Oncol. Rep. 2017, 37, 3493–3501. [Google Scholar] [CrossRef] [Green Version]
- Nead, K.T.; Sharp, S.J.; Thompson, D.J.; Painter, J.N.; Savage, D.B.; Semple, R.K.; Barker, A.; Perry, J.R.; Attia, J.; Dunning, A.M.; et al. Evidence of a causal association between insulinemia and endometrial cancer: A mendelian randomization analysis. J. Natl. Cancer Inst. 2015, 107. [Google Scholar] [CrossRef]
- Painter, J.N.; O’Mara, T.A.; Marquart, L.; Webb, P.M.; Attia, J.; Medland, S.E.; Cheng, T.; Dennis, J.; Holliday, E.G.; McEvoy, M.; et al. Genetic risk score mendelian randomization shows that obesity measured as body mass index, but not waist:hip ratio, is causal for endometrial cancer. Cancer Epidemiol. Prev. Biomark. 2016, 25, 1503–1510. [Google Scholar] [CrossRef]
- Lotta, L.A.; Wittemans, L.B.L.; Zuber, V.; Stewart, I.D.; Sharp, S.J.; Luan, J.; Day, F.R.; Li, C.; Bowker, N.; Cai, L.; et al. Association of genetic variants related to gluteofemoral vs abdominal fat distribution with type 2 diabetes, coronary disease, and cardiovascular risk factors. Jama 2018, 320, 2553–2563. [Google Scholar] [CrossRef]
- Mori, M.; Nakagami, H.; Rodriguez-Araujo, G.; Nimura, K.; Kaneda, Y. Essential role for miR-196a in brown adipogenesis of white fat progenitor cells. PLoS Biol. 2012, 10, e1001314. [Google Scholar] [CrossRef]
- Hewitt, S.C.; Deroo, B.J.; Hansen, K.; Collins, J.; Grissom, S.; Afshari, C.A.; Korach, K.S. Estrogen receptor-dependent genomic responses in the uterus mirror the biphasic physiological response to estrogen. Mol. Endocrinol. 2003, 17, 2070–2083. [Google Scholar] [CrossRef]
- Gründker, C.; Günthert, A.R.; Emons, G. Hormonal heterogeneity of endometrial cancer. In Innovative Endocrinology of Cancer; Berstein, L.M., Santen, R.J., Eds.; Springer: New York, NY, USA, 2008; pp. 166–188. [Google Scholar] [CrossRef]
- Belaghzal, H.; Dekker, J.; Gibcus, J.H. Hi-C 2.0: An optimized Hi-C procedure for high-resolution genome-wide mapping of chromosome conformation. Methods 2017, 123, 56–65. [Google Scholar] [CrossRef]
- Servant, N.; Varoquaux, N.; Lajoie, B.R.; Viara, E.; Chen, C.-J.; Vert, J.-P.; Heard, E.; Dekker, J.; Barillot, E. HiC-Pro: An optimized and flexible pipeline for Hi-C data processing. Genome Biol. 2015, 16, 259. [Google Scholar] [CrossRef]
- Lareau, C.A.; Aryee, M.J. hichipper: A preprocessing pipeline for calling DNA loops from HiChIP data. Nat. Methods 2018, 15, 155–156. [Google Scholar] [CrossRef]
- Phanstiel, D.H.; Boyle, A.P.; Heidari, N.; Snyder, M.P. Mango: A bias-correcting ChIA-PET analysis pipeline. Bioinformatics 2015, 31, 3092–3098. [Google Scholar] [CrossRef]
- Bulik-Sullivan, B.K.; Loh, P.R.; Finucane, H.K.; Ripke, S.; Yang, J.; Patterson, N.; Daly, M.J.; Price, A.L.; Neale, B.M. LD Score regression distinguishes confounding from polygenicity in genome-wide association studies. Nat. Genet. 2015, 47, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Finucane, H.K.; Bulik-Sullivan, B.; Gusev, A.; Trynka, G.; Reshef, Y.; Loh, P.R.; Anttila, V.; Xu, H.; Zang, C.; Farh, K.; et al. Partitioning heritability by functional annotation using genome-wide association summary statistics. Nat. Genet. 2015, 47, 1228–1235. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef]


{kind=link}
| Cell Line | Total Loops | Loops < 20 kb | Loops > 20 kb | Promoter-Asociated Loops | Median Span of Promoter-Associated Loops (kb) |
|---|---|---|---|---|---|
| E6E7hTERT | 162,476 | 25,133 (15.5%) | 137,343 (84.5%) | 59,658 (36.7%) | 206 |
| ARK1 | 449,157 | 45,932 (10.2%) | 403,225 (89.8%) | 155,080 (34.5%) | 282 |
| Ishikawa | 219,067 | 29,954 (13.7%) | 189,113 (86.3%) | 79,309 (36.2%) | 259 |
| JHUEM-14 | 66,092 | 10,254 (15.5%) | 55,838 (84.5%) | 26,492 (40.0%) | 209 |
| Cell Line | Enrichment (Standard Error) | p-Value |
|---|---|---|
| E6E7hTERT | 6.92 (1.70) | 1.30 × 10−04 |
| ARK1 | 4.08 (0.84) | 2.50 × 10−04 |
| JHUEM-14 | 9.61 (3.11) | 5.00 × 10−03 |
| Ishikawa | 3.23 (1.18) | 0.07 |
| Risk Locus | HiChIP Target Genes | Nearest Gene(s) to CVs 1 |
|---|---|---|
| 1p34.3 | GNL2, C1orf122 | GNL2, RSPO1 |
| 2p16.1 | BCL11A | BCL11A |
| 8q24.1 | MIR1207, PVT1, LINC00824 | LINC00824 |
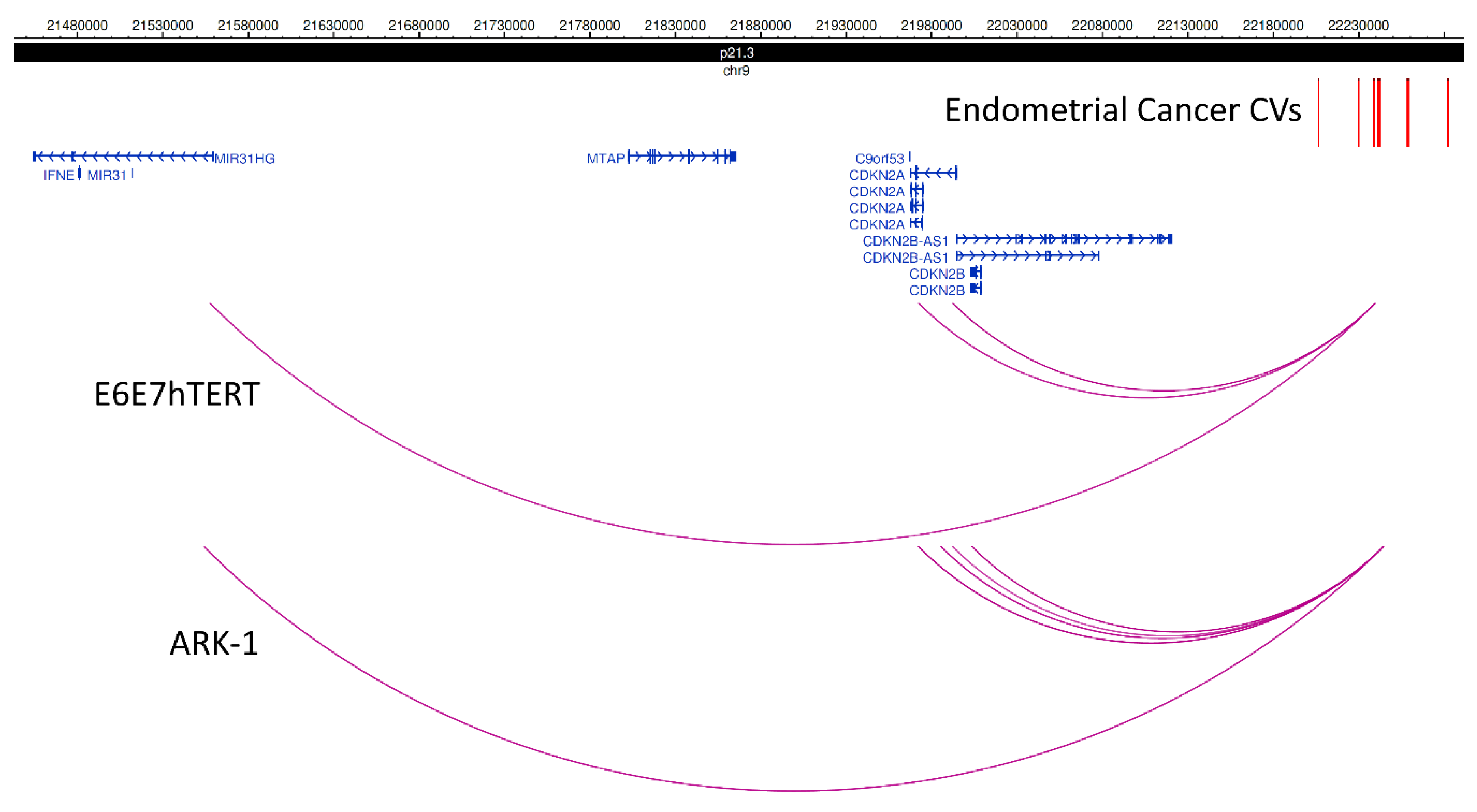
| 9p21.3 | CDKN2A, CDKN2B, CDKN2B-AS1, MIR31HG | CDKN2B-AS1 |
| 11p13 | WT1, WT1-AS, CD59, PAX6, RCN1 | WT1-AS |
| 12p12.1 | BHLHE41, PTHLH, SSPN, LRMP | SSPN |
| 12q24.11 | SH2B3, PHETA1, ACAD10, ARPC3, BRAP, IFT81, LINC02356 | SH2B3, ATXN2 |
| 12q24.21 | TBX3 | TBX3 |
| 15q15.1 | SRP14, SRP14-AS1, BMF, BAHD1, CCDC9B, GPR176, KNSTRN, PAK6, PLCB2, PLCB2-AS1, THBS1, EIF2AK4, CHST14, DISP2, FSIP1, INAFM2, PLA2G4B, RASGRP1, SPINT1, ANKRD63, PHGR1, SPINT1-AS1, C15orf56 | SRP14, SRP14-AS1, EIF2AK4 |
| 15q21.2 | DMXL2, TRPM7, TNFAIP8L3 | CYP19A1 |
| 17q11.2 | RAB11FIP4, MIR193A, TEFM, RNU6ATAC7P | RAB11FIP4, NF1, EVI2A, EVI2B |
| 17q12 | HNF1B, DUSP14, MRM1, MRPL45, SRCIN1, TBC1D3, C17orf78 | HNF1B |
| 17q21.32 | SNX11, MIR1203, SKAP1-AS1, SKAP1, CBX1, HOXB1, HOXB2, HOXB3, HOXB4, HOXB5, HOXB6, HOXB7, HOXB8, HOXB9, HOXB13, HOXB-AS1, HOXB-AS3, HOXB-AS4, PRR15L, CDK5RAP3, LRRC46, MRPL10, NFE2L1, SCRN2, CALCOCO2, COPZ2, DLX3, KPNB1, PNPO, SNF8, SP2, SP2-AS1, SP6, MIR10A, MIR152, MIR196A1, MIR3185, PHOSPHO1 | SNX11, MIR1203, SKAP1-AS1, SKAP1, CBX1 |
| Protein Encoding Gene | Similarity Score | p-Value | FDR 1 Value |
|---|---|---|---|
| TP53 | 0.60 | 3.65E−09 | 4.00E−06 |
| ESR1 | 0.54 | 5.95E−07 | 1.27E−04 |
| FOXA2 | 0.57 | 1.86E−06 | 1.99E−04 |
| EP300 | 0.41 | 8.40E−06 | 4.27E−04 |
| CTNNB1 | 0.47 | 1.35E−05 | 5.54E−04 |
| PTEN | 0.46 | 1.98E−05 | 7.05E−04 |
| CCND1 | 0.49 | 2.12E−05 | 7.42E−04 |
| FGFR2 | 0.44 | 3.97E−05 | 1.10E−03 |
| RB1 | 0.50 | 8.21E−05 | 1.91E−03 |
| MYCN | 0.44 | 1.15E−04 | 2.51E−03 |
| ERBB2 | 0.39 | 4.15E−04 | 6.28E−03 |
| AKT1 | 0.35 | 5.24E−04 | 7.31E−03 |
| ERBB3 | 0.39 | 1.12E−03 | 0.01 |
| MAX | 0.31 | 1.75E−03 | 0.02 |
| NRIP1 | 0.32 | 1.82E−03 | 0.02 |
| ATM | 0.31 | 2.05E−03 | 0.02 |
| CHD4 | 0.34 | 2.67E−03 | 0.02 |
| FBXW7 | 0.38 | 3.75E−03 | 0.03 |
| DICER1 | 0.33 | 4.44E−03 | 0.03 |
| KRAS | 0.33 | 9.91E−03 | 0.05 |
| TAF1 | 0.27 | 0.03 | 0.11 |
| ATR | 0.29 | 0.04 | 0.13 |
| PIK3R2 | 0.19 | 0.06 | 0.17 |
| POLE | 0.26 | 0.07 | 0.17 |
| CHD3 | 0.20 | 0.14 | 0.26 |
| TAB3 | 0.22 | 0.36 | 0.45 |
| METTL14 | 0.21 | 0.40 | 0.49 |
| KANSL1 | 0.09 | 0.67 | 0.67 |
| Cancer Hallmark | Related Pathway (Source) | pBonferroni |
|---|---|---|
| Evading growth suppressors | Regulation of TP53 activity (REACTOME) | 1.44E−07 |
| Avoiding immune destruction | Innate immune system (REACTOME) | 2.06E−06 |
| Enabling replicative immortality | Regulation of telomerase (Pathway Interaction Database) | 1.43E−12 |
| Tumor-promoting inflammation | Inflammation mediated by chemokine and cytokine signalling pathway (PantherDB) | 0.03 |
| Activating invasion and metastasis | Focal adhesion (KEGG) | 5.61E−15 |
| Inducing angiogenesis | VEGFA-VEGFR2 pathway (REACTOME) | 9.10E−08 |
| Genome instability and mutation | RB Tumor Suppressor/Checkpoint Signaling in response to DNA damage (MSigDB C2 BIOCARTA) | 1.05E−04 |
| Resisting cell death | Apoptosis signaling pathway (Panther DB) | 1.78E−08 |
| Deregulating cellular energetics | Choline metabolism in cancer (KEGG) | 2.10E−04 |
| Sustaining proliferative signalling | PI3K-Akt signalling pathway (KEGG) | 1.15E−18 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Mara, T.A.; Spurdle, A.B.; Glubb, D.M.; Endometrial Cancer Association Consortium. Analysis of Promoter-Associated Chromatin Interactions Reveals Biologically Relevant Candidate Target Genes at Endometrial Cancer Risk Loci. Cancers 2019, 11, 1440. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101440
O’Mara TA, Spurdle AB, Glubb DM, Endometrial Cancer Association Consortium. Analysis of Promoter-Associated Chromatin Interactions Reveals Biologically Relevant Candidate Target Genes at Endometrial Cancer Risk Loci. Cancers. 2019; 11(10):1440. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101440
Chicago/Turabian StyleO’Mara, Tracy A., Amanda B. Spurdle, Dylan M. Glubb, and Endometrial Cancer Association Consortium. 2019. "Analysis of Promoter-Associated Chromatin Interactions Reveals Biologically Relevant Candidate Target Genes at Endometrial Cancer Risk Loci" Cancers 11, no. 10: 1440. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101440