Pan Aurora Kinase Inhibitor: A Promising Targeted-Therapy in Dedifferentiated Liposarcomas With Differential Efficiency Depending on Sarcoma Molecular Profile

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
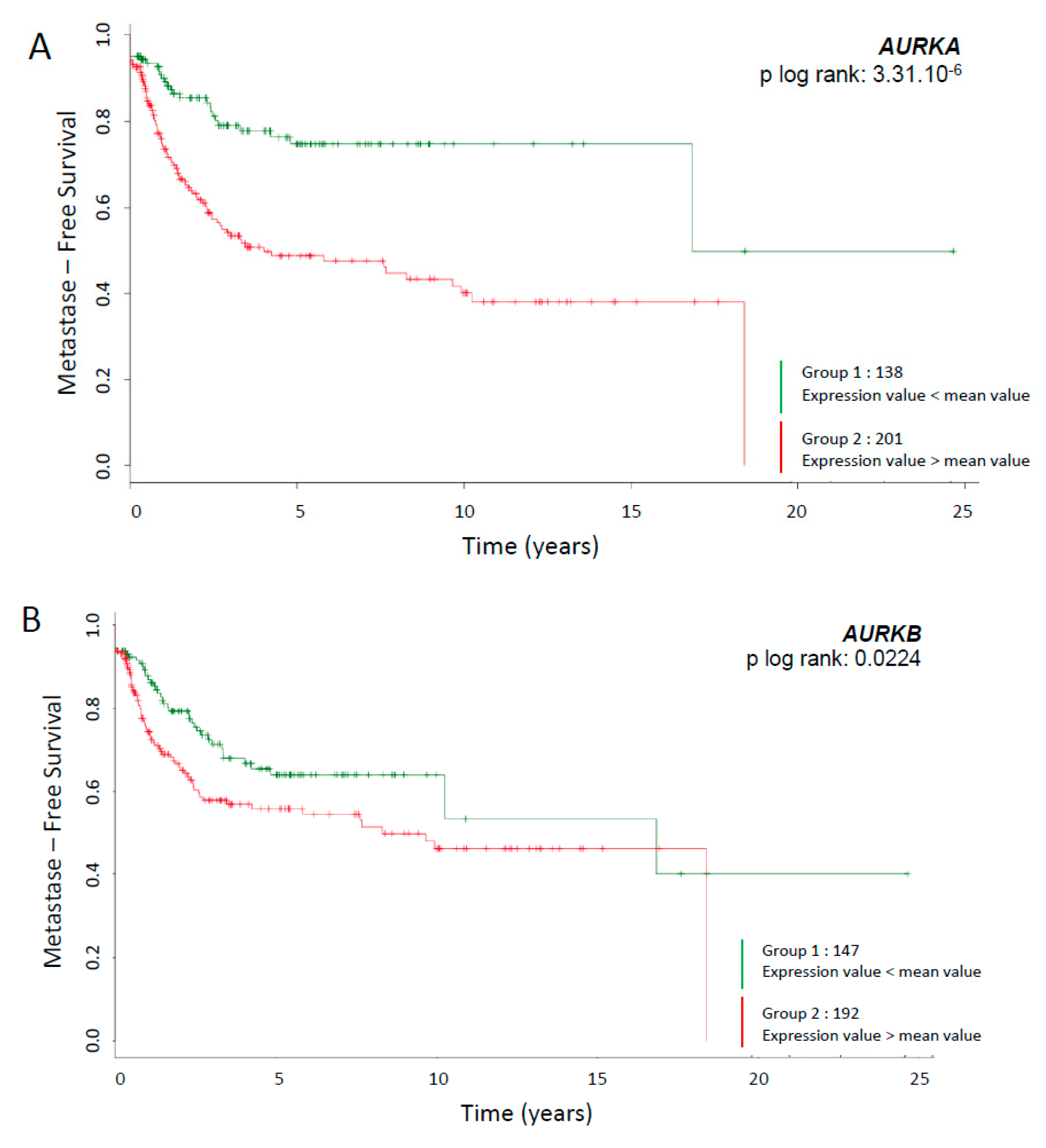
2.1. Survival Curves Specific to AURKA and AURKB mRNA Expression
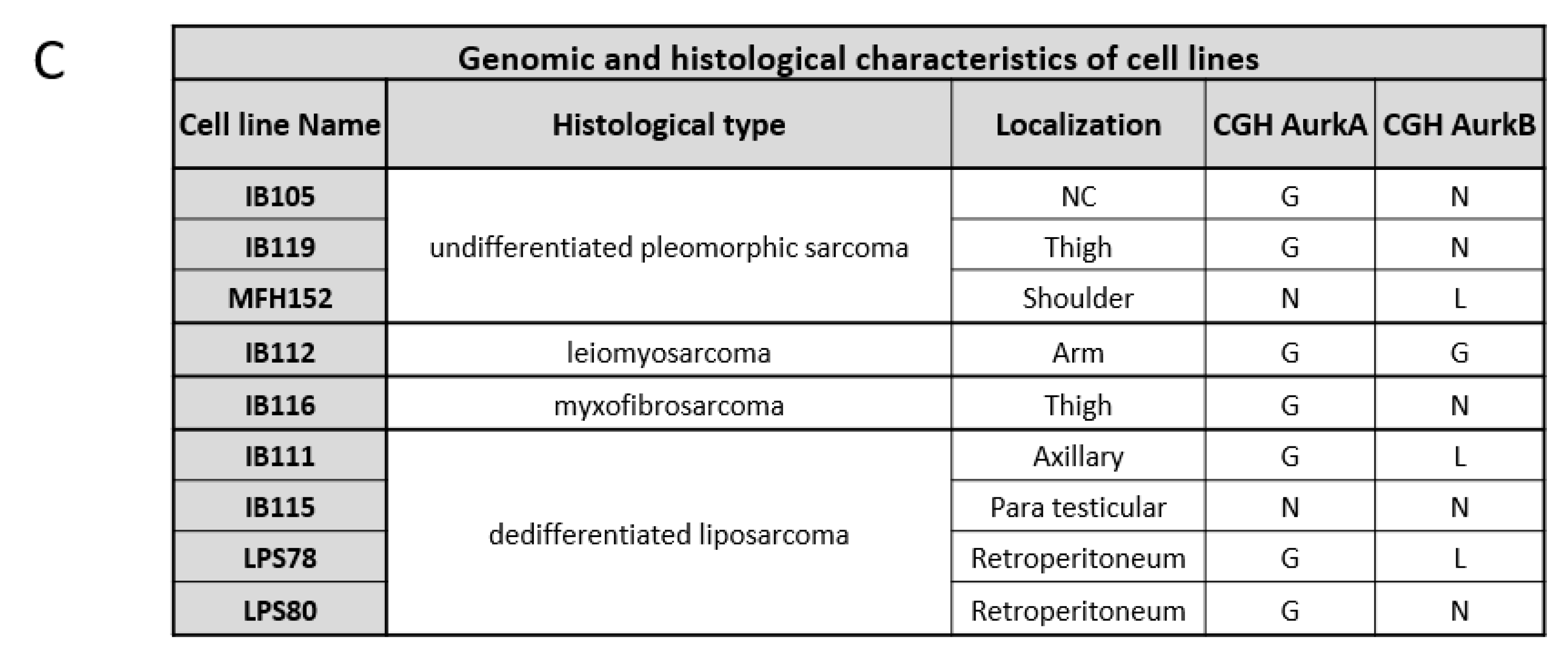
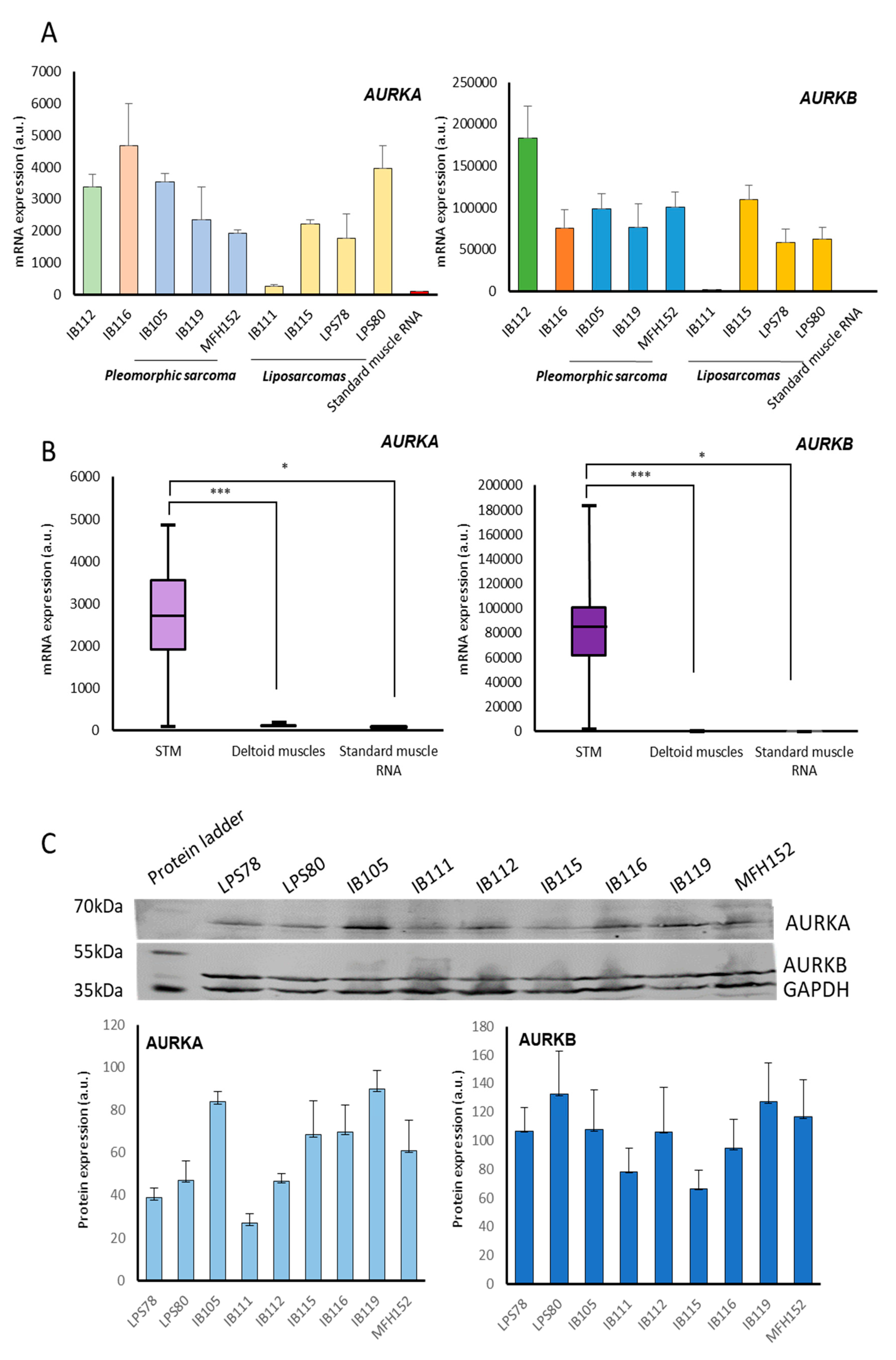
2.2. Aurora Kinase Expression in Soft Tissue Sarcomas Cell Lines
2.2.1. CGH and Gene Expression
2.2.2. Protein Expression
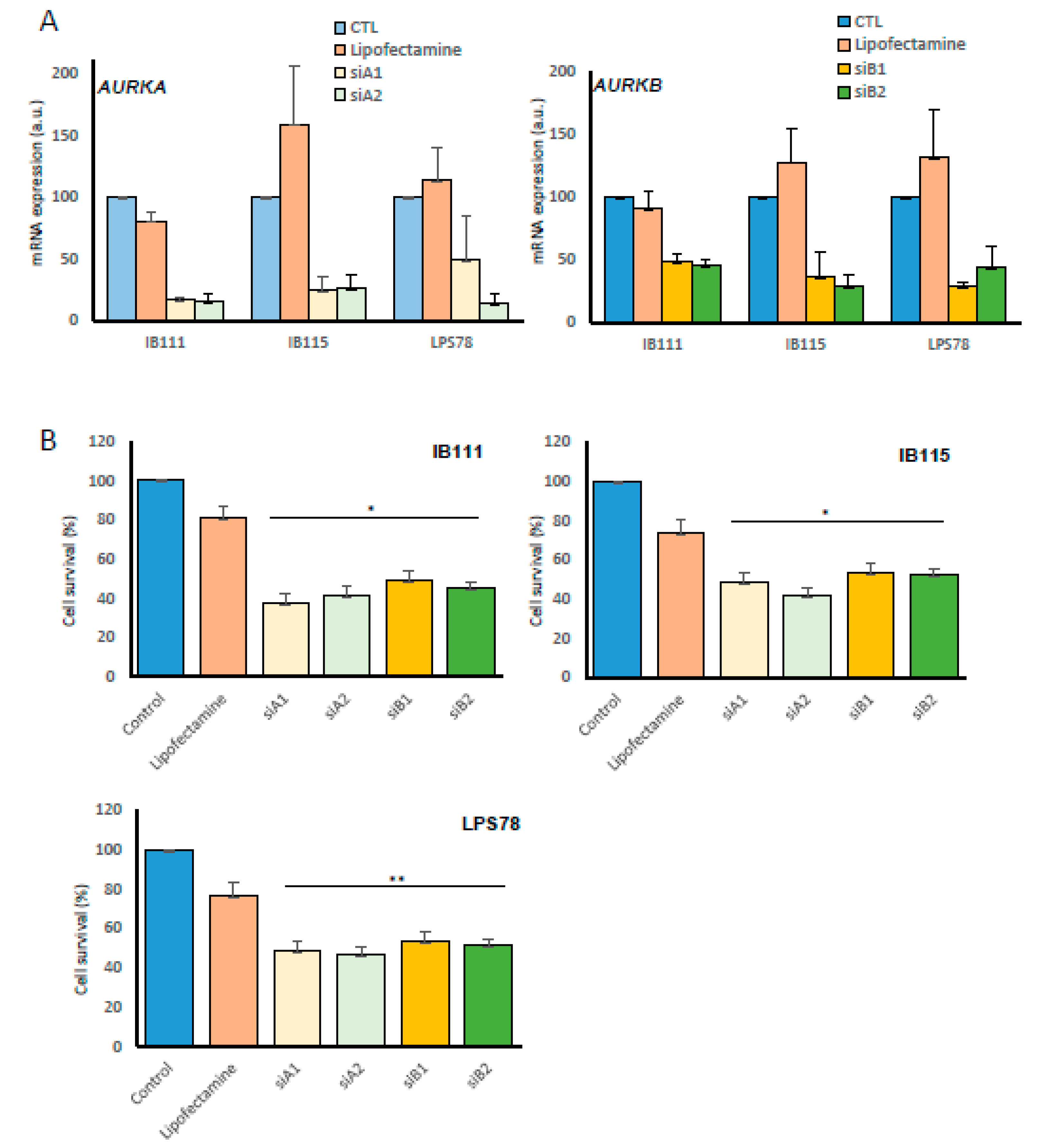
2.3. Impact of Inhibition of Aurora Kinases A and B RNAs on Liposarcoma Cell Survival
2.4. Characterisation of AMG 900 Impact on LPS
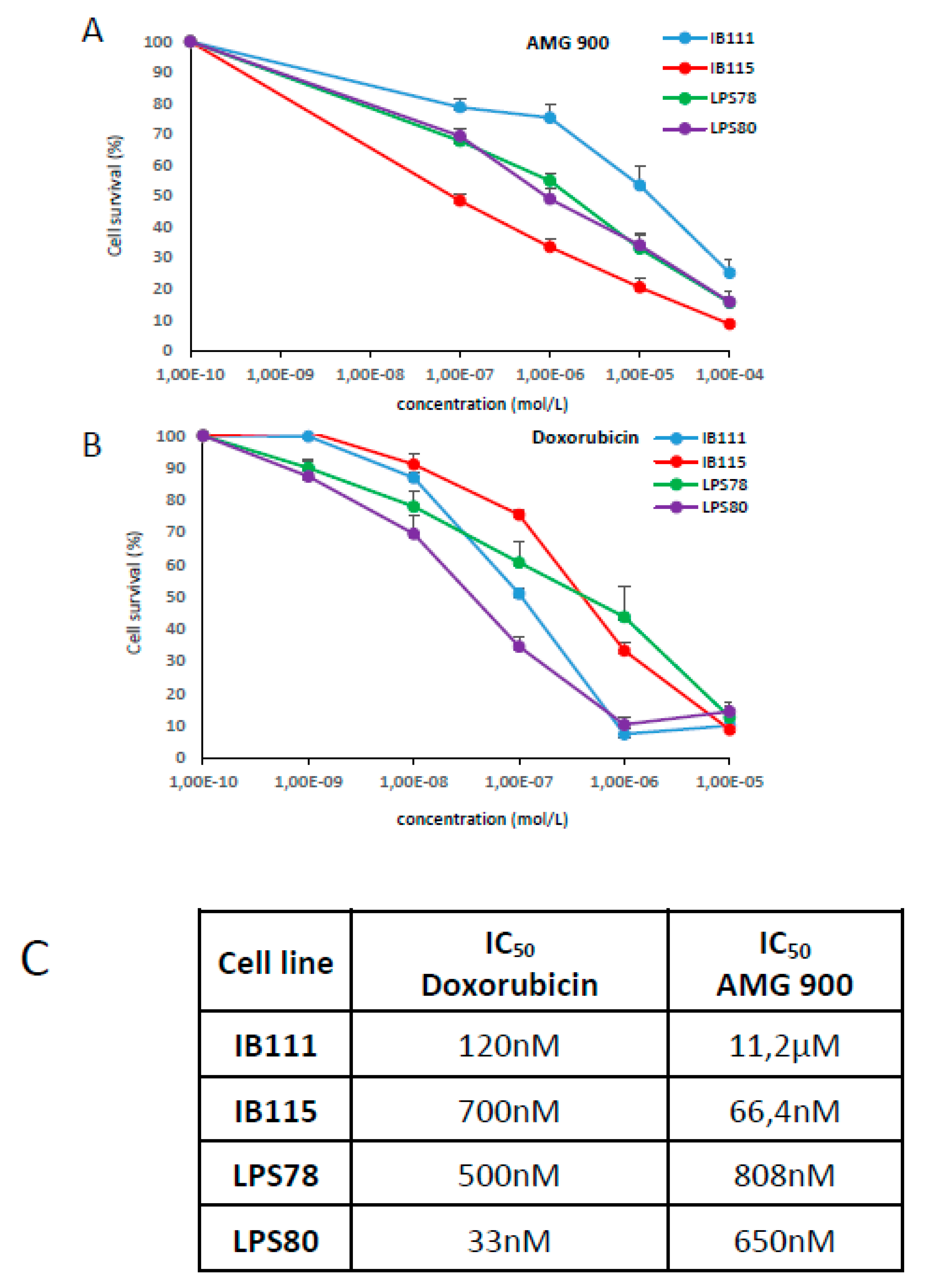
2.4.1. Cytotocic Effect of AMG 900 and Doxorubicin on Sarcoma Cell Lines
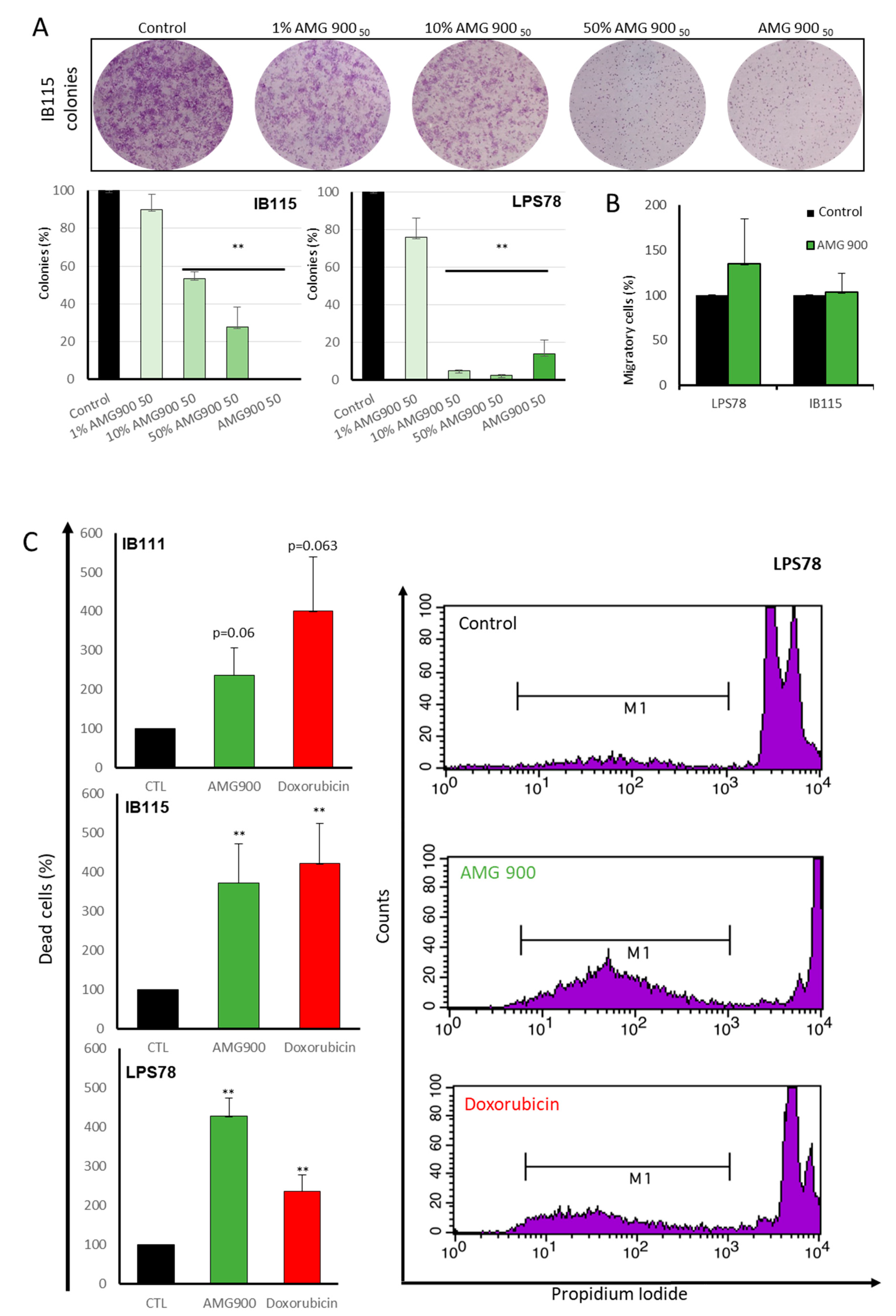
2.4.2. Tumorigenicity
2.4.3. DNA Fragmentation is Induced by AMG 900, Doxorubicin and AURKs siRNA
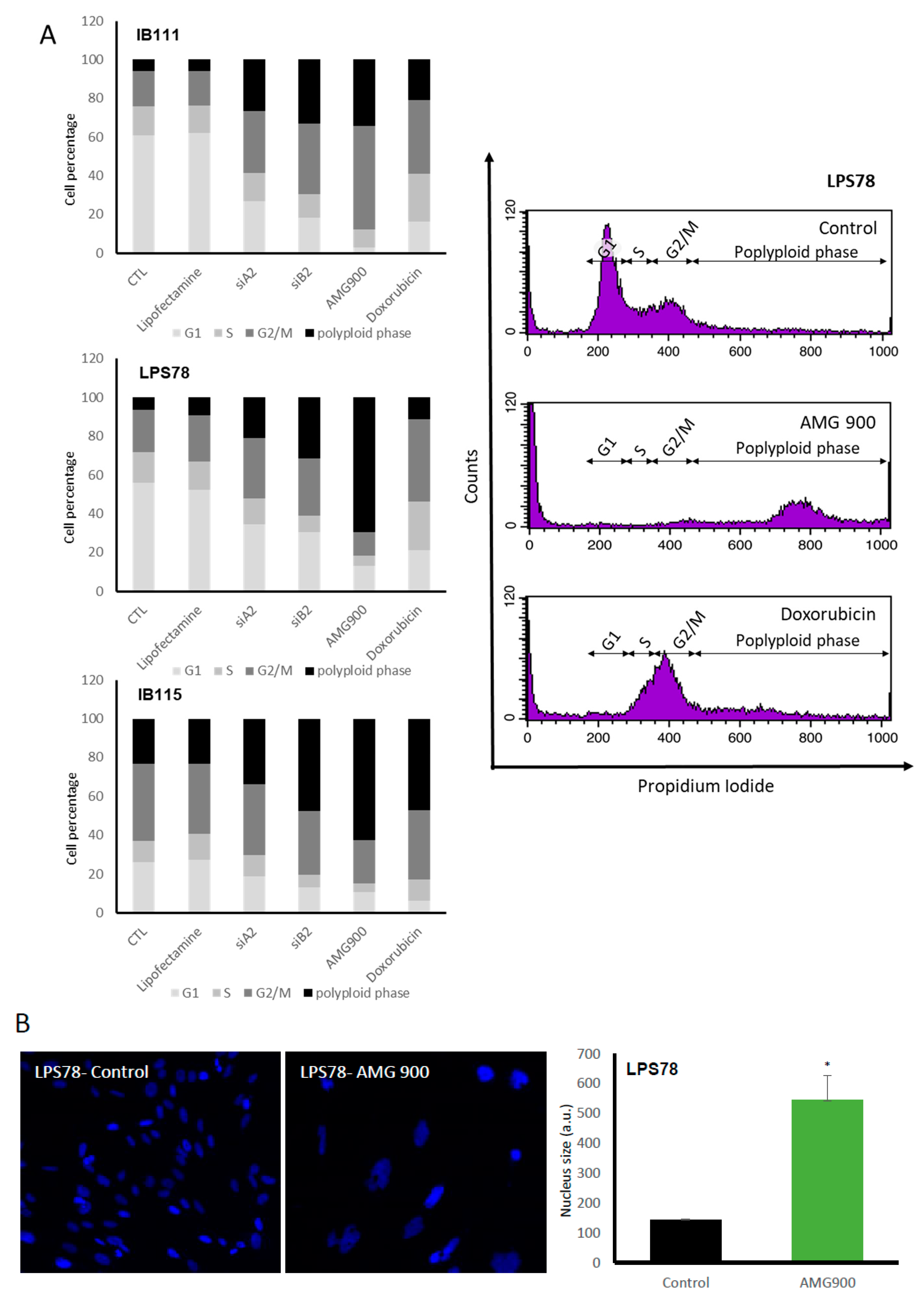
2.4.4. Cell Cycle is Differentially Modulated by AMG 900 in Different Liposarcoma Cell Lines
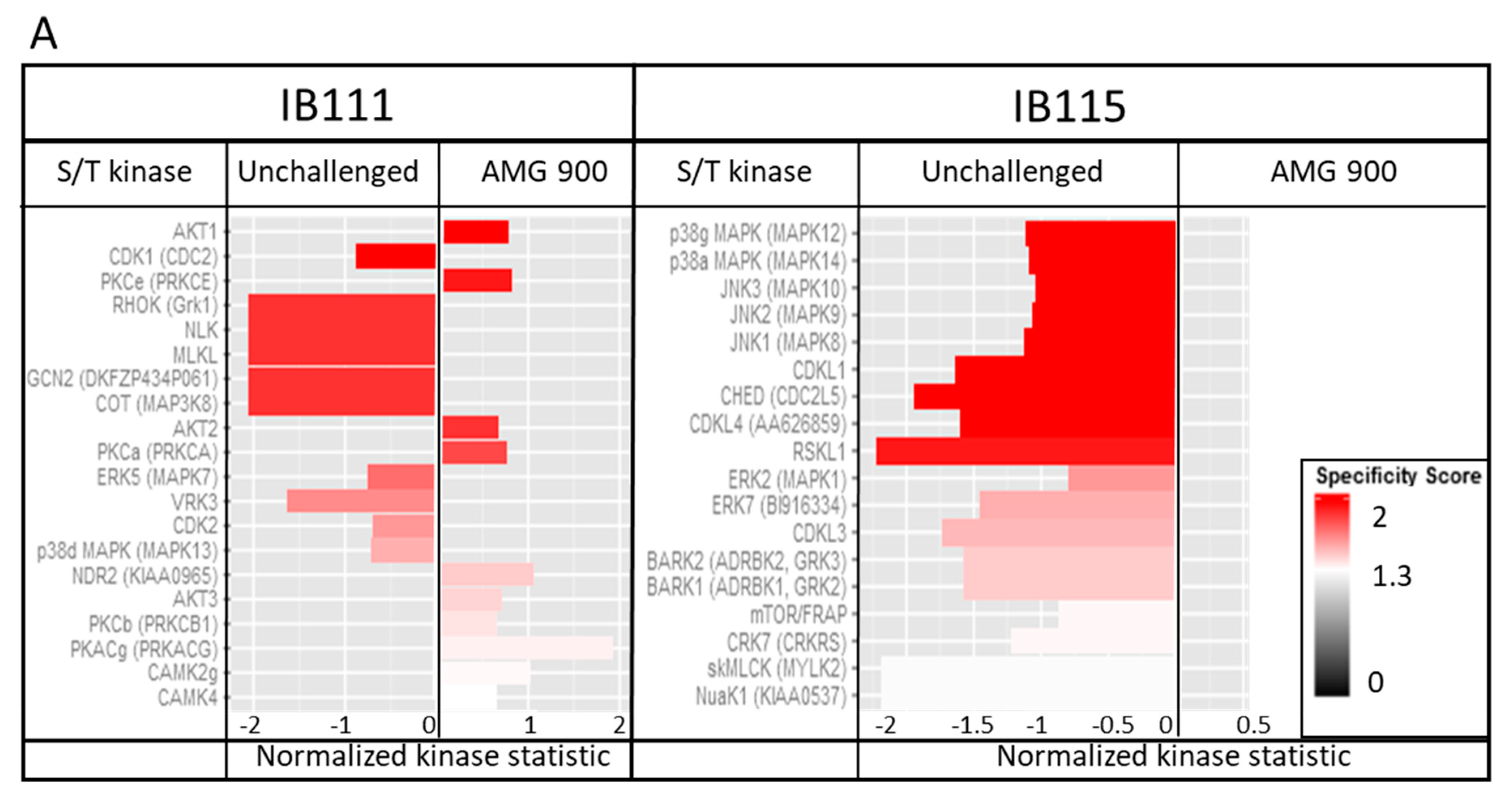
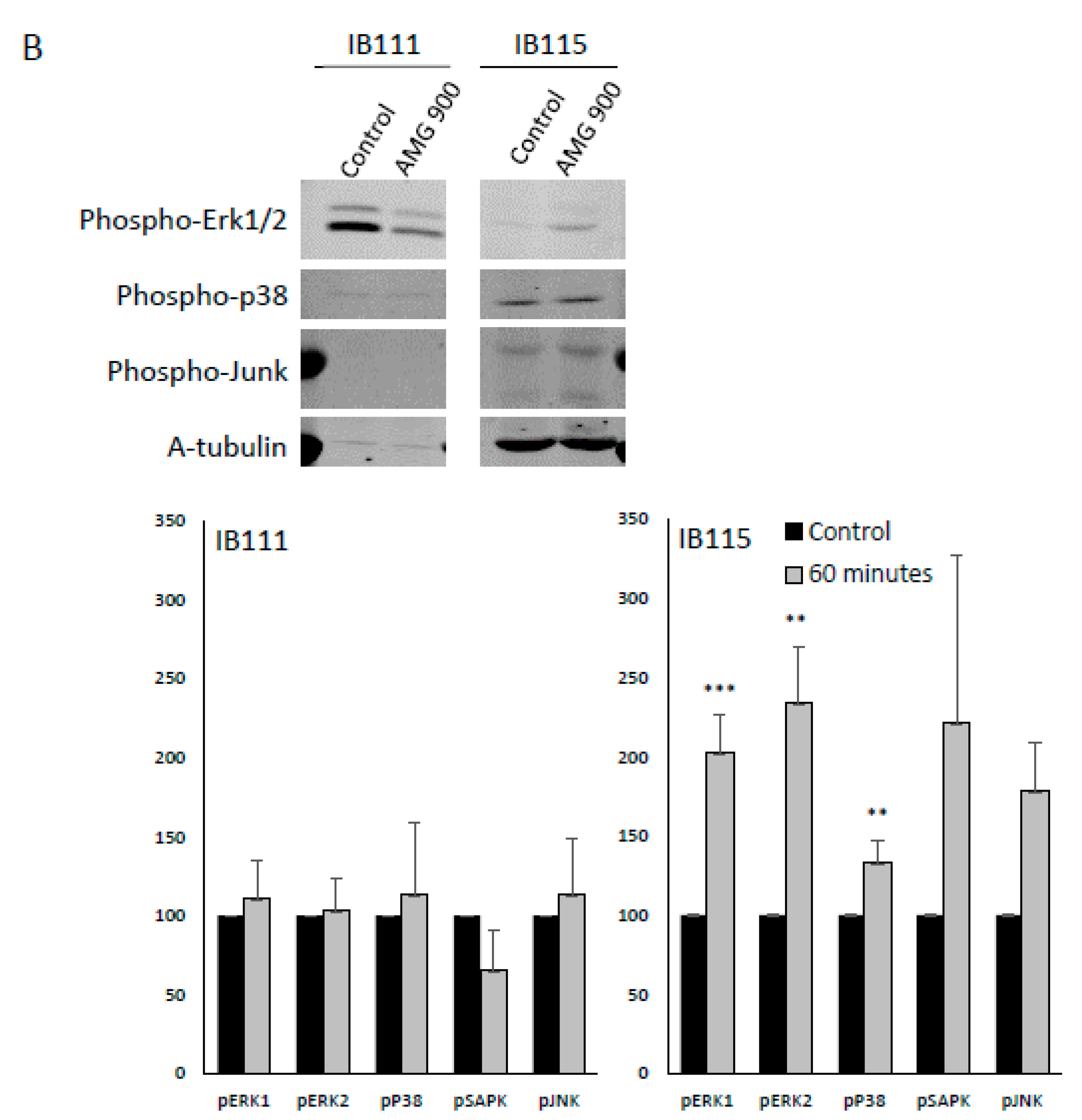
2.4.5. Kinomic Analysis: AMG 900 Action is Linked to MAPK Kinase Modulation
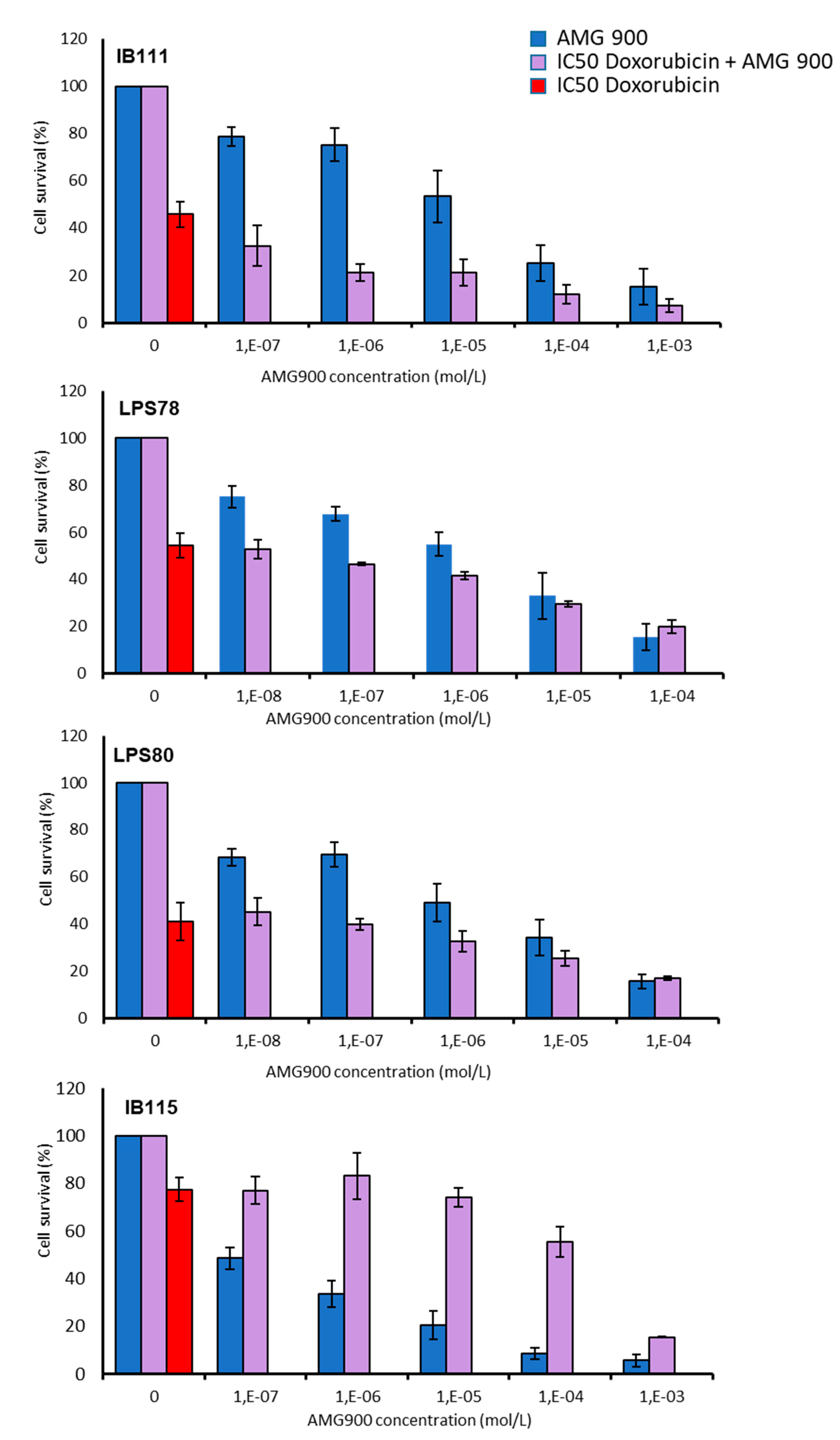
2.5. Combination of AMG 900 and Doxorubicin Treatments
3. Discussion
4. Methods
4.1. Subject and Samples
4.2. DNA Extraction, Array Comparative Genomic Hybridization Analysis and TCGA Database
4.3. STS cell Lines Establishment and Culture
4.4. MTT Assay
4.5. q-RT PCR
4.6. Western Blot
4.7. Clonogenic Assay
4.8. Migration Assay
4.9. Proliferation Assay
4.10. Cytometry Analysis
4.11. Kinome Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| STS | Soft tissue sarcoma |
| LPS | Liposarcoma |
| AURK | Aurora Kinase |
| DDLPS | dedifferentiated Liposarcoma |
References
- Bui, B.-N.; Blay, J.-Y.; Bonichon, F.; Bonvalot, S.; Chevalier-Place, A.; Coindre, J.-M.; Delannes, M.; Le Cesne, A.; Morice, P.; Ray-Coquard, I. Standards, Options et Recommandations 2006. Prise en charge des patients adultes atteints de sarcome des tissus mous, de sarcome utérin ou de tumeur stromale gastro-intestinale. Oncologie 2007, 9, 173–177. [Google Scholar]
- Les cancers en France-Edition 2014. Available online: http://www.senologie.com/wp-content/uploads/2015/02/Les-cancers-en-france-Edition-2014-Faits-et-chiffres-V8-1.pdf. (accessed on 3 March 2020).
- Cancer Facts and Statistics 2019. Available online: http://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/annual-cancer-facts-and-figures/2019/cancer-facts-and-figures-2019.pdf. (accessed on 3 March 2020).
- Jo, V.Y.; Fletcher, C.D.M. WHO classification of soft tissue tumours: An update based on the 2013 (4th) edition. Pathology (Phila.) 2014, 46, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Ray-Coquard, I.; Thiesse, P.; Ranchère-Vince, D.; Chauvin, F.; Bobin, J.-Y.; Sunyach, M.-P.; Carret, J.-P.; Mongodin, B.; Marec-Bérard, P.; Philip, T.; et al. Conformity to clinical practice guidelines, multidisciplinary management and outcome of treatment for soft tissue sarcomas. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. ESMO 2004, 15, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Giannakakou, P. Targeting microtubules for cancer chemotherapy. Curr. Med. Chem. Anti-Cancer Agents 2005, 5, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Morris, P.G.; Fornier, M.N. Microtubule active agents: Beyond the taxane frontier. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 7167–7172. [Google Scholar] [CrossRef] [Green Version]
- Keen, N.; Taylor, S. Aurora-kinase inhibitors as anticancer agents. Nat. Rev. Cancer 2004, 4, 927–936. [Google Scholar] [CrossRef]
- Warner, S.L.; Stephens, B.J.; Von Hoff, D.D. Tubulin-associated proteins: Aurora and Polo-like kinases as therapeutic targets in cancer. Curr. Oncol. Rep. 2008, 10, 122–129. [Google Scholar] [CrossRef]
- Sharma, P.S.; Sharma, R.; Tyagi, R. Inhibitors of cyclin dependent kinases: Useful targets for cancer treatment. Curr. Cancer Drug Targets 2008, 8, 53–75. [Google Scholar] [CrossRef]
- Chibon, F.; Lagarde, P.; Salas, S.; Pérot, G.; Brouste, V.; Tirode, F.; Lucchesi, C.; de Reynies, A.; Kauffmann, A.; Bui, B.; et al. Validated prediction of clinical outcome in sarcomas and multiple types of cancer on the basis of a gene expression signature related to genome complexity. Nat. Med. 2010, 16, 781–787. [Google Scholar] [CrossRef]
- Coindre, J.M.; Terrier, P.; Guillou, L.; Le Doussal, V.; Collin, F.; Ranchère, D.; Sastre, X.; Vilain, M.O.; Bonichon, F.; N’Guyen Bui, B. Predictive value of grade for metastasis development in the main histologic types of adult soft tissue sarcomas: A study of 1240 patients from the French Federation of Cancer Centers Sarcoma Group. Cancer 2001, 91, 1914–1926. [Google Scholar] [CrossRef]
- Idbaih, A.; Coindre, J.-M.; Derré, J.; Mariani, O.; Terrier, P.; Ranchère, D.; Mairal, A.; Aurias, A. Myxoid malignant fibrous histiocytoma and pleomorphic liposarcoma share very similar genomic imbalances. Lab. Investig. J. Tech. Methods Pathol. 2005, 85, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Andrews, P.D.; Knatko, E.; Moore, W.J.; Swedlow, J.R. Mitotic mechanics: The auroras come into view. Curr. Opin. Cell Biol. 2003, 15, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Penrhyn-Lowe, S.; Venkitaraman, A.R. AURORA-A amplification overrides the mitotic spindle assembly checkpoint, inducing resistance to Taxol. Cancer Cell 2003, 3, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Ewart-Toland, A.; Briassouli, P.; de Koning, J.P.; Mao, J.-H.; Yuan, J.; Chan, F.; MacCarthy-Morrogh, L.; Ponder, B.A.J.; Nagase, H.; Burn, J.; et al. Identification of Stk6/STK15 as a candidate low-penetrance tumor-susceptibility gene in mouse and human. Nat. Genet. 2003, 34, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Kufer, T.A.; Silljé, H.H.W.; Körner, R.; Gruss, O.J.; Meraldi, P.; Nigg, E.A. Human TPX2 is required for targeting Aurora-A kinase to the spindle. J. Cell Biol. 2002, 158, 617–623. [Google Scholar] [CrossRef] [Green Version]
- Payton, M.; Bush, T.L.; Chung, G.; Ziegler, B.; Eden, P.; McElroy, P.; Ross, S.; Cee, V.J.; Deak, H.L.; Hodous, B.L.; et al. Preclinical Evaluation of AMG 900, a Novel Potent and Highly Selective Pan-Aurora Kinase Inhibitor with Activity in Taxane-Resistant Tumor Cell Lines. Cancer Res. 2010, 70, 9846–9854. [Google Scholar] [CrossRef] [Green Version]
- Dickson, M.A.; Mahoney, M.R.; Tap, W.D.; D’Angelo, S.P.; Keohan, M.L.; Tine, B.A.V.; Agulnik, M.; Horvath, L.E.; Nair, J.S.; Schwartz, G.K. Phase II study of MLN8237 (Alisertib) in advanced/metastatic sarcoma. Ann. Oncol. 2016, 27, 1855–1860. [Google Scholar] [CrossRef]
- Pitts, T.M.; Bradshaw-Pierce, E.L.; Bagby, S.M.; Hyatt, S.L.; Selby, H.M.; Spreafico, A.; Tentler, J.J.; McPhillips, K.; Klauck, P.J.; Capasso, A.; et al. Antitumor activity of the aurora a selective kinase inhibitor, alisertib, against preclinical models of colorectal cancer. Oncotarget 2016, 7, 50290–50301. [Google Scholar] [CrossRef] [Green Version]
- Nair, J.S.; Schwartz, G.K. MLN-8237: A dual inhibitor of aurora A and B in soft tissue sarcomas. Oncotarget 2016, 7, 12893–12903. [Google Scholar] [CrossRef] [Green Version]
- Burgess, A.; Chia, K.M.; Haupt, S.; Thomas, D.; Haupt, Y.; Lim, E. Clinical Overview of MDM2/X-Targeted Therapies. Front. Oncol. 2016, 6. [Google Scholar] [CrossRef]
- Luke, J.J.; D’Adamo, D.R.; Dickson, M.A.; Keohan, M.L.; Carvajal, R.D.; Maki, R.G.; de Stanchina, E.; Musi, E.; Singer, S.; Schwartz, G.K. The cyclin-dependent kinase inhibitor flavopiridol potentiates doxorubicin efficacy in advanced sarcomas: Preclinical investigations and results of a phase I dose escalation clinical trial. Clin. Cancer Res. 2012, 18, 2638–2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, X.; Li, C.; He, C.; Ren, B.; Deng, Q.; Gao, W.; Wang, B. Aurora-A modulates MMP-2 expression via AKT/NF-κB pathway in esophageal squamous cell carcinoma cells. Acta Biochim. Biophys. Sin. 2016, 48, 520–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, X.-B.; Long, Z.-J.; Yan, M.; Xu, J.; Xia, L.-P.; Liu, L.; Zhao, Y.; Huang, X.-F.; Wang, X.-R.; Zhu, X.-F.; et al. Inhibition of Aurora-A suppresses epithelial–mesenchymal transition and invasion by downregulating MAPK in nasopharyngeal carcinoma cells. Carcinogenesis 2008, 29, 1930–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giet, R.; Petretti, C.; Prigent, C. Aurora kinases, aneuploidy and cancer, a coincidence or a real link? Trends Cell Biol. 2005, 15, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Kalous, O.; Conklin, D.; Desai, A.J.; Dering, J.; Goldstein, J.; Ginther, C.; Anderson, L.; Lu, M.; Kolarova, T.; Eckardt, M.A.; et al. AMG 900, pan-Aurora kinase inhibitor, preferentially inhibits the proliferation of breast cancer cell lines with dysfunctional p53. Breast Cancer Res. Treat. 2013, 141, 397–408. [Google Scholar] [CrossRef]
- Ki67, PCNA, and MCM proteins: Markers of proliferation in the diagnosis of breast cancer. Available online: http://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0065128116300848 (accessed on 10 November 2016).
- Mahankali, M.; Henkels, K.M.; Speranza, F.; Gomez-Cambronero, J. A non-mitotic role for Aurora kinase A as a direct activator of cell migration upon interaction with PLD, FAK and Src. J. Cell Sci. 2015, 128, 516–526. [Google Scholar] [CrossRef]
- Do, T.-V.; Xiao, F.; Bickel, L.E.; Klein-Szanto, A.J.; Pathak, H.B.; Hua, X.; Howe, C.; O’Brien, S.W.; Maglaty, M.; Ecsedy, J.A.; et al. Aurora kinase A mediates epithelial ovarian cancer cell migration and adhesion. Oncogene 2014, 33, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Lüpertz, R.; Wätjen, W.; Kahl, R.; Chovolou, Y. Dose- and time-dependent effects of doxorubicin on cytotoxicity, cell cycle and apoptotic cell death in human colon cancer cells. Toxicology 2010, 271, 115–121. [Google Scholar] [CrossRef]
- Ling, Y.H.; el-Naggar, A.K.; Priebe, W.; Perez-Soler, R. Cell cycle-dependent cytotoxicity, G2/M phase arrest, and disruption of p34cdc2/cyclin B1 activity induced by doxorubicin in synchronized P388 cells. Mol. Pharmacol. 1996, 49, 832–841. [Google Scholar]
- Pouysségur, J.; Volmat, V.; Lenormand, P. Fidelity and spatio-temporal control in MAP kinase (ERKs) signalling. Biochem. Pharmacol. 2002, 64, 755–763. [Google Scholar] [CrossRef]
- Cagnol, S.; Chambard, J.-C. ERK and cell death: Mechanisms of ERK-induced cell death – apoptosis, autophagy and senescence. FEBS J. 2010, 277, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Xu, S. ERK1/2 MAP kinases in cell survival and apoptosis. IUBMB Life 2006, 58, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. MMBR 2011, 75, 50–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, K.; Rainey, M.A.; Dalby, K.N. Quantifying ERK2–protein interactions by fluorescence anisotropy: PEA-15 inhibits ERK2 by blocking the binding of DEJL domains. Biochim. Biophys. Acta BBA-Proteins Proteomics 2005, 1754, 316–323. [Google Scholar] [CrossRef]
- Formstecher, E.; Ramos, J.W.; Fauquet, M.; Calderwood, D.A.; Hsieh, J.-C.; Canton, B.; Nguyen, X.-T.; Barnier, J.-V.; Camonis, J.; Ginsberg, M.H.; et al. PEA-15 Mediates Cytoplasmic Sequestration of ERK MAP Kinase. Dev. Cell 2001, 1, 239–250. [Google Scholar] [CrossRef]
- Hao, C.; Beguinot, F.; Condorelli, G.; Trencia, A.; Meir, E.G.V.; Yong, V.W.; Parney, I.F.; Roa, W.H.; Petruk, K.C. Induction and Intracellular Regulation of Tumor Necrosis Factor-related Apoptosis-inducing Ligand (TRAIL) Mediated Apotosis in Human Malignant Glioma Cells. Cancer Res. 2001, 61, 1162–1170. [Google Scholar] [PubMed]
- Kitsberg, D.; Formstecher, E.; Fauquet, M.; Kubes, M.; Cordier, J.; Canton, B.; Pan, G.; Rolli, M.; Glowinski, J.; Chneiweiss, H. Knock-Out of the Neural Death Effector Domain Protein PEA-15 Demonstrates That Its Expression Protects Astrocytes from TNFα-Induced Apoptosis. J. Neurosci. 1999, 19, 8244–8251. [Google Scholar] [CrossRef] [Green Version]
- Trencia, A.; Perfetti, A.; Cassese, A.; Vigliotta, G.; Miele, C.; Oriente, F.; Santopietro, S.; Giacco, F.; Condorelli, G.; Formisano, P.; et al. Protein Kinase B/Akt Binds and Phosphorylates PED/PEA-15, Stabilizing Its Antiapoptotic Action. Mol. Cell. Biol. 2003, 23, 4511. [Google Scholar] [CrossRef] [Green Version]
- Araujo, H.; Danziger, N.; Cordier, J.; Glowinski, J.; Chneiweiss, H. Characterization of PEA-15, a major substrate for protein kinase C in astrocytes. J. Biol. Chem. 1993, 268, 5911–5920. [Google Scholar]
- Yeh, P.Y.; Chuang, S.-E.; Yeh, K.-H.; Song, Y.C.; Chang, L.L.-Y.; Cheng, A.-L. Phosphorylation of p53 on Thr55 by ERK2 is necessary for doxorubicin-induced p53 activation and cell death. Oncogene 2004, 23, 3580–3588. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Wu, D.; Hirao, A.; Lahti, J.M.; Liu, L.; Mazza, B.; Kidd, V.J.; Mak, T.W.; Ingram, A.J. ERK Activation Mediates Cell Cycle Arrest and Apoptosis after DNA Damage Independently of p53. J. Biol. Chem. 2002, 277, 12710–12717. [Google Scholar] [CrossRef] [Green Version]
- Helfrich, B.A.; Kim, J.; Gao, D.; Chan, D.C.; Zhang, Z.; Tan, A.-C.; Bunn, P.A. Barasertib (AZD1152), a Small Molecule Aurora B Inhibitor, Inhibits the Growth of SCLC Cell Lines In Vitro and In Vivo. Mol. Cancer Ther. 2016, 15, 2314–2322. [Google Scholar] [CrossRef] [Green Version]
- Xingyu, Z.; Peijie, M.; Dan, P.; Youg, W.; Daojun, W.; Xinzheng, C.; Xijun, Z.; Yangrong, S. Quercetin suppresses lung cancer growth by targeting Aurora B kinase. Cancer Med. 2016, 5, 3156–3165. [Google Scholar] [CrossRef] [Green Version]
- Uitdehaag, J.C.M.; de Roos, J.A.D.M.; Prinsen, M.B.W.; Willemsen-Seegers, N.; de Vetter, J.R.F.; Dylus, J.; van Doornmalen, A.M.; Kooijman, J.; Sawa, M.; van Gerwen, S.J.C.; et al. Cell panel profiling reveals conserved therapeutic clusters and differentiates the mechanism of action of different PI3K/mTOR, Aurora kinase and EZH2 inhibitors. Mol. Cancer Ther. 2016, 15, 3097–3109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, J.S.; Ho, A.L.; Tse, A.N.; Coward, J.; Cheema, H.; Ambrosini, G.; Keen, N.; Schwartz, G.K. Aurora B Kinase Regulates the Postmitotic Endoreduplication Checkpoint via Phosphorylation of the Retinoblastoma Protein at Serine 780. Mol. Biol. Cell 2009, 20, 2218–2228. [Google Scholar] [CrossRef] [Green Version]
- Wiedemuth, R.; Klink, B.; Fujiwara, M.; Schröck, E.; Tatsuka, M.; Schackert, G.; Temme, A. Janus face-like effects of Aurora B inhibition: Antitumoral mode of action versus induction of aneuploid progeny. Carcinogenesis 2016, 37, 993–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent-Salomon, A.; Gruel, N.; Lucchesi, C.; MacGrogan, G.; Dendale, R.; Sigal-Zafrani, B.; Longy, M.; Raynal, V.; Pierron, G.; de Mascarel, I.; et al. Identification of typical medullary breast carcinoma as a genomic sub-group of basal-like carcinomas, a heterogeneous new molecular entity. Breast Cancer Res. 2007, 9, R24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, P.L.; Viara, E.; Hupé, P.; Pierron, G.; Liva, S.; Neuvial, P.; Brito, I.; Lair, S.; Servant, N.; Robine, N.; et al. VAMP: Visualization and analysis of array-CGH, transcriptome and other molecular profiles. Bioinformatics 2006, 22, 2066–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, E.Q.; Rosenstein, L. The Color and Ionization of Crystal-Violet. J. Am. Chem. Soc. 1914, 36, 1452–1473. [Google Scholar] [CrossRef]
- Nicoletti, I.; Migliorati, G.; Pagliacci, M.C.; Grignani, F.; Riccardi, C. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J. Immunol. Methods 1991, 139, 271–279. [Google Scholar] [CrossRef]
- Lemeer, S.; Ruijtenbeek, R.; Pinkse, M.W.H.; Jopling, C.; Heck, A.J.R.; den Hertog, J.; Slijper, M. Endogenous Phosphotyrosine Signaling in Zebrafish Embryos. Mol. Cell. Proteomics 2007, 6, 2088–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemeer, S.; Jopling, C.; Naji, F.; Ruijtenbeek, R.; Slijper, M.; Heck, A.J.R.; den Hertog, J. Protein-Tyrosine Kinase Activity Profiling in Knock Down Zebrafish Embryos. PLoS ONE 2007, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattei, J.C.; Bouvier-Labit, C.; Barets, D.; Macagno, N.; Chocry, M.; Chibon, F.; Morando, P.; Rochwerger, R.A.; Duffaud, F.; Olschwang, S.; et al. Pan Aurora Kinase Inhibitor: A Promising Targeted-Therapy in Dedifferentiated Liposarcomas With Differential Efficiency Depending on Sarcoma Molecular Profile. Cancers 2020, 12, 583. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12030583
Mattei JC, Bouvier-Labit C, Barets D, Macagno N, Chocry M, Chibon F, Morando P, Rochwerger RA, Duffaud F, Olschwang S, et al. Pan Aurora Kinase Inhibitor: A Promising Targeted-Therapy in Dedifferentiated Liposarcomas With Differential Efficiency Depending on Sarcoma Molecular Profile. Cancers. 2020; 12(3):583. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12030583
Chicago/Turabian StyleMattei, Jean Camille, Corinne Bouvier-Labit, Doriane Barets, Nicolas Macagno, Mathieu Chocry, Frédéric Chibon, Philippe Morando, Richard Alexandre Rochwerger, Florence Duffaud, Sylviane Olschwang, and et al. 2020. "Pan Aurora Kinase Inhibitor: A Promising Targeted-Therapy in Dedifferentiated Liposarcomas With Differential Efficiency Depending on Sarcoma Molecular Profile" Cancers 12, no. 3: 583. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12030583