
The Emerging Factors and Treatment Options for NAFLD-Related Hepatocellular Carcinoma


1
Department of Veterinary Pathobiology, University of Missouri, Columbia, MO 65211, USA
2
Department of Surgery, University of Missouri, Columbia, MO 65211, USA
*
Author to whom correspondence should be addressed.
Cancers 2021, 13(15), 3740; https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153740
Submission received: 2 June 2021
/
Revised: 22 July 2021
/
Accepted: 24 July 2021
/
Published: 26 July 2021
(This article belongs to the Special Issue Molecular Mechanisms Underlying Tumor Onset and Progression in Liver Cancers)
Abstract
:Simple Summary
Nonalcoholic fatty liver disease (NAFLD) is the most common chronic liver disease, and it is an increasing factor in the cause of hepatocellular carcinoma (HCC). The incidence of NAFLD has increased in recent decades, accompanied by an increase in the prevalence of other metabolic diseases, such as obesity and type 2 diabetes. However, current treatment options are limited. Both genetic factors and non-genetic factors impact the initiation and progression of NAFLD-related HCC. The early diagnosis of liver cancer predicts curative treatment and longer survival. Some key molecules play pivotal roles in the initiation and progression of NAFLD-related HCC, which can be targeted to impede HCC development. In this review, we summarize some key factors and important molecules in NAFLD-related HCC development, the latest progress in HCC diagnosis and treatment options, and some current clinical trials for NAFLD treatment.
Abstract
Hepatocellular carcinoma (HCC) is the most common type of primary liver cancer, followed by cholangiocarcinoma (CCA). HCC is the third most common cause of cancer death worldwide, and its incidence is rising, associated with an increased prevalence of obesity and nonalcoholic fatty liver disease (NAFLD). However, current treatment options are limited. Genetic factors and epigenetic factors, influenced by age and environment, significantly impact the initiation and progression of NAFLD-related HCC. In addition, both transcriptional factors and post-transcriptional modification are critically important for the development of HCC in the fatty liver under inflammatory and fibrotic conditions. The early diagnosis of liver cancer predicts curative treatment and longer survival. However, clinical HCC cases are commonly found in a very late stage due to the asymptomatic nature of the early stage of NAFLD-related HCC. The development of diagnostic methods and novel biomarkers, as well as the combined evaluation algorithm and artificial intelligence, support the early and precise diagnosis of NAFLD-related HCC, and timely monitoring during its progression. Treatment options for HCC and NAFLD-related HCC include immunotherapy, CAR T cell therapy, peptide treatment, bariatric surgery, anti-fibrotic treatment, and so on. Overall, the incidence of NAFLD-related HCC is increasing, and a better understanding of the underlying mechanism implicated in the progression of NAFLD-related HCC is essential for improving treatment and prognosis.
1. Introduction
Primary liver cancer was the sixth most commonly diagnosed and the third most common cause of cancer-related death worldwide in 2020 [1]. Hepatocellular carcinoma (HCC) comprises approximate 80% of primary liver cancer (PLC) cases [2], whereas cholangiocarcinoma (CCA) represents 10% to 15% of PLC cases [1]. Combined hepatocellular-cholangiocarcinoma (CHC) is a rare case in PLC [3,4]. Multiple factors contribute to the development of HCC, such as diet [5,6], infection with hepatitis viruses [7,8], alcohol abuse [9,10], and bioactive compounds [11,12]. Recent studies show that hepatitis C viral infection is the most common causal factor for HCC but that it shows a declining trend, whereas nonalcoholic fatty liver disease (NAFLD) or its advanced subtype, nonalcoholic steatohepatitis (NASH), is the most rapidly growing factor contributing to HCC development in the United States [13]. Myers et al. reported that in a study performed in western Switzerland, NAFLD or metabolic-associated fatty liver disease (MAFLD) was found to be an increased inducing factor for HCC incidence, being especially higher in women than in men, whereas other etiologies remained stable [14]. The early diagnosis of liver cancer is critically important for curative treatment, since early-stage HCC can be locally ablated or resected. Surgery such as laparoscopic surgery is recommended as the first-line therapy for HCC patients with an early diagnosis [15]. Unfortunately, NAFLD-related HCC progression does not have obvious clinical symptoms, which means that most cases are found in the late stage of the disease [16]. Furthermore, the increasing prevalence of NAFLD worldwide and limited therapeutic options may raise the incidence of HCC [17].
The development of HCC is associated with age, sex, geography, and etiology [2]. Men have a much higher chance of developing HCC than women. Furthermore, the diagnosis of HCC in men aged ≥60 years has dramatically increased, and milder changes have been observed in women. The sex-induced difference in the incidence of HCC is dependent not only on the hormone estrogen [18], but also on other factors, such as gut microbiota, bile acids (BAs), and microRNAs (miRNAs) [19]. Factors causing the development of NAFLD-related HCC (Figure 1), including epigenetic and genetic factors and transcriptional and post-transcriptional factors, as well as their diagnosis and treatment, are discussed in this study.
2. Genetic Factors
Genetic factors, such as patatin-like phospholipase domain-containing protein 3 (PNPLA3) [20,21], transmembrane-6 superfamily member 2 (TM6SF2) [22], and programmed cell death-1 (PDCD1) encoding PD-1, are associated with NAFLD-related HCC initiation and progression [23].
The allele alteration of rs7421861 A > G in PDCD1 gene is associated with a decreased frequency of NAFLD-HCC progression, since the wild-type A allele has been observed more in patients with NAFLD-HCC [23]. In contrast, the allele alteration of rs10204525 C > T in PDCD1 gene increased the progression of NAFLD-HCC compared to the wild-type C allele. In silico analysis showed that the rs7421861 A allele in PDCD1 gene was associated with the higher expression of PD-1 compared to the G allele, which suggests that the G allele decreases PD-1-mediated immune exhaustion to suppress HCC growth [23]. In addition, a mutation of the rs7421861 allele in PDCD1 gene, located in intron 1 with richness in regulatory and splicing sites, may cause splicing disruption, translational inhibition, and a change in the mRNA secondary structure [24].
Patients with the rs58542926 C > T genetic variant of the TM6SF2 gene, encoding the E167K amino acid substitution, showed a lower serum lipid content, but had more severe hepatic steatosis, inflammation, ballooning, and fibrosis, and were more susceptible to develop NASH [25,26]. The T allele was shown to be associated with the reduction of TM6SF2 gene and protein expression in the liver [26]. A meta-analysis study showed that the rs58542926 T allele in TM6SF2 gene had a significant association with HCC development compared to the C allele [27]. This effect may be mediated by regulating the cell cycle [28] and upregulating inflammatory cytokines, such as IL-2 and IL-6 [29].
The rs599839 A > G variant, localized in the genetic cluster of cadherin EGF LAG seven-pass G-type receptor 2 (CELSR2)-proline/serine-rich coiled-coil protein 1 (PSRC1)- sortilin 1 (SORT1), was associated with reduced severity of dyslipidemia in NAFLD patients with a higher risk of cardiovascular comorbidities [30]. This variant was associated with increased hepatic expression of CELSR2, PSRC1, and SORT1 in NAFLD patients. In addition, the data from The Cancer Genome Atlas (TCGA) showed that PSRC1-overexpression promoted HCC development [30]. However, this variant was not significantly correlated with hepatic steatosis, ballooning, lobular inflammation, or fibrosis.
In addition, the rs641738 C > T variant, near two genes encoding membrane-bound O-acyltransferase domain-containing 7 (MBOAT7) and transmembrane channel-like 4 (TMC4), was shown to be associated with the progression of NAFLD and liver fibrosis [31]. The loss of function of MBOAT7 is identified to be a factor contributing to NAFLD progression. Another study also reported that the rs641738 T allele of MBOAT7 gene is associated with NAFLD-related HCC in non-cirrhotic patients [32]. However, the rs641738 C > T variant in TMC4 gene was not found to be a genetic risk in relation to increasing the development of NAFLD [33]. Similarly, in this study, the protein level of MBOAT7 was found to be lower in the liver of NAFLD patients. The rs641738 C allele has been shown to be associated with a high expression of MBOAT7 that localizes into the membranes, which helps deliver membrane metabolites into intracellular compartments. In contrast, the T risk allele is associated with the reduction of MBOAT7, which is favorable for the increase of saturated phospholipids and triglyceride (TG) synthesis [34]. More evidence is needed to support the role of the rs641738 variant of MBOAT7/TMC4 in the susceptibility of NAFLD and NAFLD-related HCC progression.
3. Epigenetic Factors
Instead of a change in DNA sequence, epigenetic changes modulated by factors such as age and environment can also impact the progression of NAFLD-related HCC. Epigenetic factors, including DNA methylation, long non-coding RNAs (lncRNAs), and miRNAs, are considered to have profound effects on NAFLD-related HCC progression.
3.1. DNA Methylation
DNA methylation is implicated in liver fibrosis, cirrhosis, and HCC. Hypermethylation of CpG islands in genes such as CELSR1 and collapsin response mediator protein 1 (CRMP1), and hypomethylation of CpG loci in small proline-rich protein 3 (SPRR3) and tumor necrosis factor ligand superfamily member 15 (TNFSF15) genes were found in HCC and cirrhotic liver tissues compared to noncirrhotic control liver tissues [35]. Hypermethylation in promoters of genes such as Ras association domain-containing protein 1 (RASSF1A) and docking protein 1 (DOK1) was associated with the pathogenesis of hepatocarcinogenesis [36]. DNA methylation at specific CpGs within genes known to affect fibrogenesis, such as peroxisome proliferator-activated receptor alpha (PPARα), transforming growth factor-beta 1 (TGF-β1), and platelet-derived growth factor alpha (PDGFα) genes, was observed in patients with NAFLD or alcoholic liver disease (ALD) associated with the progression of fibrosis [37]. In addition to the dysregulation of DNA methylation, histone acetylation or methylation-mediated epigenetic changes can lead to cell apoptosis in the development of NAFLD and HCC. This specific subject has been well-reviewed in another published paper [38], and is thus not discussed in this paper.
3.2. Long Non-Coding RNAs
LncRNAs, defined as RNAs with a length of ≥200 nucleotides that are not translated into functional proteins, play an important role in endoplasmic reticulum (ER) stress and oxidative stress. The expression of more than 3000 lncRNAs was observed to be changed in the liver tissues of db/db mice fed with a NASH diet, and the expression of 381 lncRNAs was significantly increased during NAFLD progression to NASH [39]. Among these, LncRNA gm9795 can upregulate ER stress molecules and the nuclear factor kappa B (NF-κB)/c-Jun N-terminal kinase (JNK) signaling pathway to increase proinflammatory cytokine production, such as that of TNF-α, interleukin-6 (IL-6), and IL-1β. Increased expression of LncRNA SNHG20 was observed in the livers of NAFLD-related HCC-bearing mice and human patients with NALFD-related HCC [40]. Silencing SNHG20 can delay the progression of NAFLD to HCC [40]. In addition, lncRNAs (e.g., MYLK-AS1) can act as competitive endogenous RNA, inducing miRNAs (e.g., miR-424-5) to regulate tumor angiogenesis in HCC [41].
3.3. MicroRNAs
MiRNAs contribute to the progression of NAFLD, liver fibrosis, and HCC development. For example, microRNA-21 (miR-21) has been shown to impair lipid metabolism in mice with NAFLD and human liver cancer cell line HepG2 cells, and to inhibit the progression of xenograft tumors induced by HepG2 cells, as miR-21 knockdown can impair lipid accumulation and tumor growth by targeting HMG-Box transcription factor 1 (HBP1)-p53, part of the sterol regulatory element-binding protein 1c (SREBP1c) signaling pathway [42]. In addition, hepatic miR-21 expression has been shown to be upregulated in a methionine-choline-deficient (MCD) diet-induced mouse NASH model and in human patients with NASH [43]. Suppressing miR-21 function with antagomir-21 can reduce liver injury, inflammation, and fibrosis in low-density lipoprotein (LDL) receptor-deficient mice, but not in PPARα-deficient mice [43]. A recent study using a doxycycline-inducible transgenic zebrafish model (LmiR21) with hepatic overexpression of miR-21 showed that miR-21 overexpression contributed to liver steatosis, inflammation, and fibrosis, the broad spectrum of NAFLD [44]. Moreover, LmiR21 zebrafishes showed the NAFLD-HCC phenotype at 10 months post-fertilization, and they also showed a higher percentage of chemical-induced liver fibrosis and HCC compared to wild-types under the chemical stimuli.
Hepatocyte-specific miR-122a accounts for 70% of the total miRNAs in the liver and is downregulated in about 70% of human HCC and all HCC-derived cell lines [45]. MiR-122a-deficient mice develop reversible steatohepatitis, fibrosis, and HCC [46]. In addition, the incidence of HCC shows a sexual disparity, being 3.9 times higher in male mice compared to female mice.
The expression of miR-223 in hepatocytes is highly increased in mice when feeding on a high-fat diet (HFD) and in the liver samples of patients with NASH. Feeding with an HFD significantly enhanced the expression of proinflammatory and cancer-related genes in miR-233-knockout mice compared to that in wild-type mice [47].
In addition, some chemical RNA modifications are emerging factors of epigenetic regulation. For example, N6-methyladenosine (m6A), the most abundant chemical modification of eukaryotic mRNA, plays a critical role in regulating adipogenesis [48]. Methyltransferase-like 3 (METTL3)-mediated m6A modification can inhibit the suppressor of cytokine signaling 2 (SOCS2) to promote HCC progression [49].
4. Transcriptional Factors
Transcriptional factors, such as E2Fs transcriptional factors, hypoxia-inducible factors (HIFs), Forkhead box (FOXO), and PPARs, modulate NAFLD progression through different signaling pathways (Figure 2). For example, PPARγ can regulate lipid metabolism by regulating the Toll-like receptor 4 (TLR4)/NF-κB signaling pathway [48].
4.1. E2F1 and E2F2
The expression of transcription factors E2F1 and E2F2 is positively correlated and increased in NAFLD-related HCC. Their deficiency decreased hepatocarcinogenesis induced by HFD plus diethylnitrosamine (DEN) administration, with a reduction of lipid accumulation [50]. The molecular mechanism shows that E2F1 reversely modulates carnitine palmitoyltransferase 2 (CPT2), an essential enzyme for fatty acid oxidation, the downregulation of which promotes HCC development via acylcarnitine accumulation in a lipid-rich environment [51]. E2F1 is upstream of the transcription factor of ribosome binding protein 1 (RRBP1), which can be upregulated by high glucose. Inhibiting E2F1 expression decreased the expression of RRBP1, remarkedly reducing the proliferation and metastasis of HepG2 cells [52].
4.2. FOXOs
FOXO transcriptional factors play important roles in regulating hepatic glucose [53] and lipid homeostasis [54], cell growth and apoptosis [55], and liver inflammation and fibrosis [56]. FOXOs are the downstream signaling of protein kinase B (AKT), which can phosphorylate the serine or threonine of FOXOs to regulate multiple cellular functions. FOXO3 can activate the promoter of SREBP1c to aggravate liver TG and intrahepatic lipid accumulation [54]. However, feeding with an HFD induced more severe hepatic steatosis and fibrogenesis in Foxo1/3/4 triple knockout mice compared to wild-type mice via upregulating profibrotic genes such as C-C motif chemokine ligand 2 (CCL2), alpha-1 type I collagen (Col1A1), and TGF-β [56].
4.3. HIFs
HIFs, such as HIF-1α and HIF-2α, are transcription factors induced in response to a hypoxic environment, which plays a pivotal role in liver inflammation [57] and tumor growth [58]. Hypoxia also affects NAFLD-HCC progression, since HIF-2α was found to be increased in HCC tissues from NAFLD-HCC patients compared to tissues from non-NAFLD-HCC subjects [59]. The upregulation of HIF-2α was negatively associated with the overall survival (OS) of HCC patients and was positively associated with hepatic lipid accumulation and activation of the phosphoinositide 3-kinase (PI3K)-AKT-mechanistic target of rapamycin (mTOR) signaling pathway [59].
4.4. KLF6
As a transcription factor, Krüppel-like factor 6 (KLF6) plays essential roles in cellular processes, including cell proliferation, differentiation, and cell death [60]. In NAFLD, KLF6 regulates liver glucose and lipid metabolism by regulating the activity of PPARα and PPARα-regulated genes such as phosphoenolpyruvate carboxykinase (PEPCK) [61]. In addition, KLF6 binds the promoter of glucokinase (GCK) in NAFLD, which can regulate insulin resistance and the glucose level in the blood [62]. A mutation of a polymorphism, KLF6 intervening sequence (IVS) 1–27 G > A, was found to be positively associated with liver fibrosis in NAFLD patients [63]. Accumulating studies show that KLF6 is a tumor suppresser gene against HCC [64,65,66].
4.5. PPARs
PPARs are important transcriptional factors in modulating liver inflammation [67], lipid metabolism [68], and cancer growth [69]. All three PPAR subtypes, including PPARα [70], PPARβ/δ [71], and PPARγ [72], play important roles in lipid metabolism, either in the liver or adipose tissues. For example, hepatocyte-specific PPARα deficiency mice showed a significant increase in oleic acid and linoleic acid compared with wild-type mice in fasting, partly due to a fasting-induced increase in fibroblast growth factor 21 (FGF21) expression [73]. Metabolic syndrome, such as insulin resistance and hepatic steatosis, can be induced by activating JNK to suppress PPARα function. However, long-term JNK deficiency can result in CCA by interrupting cholesterol metabolism and bile acid homeostasis [74]. In contrast, metabolites such as 3-hydroxybutyric acid, induced by treatment with the antiangiogenic agent apatinib, can induce PPARα activation in the liver tissue to inhibit tumor growth [75].
Additionally, PPARβ/δ [76] and PPARγ [77] are implicated in liver homeostasis by regulating glucose and fatty acid metabolism. PPAR-γ agonists (e.g., pioglitazone) show clinical effects in the reduction of hepatic or visceral fat and necroinflammation in human patients [78]. PPARβ/δ activator GW501516 can prevent HFD-induced hypertriglyceridemia and hepatic fatty acid oxidation, and increase the production of 16:0/18:1-phosphatidylcholine, an endogenous ligand for PPARα in the liver [79]. A bioinformatic study showed that PPARγ was overexpressed in the livers of human patients with HCC, which was associated with poor OS [80]. However, the exact roles of PPARs in the progression of NAFLD-related HCC need to be illustrated.
4.6. SREBP-1
SREBP-1 is a transcriptional factor and plays a pivotal role in the proliferation and metastasis of liver cancer cells by regulating fatty acid synthesis and [81] and suppressing liver inflammation [82]. Other factors, such as long-chain acyl CoA synthetase 4 (ACSL4) [83], caveolin-1 (Cav1) [84], and zinc fingers and homeoboxes 2 (ZHX2) [85], can regulate lipid metabolism via the SREBP1 signaling pathway. Lipid metabolism is correlated tightly with glucose metabolism in HCC. Inhibition of SREBP-1 expression can suppress glucose metabolism in HCC cells, resulting in a synergistic anti-tumor effect combined with immunotherapy with Sorafenib on HCC in vivo [81].
5. Post-Transcriptional Modification
Post-transcriptional factors (Figure 2) such as RNA-binding proteins (RBPs) and RNA splicing factor (SF) contribute to liver damage, NAFLD development, and HCC progression [90]. Novel anti-HCC therapies can be developed based on post-transcriptional regulation, such as the administration of adenovirus-mediated trans-splicing ribozymes [91,92]. The underlying mechanism of post-transcriptional modification in NAFLD-related HCC progression needs to be investigated further.
5.1. RNA Splicing Factor
Dysregulation of RNA splicing factors contributes to the development of steatosis and NAFLD progression [93]. Silencing of some splicing machinery components in vitro, such as RNA binding motif protein 45 (RBM45) and staphylococcal nuclease domain containing 1 (SND1), can inhibit fat accumulation by modulating the expression of key de novo lipogenesis enzymes [93]. Dysregulation of RBM45 and SND1 has been associated with the progression of cancers, including HCC [94]. A splicing factor, arginine/serine-rich 10 (SFRS10) in the liver, directly regulates the splicing of lipin 1 encoded by the LPIN gene, a key regulator of lipid metabolism. SFRS10 has been shown to be downregulated in obese human livers and livers in HFD-fed mice [95]. Reducing or inhibiting SFRS10 expression can increase lipid accumulation in hepatocytes and plasma TG and very-low-density lipoprotein (VLDL) secretion by increasing the lipogenic β isoform of LPIN1 [95].
5.2. RNA-Binding Proteins
Sirtuin 1, encoded by the SIRT1 gene, can deacetylate an RNA-binding protein quaking 5 (QKI 5), which inhibits TG synthesis in vivo and in vitro via the PPARα/FoxO1 signaling pathway and suppresses NAFLD progression in mice [96]. Another study showed that a broadly expressed RNA-binding protein human antigen R (HuR) can accelerate NASH progression by increasing death receptor 5 (DR5)/caspase 8/caspase 3-mediated hepatocyte death and liver injury [97].
Aberrant expression of RBPs has been shown across many malignant tumors, including HCC [98,99]. A high score based on the expression of these RBPs was associated with poor overall survival of HCC [99]. Another study showed that aberrant expression of four key RBPs, including mitochondrial ribosomal protein L54 (MRPL54), enhancer of zeste homolog 2 (EZH2), PPARγ coactivator 1 alpha (PPARGC1A), and eukaryotic translation initiation factor 2-alpha kinase 4 (EIF2AK4), can be applied to HCC prognosis [100].
5.3. RNA Editing
The inactivation or low activity of apolipoprotein B (ApoB) is associated with poor prognosis of HCC, upregulation of oncogenic and metastatic factors, and the inhibition of tumor suppressor genes [101,102], such as p53 and phosphatase and tensin homolog (PTEN). The ApoB mRNA editing enzyme, catalytic polypeptide 1 (APOBEC1) complementation factor (A1CF) regulates posttranscriptional ApoB mRNA editing (C > U). Aged hepatocyte-specific A1CF-transgenic mice can spontaneously develop hepatic fibrosis and HCC, and disease can be accelerated when those mice are fed a high-fructose high-fat diet [103]. In addition, the expression of A1CF was associated with advanced fibrosis and low survival in NAFLD-related HCC patients [103].
Overall, both genetic and epigenetic factors play critical roles in NAFLD-related HCC progression. Furthermore, recent studies show that NAFLD/NASH modulates intrahepatic immune responses to inhibit anti-tumor immunity, resulting in the progression of HCC. For example, Tim Greten et al. reported that dysregulation of lipid metabolism, specifically for linoleic acid, causes more oxidative damage in mitochondria during NAFLD, resulting in a dramatic reduction of CD4+ T cells and promoting hepatocarcinogenesis [104]. Their further study showed that NASH can impair the effects of M3-RNA vaccine and anti-OX40 antibody treatment against mouse liver tumors induced by intrahepatic injections of B16 melanoma and CT26 colon cancer cells [105]. In contrast, the administration of N-acetylcysteine in NASH mice restored CD4+ T cells and resulted in improved effects of the M3-RNA vaccine and anti-OX40 antibody. Moreover, immunotherapy including anti-PD-1 or anti-PD-L1 treatment reduced the overall survival of NASH-HCC patients compared to HCC patients induced with other etiologies [106]. Preclinical mouse model study showed that anti-PD-1 expanded the exhausted and unconventionally activated T cells, such as CXCR6+PD-1+CD8+ T cells, which lost the immune surveillance function and promoted NASH-HCC progression. PD-1+CD8+ T cells have been reported to be correlated with a poor clinical outcome in HCC patients [107,108]. Thus, the development of NAFLD/NASH modulates intrahepatic immunity to impair the anti-HCC immune response.
6. Potential Diagnosis of NAFLD-Related HCC
The development of NAFLD-related HCC is caused by multiple factors. For clinical diagnosis, liver biopsy is still considered the gold standard for clinical decisions. However, it is invasive and may not represent the tumor heterogeneity due to the sample size of the biopsy [109]. In addition, it is not appropriate to be applied to monitor the dynamic progression of HCC. Moreover, the diagnosis of liver fibrosis and cirrhosis may not be helpful in predicting all cases of NAFLD-related HCC, since some patients have developed HCC in the background of NAFLD/NASH without the progression of liver fibrosis and cirrhosis [110,111]. Currently, imaging techniques are essential for the diagnosis of chronic liver disease and HCC. Dynamic multiphase contrast-enhanced computed tomography (CT) scanning and magnetic resonance imaging (MRI) are the most commonly used methods to detect HCC in clinical diagnosis [112,113]. Still, there are some challenges, such as the lack of standardization in image acquisition protocols and optimization of the radiomics analysis procedure [114]. Additional non-invasive or less harmful diagnostic methods, such as biomarkers, score systems, and algorithms, have been investigated to improve the diagnosis of HCC, including its initiation, progression, and potential recurrence. In Table 1, we summarize some developed biomarkers and imaging techniques that provide assistance in the diagnosis of NAFLD-HCC. The combination of different markers and detection methods is helpful in order to provide a precise diagnosis and to make clinical decisions relating to treatment.
7. Treatment Options against HCC
Treatment options for HCC can be broadly classified into surgical resection and non-surgical therapies dependent upon the stage of the disease, liver function, availability of donor organs, cost of treatment, and so on [128]. The Westernized diet and sedentary lifestyle promote the progression of NAFLD [129,130]. A cohort study in Europe also showed that physical activity is inversely associated with the risk of HCC [131]. However, excessive exercise can impact the host metabolism to reduce glucose control by impairing mitochondrial function [132]. Here, therapies for regular HCC, NAFLD, and NAFLD-related HCC, including immunotherapy, CAR T cell therapy, peptide treatment, bariatric surgery, and treatment for liver fibrosis, are discussed.
7.1. Treatments against HCC
7.1.1. Systemic Therapies and Immunotherapy
The recurrence of HCC is a big concern after surgical operations. Immunotherapy is helpful to reduce the recurrence of HCC and provides treatment options for advanced HCC that is not suitable for surgical resection. Here, we first briefly summarize some approved first- and second-line treatment options for regular HCC, which may be applied in NAFLD-related HCC treatment.
Sorafenib, a multi-kinase inhibitor, was the first systemic therapy approved by the U.S. Food and Drug Administration (FDA) for patients with unresectable HCC in 2008 [133], and has been approved for the treatment of advanced renal cell carcinoma (RCC) [134]. Lenvatinib is another FDA-approved systemic treatment for unresectable advanced HCC, approved in 2018 [135]. Phase 3 non-inferiority trial data showed that the median survival time for lenvatinib-treated patients was 13.6 months (95% CI, 12.1–14.9 months) compared with a median survival time of 12.3 months (95% CI, 10.4–13.9 months) for patients with sorafenib treatment [135]. In 2020, the FDA approved treatment with atezolizumab in combination with bevacizumab for adult patients with unresectable locally advanced or metastatic HCC without prior systemic therapy [136], since the combined treatment improved OS and progression-free survival compared to treatment with sorafenib [137].
Since 2017, the U.S. FDA has approved several drugs for HCC treatment as the second line after sorafenib treatment, including kinase inhibitors (regorafenib, lenvatinib, cabozantinib, and ramucirumab), immune checkpoint inhibitors (nivolumab and pembrolizumab), and monoclonal antibodies (atezolizumab plus bevacizumab). In 2017, regorafenib was approved as the first drug by the FDA for the treatment of advanced HCC in patients who had previously been treated with sorafenib [138]. In 2018, pembrolizumab, a monoclonal antibody against PD-1, was approved by the FDA for the treatment of patients with HCC who have been previously treated with sorafenib. In 2019, the FDA approved another drug, cabozantinib, for patients with HCC with prior treatment with sorafenib, since phase 3 trial results indicated that cabozantinib treatment resulted in longer overall survival and progression-free survival than a placebo in previously treated patients with advanced HCC [139]. Following up, the FDA approved ramucirumab as a sole treatment for HCC patients who have serum AFP levels ≥ 400 ng/mL with prior treatment with sorafenib. In addition, there are some combined treatments, such as nivolumab and ipilimumab, that may improve outcomes [140].
Additionally, there are increasing numbers of clinical trials evaluating different therapies in a variety of combinations for the systemic treatment of HCC at different disease stages with the Barcelona Clinic for Liver Cancer (BCLC) criteria, which is well summarized in another review [141]. The FDA-approved treatments for HCC are summarized in Table 2.
7.1.2. CAR T Cells
Chimeric antigen receptor (CAR) T cells have been tested in a variety of diseases, including aging [142], autoimmune disease [143], and tumors such as B-cell acute lymphatic leukemia [144] and myeloma [145]. The principle of CAR T cell immunotherapy is to engineer T cells to express CARs, which consist of an extracellular antigen recognition domain fused to intracellular T-cell receptor (TCR) signaling and co-stimulatory domains [146]. Genetically engineered CAR T cells recognize antigens on malignancy cells to effectively damage them and overcome tolerance. A phase 1 trial showed that CAR-glypican-3 (GPC-3) T-cell therapy showed some early signs of effectiveness against advanced HCC in patients [147], such as decreased counts of different lymphocytes. In addition, there are other tumor-targeting antigens against HCC, such as AFP [148] and New York esophageal squamous cell carcinoma-1 (NY-ESO-1) [120]. In addition, these antigens including AFP and GPC-3 are observed in NAFLD-related HCC [149].
7.1.3. Peptides
Peptide vaccination has been applied in the clinic to treat HCC. In a study of vaccination with human telomerase reverse transcriptase-derived peptide 461 (hTERT461, VYGFVRACL) in a total of 14 HCC patients, 11 patients (71.4%) showed hTERT461-specific cytotoxic T cells (CTLs) in blood post-vaccination at 4 weeks [161]. Of the response patients, 57.1% of patients (five of them) did not show HCC recurrence. In contrast, HCC recurred in all three patients (100.0%) without hTERT461-specific CTLs. In 15 HCC patients after vaccination with AFP-derived peptide 357 (AFP357) vaccine, 53.3% of patients showed slow tumor growth and 26.7% had AFP357-specific CD8 T cells [162]. Only one patient had a complete response more than 2 years; the functional T cells in this patient expressed a high avidity for AFP-specific T-cell receptors. Anti-cancer peptides are also potential treatment options for liver cancer [163].
7.2. Treatment against NAFLD
Bariatric surgery (BS) or weight loss surgery has been well established to provide excellent weight loss outcomes and improvements in comorbid medical conditions, including diabetes [164], NAFLD [165], cardiac function [166], and cancer [167]. Multiple mechanisms are involved in the effectiveness of weight loss surgery, including restriction (sleeve gastrectomy and gastric bypass) and malabsorption (gastric bypass and biliopancreatic diversion with duodenal switch) and through gut enteroendocrine hormonal effects [168] (sleeve gastrectomy, gastric bypass, and biliopancreatic diversion with duodenal switch). Weight loss surgery alters the gut microbiota, as well as circulating bile acids [169] and many blood metabolites [170]. These mechanisms are likely intricately related and function to regulate the effects of weight loss surgery on a person’s health [169]. However, there is a lack of evidence for the treatment of NAFLD-related HCC in clinical trials.
Many treatment agents have been tested in clinical trials for the treatment of NAFLD or NASH with promising effects, including polyphenols, bile acids, diet intervention, herb medicines, anti-inflammatory or antioxidant agents, hormones, and symbiotics (Table 3).
7.3. Treatment against Liver Fibrosis
TGF-β is a predominant profibrotic gene that causes the activation of hepatic stellate cells (HSCs) in the liver independently of causal factors such as a high-fat diet, alcohol, and other toxins such as carbon tetrachloride (CCL4). Strategies that block the TGF-β signaling pathway can inhibit the progression of liver fibrosis [181]. Bone morphogenetic proteins (BMPs) belonging to the large TGF-β family play an important role in tissue homeostasis. Accumulating evidence indicates that BMPs are involved in the development and progression of liver fibrosis and liver regeneration [182,183], becoming a new target for liver fibrosis. In addition, some other drugs can ameliorate NAFLD/NASH-associated liver fibrosis, such as the farnesoid X receptor (FXR) agonists obeticholic acid and isotschimgine [184,185]. Furthermore, changing lifestyles, such as the consumption of a healthy diet and appropriate exercise, also can prevent fibrosis progression in NAFLD patients.
Overall, the treatment options (Figure 3) for NAFLD-related HCC, targeting liver fibrosis, lipid accumulation, cancer cells, and immune responses, can be selected according to the pathogenesis of the liver disease and the health condition of the patient.
8. Conclusions
The prevalence of NAFLD-related HCC is increasing in developed countries and developing countries due to the consumption of fast food or a Western-like diet, less physical exercise, and an increased prevalence of obesity and diabetes. Patients with advanced NAFLD or NASH and liver fibrosis progression are at a higher risk of developing HCC. Imaging is the most commonly used technique for HCC diagnosis. The development of other non-invasive diagnostic methods is also critically important for better clinical treatment and prevention of HCC. Combined diagnosis with multiple diagnostic methods provides higher sensitivity and specificity for monitoring NAFLD-related HCC. Although there are several FDA-approved drug treatments, the overall survival rate and survival time are still not promising. In addition, some treatments such as bariatric surgery are more beneficial at the early stage of NAFLD but show no evidence of helping HCC therapy in clinical trials. With the development of large databases, along with artificial intelligence and machine learning, precision medicine in the future will improve the diagnosis of NAFLD-related HCC and provide better options for personal precise treatment of the disease.
Author Contributions
Conceptualization, C.Z. and M.Y.; data collection, C.Z. and M.Y.; writing—original draft preparation, C.Z. and M.Y.; writing—review and editing, C.Z. and M.Y. They contribute equally. All authors have read and agreed to the published version of the manuscript.
Funding
This manuscript received no external funding.
Data Availability Statement
All the data supporting the reported results can be found in the references.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Liu, Z.; Suo, C.; Mao, X.; Jiang, Y.; Jin, L.; Zhang, T.; Chen, X. Global incidence trends in primary liver cancer by age at diagnosis, sex, region, and etiology, 1990–2017. Cancer 2020, 126, 2267–2278. [Google Scholar] [CrossRef]
- He, C.; Mao, Y.; Wang, J.; Song, Y.; Huang, X.; Lin, X.; Li, S. The Predictive Value of Staging Systems and Inflammation Scores for Patients with Combined Hepatocellular Cholangiocarcinoma After Surgical Resection: A Retrospective Study. J. Gastrointest. Surg. 2018, 22, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Schizas, D.; Mastoraki, A.; Routsi, E.; Papapanou, M.; Tsapralis, D.; Vassiliu, P.; Toutouzas, K.; Felekouras, E. Combined hepatocellular-cholangiocarcinoma: An update on epidemiology, classification, diagnosis and management. Hepatobiliary Pancreat. Dis. Int. 2020, 19, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Simoni-Nieves, A.; Salas-Silva, S.; Chávez-Rodríguez, L.; Escobedo-Calvario, A.; Desoteux, M.; Bucio, L.; Souza, V.; Miranda-Labra, R.U.; Muñoz-Espinosa, L.E.; Coulouarn, C.; et al. The Consumption of Cholesterol-Enriched Diets Conditions the Development of a Subtype of HCC with High Aggressiveness and Poor Prognosis. Cancers 2021, 13, 1721. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Coker, O.O.; Chu, E.S.; Fu, K.; Lau, H.C.H.; Wang, Y.X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary cholesterol drives fatty liver-associated liver cancer by modulating gut microbiota and metabolites. Gut 2021, 70, 761–774. [Google Scholar] [CrossRef]
- Virzì, A.; Gonzalez-Motos, V.; Tripon, S.; Baumert, T.F.; Lupberger, J. Profibrotic Signaling and HCC Risk during Chronic Viral Hepatitis: Biomarker Development. J. Clin. Med. 2021, 10, 977. [Google Scholar] [CrossRef]
- Yang, M.; Parikh, N.D.; Liu, H.; Wu, E.; Rao, H.; Feng, B.; Lin, A.; Wei, L.; Lok, A.S. Incidence and risk factors of hepatocellular carcinoma in patients with hepatitis C in China and the United States. Sci. Rep. 2020, 10, 20922. [Google Scholar] [CrossRef] [PubMed]
- Petrick, J.L.; Campbell, P.T.; Koshiol, J.; Thistle, J.E.; Andreotti, G.; Beane-Freeman, L.E.; Buring, J.E.; Chan, A.T.; Chong, D.Q.; Doody, M.M.; et al. Tobacco, alcohol use and risk of hepatocellular carcinoma and intrahepatic cholangiocarcinoma: The Liver Cancer Pooling Project. Br. J. Cancer 2018, 118, 1005–1012. [Google Scholar] [CrossRef]
- Ochiai, Y.; Kawamura, Y.; Kobayashi, M.; Shindoh, J.; Kobayashi, Y.; Okubo, S.; Muraishi, N.; Kajiwara, A.; Iritani, S.; Fujiyama, S.; et al. Effects of alcohol consumption on multiple hepatocarcinogenesis in patients with fatty liver disease. Hepatol. Res. 2021, 51, 62–68. [Google Scholar] [CrossRef]
- Ricart, A.D. Drug-induced liver injury in Oncology. Ann. Oncol. 2017, 28, 2013–2020. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, L.N.Z.; Porta, L.D.; Rosim, R.E.; Petta, T.; Augusto, M.J.; Silva, D.M.; Ramalho, F.S.; Oliveira, C.A.F. Aflatoxin B(1) residues in human livers and their relationship with markers of hepatic carcinogenesis in São Paulo, Brazil. Toxicol. Rep. 2018, 5, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Stepanova, M.; Ong, J.P.; Jacobson, I.M.; Bugianesi, E.; Duseja, A.; Eguchi, Y.; Wong, V.W.; Negro, F.; Yilmaz, Y.; et al. Nonalcoholic Steatohepatitis Is the Fastest Growing Cause of Hepatocellular Carcinoma in Liver Transplant Candidates. Clin. Gastroenterol. Hepatol. 2019, 17, 748–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, S.; Neyroud-Caspar, I.; Spahr, L.; Gkouvatsos, K.; Fournier, E.; Giostra, E.; Magini, G.; Frossard, J.L.; Bascaron, M.E.; Vernaz, N.; et al. NAFLD and MAFLD as emerging causes of HCC: A populational study. JHEP Rep. 2021, 3, 100231. [Google Scholar] [CrossRef]
- Vitale, A.; Peck-Radosavljevic, M.; Giannini, E.G.; Vibert, E.; Sieghart, W.; Van Poucke, S.; Pawlik, T.M. Personalized treatment of patients with very early hepatocellular carcinoma. J. Hepatol. 2017, 66, 412–423. [Google Scholar] [CrossRef] [Green Version]
- Pocha, C.; Kolly, P.; Dufour, J.F. Nonalcoholic Fatty Liver Disease-Related Hepatocellular Carcinoma: A Problem of Growing Magnitude. Semin. Liver Dis. 2015, 35, 304–317. [Google Scholar] [CrossRef]
- Bertot, L.C.; Adams, L.A. Trends in hepatocellular carcinoma due to non-alcoholic fatty liver disease. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 179–187. [Google Scholar] [CrossRef]
- Liu, P.; Xie, S.H.; Hu, S.; Cheng, X.; Gao, T.; Zhang, C.; Song, Z. Age-specific sex difference in the incidence of hepatocellular carcinoma in the United States. Oncotarget 2017, 8, 68131–68137. [Google Scholar] [CrossRef] [Green Version]
- Xie, G.; Wang, X.; Zhao, A.; Yan, J.; Chen, W.; Jiang, R.; Ji, J.; Huang, F.; Zhang, Y.; Lei, S.; et al. Sex-dependent effects on gut microbiota regulate hepatic carcinogenic outcomes. Sci. Rep. 2017, 7, 45232. [Google Scholar] [CrossRef] [Green Version]
- Grimaudo, S.; Pipitone, R.M.; Pennisi, G.; Celsa, C.; Cammà, C.; Di Marco, V.; Barcellona, M.R.; Boemi, R.; Enea, M.; Giannetti, A.; et al. Association Between PNPLA3 rs738409 C > G Variant and Liver-Related Outcomes in Patients With Nonalcoholic Fatty Liver Disease. Clin. Gastroenterol. Hepatol. 2020, 18, 935–944.e3. [Google Scholar] [CrossRef]
- Liu, Y.L.; Patman, G.L.; Leathart, J.B.; Piguet, A.C.; Burt, A.D.; Dufour, J.F.; Day, C.P.; Daly, A.K.; Reeves, H.L.; Anstee, Q.M. Carriage of the PNPLA3 rs738409 C >G polymorphism confers an increased risk of non-alcoholic fatty liver disease associated hepatocellular carcinoma. J. Hepatol. 2014, 61, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Z.; Xia, H.H.; Xin, Y.N.; Lin, Z.H.; Xuan, S.Y. TM6SF2 E167K Variant, a Novel Genetic Susceptibility Variant, Contributing to Nonalcoholic Fatty Liver Disease. J. Clin. Transl. Hepatol. 2015, 3, 265–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldafashi, N.; Darlay, R.; Shukla, R.; McCain, M.V.; Watson, R.; Liu, Y.L.; McStraw, N.; Fathy, M.; Fawzy, M.A.; Zaki, M.Y.W.; et al. A PDCD1 Role in the Genetic Predisposition to NAFLD-HCC? Cancers 2021, 13, 1412. [Google Scholar] [CrossRef] [PubMed]
- Salmaninejad, A.; Khoramshahi, V.; Azani, A.; Soltaninejad, E.; Aslani, S.; Zamani, M.R.; Zal, M.; Nesaei, A.; Hosseini, S.M. PD-1 and cancer: Molecular mechanisms and polymorphisms. Immunogenetics 2018, 70, 73–86. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Petta, S.; Maglio, C.; Fracanzani, A.L.; Pipitone, R.; Mozzi, E.; Motta, B.M.; Kaminska, D.; Rametta, R.; Grimaudo, S.; et al. Transmembrane 6 superfamily member 2 gene variant disentangles nonalcoholic steatohepatitis from cardiovascular disease. Hepatology 2015, 61, 506–514. [Google Scholar] [CrossRef]
- Sookoian, S.; Castaño, G.O.; Scian, R.; Mallardi, P.; Fernández Gianotti, T.; Burgueño, A.L.; San Martino, J.; Pirola, C.J. Genetic variation in transmembrane 6 superfamily member 2 and the risk of nonalcoholic fatty liver disease and histological disease severity. Hepatology 2015, 61, 515–525. [Google Scholar] [CrossRef]
- Tang, S.; Zhang, J.; Mei, T.T.; Guo, H.Q.; Wei, X.H.; Zhang, W.Y.; Liu, Y.L.; Liang, S.; Fan, Z.P.; Ma, L.X.; et al. Association of TM6SF2 rs58542926 T/C gene polymorphism with hepatocellular carcinoma: A meta-analysis. BMC Cancer 2019, 19, 1128. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Lu, L.; Miao, Y.; Jin, W.; Li, C.; Xin, Y.; Xuan, S. E167K polymorphism of TM6SF2 gene affects cell cycle of hepatocellular carcinoma cell HEPA 1–6. Lipids Health Dis. 2017, 16, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, S.; Liao, S.; Liu, S.; Xin, Y. TM6SF2 E167K Variant Overexpression Promotes Expression of Inflammatory Cytokines in the HCC Cell Line HEPA 1–6. J. Clin. Transl. Hepatol. 2019, 7, 27–31. [Google Scholar] [CrossRef]
- Meroni, M.; Longo, M.; Paolini, E.; Alisi, A.; Miele, L.; De Caro, E.R.; Pisano, G.; Maggioni, M.; Soardo, G.; Valenti, L.V.; et al. The rs599839 A>G Variant Disentangles Cardiovascular Risk and Hepatocellular Carcinoma in NAFLD Patients. Cancers 2021, 13, 1783. [Google Scholar] [CrossRef]
- Mancina, R.M.; Dongiovanni, P.; Petta, S.; Pingitore, P.; Meroni, M.; Rametta, R.; Borén, J.; Montalcini, T.; Pujia, A.; Wiklund, O.; et al. The MBOAT7-TMC4 Variant rs641738 Increases Risk of Nonalcoholic Fatty Liver Disease in Individuals of European Descent. Gastroenterology 2016, 150, 1219–1230.e6. [Google Scholar] [CrossRef] [Green Version]
- Donati, B.; Dongiovanni, P.; Romeo, S.; Meroni, M.; McCain, M.; Miele, L.; Petta, S.; Maier, S.; Rosso, C.; De Luca, L.; et al. MBOAT7 rs641738 variant and hepatocellular carcinoma in non-cirrhotic individuals. Sci. Rep. 2017, 7, 4492. [Google Scholar] [CrossRef] [Green Version]
- Sookoian, S.; Flichman, D.; Garaycoechea, M.E.; Gazzi, C.; Martino, J.S.; Castaño, G.O.; Pirola, C.J. Lack of evidence supporting a role of TMC4-rs641738 missense variant-MBOAT7-intergenic downstream variant-in the Susceptibility to Nonalcoholic Fatty Liver Disease. Sci. Rep. 2018, 8, 5097. [Google Scholar] [CrossRef] [Green Version]
- Meroni, M.; Longo, M.; Fracanzani, A.L.; Dongiovanni, P. MBOAT7 down-regulation by genetic and environmental factors predisposes to MAFLD. EBioMedicine 2020, 57, 102866. [Google Scholar] [CrossRef]
- Ammerpohl, O.; Pratschke, J.; Schafmayer, C.; Haake, A.; Faber, W.; von Kampen, O.; Brosch, M.; Sipos, B.; von Schönfels, W.; Balschun, K.; et al. Distinct DNA methylation patterns in cirrhotic liver and hepatocellular carcinoma. Int. J. Cancer 2012, 130, 1319–1328. [Google Scholar] [CrossRef]
- Araújo, O.C.; Rosa, A.S.; Fernandes, A.; Niel, C.; Villela-Nogueira, C.A.; Pannain, V.; Araujo, N.M. RASSF1A and DOK1 Promoter Methylation Levels in Hepatocellular Carcinoma, Cirrhotic and Non-Cirrhotic Liver, and Correlation with Liver Cancer in Brazilian Patients. PLoS ONE 2016, 11, e0153796. [Google Scholar] [CrossRef] [PubMed]
- Zeybel, M.; Hardy, T.; Robinson, S.M.; Fox, C.; Anstee, Q.M.; Ness, T.; Masson, S.; Mathers, J.C.; French, J.; White, S.; et al. Differential DNA methylation of genes involved in fibrosis progression in non-alcoholic fatty liver disease and alcoholic liver disease. Clin. Epigenetics 2015, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajan, P.K.; Udoh, U.A.; Sanabria, J.D.; Banerjee, M.; Smith, G.; Schade, M.S.; Sanabria, J.; Sodhi, K.; Pierre, S.; Xie, Z.; et al. The Role of Histone Acetylation-/Methylation-Mediated Apoptotic Gene Regulation in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2020, 21, 8894. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhao, D.; Xu, Y.; Lin, J.; Xu, J.; Wang, K.; Ye, Z.; Luo, Y.; Liu, S.; Yang, H. LncRNA-Gm9795 promotes inflammation in non-alcoholic steatohepatitis via NF-κB/JNK pathway by endoplasmic reticulum stress. J. Transl. Med. 2021, 19, 101. [Google Scholar] [CrossRef]
- Wang, B.; Li, X.; Hu, W.; Zhou, Y.; Din, Y. Silencing of lncRNA SNHG20 delays the progression of nonalcoholic fatty liver disease to hepatocellular carcinoma via regulating liver Kupffer cells polarization. IUBMB Life 2019, 71, 1952–1961. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Zhang, J.X.; Chang, Q.M.; Wu, X.B.; Tang, W.G.; Wang, J.F.; Feng, J.F.; Zhang, Z.P.; Hu, Z.Q. LncRNA MYLK-AS1 facilitates tumor progression and angiogenesis by targeting miR-424-5p/E2F7 axis and activating VEGFR-2 signaling pathway in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 235. [Google Scholar] [CrossRef]
- Wu, H.; Ng, R.; Chen, X.; Steer, C.J.; Song, G. MicroRNA-21 is a potential link between non-alcoholic fatty liver disease and hepatocellular carcinoma via modulation of the HBP1-p53-Srebp1c pathway. Gut 2016, 65, 1850–1860. [Google Scholar] [CrossRef] [Green Version]
- Loyer, X.; Paradis, V.; Hénique, C.; Vion, A.C.; Colnot, N.; Guerin, C.L.; Devue, C.; On, S.; Scetbun, J.; Romain, M.; et al. Liver microRNA-21 is overexpressed in non-alcoholic steatohepatitis and contributes to the disease in experimental models by inhibiting PPARα expression. Gut 2016, 65, 1882–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.Y.; Yeh, K.Y.; Lin, C.Y.; Hsieh, Y.W.; Lai, H.H.; Chen, J.R.; Hsu, C.C.; Her, G.M. MicroRNA-21 Plays Multiple Oncometabolic Roles in the Process of NAFLD-Related Hepatocellular Carcinoma via PI3K/AKT, TGF-β, and STAT3 Signaling. Cancers 2021, 13, 940. [Google Scholar] [CrossRef]
- Gramantieri, L.; Ferracin, M.; Fornari, F.; Veronese, A.; Sabbioni, S.; Liu, C.G.; Calin, G.A.; Giovannini, C.; Ferrazzi, E.; Grazi, G.L.; et al. Cyclin G1 is a target of miR-122a, a microRNA frequently down-regulated in human hepatocellular carcinoma. Cancer Res. 2007, 67, 6092–6099. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.C.; Hsu, S.D.; Hsu, C.S.; Lai, T.C.; Chen, S.J.; Shen, R.; Huang, Y.; Chen, H.C.; Lee, C.H.; Tsai, T.F.; et al. MicroRNA-122 plays a critical role in liver homeostasis and hepatocarcinogenesis. J. Clin. Invest. 2012, 122, 2884–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Hwang, S.; Cai, Y.; Kim, S.J.; Xu, M.; Yang, D.; Guillot, A.; Feng, D.; Seo, W.; Hou, X.; et al. MicroRNA-223 Ameliorates Nonalcoholic Steatohepatitis and Cancer by Targeting Multiple Inflammatory and Oncogenic Genes in Hepatocytes. Hepatology 2019, 70, 1150–1167. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Ni, X.X.; Xu, Q.Y.; Wang, Q.; Li, X.Y.; Hua, J. Regulation of lipid-induced macrophage polarization through modulating peroxisome proliferator-activated receptor-gamma activity affects hepatic lipid metabolism via a Toll-like receptor 4/NF-κB signaling pathway. J. Gastroenterol. Hepatol. 2020, 35, 1998–2008. [Google Scholar] [CrossRef]
- Chen, M.; Wei, L.; Law, C.T.; Tsang, F.H.; Shen, J.; Cheng, C.L.; Tsang, L.H.; Ho, D.W.; Chiu, D.K.; Lee, J.M.; et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology 2018, 67, 2254–2270. [Google Scholar] [CrossRef]
- Gonzalez-Romero, F.; Mestre, D.; Aurrekoetxea, I.; O’Rourke, C.J.; Andersen, J.B.; Woodhoo, A.; Tamayo-Caro, M.; Varela Rey, M.; Palomo-Irigoyen, M.; Gómez-Santos, B.; et al. E2F1 and E2F2-mediated repression of CPT2 establishes a lipid-rich tumor-promoting environment. Cancer Res. 2021. [Google Scholar] [CrossRef]
- Fujiwara, N.; Nakagawa, H.; Enooku, K.; Kudo, Y.; Hayata, Y.; Nakatsuka, T.; Tanaka, Y.; Tateishi, R.; Hikiba, Y.; Misumi, K.; et al. CPT2 downregulation adapts HCC to lipid-rich environment and promotes carcinogenesis via acylcarnitine accumulation in obesity. Gut 2018, 67, 1493–1504. [Google Scholar] [CrossRef]
- He, Y.; Huang, S.; Cheng, T.; Wang, Y.; Zhou, S.J.; Zhang, Y.M.; Yu, P. High glucose may promote the proliferation and metastasis of hepatocellular carcinoma via E2F1/RRBP1 pathway. Life Sci. 2020, 252, 117656. [Google Scholar] [CrossRef]
- Sekine, K.; Chen, Y.R.; Kojima, N.; Ogata, K.; Fukamizu, A.; Miyajima, A. Foxo1 links insulin signaling to C/EBP alpha and regulates gluconeogenesis during liver development. Embo J. 2007, 26, 3607–3615. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhu, X.; Sun, X.; Yang, X.; Chang, X.; Xia, M.; Lu, Y.; Xia, P.; Yan, H.; Bian, H.; et al. FoxO3 regulates hepatic triglyceride metabolism via modulation of the expression of sterol regulatory-element binding protein 1c. Lipids Health Dis. 2019, 18, 197. [Google Scholar] [CrossRef] [Green Version]
- Lou, K.; Chen, N.; Li, Z.; Zhang, B.; Wang, X.; Chen, Y.; Xu, H.; Wang, D.; Wang, H. MicroRNA-142-5p Overexpression Inhibits Cell Growth and Induces Apoptosis by Regulating FOXO in Hepatocellular Carcinoma Cells. Oncol. Res. 2017, 25, 65–73. [Google Scholar] [CrossRef]
- Pan, X.; Zhang, Y.; Kim, H.G.; Liangpunsakul, S.; Dong, X.C. FOXO transcription factors protect against the diet-induced fatty liver disease. Sci. Rep. 2017, 7, 44597. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Riopel, M.; Cabrales, P.; Bandyopadhyay, G.K. Hepatocyte-specific HIF-1α ablation improves obesity-induced glucose intolerance by reducing first-pass GLP-1 degradation. Sci. Adv. 2019, 5, eaaw4176. [Google Scholar] [CrossRef] [Green Version]
- Vanderborght, B.; De Muynck, K.; Lefere, S.; Geerts, A.; Degroote, H.; Verhelst, X.; Van Vlierberghe, H.; Devisscher, L. Effect of isoform-specific HIF-1α and HIF-2α antisense oligonucleotides on tumorigenesis, inflammation and fibrosis in a hepatocellular carcinoma mouse model. Oncotarget 2020, 11, 4504–4520. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, J.; Huang, J.; Li, Z.; Gong, Y.; Zou, B.; Liu, X.; Ding, L.; Li, P.; Zhu, Z.; et al. HIF-2α upregulation mediated by hypoxia promotes NAFLD-HCC progression by activating lipid synthesis via the PI3K-AKT-mTOR pathway. Aging 2019, 11, 10839–10860. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, M.E.; Castellaro, A.; Racca, A.C.; Carbajosa González, S.; Pansa, M.F.; Soria, G.; Bocco, J.L. Krüppel-Like Factor 6 Is Required for Oxidative and Oncogene-Induced Cellular Senescence. Front. Cell Dev. Biol. 2019, 7, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechmann, L.P.; Vetter, D.; Ishida, J.; Hannivoort, R.A.; Lang, U.E.; Kocabayoglu, P.; Fiel, M.I.; Muñoz, U.; Patman, G.L.; Ge, F.; et al. Post-transcriptional activation of PPAR alpha by KLF6 in hepatic steatosis. J. Hepatol. 2013, 58, 1000–1006. [Google Scholar] [CrossRef] [Green Version]
- Bechmann, L.P.; Gastaldelli, A.; Vetter, D.; Patman, G.L.; Pascoe, L.; Hannivoort, R.A.; Lee, U.E.; Fiel, I.; Muñoz, U.; Ciociaro, D.; et al. Glucokinase links Krüppel-like factor 6 to the regulation of hepatic insulin sensitivity in nonalcoholic fatty liver disease. Hepatology 2012, 55, 1083–1093. [Google Scholar] [CrossRef]
- Miele, L.; Beale, G.; Patman, G.; Nobili, V.; Leathart, J.; Grieco, A.; Abate, M.; Friedman, S.L.; Narla, G.; Bugianesi, E.; et al. The Kruppel-like factor 6 genotype is associated with fibrosis in nonalcoholic fatty liver disease. Gastroenterology 2008, 135, 282–291.e1. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.P.; Zhou, H.J.; Chen, X.P.; Ren, G.Y.; Ruan, X.X.; Zhang, Y.; Zhang, R.L.; Chen, J. Loss of expression of Kruppel-like factor 6 in primary hepatocellular carcinoma and hepatoma cell lines. J. Exp. Clin. Cancer Res. 2007, 26, 117–124. [Google Scholar] [PubMed]
- He, A.D.; Xie, W.; Song, W.; Ma, Y.Y.; Liu, G.; Liang, M.L.; Da, X.W.; Yao, G.Q.; Zhang, B.X.; Gao, C.J.; et al. Platelet releasates promote the proliferation of hepatocellular carcinoma cells by suppressing the expression of KLF6. Sci. Rep. 2017, 7, 3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narla, G.; Kremer-Tal, S.; Matsumoto, N.; Zhao, X.; Yao, S.; Kelley, K.; Tarocchi, M.; Friedman, S.L. In vivo regulation of p21 by the Kruppel-like factor 6 tumor-suppressor gene in mouse liver and human hepatocellular carcinoma. Oncogene 2007, 26, 4428–4434. [Google Scholar] [CrossRef] [Green Version]
- Jordan, S.; Tung, N.; Casanova-Acebes, M.; Chang, C.; Cantoni, C.; Zhang, D.; Wirtz, T.H.; Naik, S.; Rose, S.A.; Brocker, C.N.; et al. Dietary Intake Regulates the Circulating Inflammatory Monocyte Pool. Cell 2019, 178, 1102–1114.e17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nakajima, T.; Gonzalez, F.J.; Tanaka, N. PPARs as Metabolic Regulators in the Liver: Lessons from Liver-Specific PPAR-Null Mice. Int. J. Mol. Sci. 2020, 21, 2061. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Dai, W.; Mao, Y.; Wu, L.; Li, J.; Chen, K.; Yu, Q.; Kong, R.; Li, S.; Zhang, J.; et al. Simvastatin re-sensitizes hepatocellular carcinoma cells to sorafenib by inhibiting HIF-1α/PPAR-γ/PKM2-mediated glycolysis. J. Exp. Clin. Cancer Res. 2020, 39, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.; Jeong, I.K.; Ahn, K.J.; Chung, H.Y.; Hwang, Y.C. Fenofibrate, a PPARα agonist, reduces hepatic fat accumulation through the upregulation of TFEB-mediated lipophagy. Metabolism 2021, 120, 154798. [Google Scholar] [CrossRef]
- Zarei, M.; Barroso, E.; Palomer, X.; Dai, J.; Rada, P.; Quesada-López, T.; Escolà-Gil, J.C.; Cedó, L.; Zali, M.R.; Molaei, M.; et al. Hepatic regulation of VLDL receptor by PPARβ/δ and FGF21 modulates non-alcoholic fatty liver disease. Mol. Metab. 2018, 8, 117–131. [Google Scholar] [CrossRef]
- Romano, A.; Friuli, M.; Del Coco, L.; Longo, S.; Vergara, D.; Del Boccio, P.; Valentinuzzi, S.; Cicalini, I.; Fanizzi, F.P.; Gaetani, S.; et al. Chronic Oleoylethanolamide Treatment Decreases Hepatic Triacylglycerol Level in Rat Liver by a PPARγ/SREBP-Mediated Suppression of Fatty Acid and Triacylglycerol Synthesis. Nutrients 2021, 13, 394. [Google Scholar] [CrossRef] [PubMed]
- Montagner, A.; Polizzi, A.; Fouché, E.; Ducheix, S.; Lippi, Y.; Lasserre, F.; Barquissau, V.; Régnier, M.; Lukowicz, C.; Benhamed, F.; et al. Liver PPARα is crucial for whole-body fatty acid homeostasis and is protective against NAFLD. Gut 2016, 65, 1202–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manieri, E.; Folgueira, C.; Rodríguez, M.E.; Leiva-Vega, L.; Esteban-Lafuente, L.; Chen, C.; Cubero, F.J.; Barrett, T.; Cavanagh-Kyros, J.; Seruggia, D.; et al. JNK-mediated disruption of bile acid homeostasis promotes intrahepatic cholangiocarcinoma. Proc. Natl. Acad. Sci. USA 2020, 117, 16492–16499. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Wang, H.; Wang, Y.; Sun, R.; Xie, Y.; Zhou, Z.; Wang, H.; Aa, J.; Zhou, F.; Wang, G. Apatinib induces 3-hydroxybutyric acid production in the liver of mice by peroxisome proliferator-activated receptor α activation to aid its antitumor effect. Cancer Sci. 2019, 110, 3328–3339. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Hatano, B.; Zhao, M.; Yen, C.C.; Kang, K.; Reilly, S.M.; Gangl, M.R.; Gorgun, C.; Balschi, J.A.; Ntambi, J.M.; et al. Role of peroxisome proliferator-activated receptor {delta}/{beta} in hepatic metabolic regulation. J. Biol. Chem. 2011, 286, 1237–1247. [Google Scholar] [CrossRef] [Green Version]
- Gavrilova, O.; Haluzik, M.; Matsusue, K.; Cutson, J.J.; Johnson, L.; Dietz, K.R.; Nicol, C.J.; Vinson, C.; Gonzalez, F.J.; Reitman, M.L. Liver peroxisome proliferator-activated receptor gamma contributes to hepatic steatosis, triglyceride clearance, and regulation of body fat mass. J. Biol. Chem. 2003, 278, 34268–34276. [Google Scholar] [CrossRef] [Green Version]
- Gastaldelli, A.; Sabatini, S.; Carli, F.; Gaggini, M.; Bril, F.; Belfort-DeAguiar, R.; Positano, V.; Barb, D.; Kadiyala, S.; Harrison, S.; et al. PPAR-γ-induced Changes in Visceral Fat and Adiponectin Levels are Associated with Improvement of Steatohepatitis in Patients with NASH. Liver Int. 2021. [Google Scholar] [CrossRef] [PubMed]
- Barroso, E.; Rodríguez-Calvo, R.; Serrano-Marco, L.; Astudillo, A.M.; Balsinde, J.; Palomer, X.; Vázquez-Carrera, M. The PPARβ/δ activator GW501516 prevents the down-regulation of AMPK caused by a high-fat diet in liver and amplifies the PGC-1α-Lipin 1-PPARα pathway leading to increased fatty acid oxidation. Endocrinology 2011, 152, 1848–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Chi, Y.; Dong, Z.; Tao, T.; Zhang, X.; Pan, W.; Wang, Y. A nomogram combining PPARγ expression profiles and clinical factors predicts survival in patients with hepatocellular carcinoma. Oncol. Lett. 2021, 21, 319. [Google Scholar] [CrossRef]
- Yin, F.; Feng, F.; Wang, L.; Wang, X.; Li, Z.; Cao, Y. SREBP-1 inhibitor Betulin enhances the antitumor effect of Sorafenib on hepatocellular carcinoma via restricting cellular glycolytic activity. Cell Death Dis. 2019, 10, 672. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhou, Z.S.; Shen, Y.; Xu, J.; Miao, H.H.; Xiong, Y.; Xu, F.; Li, B.L.; Luo, J.; Song, B.L. Inhibition of the sterol regulatory element-binding protein pathway suppresses hepatocellular carcinoma by repressing inflammation in mice. Hepatology 2017, 65, 1936–1947. [Google Scholar] [CrossRef]
- Chen, J.; Ding, C.; Chen, Y.; Hu, W.; Yu, C.; Peng, C.; Feng, X.; Cheng, Q.; Wu, W.; Lu, Y.; et al. ACSL4 reprograms fatty acid metabolism in hepatocellular carcinoma via c-Myc/SREBP1 pathway. Cancer Lett. 2021, 502, 154–165. [Google Scholar] [CrossRef]
- Ma, A.P.Y.; Yeung, C.L.S.; Tey, S.K.; Mao, X.; Wong, S.W.K.; Ng, T.H.; Ko, F.C.F.; Kwong, E.M.L.; Tang, A.H.N.; Ng, I.O.; et al. Suppression of ACADM-mediated fatty acid oxidation promotes hepatocellular carcinoma via aberrant Cav1/SREBP-1 signaling. Cancer Res. 2021. [Google Scholar] [CrossRef]
- Yu, X.; Lin, Q.; Wu, Z.; Zhang, Y.; Wang, T.; Zhao, S.; Song, X.; Chen, C.; Wang, Z.; Xu, L.; et al. ZHX2 inhibits SREBP1c-mediated de novo lipogenesis in hepatocellular carcinoma via miR-24-3p. J. Pathol. 2020, 252, 358–370. [Google Scholar] [CrossRef]
- Kumar, D.P.; Santhekadur, P.K.; Seneshaw, M.; Mirshahi, F.; Uram-Tuculescu, C.; Sanyal, A.J. A Regulatory Role of Apoptosis Antagonizing Transcription Factor in the Pathogenesis of Nonalcoholic Fatty Liver Disease and Hepatocellular Carcinoma. Hepatology 2019, 69, 1520–1534. [Google Scholar] [CrossRef]
- Liu, J.; Lu, J.; Ma, Z.; Li, W. A Nomogram Based on a Three-Gene Signature Derived from AATF Coexpressed Genes Predicts Overall Survival of Hepatocellular Carcinoma Patients. Biomed. Res. Int. 2020, 2020, 7310768. [Google Scholar] [CrossRef] [PubMed]
- Benhamed, F.; Denechaud, P.D.; Lemoine, M.; Robichon, C.; Moldes, M.; Bertrand-Michel, J.; Ratziu, V.; Serfaty, L.; Housset, C.; Capeau, J.; et al. The lipogenic transcription factor ChREBP dissociates hepatic steatosis from insulin resistance in mice and humans. J. Clin. Invest. 2012, 122, 2176–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Y.; Hu, Q.; Gu, J. Expressions of Carbohydrate Response Element Binding Protein and Glucose Transporters in Liver Cancer and Clinical Significance. Pathol. Oncol. Res. 2020, 26, 1331–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Lekbaby, B.; Fares, N.; Augustin, J.; Attout, T.; Schnuriger, A.; Cassard, A.M.; Panasyuk, G.; Perlemuter, G.; Bieche, I.; et al. Alteration of splicing factors’ expression during liver disease progression: Impact on hepatocellular carcinoma outcome. Hepatol. Int. 2019, 13, 454–467. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Won, R.; Ban, G.; Ju, M.H.; Cho, K.S.; Young Han, S.; Jeong, J.S.; Lee, S.W. Targeted Regression of Hepatocellular Carcinoma by Cancer-Specific RNA Replacement through MicroRNA Regulation. Sci. Rep. 2015, 5, 12315. [Google Scholar] [CrossRef] [Green Version]
- Han, S.R.; Lee, C.H.; Im, J.Y.; Kim, J.H.; Kim, J.H.; Kim, S.J.; Cho, Y.W.; Kim, E.; Kim, Y.; Ryu, J.H.; et al. Targeted suicide gene therapy for liver cancer based on ribozyme-mediated RNA replacement through post-transcriptional regulation. Mol. Ther. Nucleic Acids 2021, 23, 154–168. [Google Scholar] [CrossRef]
- Del Río-Moreno, M.; Alors-Pérez, E.; González-Rubio, S.; Ferrín, G.; Reyes, O.; Rodríguez-Perálvarez, M.; Sánchez-Frías, M.E.; Sánchez-Sánchez, R.; Ventura, S.; López-Miranda, J.; et al. Dysregulation of the Splicing Machinery Is Associated to the Development of Nonalcoholic Fatty Liver Disease. J. Clin. Endocrinol. Metab. 2019, 104, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.; Wang, Y.; Bedford, M.T. The Role of the PRMT5-SND1 Axis in Hepatocellular Carcinoma. Epigenomes 2021, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Pihlajamäki, J.; Lerin, C.; Itkonen, P.; Boes, T.; Floss, T.; Schroeder, J.; Dearie, F.; Crunkhorn, S.; Burak, F.; Jimenez-Chillaron, J.C.; et al. Expression of the splicing factor gene SFRS10 is reduced in human obesity and contributes to enhanced lipogenesis. Cell Metab. 2011, 14, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Sun, Y.; Liu, W.; Dong, J.; Chen, J. SIRT1 mediates the role of RNA-binding protein QKI 5 in the synthesis of triglycerides in non-alcoholic fatty liver disease mice via the PPARα/FoxO1 signaling pathway. Int. J. Mol. Med. 2019, 43, 1271–1280. [Google Scholar] [CrossRef]
- Li, X.; Yuan, B.; Yao, Z.; Sun, X.; Guo, W.; Chen, Z. The RNA-binding protein HuR promotes nonalcoholic steatohepatitis (NASH) progression by enhancing death signaling pathway. bioRxiv 2020. [Google Scholar] [CrossRef]
- Yang, S.; Lin, S.; Liu, K.; Liu, Y.; Xu, P.; Zheng, Y.; Deng, Y.; Zhang, D.; Zhai, Z.; Li, N.; et al. Identification of an immune-related RNA-binding protein signature to predict survival and targeted therapy responses in liver cancer. Genomics 2021, 113, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Z.; Li, Y.; Wan, Y.; Xing, B. Integrated bioinformatic analysis of RNA binding proteins in hepatocellular carcinoma. Aging 2020, 13, 2480–2505. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, F.; Wei, K.; Wang, J.; Zhou, G.; Wu, C.; Yin, G. Development and Validation of a RNA Binding Protein-Associated Prognostic Model for Hepatocellular Carcinoma. Technol. Cancer Res. Treat. 2021, 20. [Google Scholar] [CrossRef]
- Lee, G.; Jeong, Y.S.; Kim, D.W.; Kwak, M.J.; Koh, J.; Joo, E.W.; Lee, J.S.; Kah, S.; Sim, Y.E.; Yim, S.Y. Clinical significance of APOB inactivation in hepatocellular carcinoma. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yao, M.; Wen, X.; Zhu, Y.; Zhao, E.; Qian, X.; Chen, X.; Lu, W.; Lv, Q.; Zhang, L.; et al. Elevated apolipoprotein B predicts poor postsurgery prognosis in patients with hepatocellular carcinoma. Onco Targets Ther. 2019, 12, 1957–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, V.; Riordan, J.D.; Soleymanjahi, S.; Nadeau, J.H.; Nalbantoglu, I.; Xie, Y.; Molitor, E.A.; Madison, B.B.; Brunt, E.M.; Mills, J.C.; et al. Apobec1 complementation factor overexpression promotes hepatic steatosis, fibrosis, and hepatocellular cancer. J. Clin. Invest. 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Kesarwala, A.H.; Eggert, T.; Medina-Echeverz, J.; Kleiner, D.E.; Jin, P.; Stroncek, D.F.; Terabe, M.; Kapoor, V.; ElGindi, M.; et al. NAFLD causes selective CD4(+) T lymphocyte loss and promotes hepatocarcinogenesis. Nature 2016, 531, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, B.; Brown, Z.J.; Diggs, L.P.; Vormehr, M.; Ma, C.; Subramanyam, V.; Rosato, U.; Ruf, B.; Walz, J.S.; McVey, J.C.; et al. Steatohepatitis Impairs T-cell-Directed Immunotherapies Against Liver Tumors in Mice. Gastroenterology 2021, 160, 331–345.e6. [Google Scholar] [CrossRef]
- Pfister, D.; Núñez, N.G.; Pinyol, R.; Govaere, O.; Pinter, M.; Szydlowska, M.; Gupta, R.; Qiu, M.; Deczkowska, A.; Weiner, A.; et al. NASH limits anti-tumour surveillance in immunotherapy-treated HCC. Nature 2021, 592, 450–456. [Google Scholar] [CrossRef]
- Ma, J.; Zheng, B.; Goswami, S.; Meng, L.; Zhang, D.; Cao, C.; Li, T.; Zhu, F.; Ma, L.; Zhang, Z.; et al. PD1(Hi) CD8(+) T cells correlate with exhausted signature and poor clinical outcome in hepatocellular carcinoma. J. Immunother. Cancer 2019, 7, 331. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.D.; Song, G.W.; Park, S.; Jung, M.K.; Kim, M.H.; Kang, H.J.; Yoo, C.; Yi, K.; Kim, K.H.; Eo, S.; et al. Association Between Expression Level of PD1 by Tumor-Infiltrating CD8(+) T Cells and Features of Hepatocellular Carcinoma. Gastroenterology 2018, 155, 1936–1950.e17. [Google Scholar] [CrossRef]
- Ye, Q.; Ling, S.; Zheng, S.; Xu, X. Liquid biopsy in hepatocellular carcinoma: Circulating tumor cells and circulating tumor DNA. Mol. Cancer 2019, 18, 114. [Google Scholar] [CrossRef]
- Paradis, V.; Zalinski, S.; Chelbi, E.; Guedj, N.; Degos, F.; Vilgrain, V.; Bedossa, P.; Belghiti, J. Hepatocellular carcinomas in patients with metabolic syndrome often develop without significant liver fibrosis: A pathological analysis. Hepatology 2009, 49, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Perumpail, R.B.; Wong, R.J.; Ahmed, A.; Harrison, S.A. Hepatocellular Carcinoma in the Setting of Non-cirrhotic Nonalcoholic Fatty Liver Disease and the Metabolic Syndrome: US Experience. Dig. Dis. Sci. 2015, 60, 3142–3148. [Google Scholar] [CrossRef] [PubMed]
- Osho, A.; Rich, N.E.; Singal, A.G. Role of imaging in management of hepatocellular carcinoma: Surveillance, diagnosis, and treatment response. Hepatoma Res. 2020, 6. [Google Scholar] [CrossRef]
- Hennedige, T.; Venkatesh, S.K. Imaging of hepatocellular carcinoma: Diagnosis, staging and treatment monitoring. Cancer Imaging 2013, 12, 530–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masokano, I.B.; Liu, W.; Xie, S.; Marcellin, D.F.H.; Pei, Y.; Li, W. The application of texture quantification in hepatocellular carcinoma using CT and MRI: A review of perspectives and challenges. Cancer Imaging 2020, 20, 67. [Google Scholar] [CrossRef] [PubMed]
- Su, T.H.; Wu, C.H.; Kao, J.H. Artificial intelligence in precision medicine in hepatology. J. Gastroenterol. Hepatol. 2021, 36, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Spieler, B.; Sabottke, C.; Moawad, A.W.; Gabr, A.M.; Bashir, M.R.; Do, R.K.G.; Yaghmai, V.; Rozenberg, R.; Gerena, M.; Yacoub, J.; et al. Artificial intelligence in assessment of hepatocellular carcinoma treatment response. Abdom. Radiol. 2021. [Google Scholar] [CrossRef]
- Lupsor-Platon, M.; Serban, T.; Silion, A.I.; Tirpe, G.R.; Tirpe, A.; Florea, M. Performance of Ultrasound Techniques and the Potential of Artificial Intelligence in the Evaluation of Hepatocellular Carcinoma and Non-Alcoholic Fatty Liver Disease. Cancers 2021, 13, 790. [Google Scholar] [CrossRef] [PubMed]
- Bianco, C.; Jamialahmadi, O.; Pelusi, S.; Baselli, G.; Dongiovanni, P.; Zanoni, I.; Santoro, L.; Maier, S.; Liguori, A.; Meroni, M.; et al. Non-invasive stratification of hepatocellular carcinoma risk in non-alcoholic fatty liver using polygenic risk scores. J. Hepatol. 2021, 74, 775–782. [Google Scholar] [CrossRef]
- Gellert-Kristensen, H.; Richardson, T.G.; Davey Smith, G.; Nordestgaard, B.G.; Tybjaerg-Hansen, A.; Stender, S. Combined Effect of PNPLA3, TM6SF2, and HSD17B13 Variants on Risk of Cirrhosis and Hepatocellular Carcinoma in the General Population. Hepatology 2020, 72, 845–856. [Google Scholar] [CrossRef]
- Nakamura, N.; Hatano, E.; Iguchi, K.; Sato, M.; Kawaguchi, H.; Ohtsu, I.; Sakurai, T.; Aizawa, N.; Iijima, H.; Nishiguchi, S.; et al. Elevated levels of circulating ITIH4 are associated with hepatocellular carcinoma with nonalcoholic fatty liver disease: From pig model to human study. BMC Cancer 2019, 19, 621. [Google Scholar] [CrossRef]
- Jampoka, K.; Muangpaisarn, P.; Khongnomnan, K.; Treeprasertsuk, S.; Tangkijvanich, P.; Payungporn, S. Serum miR-29a and miR-122 as Potential Biomarkers for Non-Alcoholic Fatty Liver Disease (NAFLD). Microrna 2018, 7, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zheng, Y.; Xiao, X.; Liu, C.; Lin, J.; Zheng, S.; Yang, B.; Ou, Q. A Circulating Long Noncoding RNA Panel Serves as a Diagnostic Marker for Hepatocellular Carcinoma. Dis. Markers 2020, 2020, 5417598. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Gao, S.; Zhu, M.; Luo, P.; Jing, X.; Chai, H.; Tu, J. Potential diagnostic value of lncRNA SPRY4-IT1 in hepatocellular carcinoma. Oncol. Rep. 2016, 36, 1085–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, J.; Bilgi, H.; Heider, D.; Schotten, C.; Manka, P.; Bedreli, S.; Gorray, M.; Ertle, J.; van Grunsven, L.A.; Dechêne, A. The GALAD scoring algorithm based on AFP, AFP-L3, and DCP significantly improves detection of BCLC early stage hepatocellular carcinoma. Z. Gastroenterol. 2016, 54, 1296–1305. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Wu, R.; Liu, X.; Xu, H.; Chi, X.; Wang, X.; Zhan, M.; Wang, B.; Peng, F.; Gao, X.; et al. Validation of the GALAD Model and Establishment of GAAP Model for Diagnosis of Hepatocellular Carcinoma in Chinese Patients. J. Hepatocell. Carcinoma 2020, 7, 219–232. [Google Scholar] [CrossRef]
- Singal, A.G.; Tayob, N.; Mehta, A.; Marrero, J.A.; Jin, Q.; Lau, J.; Parikh, N.D. Doylestown Plus and GALAD Demonstrate High Sensitivity for HCC Detection in Patients with Cirrhosis. Clin. Gastroenterol. Hepatol. 2021. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, Y.; Yang, G.; He, S.; Qiu, X.; Zhang, L.; Deng, Q.; Zheng, F. Using circular RNA SMARCA5 as a potential novel biomarker for hepatocellular carcinoma. Clin. Chim. Acta 2019, 492, 37–44. [Google Scholar] [CrossRef]
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef]
- Chhimwal, J.; Patial, V.; Padwad, Y. Beverages and Non-alcoholic fatty liver disease (NAFLD): Think before you drink. Clin. Nutr. 2021, 40, 2508–2519. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.V.; Zandvakili, I.; Thaiss, C.A.; Schneider, K.M. Physical activity is associated with reduced risk of liver disease in the prospective UK Biobank cohort. JHEP Rep. 2021, 3, 100263. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, S.E.; Schlesinger, S.; Aleksandrova, K.; Jochem, C.; Jenab, M.; Gunter, M.J.; Overvad, K.; Tjønneland, A.; Boutron-Ruault, M.C.; Carbonnel, F.; et al. Association between physical activity and risk of hepatobiliary cancers: A multinational cohort study. J. Hepatol. 2019, 70, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Flockhart, M.; Nilsson, L.C.; Tais, S.; Ekblom, B.; Apró, W.; Larsen, F.J. Excessive exercise training causes mitochondrial functional impairment and decreases glucose tolerance in healthy volunteers. Cell Metab. 2021, 33, 957–970. [Google Scholar] [CrossRef]
- Kane, R.C.; Farrell, A.T.; Madabushi, R.; Booth, B.; Chattopadhyay, S.; Sridhara, R.; Justice, R.; Pazdur, R. Sorafenib for the treatment of unresectable hepatocellular carcinoma. Oncologist 2009, 14, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Kane, R.C.; Farrell, A.T.; Saber, H.; Tang, S.; Williams, G.; Jee, J.M.; Liang, C.; Booth, B.; Chidambaram, N.; Morse, D.; et al. Sorafenib for the treatment of advanced renal cell carcinoma. Clin. Cancer Res. 2006, 12, 7271–7278. [Google Scholar] [CrossRef] [Green Version]
- Kudo, M.; Finn, R.S.; Qin, S.; Han, K.H.; Ikeda, K.; Piscaglia, F.; Baron, A.; Park, J.W.; Han, G.; Jassem, J.; et al. Lenvatinib versus sorafenib in first-line treatment of patients with unresectable hepatocellular carcinoma: A randomised phase 3 non-inferiority trial. Lancet 2018, 391, 1163–1173. [Google Scholar] [CrossRef] [Green Version]
- Casak, S.J.; Donoghue, M.; Fashoyin-Aje, L.; Jiang, X.; Rodriguez, L.; Shen, Y.L.; Xu, Y.; Jiang, X.; Liu, J.; Zhao, H.; et al. FDA Approval Summary: Atezolizumab Plus Bevacizumab for the Treatment of Patients with Advanced Unresectable or Metastatic Hepatocellular Carcinoma. Clin. Cancer Res. 2021, 27, 1836–1841. [Google Scholar] [CrossRef]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905. [Google Scholar] [CrossRef] [PubMed]
- Pelosof, L.; Lemery, S.; Casak, S.; Jiang, X.; Rodriguez, L.; Pierre, V.; Bi, Y.; Liu, J.; Zirkelbach, J.F.; Patel, A.; et al. Benefit-Risk Summary of Regorafenib for the Treatment of Patients with Advanced Hepatocellular Carcinoma That Has Progressed on Sorafenib. Oncologist 2018, 23, 496–500. [Google Scholar] [CrossRef] [Green Version]
- Abou-Alfa, G.K.; Meyer, T.; Cheng, A.L.; El-Khoueiry, A.B.; Rimassa, L.; Ryoo, B.Y.; Cicin, I.; Merle, P.; Chen, Y.; Park, J.W.; et al. Cabozantinib in Patients with Advanced and Progressing Hepatocellular Carcinoma. N. Engl. J. Med. 2018, 379, 54–63. [Google Scholar] [CrossRef]
- Yau, T.; Kang, Y.K.; Kim, T.Y.; El-Khoueiry, A.B.; Santoro, A.; Sangro, B.; Melero, I.; Kudo, M.; Hou, M.M.; Matilla, A.; et al. Efficacy and Safety of Nivolumab Plus Ipilimumab in Patients With Advanced Hepatocellular Carcinoma Previously Treated With Sorafenib: The CheckMate 040 Randomized Clinical Trial. JAMA Oncol. 2020, 6, e204564. [Google Scholar] [CrossRef] [PubMed]
- Foerster, F.; Galle, P.R. The Current Landscape of Clinical Trials for Systemic Treatment of HCC. Cancers 2021, 13, 1962. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A.G.; Lee, M.Y.; Hunter, R.; Ank, R.S.; Story, J.Y.; Talekar, G.; Sisroe, T.; Ballak, D.B.; Fedanov, A.; Porter, C.C.; et al. Interleukin-37 improves T-cell-mediated immunity and chimeric antigen receptor T-cell therapy in aged backgrounds. Aging Cell 2021, 20, e13309. [Google Scholar] [CrossRef]
- Ellebrecht, C.T.; Bhoj, V.G.; Nace, A.; Choi, E.J.; Mao, X.; Cho, M.J.; Di Zenzo, G.; Lanzavecchia, A.; Seykora, J.T.; Cotsarelis, G.; et al. Reengineering chimeric antigen receptor T cells for targeted therapy of autoimmune disease. Science 2016, 353, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Ho, J.Y.; Du, H.; Xuan, F.; Wu, X.; Wang, Q.; Wang, L.; Liu, Y.; Ba, M.; Wang, Y.; et al. Evidence of long-lasting anti-CD19 activity of engrafted CD19 chimeric antigen receptor-modified T cells in a phase I study targeting pediatrics with acute lymphoblastic leukemia. Hematol. Oncol. 2019, 37, 601–608. [Google Scholar] [CrossRef]
- Rapoport, A.P.; Stadtmauer, E.A.; Binder-Scholl, G.K.; Goloubeva, O.; Vogl, D.T.; Lacey, S.F.; Badros, A.Z.; Garfall, A.; Weiss, B.; Finklestein, J.; et al. NY-ESO-1-specific TCR-engineered T cells mediate sustained antigen-specific antitumor effects in myeloma. Nat. Med. 2015, 21, 914–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.A.; June, C.H. The Principles of Engineering Immune Cells to Treat Cancer. Cell 2017, 168, 724–740. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Shi, Y.; Kaseb, A.O.; Qi, X.; Zhang, Y.; Chi, J.; Lu, Q.; Gao, H.; Jiang, H.; Wang, H.; et al. Chimeric Antigen Receptor-Glypican-3 T-Cell Therapy for Advanced Hepatocellular Carcinoma: Results of Phase I Trials. Clin. Cancer Res. 2020, 26, 3979–3989. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Peng, Y.; Wang, L.; Hong, Y.; Jiang, X.; Li, Q.; Liu, H.; Huang, L.; Wu, J.; Celis, E.; et al. Identification of α-fetoprotein-specific T-cell receptors for hepatocellular carcinoma immunotherapy. Hepatology 2018, 68, 574–589. [Google Scholar] [CrossRef] [Green Version]
- Caviglia, G.P.; Armandi, A.; Rosso, C.; Gaia, S.; Aneli, S.; Rolle, E.; Abate, M.L.; Olivero, A.; Nicolosi, A.; Guariglia, M.; et al. Biomarkers of Oncogenesis, Adipose Tissue Dysfunction and Systemic Inflammation for the Detection of Hepatocellular Carcinoma in Patients with Nonalcoholic Fatty Liver Disease. Cancers 2021, 13, 2305. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.M.; Carter, C.; Tang, L.; Wilkie, D.; McNabola, A.; Rong, H.; Chen, C.; Zhang, X.; Vincent, P.; McHugh, M.; et al. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004, 64, 7099–7109. [Google Scholar] [CrossRef] [Green Version]
- Abou-Alfa, G.K.; Schwartz, L.; Ricci, S.; Amadori, D.; Santoro, A.; Figer, A.; De Greve, J.; Douillard, J.Y.; Lathia, C.; Schwartz, B.; et al. Phase II study of sorafenib in patients with advanced hepatocellular carcinoma. J. Clin. Oncol. 2006, 24, 4293–4300. [Google Scholar] [CrossRef] [Green Version]
- Mross, K.; Frost, A.; Steinbild, S.; Hedbom, S.; Büchert, M.; Fasol, U.; Unger, C.; Krätzschmar, J.; Heinig, R.; Boix, O.; et al. A phase I dose-escalation study of regorafenib (BAY 73-4506), an inhibitor of oncogenic, angiogenic, and stromal kinases, in patients with advanced solid tumors. Clin. Cancer Res. 2012, 18, 2658–2667. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Cabrera, R. Systemic Treatment of Patients with Advanced, Unresectable Hepatocellular Carcinoma: Emergence of Therapies. J. Gastrointest. Cancer 2018, 49, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Kudo, M.; Kawazoe, S.; Osaki, Y.; Ikeda, M.; Okusaka, T.; Tamai, T.; Suzuki, T.; Hisai, T.; Hayato, S.; et al. Phase 2 study of lenvatinib in patients with advanced hepatocellular carcinoma. J. Gastroenterol. 2017, 52, 512–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.S.; Ryoo, B.Y.; Merle, P.; Kudo, M.; Bouattour, M.; Lim, H.Y.; Breder, V.; Edeline, J.; Chao, Y.; Ogasawara, S.; et al. Pembrolizumab As Second-Line Therapy in Patients With Advanced Hepatocellular Carcinoma in KEYNOTE-240: A Randomized, Double-Blind, Phase III Trial. J. Clin. Oncol. 2020, 38, 193–202. [Google Scholar] [CrossRef]
- Kelley, R.K.; Verslype, C.; Cohn, A.L.; Yang, T.S.; Su, W.C.; Burris, H.; Braiteh, F.; Vogelzang, N.; Spira, A.; Foster, P.; et al. Cabozantinib in hepatocellular carcinoma: Results of a phase 2 placebo-controlled randomized discontinuation study. Ann. Oncol. 2017, 28, 528–534. [Google Scholar] [CrossRef]
- Zhu, A.X.; Kang, Y.K.; Yen, C.J.; Finn, R.S.; Galle, P.R.; Llovet, J.M.; Assenat, E.; Brandi, G.; Pracht, M.; Lim, H.Y.; et al. Ramucirumab after sorafenib in patients with advanced hepatocellular carcinoma and increased α-fetoprotein concentrations (REACH-2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Oncol. 2019, 20, 282–296. [Google Scholar] [CrossRef]
- Saung, M.T.; Pelosof, L.; Casak, S.; Donoghue, M.; Lemery, S.; Yuan, M.; Rodriguez, L.; Schotland, P.; Chuk, M.; Davis, G.; et al. FDA Approval Summary: Nivolumab plus ipilimumab for the treatment of patients with hepatocellular carcinoma previously treated with sorafenib. Oncologist 2021. [Google Scholar] [CrossRef] [PubMed]
- Pinato, D.J.; Cortellini, A.; Sukumaran, A.; Cole, T.; Pai, M.; Habib, N.; Spalding, D.; Sodergren, M.H.; Martinez, M.; Dhillon, T.; et al. PRIME-HCC: Phase Ib study of neoadjuvant ipilimumab and nivolumab prior to liver resection for hepatocellular carcinoma. BMC Cancer 2021, 21, 301. [Google Scholar] [CrossRef]
- Hack, S.P.; Spahn, J.; Chen, M.; Cheng, A.L.; Kaseb, A.; Kudo, M.; Lee, H.C.; Yopp, A.; Chow, P.; Qin, S. IMbrave 050: A Phase III trial of atezolizumab plus bevacizumab in high-risk hepatocellular carcinoma after curative resection or ablation. Future Oncol. 2020, 16, 975–989. [Google Scholar] [CrossRef]
- Mizukoshi, E.; Nakagawa, H.; Kitahara, M.; Yamashita, T.; Arai, K.; Sunagozaka, H.; Fushimi, K.; Kobayashi, E.; Kishi, H.; Muraguchi, A.; et al. Immunological features of T cells induced by human telomerase reverse transcriptase-derived peptides in patients with hepatocellular carcinoma. Cancer Lett. 2015, 364, 98–105. [Google Scholar] [CrossRef]
- Nakagawa, H.; Mizukoshi, E.; Kobayashi, E.; Tamai, T.; Hamana, H.; Ozawa, T.; Kishi, H.; Kitahara, M.; Yamashita, T.; Arai, K.; et al. Association Between High-Avidity T-Cell Receptors, Induced by α-Fetoprotein-Derived Peptides, and Anti-Tumor Effects in Patients With Hepatocellular Carcinoma. Gastroenterology 2017, 152, 1395–1406.e10. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M.; Ericsson, A.C. Antimicrobial Peptides: Potential Application in Liver Cancer. Front. Microbiol. 2019, 10, 1257. [Google Scholar] [CrossRef] [PubMed]
- Schauer, P.R.; Bhatt, D.L.; Kirwan, J.P.; Wolski, K.; Brethauer, S.A.; Navaneethan, S.D.; Aminian, A.; Pothier, C.E.; Kim, E.S.; Nissen, S.E.; et al. Bariatric surgery versus intensive medical therapy for diabetes-3-year outcomes. N. Engl. J. Med. 2014, 370, 2002–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blond, E.; Disse, E.; Cuerq, C.; Drai, J.; Valette, P.J.; Laville, M.; Thivolet, C.; Simon, C.; Caussy, C. EASL-EASD-EASO clinical practice guidelines for the management of non-alcoholic fatty liver disease in severely obese people: Do they lead to over-referral? Diabetologia 2017, 60, 1218–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Witte, D.; Wijngaarden, L.H.; van Houten, V.A.A.; van den Dorpel, M.A.; Bruning, T.A.; van der Harst, E.; Klaassen, R.A.; Niezen, R.A. Improvement of Cardiac Function After Roux-en-Y Gastric Bypass in Morbidly Obese Patients Without Cardiac History Measured by Cardiac MRI. Obes. Surg. 2020, 30, 2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauer, D.P.; Feigelson, H.S.; Koebnick, C.; Caan, B.; Weinmann, S.; Leonard, A.C.; Powers, J.D.; Yenumula, P.R.; Arterburn, D.E. Bariatric Surgery and the Risk of Cancer in a Large Multisite Cohort. Ann. Surg. 2019, 269, 95–101. [Google Scholar] [CrossRef]
- Gribble, F.M.; Reimann, F. Function and mechanisms of enteroendocrine cells and gut hormones in metabolism. Nat. Rev. Endocrinol. 2019, 15, 226–237. [Google Scholar] [CrossRef]
- Liu, H.; Hu, C.; Zhang, X.; Jia, W. Role of gut microbiota, bile acids and their cross-talk in the effects of bariatric surgery on obesity and type 2 diabetes. J. Diabetes Investig 2018, 9, 13–20. [Google Scholar] [CrossRef]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Mirhafez, S.R.; Azimi-Nezhad, M.; Dehabeh, M.; Hariri, M.; Naderan, R.D.; Movahedi, A.; Abdalla, M.; Sathyapalan, T.; Sahebkar, A. The Effect of Curcumin Phytosome on the Treatment of Patients with Non-alcoholic Fatty Liver Disease: A Double-Blind, Randomized, Placebo-Controlled Trial. Adv. Exp. Med. Biol. 2021, 1308, 25–35. [Google Scholar] [CrossRef]
- Nadinskaia, M.; Maevskaya, M.; Ivashkin, V.; Kodzoeva, K.; Pirogova, I.; Chesnokov, E.; Nersesov, A.; Kaibullayeva, J.; Konysbekova, A.; Raissova, A.; et al. Ursodeoxycholic acid as a means of preventing atherosclerosis, steatosis and liver fibrosis in patients with nonalcoholic fatty liver disease. World J. Gastroenterol. 2021, 27, 959–975. [Google Scholar] [CrossRef]
- Yoneda, M.; Honda, Y.; Ogawa, Y.; Kessoku, T.; Kobayashi, T.; Imajo, K.; Ozaki, A.; Nogami, A.; Taguri, M.; Yamanaka, T.; et al. Comparing the effects of tofogliflozin and pioglitazone in non-alcoholic fatty liver disease patients with type 2 diabetes mellitus (ToPiND study): A randomized prospective open-label controlled trial. BMJ Open Diabetes Res. Care 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Franco, I.; Bianco, A.; Mirizzi, A.; Campanella, A.; Bonfiglio, C.; Sorino, P.; Notarnicola, M.; Tutino, V.; Cozzolongo, R.; Giannuzzi, V.; et al. Physical Activity and Low Glycemic Index Mediterranean Diet: Main and Modification Effects on NAFLD Score. Results from a Randomized Clinical Trial. Nutrients 2020, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, S.; Shidfar, F.; Ehsani, S.; Adibi, P.; Janani, L.; Eslami, O. The effects of sumac (Rhus coriaria L.) powder supplementation in patients with non-alcoholic fatty liver disease: A randomized controlled trial. Complement. Ther. Clin. Pract. 2020, 41, 101259. [Google Scholar] [CrossRef]
- Soleimani, D.; Rezaie, M.; Rajabzadeh, F.; Gholizadeh Navashenaq, J.; Abbaspour, M.; Miryan, M.; Razmpour, F.; Ranjbar, G.; Rezvani, R.; Jarahi, L.; et al. Protective effects of propolis on hepatic steatosis and fibrosis among patients with nonalcoholic fatty liver disease (NAFLD) evaluated by real-time two-dimensional shear wave elastography: A randomized clinical trial. Phytother. Res. 2021, 35, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, M.; Cheraghpour, M.; Jafarirad, S.; Alavinejad, P.; Asadi, F.; Hekmatdoost, A.; Mohammadi, M.; Yari, Z. The effect of melatonin on treatment of patients with non-alcoholic fatty liver disease: A randomized double blind clinical trial. Complement. Ther. Med. 2020, 52, 102452. [Google Scholar] [CrossRef] [PubMed]
- Podszun, M.C.; Alawad, A.S.; Lingala, S.; Morris, N.; Huang, W.A.; Yang, S.; Schoenfeld, M.; Rolt, A.; Ouwerkerk, R.; Valdez, K.; et al. Vitamin E treatment in NAFLD patients demonstrates that oxidative stress drives steatosis through upregulation of de-novo lipogenesis. Redox Biol. 2020, 37, 101710. [Google Scholar] [CrossRef]
- Ratziu, V.; Sanyal, A.; Harrison, S.A.; Wong, V.W.; Francque, S.; Goodman, Z.; Aithal, G.P.; Kowdley, K.V.; Seyedkazemi, S.; Fischer, L.; et al. Cenicriviroc Treatment for Adults With Nonalcoholic Steatohepatitis and Fibrosis: Final Analysis of the Phase 2b CENTAUR Study. Hepatology 2020, 72, 892–905. [Google Scholar] [CrossRef] [Green Version]
- Scorletti, E.; Afolabi, P.R.; Miles, E.A.; Smith, D.E.; Almehmadi, A.; Alshathry, A.; Childs, C.E.; Del Fabbro, S.; Bilson, J.; Moyses, H.E.; et al. Synbiotics Alter Fecal Microbiomes, But Not Liver Fat or Fibrosis, in a Randomized Trial of Patients With Nonalcoholic Fatty Liver Disease. Gastroenterology 2020, 158, 1597–1610.e7. [Google Scholar] [CrossRef]
- Cheng, Q.; Li, C.; Yang, C.F.; Zhong, Y.J.; Wu, D.; Shi, L.; Chen, L.; Li, Y.W.; Li, L. Methyl ferulic acid attenuates liver fibrosis and hepatic stellate cell activation through the TGF-β1/Smad and NOX4/ROS pathways. Chem. Biol. Interact. 2019, 299, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, C. The role of bone morphogenetic proteins in liver fibrosis. Gastroenterol. Hepatol. Open Access 2021, 12. [Google Scholar] [CrossRef]
- Vacca, M.; Leslie, J.; Virtue, S.; Lam, B.Y.H.; Govaere, O.; Tiniakos, D.; Snow, S.; Davies, S.; Petkevicius, K.; Tong, Z.; et al. Bone morphogenetic protein 8B promotes the progression of non-alcoholic steatohepatitis. Nat. Metab. 2020, 2, 514–531. [Google Scholar] [CrossRef]
- Li, J.; Liu, C.; Zhou, Z.; Dou, B.; Huang, J.; Huang, L.; Zheng, P.; Fan, S.; Huang, C. Isotschimgine alleviates nonalcoholic steatohepatitis and fibrosis via FXR agonism in mice. Phytother. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.A.; Kowdley, K.V. Obeticholic acid for the treatment of nonalcoholic steatohepatitis. Expert Rev. Gastroenterol. Hepatol. 2020, 14, 311–321. [Google Scholar] [CrossRef]
Figure 1.
Factors causing the development and progression of NAFLD-related HCC and other etiologies causing HCC aside from NAFLD. Abbreviations: DEN: diethylnitrosamine; ECM: extracellular matrix proteins; HCC: hepatocellular carcinoma; HSC: hepatic stellate cell; NAFLD: nonalcoholic fatty liver disease; NASH: nonalcoholic steatohepatitis.
Figure 1.
Factors causing the development and progression of NAFLD-related HCC and other etiologies causing HCC aside from NAFLD. Abbreviations: DEN: diethylnitrosamine; ECM: extracellular matrix proteins; HCC: hepatocellular carcinoma; HSC: hepatic stellate cell; NAFLD: nonalcoholic fatty liver disease; NASH: nonalcoholic steatohepatitis.

Figure 2.
Transcriptional and post-transcriptional factors mediated the progression of NAFLD and HCC. Abbreviations: AKT: protein kinase B; ApoB: apolipoprotein B; A1CF: catalytic polypeptide 1 complementation factor; DR5: death receptor 5; FOXO: Forkhead box; HIF-2α: hypoxia-inducible factor-2 alpha; HuR: human antigen R; LPIN: lipin; mTOR: mechanistic target of rapamycin; PI3K: phosphoinositide 3-kinase; PPARs: peroxisome prolifera-tor-activated receptors; SFR10: splicing factor, arginine/serine-rich 10; SREBP-1: sterol regulatory element-binding transcription factor-1; TGF-β: transforming growth factor-beta.
Figure 2.
Transcriptional and post-transcriptional factors mediated the progression of NAFLD and HCC. Abbreviations: AKT: protein kinase B; ApoB: apolipoprotein B; A1CF: catalytic polypeptide 1 complementation factor; DR5: death receptor 5; FOXO: Forkhead box; HIF-2α: hypoxia-inducible factor-2 alpha; HuR: human antigen R; LPIN: lipin; mTOR: mechanistic target of rapamycin; PI3K: phosphoinositide 3-kinase; PPARs: peroxisome prolifera-tor-activated receptors; SFR10: splicing factor, arginine/serine-rich 10; SREBP-1: sterol regulatory element-binding transcription factor-1; TGF-β: transforming growth factor-beta.

Figure 3.
Treatment options for NAFLD-related HCC. Abbreviations: NAFLD: nonalcoholic fatty liver disease; NASH: nonalcoholic steatohepatitis; HCC: hepatocellular carcinoma.
Figure 3.
Treatment options for NAFLD-related HCC. Abbreviations: NAFLD: nonalcoholic fatty liver disease; NASH: nonalcoholic steatohepatitis; HCC: hepatocellular carcinoma.


{kind=link}
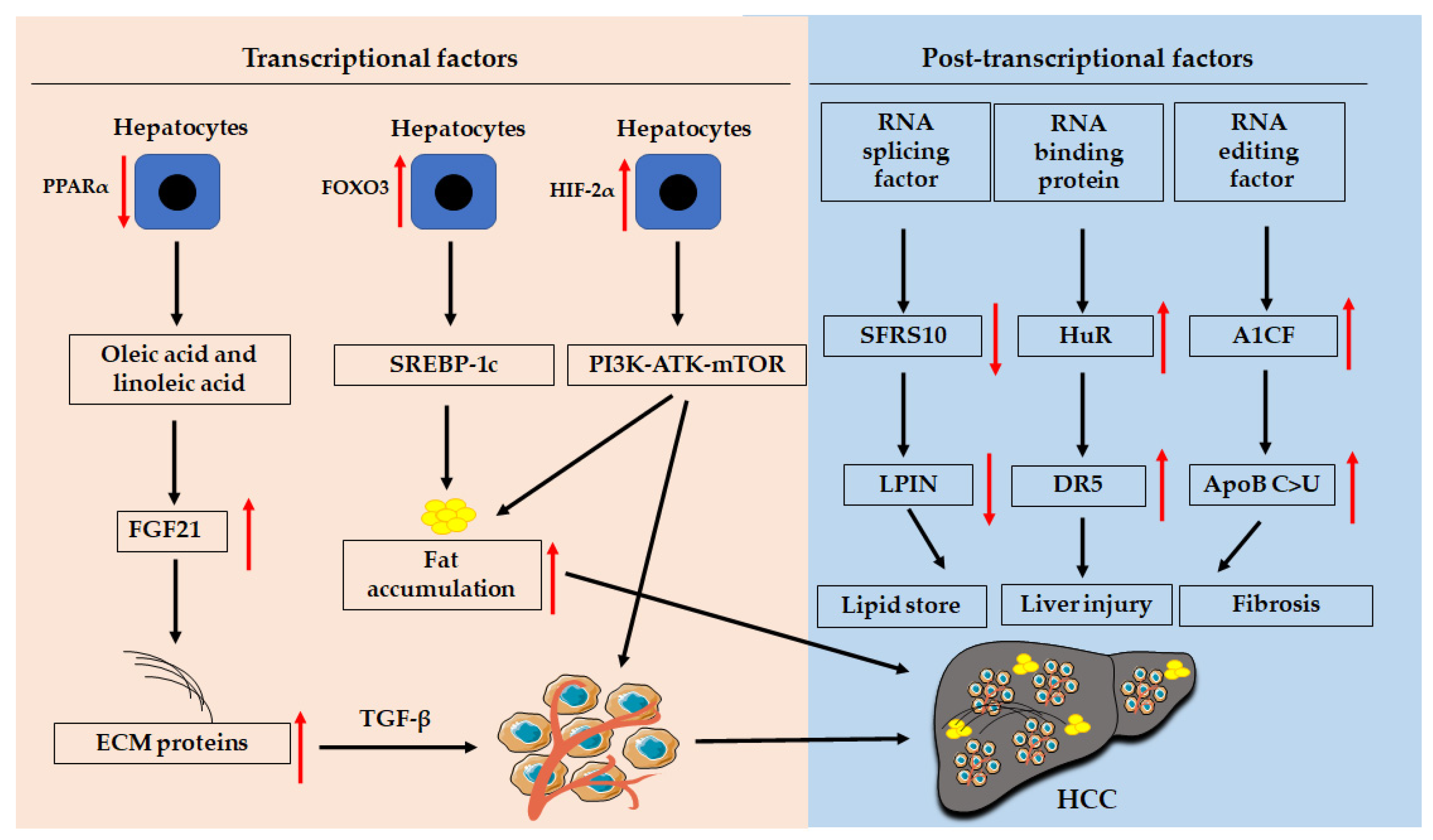{kind=link}
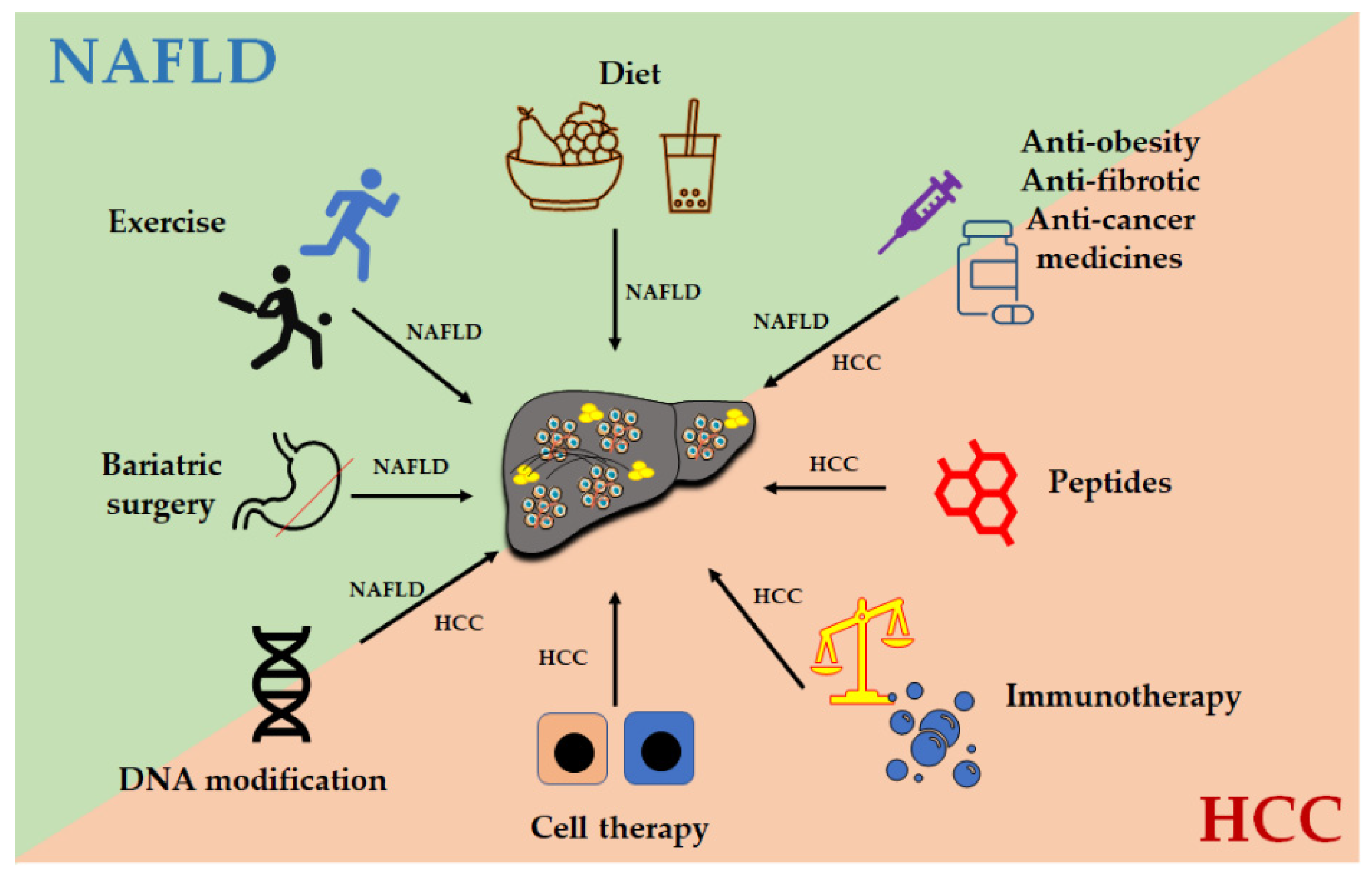{kind=link}
Table 1.
The potential HCC diagnostic methods.
| Methods | Target | Disease | References |
|---|---|---|---|
| Imaging techniques 1 | Various data in clinical records, including images from abdominal ultrasonography, computed tomography, magnetic resonance imaging, electronic health records, liver pathology, data from wearable devices, and multi-omics measurements can be used to predict liver fibrosis, cirrhosis, NAFLD, and for the differentiation of benign tumors from HCC. | NAFLD, liver fibrosis, cirrhosis, and HCC | [115,116,117] |
| Polygenic risk scores (PRS) | Variants in PNPLA3-TM6SF2-GCKR-MBOAT7 are combined in a hepatic fat PRS (PRS-HFC), and then adjusted for HSD17B13 (PRS-5). Similarly, a genetic risk score (GRS) based on three genetic variants of PNPLA3-TM6SF2-HSD17B13 is applied to evaluate how fatty liver disease influences the risk of cirrhosis and HCC. | NAFLD-related HCC | [118,119] |
| Biomarkers | For example, serum levels of inter-alpha-trypsin inhibitor heavy chain 4 are significantly increased in HCC-NAFLD patients compared to those in patients with simple steatosis, NASH, and virus-related HCC. There are some other biomarkers, such as miRNAs, lncRNAs, and circulating tumor DNA (ctDNA). | NAFLD-related HCC | [109,120,121,122,123] |
| Biomarker-based diagnostic algorithm (GALAD) | A GALAD score based on gender, age, and biomarker alpha-fetoprotein (AFP), AFP-L3, and Des-gamma-carboxy prothrombin (DCP). | Early-stage of HCC | [124,125,126] |
| Epigenetic factors with or without combined biomarkers | Epigenetic factors such as circular RNA SMARCA5 can be used as potential new biomarkers for hepatocellular carcinoma. In combination with serum lncRNA linc00152, UCA1 and AFP can show better predictive ability, with areas under the curve (AUC) of 0.912% and 82.9% sensitivity and 88.2% specificity. | Hepatitis, cirrhosis, and HCC | [122,127] |
1 Imaging techniques, especially dynamic multiphase contrast-enhanced computed tomography (CT) scanning and magnetic resonance imaging (MRI), are the most commonly used methods for the clinical diagnosis of HCC.
Table 2.
FDA-approved treatments for HCC.
| Approval Year | Selection | Treatment | Targets | References |
|---|---|---|---|---|
| 2008 | First-line | Sorafenib | Vascular endothelial growth factor receptor (VEGFR)-2 and -3, platelet-derived growth factor receptor (PDGFRβ), receptor tyrosine kinases RET and KIT, and Raf kinase | [150,151] |
| 2017 | Second-line | Regorafenib | VEGFR1-3, TEK receptor tyrosine kinase or angiopoietin-1 receptor (TIE2), fibroblast growth factor receptor 1 (FGFR1) and PDGFRβ, oncogenic kinases c-KIT, RET, and c-RAF/RAF-1 | [138,152] |
| 2017 | Second-line | Nivolumab | PD-1 | [153] |
| 2018 | First-line | Lenvatinib | An oral inhibitor of VEGFR 1-3, FGFR 1-4, PDGFRα, receptor tyrosine kinases RET and KIT | [135,154] |
| 2018 | Second-line | Pembrolizumab | PD-1 | [155] |
| 2019 | Second-line | Cabozantinib | Tyrosine kinases, including MET, AXL, and VEGFR 1-3 | [139,156] |
| 2019 | Second-line | Ramucirumab | VEGFR2 | [157] |
| 2020 | Second-line | Nivolumab and Ipilimumab | PD-1 and CTLA-4 | [158,159] |
| 2020 | First-line | Atezolizumab plus Bevacizumab | PD-L1 and VEGF | [136,160] |
Table 3.
The completed clinical trials for NAFLD/NASH treatment.
| Liver Disease | Treatment | Class | Effect | Trial | References |
|---|---|---|---|---|---|
| NAFLD | Curcumin | Polyphenol | Daily supplementation of curcumin (250 mg) for 2 months caused a significant reduction in hepatic steatosis and enzymes in patients with NAFLD compared to placebo. | A double-blind, randomized, placebo-controlled trial. | [171] |
| NAFLD | Ursodeoxycholic acid (UDCA) | Bile acid | Treatment with UDCA (15 mg/kg/d) for 3 months, the level of enzymes alanine aminotransferase (ALT), aspartate transaminase (AST), and glutamyltransferase decreased. After 6-month treatment, body weight, fatty liver index, total cholesterol, low-density lipoprotein, and triglyceride were significantly reduced. | An open-label, multicenter, international noncomparative trial. | [172] |
| NAFLD with T2DM | Tofogliflozin | A selective sodium-glucose cotransporter 2 inhibitor | Oral treatment with 20 mg tofogliflozin or 15–30 mg daily for 24 weeks significantly decreased body weight and hepatic steatosis. | A randomized prospective open-label controlled trial. | [173] |
| NAFLD | Diet and activity | Physical activity (PA), a low glycemic index Mediterranean diet (LGIMD), or their combined effect. | After 45 days, there was a statistically significant reduction in the NAFLD score in each group except for the control diet-treated group. After 90 days, the combined treatment showed the best effect on the reduction of NAFLD score. | A randomized clinical trial. | [174] |
| NAFLD | Sumac | Herbal medicine | After a 12-week intervention, the sumac-treated group displayed a greater reduction in hepatic fibrosis and enzymes ALT and AST, fasting blood sugar, serum insulin, and HOMA-IR (insulin resistance index), as well as higher QUICKI (insulin sensitivity index) compared to the placebo. | A randomized controlled trial. | [175] |
| NAFLD | Propolis | An anti-inflammatory agent | Patients with propolis at a dose of 250 mg twice daily for 4 months compared to placebo showed a significant improvement in hepatic steatosis and a significant reduction of liver stiffness, as well as serum high-sensitivity C-reactive protein (hs-CRP). | A randomized clinical trial. | [176] |
| NAFLD | Melatonin | A hormone | Administration of 6 mg/day melatonin significantly reduced body weight, waist and abdominal circumference systolic and diastolic blood pressure, serum levels of leptin, ALT, AST, and hs-CRP, and the grade of fatty liver compared with the placebo. | A randomized double-blind clinical trial. | [177] |
| NAFLD | Vitamin E | An antioxidant vitamin | Treatment with vitamin E improved liver injury and steatosis. | A single-center prospective trial. | [178] |
| NASH | Cenicriviroc | A dual antagonist C-C chemokine receptor type 2 (CCR2) and CCR5 | Therapy with cenicriviroc showed an antifibrotic effect without impacting steatohepatitis at year 1 in responders, which was maintained in year 2 with a greater effect in advanced fibrosis. | A randomized, controlled study. | [179] |
| NAFLD | Synbiotic | A mixture of probiotics and prebiotics | Administration of a synbiotic combination of probiotic and prebiotic for one year changed gut microbiota but did not reduce liver fat content or markers of liver fibrosis. | A double-blind phase 2 trial. | [180] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, C.; Yang, M. The Emerging Factors and Treatment Options for NAFLD-Related Hepatocellular Carcinoma. Cancers 2021, 13, 3740. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153740
AMA Style
Zhang C, Yang M. The Emerging Factors and Treatment Options for NAFLD-Related Hepatocellular Carcinoma. Cancers. 2021; 13(15):3740. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153740
Chicago/Turabian StyleZhang, Chunye, and Ming Yang. 2021. "The Emerging Factors and Treatment Options for NAFLD-Related Hepatocellular Carcinoma" Cancers 13, no. 15: 3740. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153740
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.