
A Universal Approach to Analyzing Transmission Electron Microscopy with ImageJ

 , , , , , and
, , , , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
2.1. Animal Care & Maintenance
2.2. Isolation of Satellite Cells
2.3. Cell Culture & Differentiation
2.4. Thapsigargin Treatment
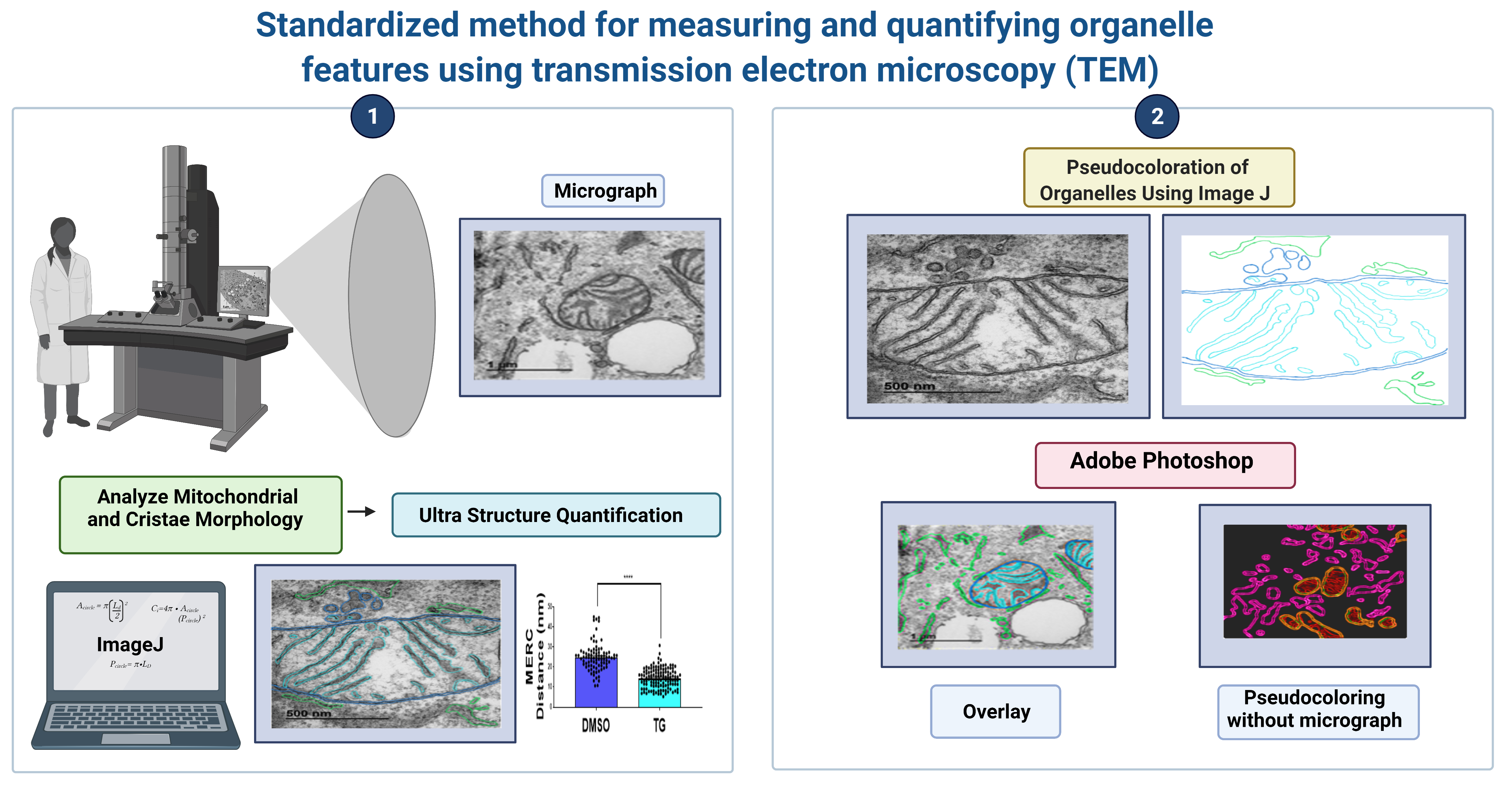
2.5. Systematic ImageJ Analysis for TEM Measurements of Organelle Morphology
2.5.1. ImageJ Quantification
- (1)
- Length (L): The length is measured by applying the Pythagorean Theorem to the coordinate pixels. Thus, where Lp1,p2 denotes the Euclidean distance between pixel 1 and pixel 2. Length is given by (Equation: L(p1 + p2) = √(x1 − x2)2 + (y1 − y2)2)
- (2)
- Area: This is calculated similarly. The area calculated by ImageJ is the area of pixels.
- i.
- Rectangles (Arectangle): Let L1 be the length (as defined previously) of one side of the rectangle and let L2 be the length of the other side; the area is then given by (Equation: Arectangle = L1 × L2)
- ii.
- Circles (A circle): Once the circle is traced, the software chooses two opposite pixels on the circle, pixel 1 and pixel 2. Equation then gives the diameter LD. Circle area is given by (Equation: Acircle = π(LD/2)2)
- (3)
- Perimeter:
- i.
- Rectangles (P rectangle): Consider L1 and L2 as defined in Section 2, rectangle perimeter is given by (Equation Prectangle = 4L1 × L2)
- ii.
- Circles (P circle): Consider LD as defined in 2.ii., circle perimeter is given by (Equation: Pcircle = π × LD)
- (4)
- Circularity Index (Ci): Consider P circle and A circle as in 3.ii and 2.ii, respectively. The circularity index is given by (Equation: Ci = 4π × Acircle/(Pcircle)2)
- (5)
- Volume: Surface area of substructure features are divided by the area of the structure (SA:A) to provide an estimate volume.
2.5.2. ImageJ Parameters
3. Protocol for ImageJ Installation
3.1. Installing and Preparing ImageJ Software for Analysis
- i.
- Download ImageJ software from the NIH website (Download (nih.gov, accessed on 17 June 2021).
- ii.
- Install and open the ImageJ software
- iii.
- Open the Region of Interest (ROI) Manager, which is used to record and keep track of measurements, by selecting Analyze ➧ Tools ➧ ROI Manager (Figure 1A).
- iv.
- Click on Analyze ➧ Set Measurements to input the measurements that ImageJ will make, such as area, circularity, and perimeter (Figure 1A).
- v.
- Drag the image to be analyzed (TIFF or DM3 file) directly into ImageJ (Figure 1B).
3.2. Analyzing Mitochondrial Morphology
- i.
- Measuring mitochondrial area, circularity, and perimeter.
- ii.
- Click on Freehand Selections on the toolbar to access the Freehand tool.
- iii.
- Trace the outline of the entire cell. To store the measurement, click Add on the ROI Manager. This will be used to normalize later measurements.
- iv.
- Trace the outer mitochondrial membrane of each mitochondrion. Add the shape to the ROI Manager (Figure 2A).
- v.
- Click on Measure in the ROI Manager to obtain the measurements for each shape.
3.3. Measuring Mitochondrial Length and Width
3.4. Analyzing Cristae Morphology
- i.
- Split each image into four quadrants and randomly pick two quadrants to analyze; use the same quadrants for all images.
- ii.
- Using the Freehand tool in ImageJ, outline the outer membrane of a mitochondrion.
- iii.
- Add the measurement to the ROI Manager.
- iv.
- Trace the outline of each crista within the mitochondrion. Add each measurement to the ROI Manager (Figure 2B).
- v.
- Click on Measure in the ROI Manager to obtain the area of each crista, and cristae surface area is the sum of the areas of all the cristae in a single mitochondrion.
- vi.
- To calculate cristae estimated volume density, cristae surface area is divided by the area of the mitochondrion.
- vii.
- To determine the cristae number, count the number of cristae in each mitochondrion.Alternatively, you can use the number of cristae measurements obtained earlier.
3.5. Analyzing Mitochondria-ER Contacts (MERCs)
- i.
- Split each image into four quadrants and randomly pick two quadrants to analyze; use the same quadrants for all images.
- ii.
- Identify a Mitochondrion-ER contact (MERC), a mitochondrion in close contact with the ER membrane, in your TEM image.
- iii.
- Using the Freehand Selections tool, trace both the outer membrane of the mitochondrion and the ER membrane at a contact point (Figure 2C).
- iv.
- To obtain the MERC contact length, click on the Straight, Segmented, or Freehand Lines tool on the toolbar, select Freehand Line, and draw a line spanning the length of the contact site (Figure 2C).
- v.
- Add the measurement to the ROI Manager and use the Measure function to determine the length of the contact.
- vi.
- To measure the MERC distance, use the Freehand Line tool to draw a line between the two organelles (Figure 2D).
- vii.
- Add the measurement to the ROI Manager and use the Measure function to determine the distance.
- viii.
- To calculate percent coverage, divide the contact length by either the mitochondrial surface area (mitochondrial percent coverage) or ER surface (ER percent coverage), and multiply the value by 100 to obtain a percentage.Similar measurements can be made for other organelle interactions using the same steps detailed here.
3.6. (Optional) Pseudo-Coloration of Organelles Using ImageJ
- i.
- Open ImageJ
- ii.
- Select File ➧ Open and select a desired TIF (or alternative) image.
- iii.
- Select Image ➧ Type ➧ RGB Color to allow for visible colors (Supplementary Figure S1A).
- iv.
- Select the Color Picker Tool from the toolbar and select the desired color for the organelle (Supplementary Figure S1B)
- v.
- Outline the organelle
- vi.
- On the right-hand side of the tool bar, select the Paintbrush Overlay Tool.
- vii.
- Double click the Paintbrush Overlay Tool and set the width to 1, or a small enough width to outline, and the transparency to 0 (Supplementary Figure S1C)
- viii.
- Trace the outlines of the organelles with the brush tool and mouse (Supplementary Figure S1D).
- ix.
- Use the +/− buttons to better zoom in to the picture as needed.
- x.
- Color the organelle
- xi.
- Double click the Paintbrush Overlay Tool and set the width to 3–15, depending on the size of the organelle to color in.
- xii.
- Use the mouse to fill in the previously made outline (Supplementary Figure S1E).
- xiii.
- Repeat Steps 5.1.5 and 5.1.6 with new colors as many times as necessary for each organelle or structure that needs to be colored (Supplementary Figure S1F).
3.7. (Optional) Pseudo-Coloration of Organelles Using Adobe Photoshop
- i.
- Install and Open Adobe Photoshop
- ii.
- Click File ➧ Open and select the saved ImageJ export file.
- iii.
- Under the Layers menu click New ➧ Layer and ensure the new layer is selected and above the layer with the image.
- iv.
- Outline the organelle (Supplementary Figure S2).
- v.
- On the left tool sidebar, select the Freeform Pen Tool. Use tiny increments to slowly outline. If mistakes are made, use the eraser tool to fix them.Ensure one continuous line is used for the outline of organelles.
- vi.
- Right-click the line drawn and click Stroke Path, set to pencil or paintbrush to create a continuous and clean line (Supplementary Figure S2A).
- vii.
- Right-click the original line drawn and click Delete Path to remove the original line and only be left with the clean version (Supplementary Figure S2B).
- viii.
- Clean up the outline
- ix.
- On the left tool sidebar, select the Eraser.
- x.
- Use the eraser to manually erase parts where the lines are uneven or unrepresentative of the organelle.
- xi.
- Under the Layers menu, click New ➧ Layer and ensure the new layer is selected and below the layer with the outline.
- xii.
- Color the organelle
- xiii.
- On the left-hand side, select the Paint Brush tool, and at the bottom select a color Supplementary Figure S2C).
- xiv.
- Use the mouse to color in the area outlined. This does not need to be precise as it will later be adjusted (Supplementary Figure S2D).
- xv.
- To fix uneven coloring, once fully colored, for each organelle, select the Magic Wand. Go to the outline layer and click onto each organelle to cause the coloration to fit evenly inside the outline (Supplementary Figure S2E).
- xvi.
- Delete the Color layer, so the only layer left is the Outline layer with the new colors and the background image (Supplementary Figure S2F).
- xvii.
- During the coloration process, so long as the outline is continuous, one can use the Magic Wand from the tool menu to instantly fill in the area with one color.
- xviii.
- Click File ➧ Save As… and save as a desired export file for further analysis, if necessary.
4. Representative Results
4.1. OPA1 Knockdown Decreases Mitochondrial Coverage
4.2. OPA1 Effects on Cristae Structure
4.3. Thapsigargin Treatment Increases MERCs
5. Discussion
6. Perspectives on TEM Measurements
7. Perspectives on TEM Standardization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brand, M.; Orr, A.; Perevoshchikova, I.; Quinlan, C. The role of mitochondrial function and cellular bioenergetics in ageing and disease. Br. J. Dermatol. 2013, 169, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, D.S.; Blower, M.D. The endoplasmic reticulum: Structure, function and response to cellular signaling. Cell. Mol. Life Sci. 2016, 73, 79–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bone, R.; Oyebamiji, O.; Talware, S.; Selvaraj, S.; Krishnan, P.; Syed, F.; Wu, H.; Evans-Molina, C. A Computational Approach for Defining a Signature of β-Cell Golgi Stress in Diabetes. Diabetes 2020, 69, 2364–2376. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: A dawn for evolutionary medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, L.; Tabas, I. Role of endoplasmic reticulum stress in metabolic disease and other disorders. Annu. Rev. Med. 2012, 63, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhao, F.; Ma, X.; Perry, G.; Zhu, X. Mitochondria dysfunction in the pathogenesis of Alzheimer’s disease: Recent advances. Mol. Neurodegener. 2020, 15, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Facility, The University of Iowa’s Central Microscopy Research Facility Methodology. Available online: https://cmrf.research.uiowa.edu/ (accessed on 17 June 2021).
- Franken, L.E.; Grünewald, K.; Boekema, E.J.; Stuart, M.C.A. A Technical Introduction to Transmission Electron Microscopy for Soft-Matter: Imaging, Possibilities, Choices, and Technical Developments. Small 2020, 16, e1906198. [Google Scholar] [CrossRef]
- Pereira, R.O.; Tadinada, S.M.; Zasadny, F.M.; Oliveira, K.J.; Pires, K.M.P.; Olvera, A.; Jeffers, J.; Souvenir, R.; McGlauflin, R.; Seei, A.; et al. OPA1 deficiency promotes secretion of FGF21 from muscle that prevents obesity and insulin resistance. EMBO J. 2017, 36, 2126–2145. [Google Scholar] [CrossRef]
- Kumar, A.; Xie, L.; Ta, C.M.; Hinton, O.A.; Gunasekar, S.K.; Minerath, A.R.; Shen, K.; Maurer, J.M.; Grueter, E.C.; Abel, E.D.; et al. SWELL1 regulates skeletal muscle cell size, intracellular signaling, adiposity and glucose metabolism. eLife 2020, 9, e58941. [Google Scholar] [CrossRef]
- Han, J.; Back, S.H.; Hur, J.; Lin, Y.-H.; Gildersleeve, R.; Shan, J.; Yuan, C.L.; Krokowski, D.; Wang, S.; Hatzoglou, M.; et al. ER-stress-induced transcriptional regulation increases protein synthesis leading to cell death. Nat. Cell Biol. 2013, 15, 481–490. [Google Scholar] [CrossRef]
- Parra, V.; Verdejo, H.E.; Iglewski, M.; Del Campo, A.; Troncoso, R.; Jones, D.; Zhu, Y.; Kuzmicic, J.; Pennanen, C.; Lopez-Crisosto, C.; et al. Insulin stimulates mitochondrial fusion and function in cardiomyocytes via the AktmTOR-NFkB-Opa-1 signaling pathway. Diabetes 2014, 63, 75–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponce, J.M.; Coen, G.; Spitler, K.M.; Dragisic, N.; Martins, I.; Hinton, A., Jr.; Mungai, M.; Tadinada, S.M.; Zhang, H.; Oudit, G.Y.; et al. Stress-Induced Cyclin C Translocation Regulates Cardiac Mitochondrial Dynamics. J. Am. Heart Assoc. 2020, 9, e014366. [Google Scholar] [CrossRef]
- Carter, C.S.; Huang, S.C.; Searby, C.C.; Cassaidy, B.; Miller, M.J.; Grzesik, W.J.; Piorczynski, T.B.; Pak, T.K.; Walsh, S.A.; Acevedo, M.; et al. Exposure to static magnetic and electric fields treats type 2 diabetes. Cell Metab. 2020, 32, 561–574.e7. [Google Scholar] [CrossRef]
- Tsushima, K.; Bugger, H.; Wende, A.; Soto, J.; Jenson, G.A.; Tor, A.R.; McGlauflin, R.; Kenny, H.C.; Zhang, Y.; Souvenir, R.; et al. Mitochondrial reactive oxygen species in lipotoxic hearts induce post-translational modifications of AKAP121, DRP1, and OPA1 that promote mitochondrial fission. Circ. Res. 2018, 122, 58–73. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Amini, P.; Stojkov, D.; Felser, A.; Jackson, C.B.; Courage, C.; Schaller, A.; Gelman, L.; Soriano, M.E.; Nuoffer, J.-M.; Scorrano, L.; et al. Neutrophil Extracellular Trap Formation Requires OPA1-Dependent Glycolytic ATP Production. Nat. Commun. 2018, 9, 2958. [Google Scholar] [CrossRef] [Green Version]
- Pamenter, M.E.; Perkins, G.A.; Gu, X.Q.; Ellisman, M.H.; Haddad, G.G. DIDS (4,4-Diisothiocyanatostilbenedisulphonic Acid) Induces Apoptotic Cell Death in a Hippocampal Neuronal Cell Line and Is Not Neuroprotective against Ischemic Stress. PLoS ONE 2013, 8, e60804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, M.; Mahata, S.K.; Padhan, D.K.; Sen, M. CCN6 (Wnt Induced Signaling Protein–3) Regulates Mitochondrial Function. J. Cell Sci. 2016, 129, 2841–2851. [Google Scholar] [CrossRef] [Green Version]
- Shults, N.V.; Kanovka, S.S.; Ten Eyck, J.E.; Rybka, V.; Suzuki, Y.J. Ultrastructural Changes of the Right Ventricular Myocytes in Pulmonary Arterial Hypertension. JAHA 2019, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Broeke, J.; Mateos Pérez, J.M.; Pascau, J. Image Processing with ImageJ: Extract and Analyze Data from Complex Images with ImageJ, the World’s Leading Image Processing Tool, 2nd ed.; Community Experience Distilled; Packt Publishing Open Source: Birmingham Mumbai, UK, 2015; ISBN 978-1-78588-983-7. [Google Scholar]
- Eisner, V.; Cupo, R.R.; Gao, E.; Csordás, G.; Slovinsky, W.S.; Paillard, M.; Cheng, L.; Ibetti, J.; Chen, S.R.W.; Chuprun, J.K.; et al. Mitochondrial fusion dynamics is robust in the heart and depends on calcium oscillations and contractile activity. Proc. Natl. Acad. Sci. USA 2017, 114, E859–E868. [Google Scholar] [CrossRef] [Green Version]
- Zorzano, A.; Liesa, M.; Palacín, M. Role of mitochondrial dynamics proteins in the pathophysiology of obesity and type 2 diabetes. Int. J. Biochem. Cell Biol. 2009, 41, 1846–1854. [Google Scholar] [CrossRef] [PubMed]
- Frezza, C.; Cipolat, S.; De Brito, O.M.; Micaroni, M.; Beznoussenko, G.V.; Rudka, T.; Bartoli, D.; Polishuck, R.S.; Danial, N.N.; De Strooper, B.; et al. OPA1 Controls Apoptotic Cristae Remodeling Independently from Mitochondrial Fusion. Cell 2006, 126, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moltedo, O.; Remondelli, P.; Amodio, G. The mitochondria–endoplasmic reticulum contacts and their critical role in aging and age-associated diseases. Front. Cell Dev. Biol. 2019, 7, 172. [Google Scholar] [CrossRef] [PubMed]
- Lindner, P.; Christensen, S.B.; Nissen, P.; Møller, J.V.; Engedal, N. Cell death induced by the ER stressor thapsigargin involves death receptor 5, a non-autophagic function of MAP1LC3B, and distinct contributions from unfolded protein response components. Cell Commun. Signal. 2020, 18, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, R.; Vicencio, J.M.; Parra, V.; Troncoso, R.; Munoz, J.P.; Bui, M.; Quiroga, C.; Rodriguez, A.E.; Verdejo, H.E.; Ferreira, J.; et al. Increased ER-mitochondrial coupling promotes mitochondrial respiration and bioenergetics during early phases of ER stress. J. Cell Sci. 2011, 124, 2511. [Google Scholar] [CrossRef] [Green Version]
- Howard, V.; Reed, M. Unbiased Stereology: Three-Dimensional Measurement in Microscopy; Garland Science: New York, NY, USA, 2004. [Google Scholar]
- Ware, C.I. A simple method to ‘point count’ silt using scanning electron microscopy aided by image analysis. J. Microsc. 2003, 212, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Bravo-Sagua, R.; López-Crisosto, C.; Parra, V.; Rodriguez-Peña, M.; Rothermel, B.A.; Quest, A.F.; Lavandero, S. mTORC1 inhibitor rapamycin and ER stressor tunicamycin induce differential patterns of ER-mitochondria coupling. Sci. Rep. 2016, 6, 36394. [Google Scholar] [CrossRef]
- Bleck, C.; Kim, Y.; Willingham, T.B.; Glancy, B. Subcellular connectomic analyses of energy networks in striated muscle. Nat. Commun. 2018, 9, 5111. [Google Scholar] [CrossRef]
- Mouton, P.R. Applications of unbiased stereology to neurodevelopmental toxicology. In Developmental Neurotoxicology Research; Wiley: Hoboken, NJ, USA, 2011; pp. 53–75. [Google Scholar]
- Mouton, P.R. Principles and Practices of Unbiased Stereology: An Introduction for Bioscientists; Johns Hopkins University Press: Baltimore, MD, USA, 2002; ISBN 978-0-8018-6797-2. [Google Scholar]
- Wernitznig, S.; Reichmann, F.; Sele, M.; Birkl, C.; Haybäck, J.; Kleinegger, F.; Birkl-Töglhofer, A.; Krassnig, S.; Wodlej, C.; Holzer, P.; et al. An Unbiased Approach of Sampling TEM Sections in Neuroscience. JoVE J. Vis. Exp. 2019, 146, e58745. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, J.; Katti, P.; Biete, M.; Mungai, M.; AshShareef, S.; Neikirk, K.; Garza Lopez, E.; Vue, Z.; Christensen, T.A.; Beasley, H.K.; et al. A Universal Approach to Analyzing Transmission Electron Microscopy with ImageJ. Cells 2021, 10, 2177. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092177
Lam J, Katti P, Biete M, Mungai M, AshShareef S, Neikirk K, Garza Lopez E, Vue Z, Christensen TA, Beasley HK, et al. A Universal Approach to Analyzing Transmission Electron Microscopy with ImageJ. Cells. 2021; 10(9):2177. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092177
Chicago/Turabian StyleLam, Jacob, Prasanna Katti, Michelle Biete, Margaret Mungai, Salma AshShareef, Kit Neikirk, Edgar Garza Lopez, Zer Vue, Trace A. Christensen, Heather K. Beasley, and et al. 2021. "A Universal Approach to Analyzing Transmission Electron Microscopy with ImageJ" Cells 10, no. 9: 2177. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092177