
Gut-Kidney Impairment Process of Adenine Combined with Folium sennae-Induced Diarrhea: Association with Interactions between Lactobacillus intestinalis, Bacteroides acidifaciens and Acetic Acid, Inflammation, and Kidney Function

Abstract
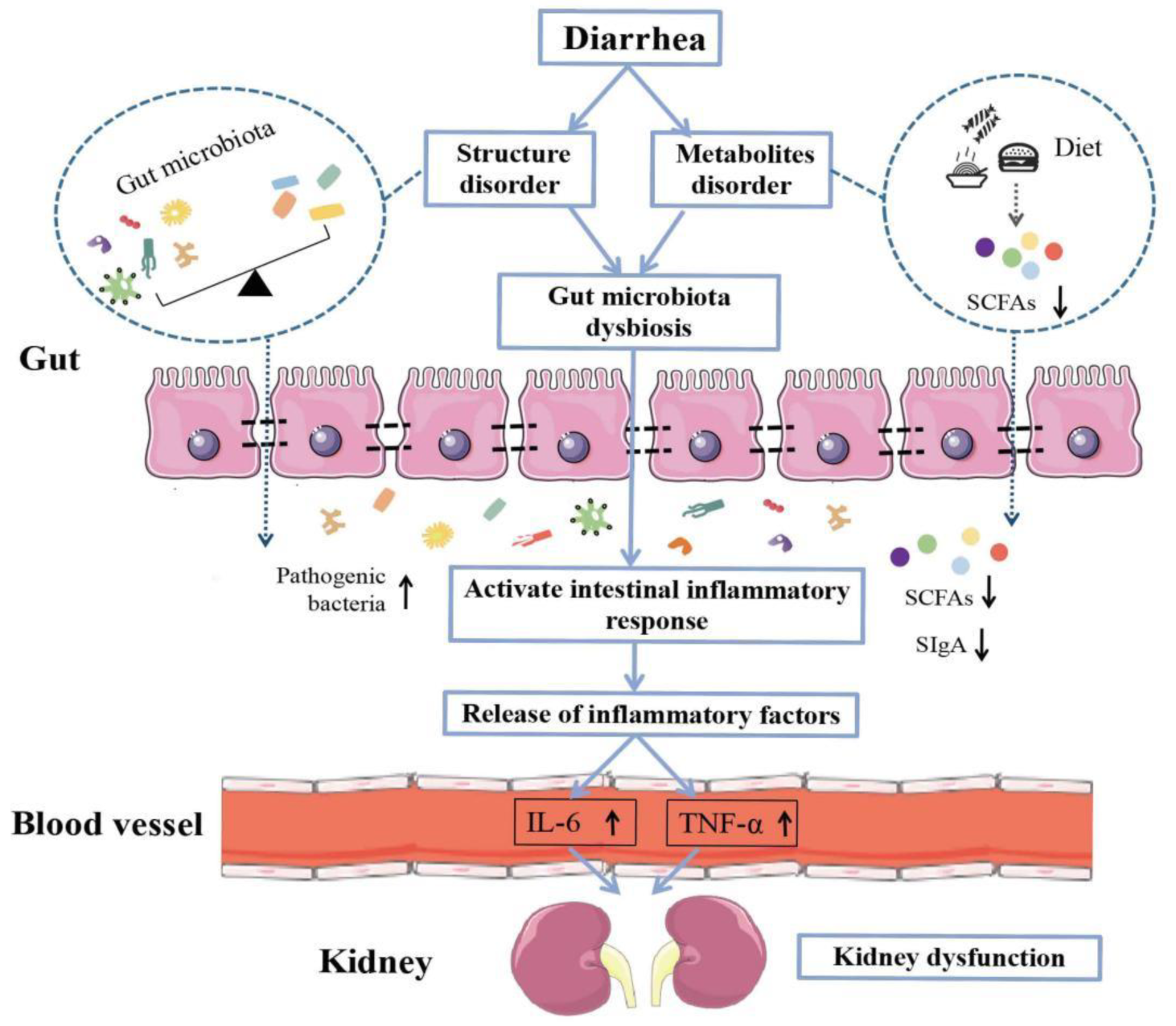
:1. Introduction
2. Materials and Methods
2.1. Medicine
2.2. Reagents
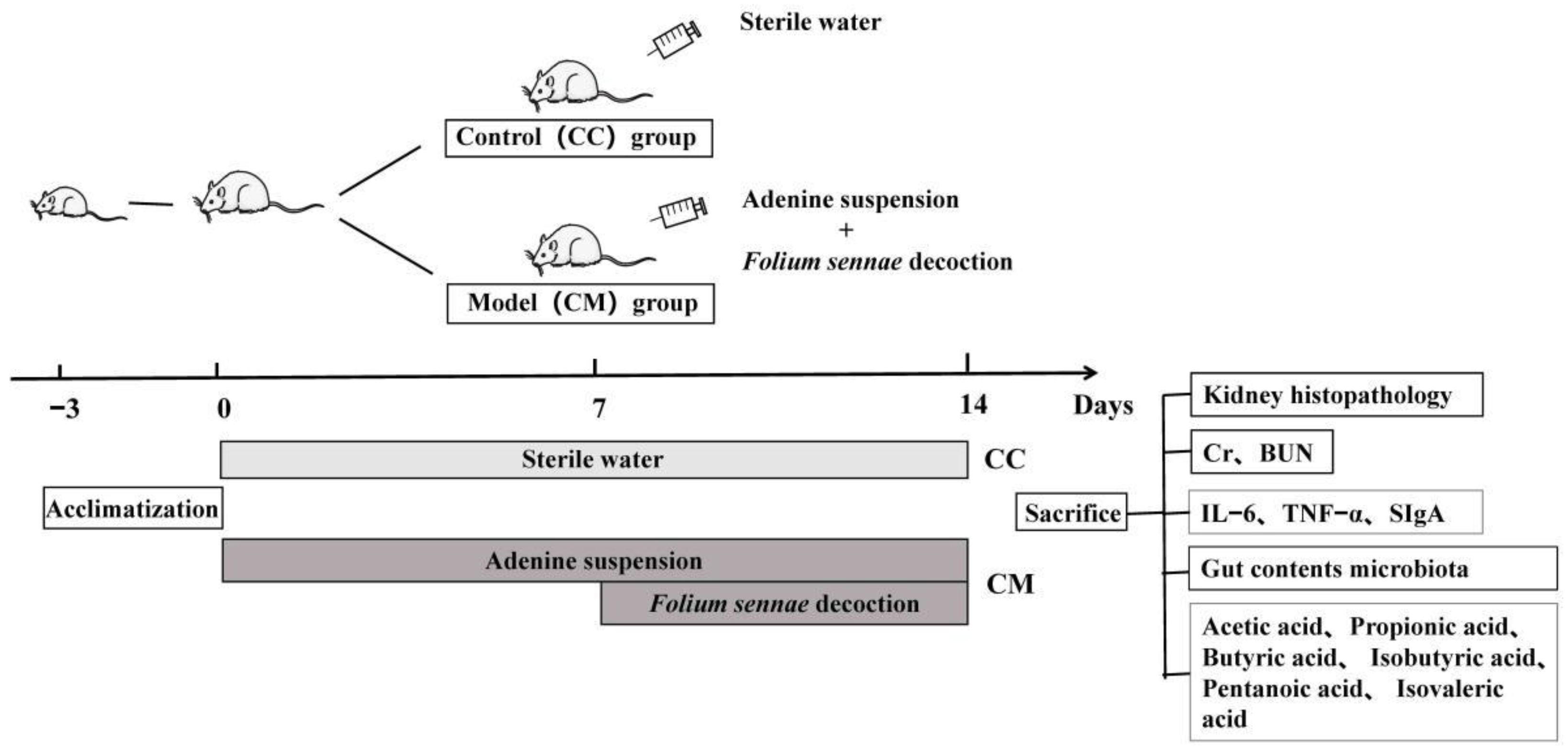
2.3. Animals
2.4. Animal Treatment
2.5. Sample Collection
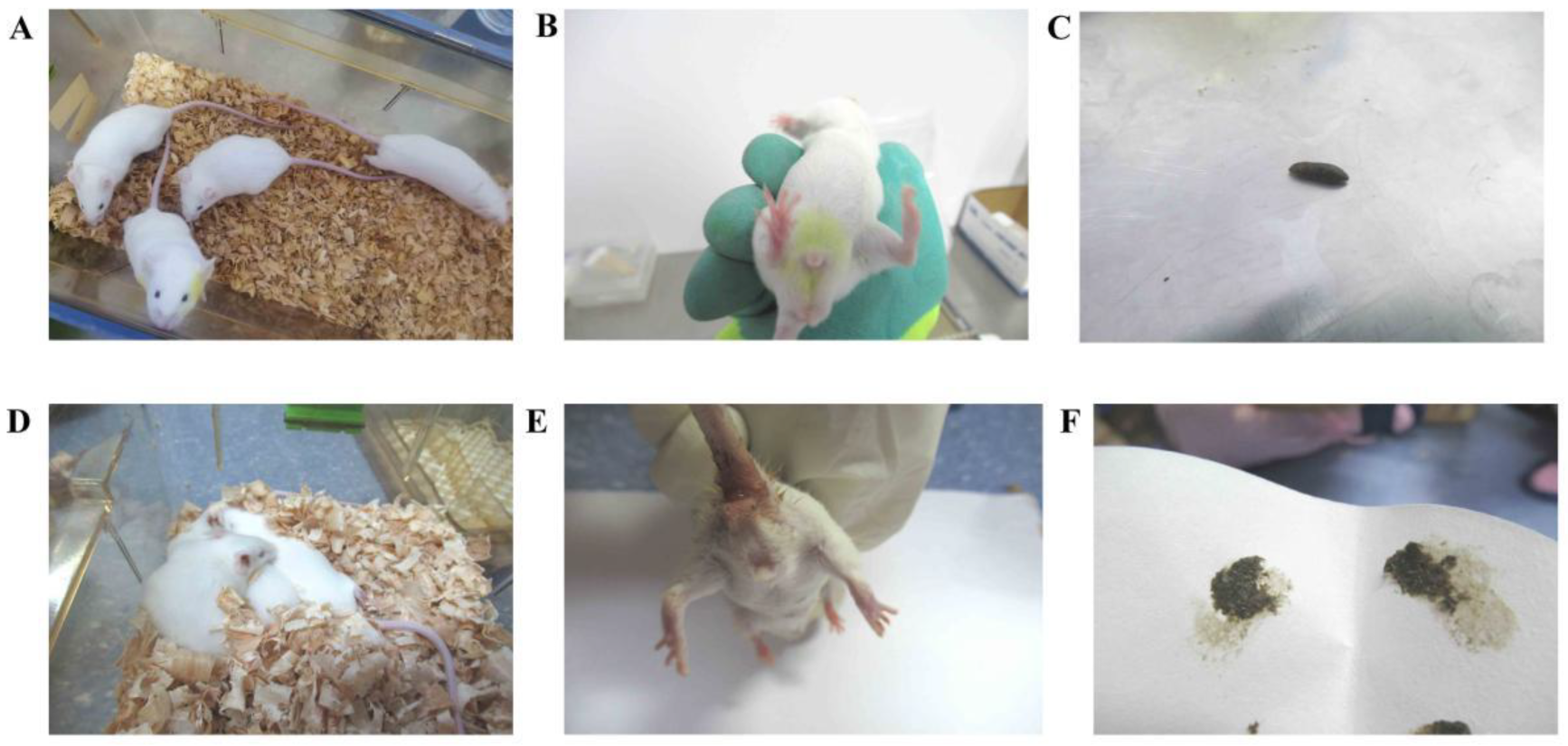
2.6. General Behavioral Observations
2.7. Histological Observation of the Kidneys
2.8. Blood Biochemical Measurement
2.9. ELISA Analysis
2.10. Preparation of the Total DNA and the 16S rRNA High-Throughput Sequencing
2.11. Gas Chromatography-Mass Spectrometry (GC-MS)
2.12. Bioinformatics Analysis
2.13. Statistical Analysis
3. Results
3.1. Behavioral Changes in the Mice
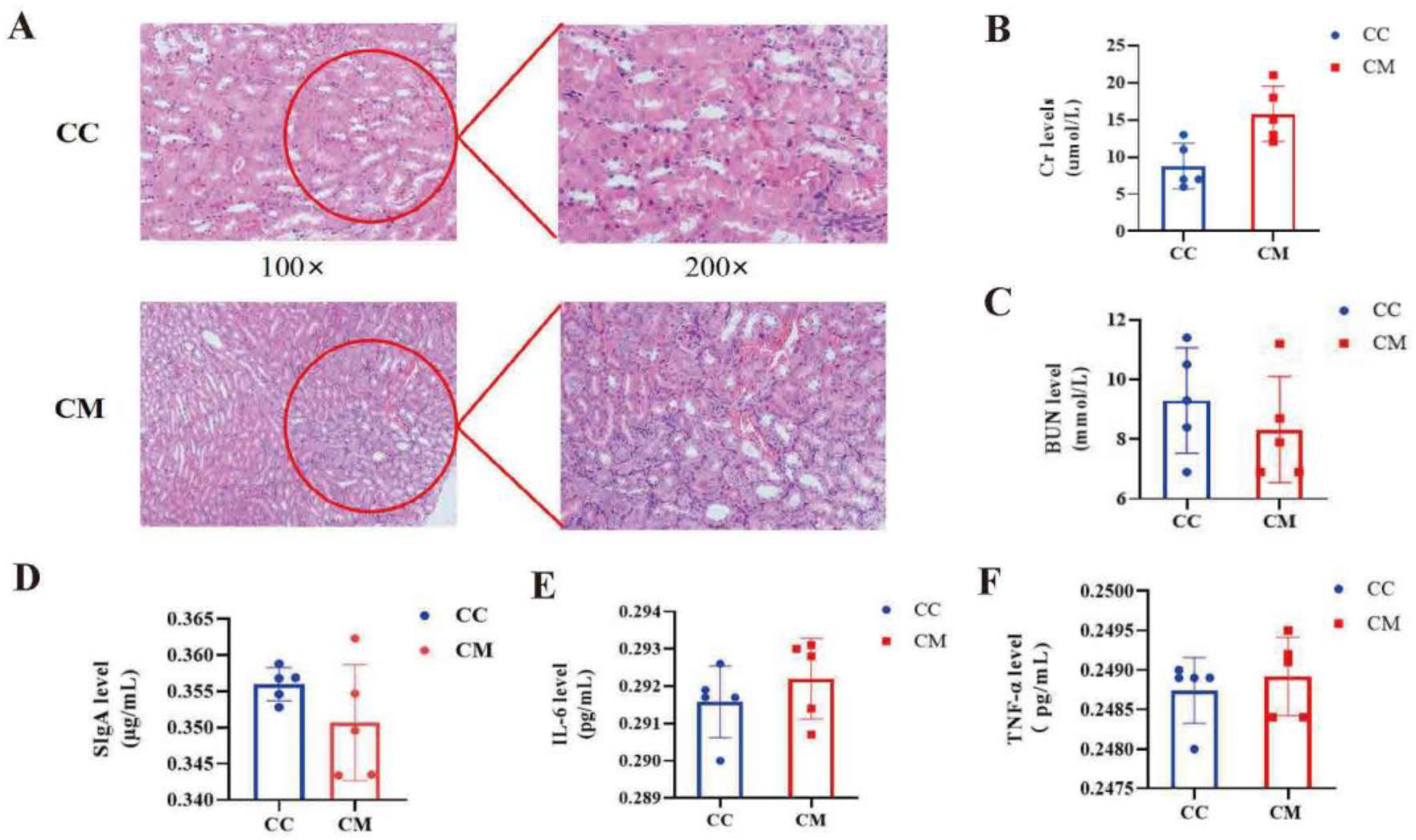
3.2. Changes in the Kidney Structure and Function in the Mice
3.3. Analysis of the Intestinal Inflammatory Reaction in the Mice
3.4. Analysis of the Structure and Function of the Gut Content Microbiota in the Mice
3.4.1. Sequencing Data Quality Assessment
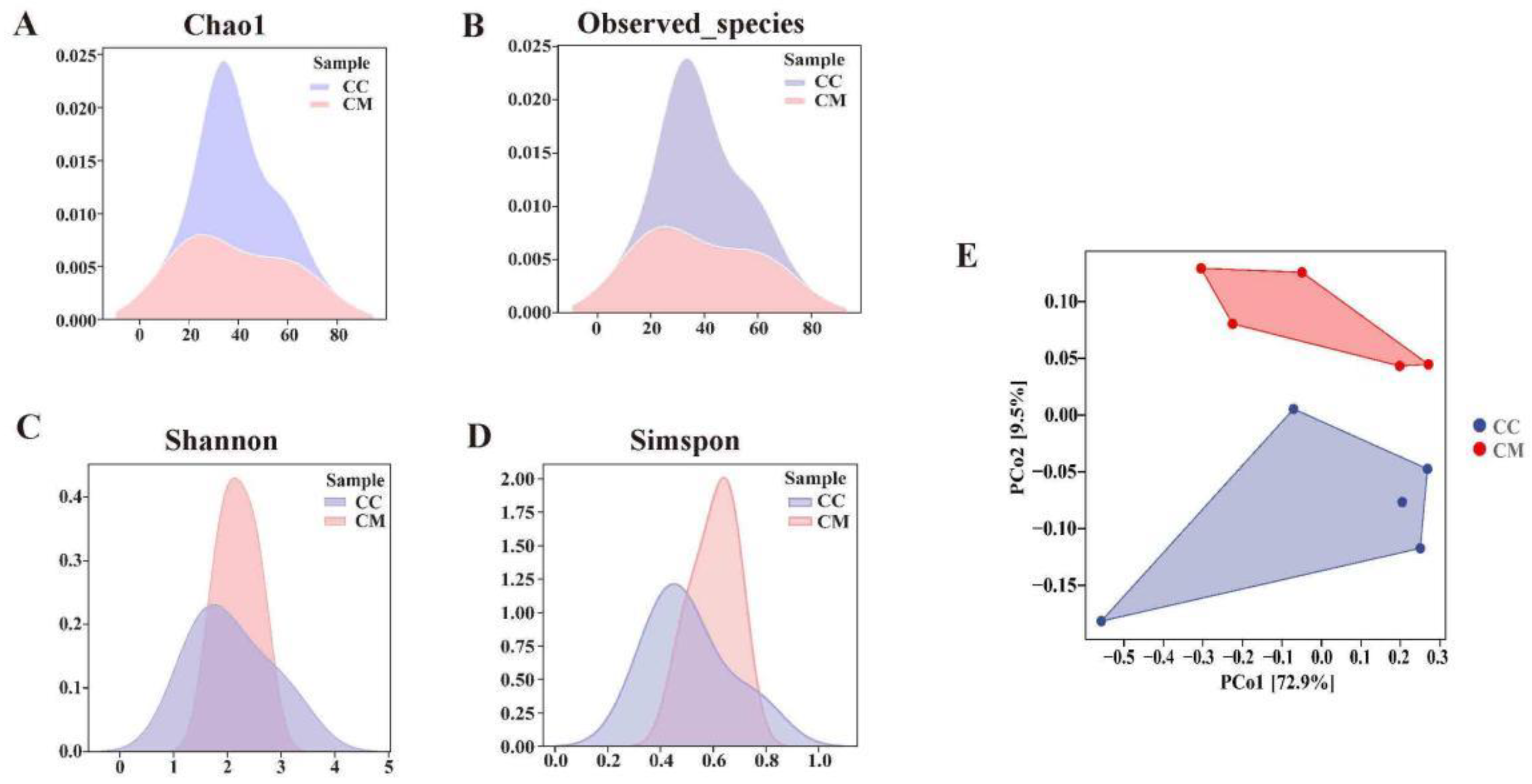
3.4.2. Analysis of the Diversity, Richness and Microbiota Structure of the Gut Content Microbiota in Mice
3.4.3. Analysis of the Dominant Bacteria of Gut Contents in the Mice
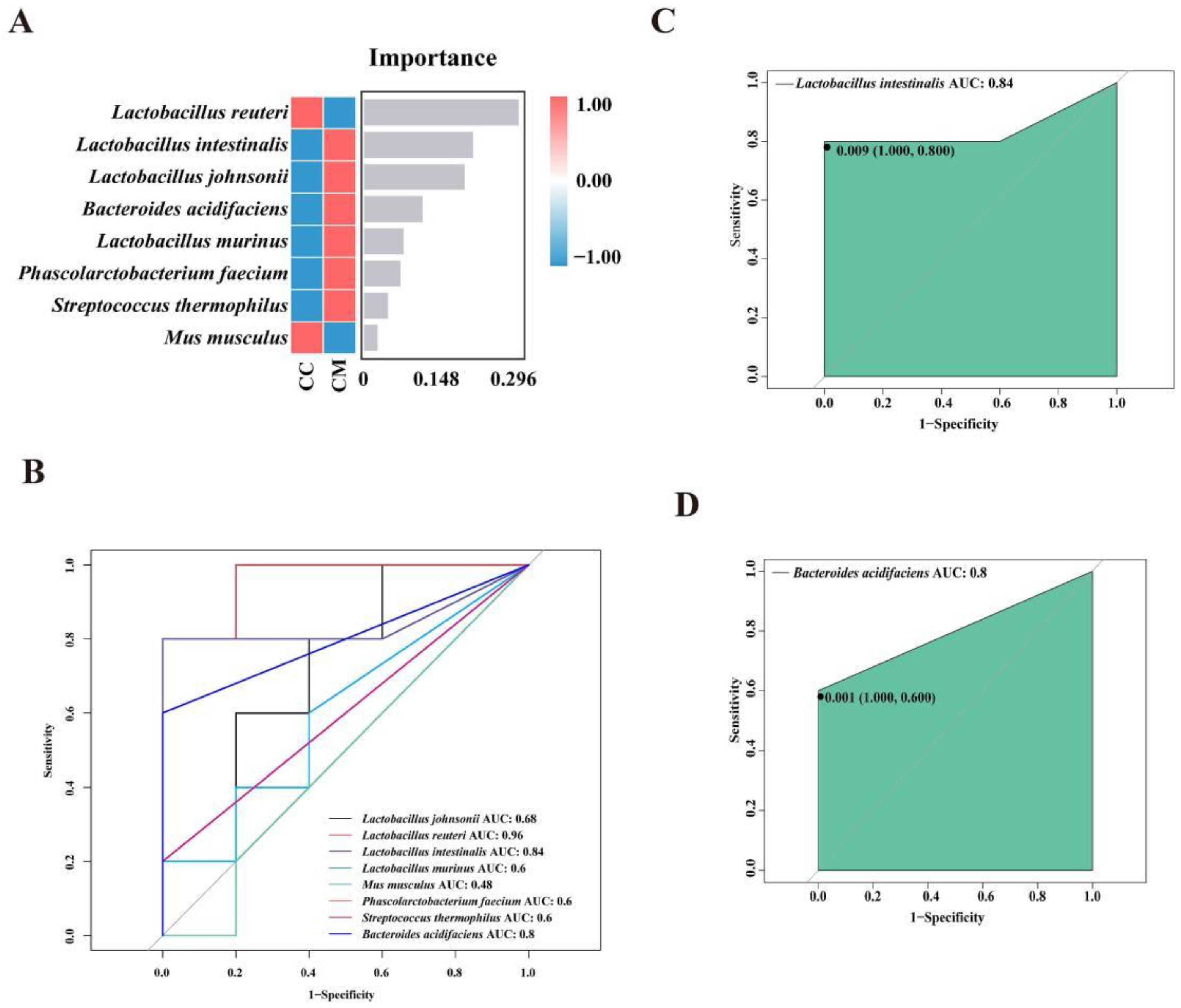
3.4.4. Analysis of the Characteristic Bacteria of the Gut Contents in the Mice
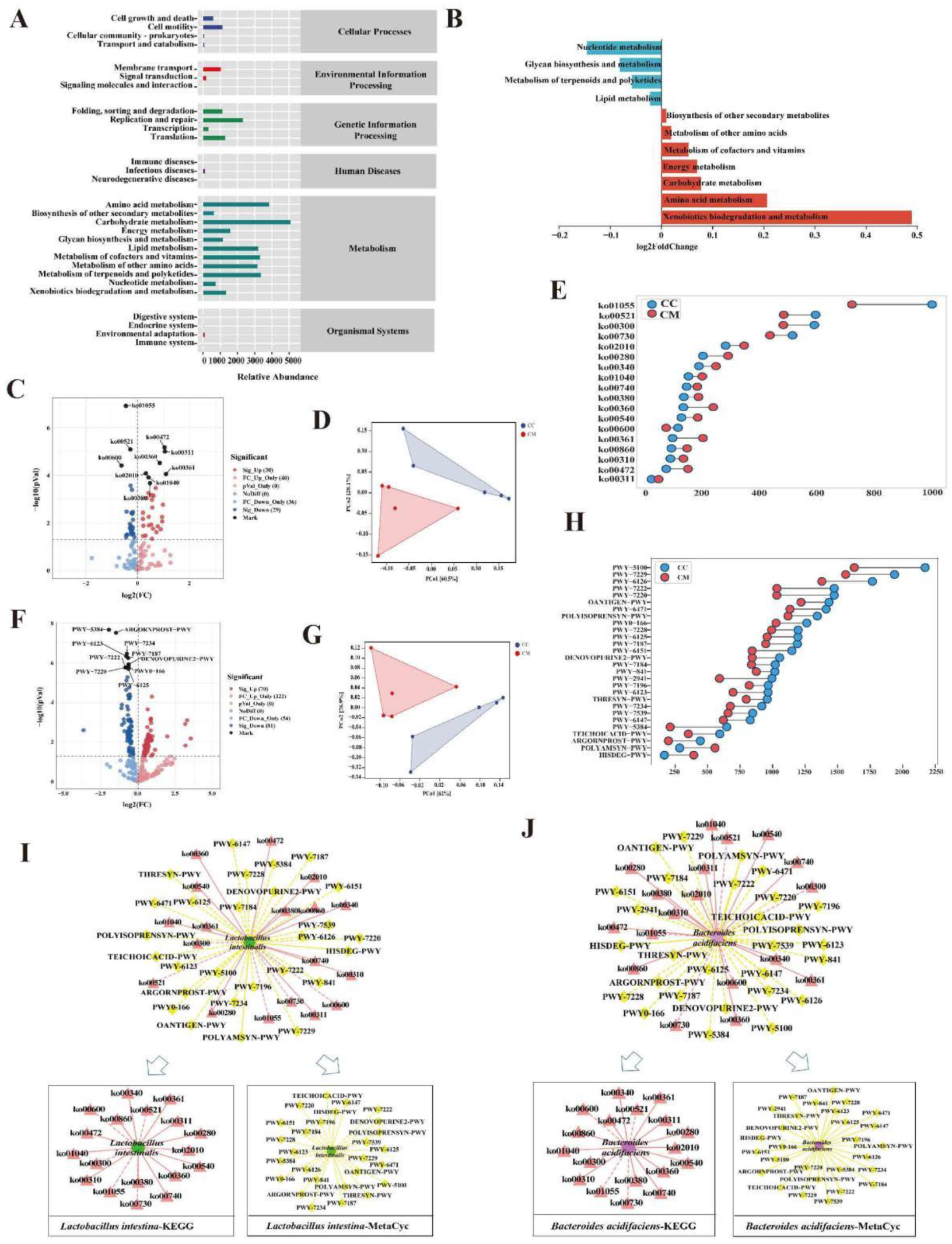
3.4.5. Functional Analysis of the Gut Content Microbiota in the Mice
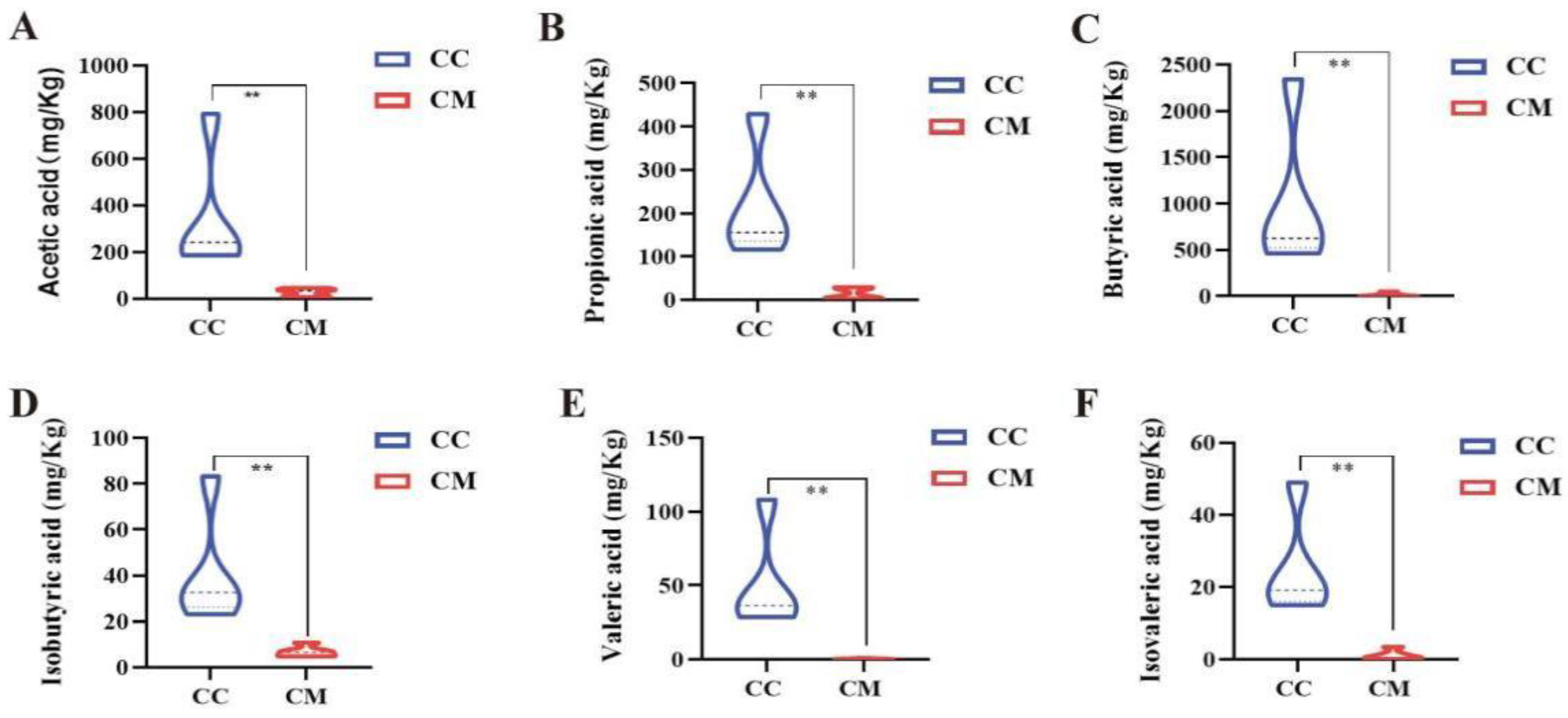
3.5. Changes in the Acetic Acid, Propionic Acid, Butyric Acid, Valeric Acid, Isobutyric Acid and Isovaleric Acid of Gut Contents in the Mice
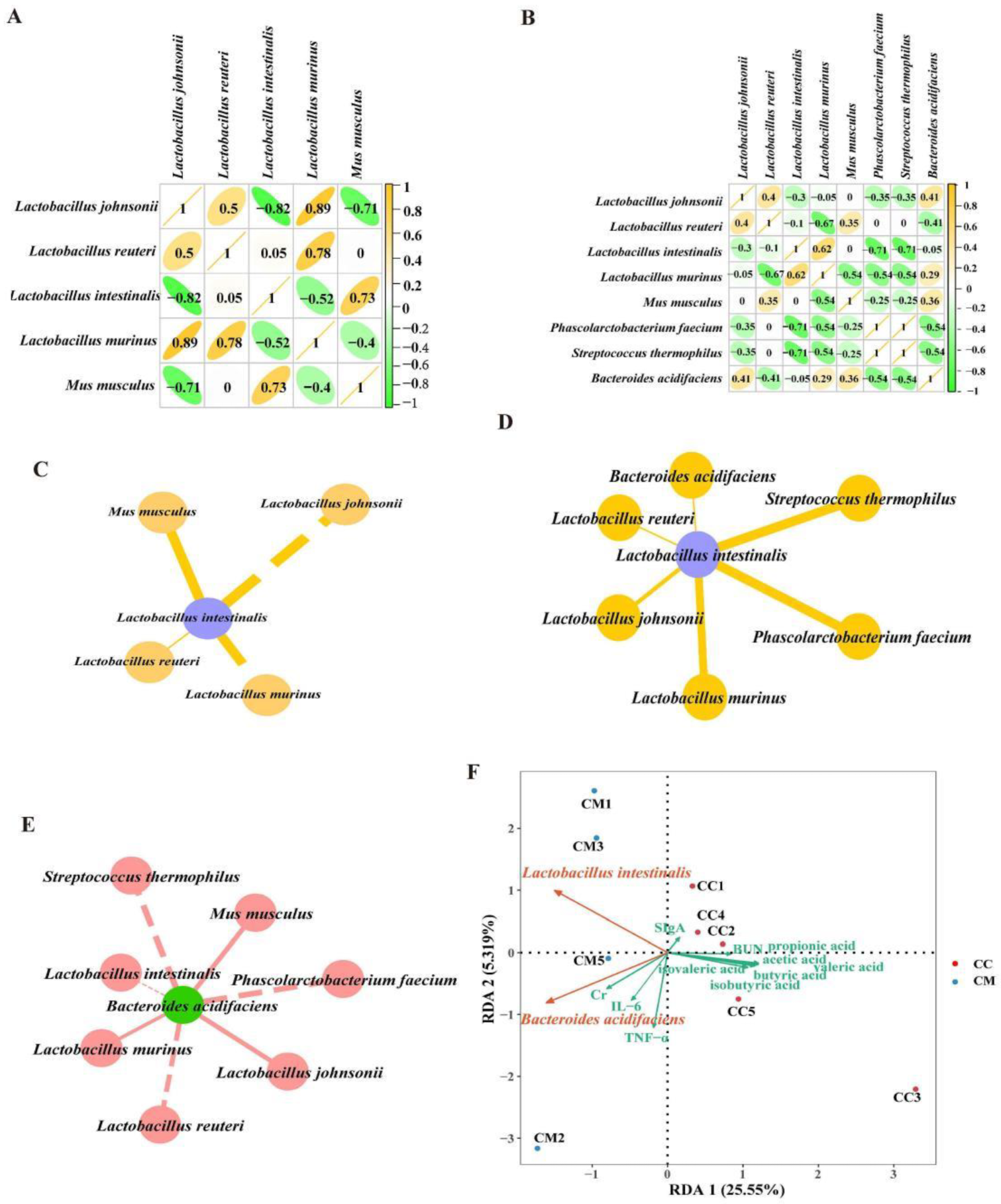
3.6. Correlation Analysis between the Characteristic Bacteria, SCFAs, Intestinal Inflammatory Factors and Kidney Function
4. Discussion
4.1. Disturbed Gut Contents Microbiota in Mice Could Be an Important Factor in the Adenine Combined with Folium sennae-Induced Diarrhea
4.2. Closely Related among the Characteristic Bacteria, SCFAs, Intestinal Inflammatory Response and Kidney Function
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sangnes, D.A.; Dimcevski, G.; Frey, J.; Softeland, E. Diabetic diarrhoea: A study on gastrointestinal motility, pH levels and autonomic function. J. Intern. Med. 2021, 290, 1206–1218. [Google Scholar] [CrossRef] [PubMed]
- Tolaney, S.M.; Beeram, M.; Beck, J.T.; Conlin, A.; Dees, E.C.; Puhalla, S.L.; Rexer, B.N.; Burris, H.A.; Jhaveri, K.; Helsten, T. Abemaciclib in Combination With Endocrine Therapy for patients with hormone receptor-positive, HER2-negative metastatic breast cancer: A phase 1b study. Front Oncol. 2022, 11, 810023. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Li, H.Y.; Ma, R.; Ren, M.H.; Li, Y.; Li, J.X.; Chen, H.; Chen, Z.P.; Gong, D.Y.; Wang, J. Effect of coptis chinensis franch and Magnolia officinalis on intestinal flora and intestinal barrier in a TNBS-induced ulcerative colitis rats model. Phytomedicine 2022, 97, 153927. [Google Scholar] [CrossRef]
- Li, Y.L.; Wu, Y.; Yuan, Z.Y.; Tan, Z.J. Analysis of “treating the same disease with different therapies” in patients with diarrhea. World Chin. J. Digestol. 2022, 28, 1090–1095. [Google Scholar] [CrossRef]
- Pan, J.; Liu, L.H.; Mu, J.W. Research progress of gut microbiota and human health. J. Shandong Normal Univ. 2021, 36, 337–365. [Google Scholar]
- Li, X.Y.; Zhang, C.Y.; Hui, H.Y.; Tan, Z.J. Effect of gegenqinlian decoction on intestinal mucosal flora in mice with diarrhea induced by high temperature and humidity treatment. 3 Biotech. 2021, 11, 83. [Google Scholar] [CrossRef] [PubMed]
- He, H.B.; Wang, M.R.; Chen, L.J.; Liu, S.H.; Chen, S.Z. Effect of Shenling Baizhu Powder on functional diarrhea of spleen and stomach weakness and its influence on intestinal flora. Pract. Clin. J. Integrated Tradit. Chin. West Med. 2021, 21, 16–17. [Google Scholar]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Bilotta, A.J.; Ma, C.; Yang, W.; Yu, Y.; Yu, Y.; Zhao, X.; Zhou, Z.; Yao, S.; Dann, S.M.; Cong, Y. Propionate enhances cell speed and persistence to promote intestinal epithelial turnover and repair. Cell Mol. Gastroenterol. Hepatol. 2021, 11, 1023–1044. [Google Scholar] [CrossRef]
- Jeffery, I.B.; Das, A.O.; Herlihy, E.; Coughlan, S.; Cisek, K.; Moore, M.; Bradley, F.; Carty, T.; Pradhan, M.; Dwibedi, C.; et al. Differences in fecal microbiomes and metabolomes of people with vs without irritable bowel syndrome and bile acid malabsorption. Gastroenterology 2020, 158, 1016–1028.e8. [Google Scholar] [CrossRef] [PubMed]
- Luu, M.; Pautz, S.; Kohl, V.; Singh, R.; Romero, R.; Lucas, S.; Hofmann, J.; Raifer, H.; Vachharajani, N.; Carrascosa, L.C.; et al. The short chain fatty acid pentanoate suppresses autoimmunity by modulating the met-abolic-epigenetic crosstalk in lymphocytes. Nat. Commu. 2019, 10, 760. [Google Scholar]
- Zhang, X.F.; Wang, X.; Wang, H.Q.; Ke, L.J.; Gao, G.Z.; Zhu, L.Y. Human intestinal flora metabolite short-chain fatty acids produced from amino acids and effect on cell permeability. J. Chin. Inst. Food Sci. Technol. 2021, 21, 60–67. [Google Scholar]
- Xu, S.; Lin, L.L.; Wang, S.W. Meetabolic regulation of Wenyun granules on fecal short chain fatty acids in infants witj spleen-deficiency diarrhea. J. Nanjing Univ. Tradit. Chin. Med. 2020, 36, 859–862. [Google Scholar]
- Li, Y.; Hong, G.; Yang, M.; Li, G.; Jin, Y.; Xiong, H.; Qian, W.; Hou, X. Fecal bacteria can predict the efficacy of rifaximin in patients with diarrhea-predominant irritable bowel syndrome. Pharmacol. Res. 2020, 159, 104936. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Chi, L.; Zhu, Y.; Shi, X.; Tu, P.; Li, B.; Yin, J.; Gao, N.; Shen, W.; Schnabl, B. An introduction to next generation sequencing bioinformatic analysis in gut microbiome studies. Biomolecules 2021, 11, 530. [Google Scholar] [CrossRef] [PubMed]
- Ahlawat, S.; Asha; Sharma, K.K. Gut-organ axis: A microbial outreach and networking. Lett. Appl. Microbiol. 2021, 72, 636–668. [Google Scholar] [CrossRef]
- Huang, T.Y.; Li, C.Y.; Liu, D.J. Effect of serum LPa expression level in patients with chronic renal failure on the microecological changes of intestinal flora. Contemp. Med. 2021, 27, 114–115. [Google Scholar]
- Li, J.; Zhu, D.X.; Lu, J.Y.; Zhang, Y.D.; Lu, W.; Lu, J. Alteration of short-chain fatty acids produced by gut microbiota in dogs with chronic renal failure and its effect on renal function. Acta Veterinaria Zootechnica Sinica 2021, 52, 2334–2343. [Google Scholar]
- Jia, K.J.; Jia, T.Z. Comparison of Antidiuretic Activity of Ootheca Mantidis before and after Processing and Its Medicinal Part against Insufficiency of Kidney-Yang and Diuresis Rats. China Pharm. 2016, 27, 879–882. [Google Scholar]
- Zhang, C.Y.; Shao, H.Q.; Li, D.D.; Xiao, N.Q.; Tan, Z.J. Role of tryptophan-metabolizing microbiota in mice diarrhea caused by Folium sennae extracts. BMC Microbiol. 2020, 20, 185. [Google Scholar] [CrossRef]
- Li, X.Y.; Zhu, J.Y.; Wu, Y.; Tan, Z.J. Correlation Between Kidney Function and Intestinal Biological Characteristics of Adenine and Folium Sennae-Induced Diarrhea Model in Mice. Turkish J. Gastroenterol. 2022. [CrossRef]
- Li, X.Y.; Zhu, J.Y.; Wu, Y.; Liu, Y.W.; Hui, H.Y.; Tan, Z.J. Model Building and Validation of Diarrhea Mice with Kidney-yang Depletion Syndrome. J. Tradit. Chin. Med. 2022, 63, 1368–1373. [Google Scholar]
- Xiao, J.; He, L.Q.; Gao, J.D.; Huang, D. Comparison of the model establishment method of syndrome of deficiency of Kidney-yang between adenine and hydrocortisone. Chin. J. Comp. Med. 2008, 18, 77–80. [Google Scholar]
- Li, Y.K. Experimental Methodology of Pharmacology of Traditional Chinese Medicine; Shanghai Science and Technology Press: Shanghai, China, 2006; p. 119. [Google Scholar]
- Wu, Y.; Peng, X.X.; Li, X.Y.; Li, D.D.; Tan, Z.J.; Yu, R. Sex hormones influences intestinal microbiota composition in mice. Front. Microbiol. 2022. [Google Scholar] [CrossRef]
- Xiao, X.Y.; Deng, Y.L.; Liu, Y.J.; Li, D.D.; Tan, Z.J. Effects of folium sennae on blood routine in rats with diarrhea of splenic deficiency type. J. Hubei Univ. Chin. Med. 2016, 18, 49–51. [Google Scholar]
- Yuan, Z.; Zhang, C.Y.; Peng, X.X.; Shu, L.; Long, C.X.; Tan, Z.J. Intestinal microbiota characteristics of mice treated with Folium senna decoction gavage combined with restraint and tail pinch stress. 3 Biotech. 2020, 10, 180. [Google Scholar] [CrossRef]
- Li, X.Y.; Deng, N.; Zheng, T.; Qiao, B.; Peng, M.J.; Xiao, N.Q.; Tan, Z.J. Importance of Dendrobium officinale in improving the adverse effects of high-fat diet on mice associated with intestinal contents microbiota. Front Nutr. 2022, 9, 957334. [Google Scholar] [CrossRef]
- D’Amico, G. Natural history of idiopathic IgA nephropathy: Role of clinical and histological prognostic factors. Am. J. Kidney Dis. 2000, 36, 227–237. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach Learn 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.C.; Yang, C.; Guo, L.Q.; Chen, J.; Xiong, D.X. Prediction model of acute exacerbation of chronic obstructive pulmonary disease based on machine learning. Chin. J. Rehabil. Theory Pract. 2022, 28, 678–683. [Google Scholar]
- Zhang, C.Y.; Peng, X.X.; Shao, H.Q.; Li, X.Y.; Wu, Y.; Tan, Z.J. Gut Microbiota Comparison Between Intestinal Contents and Mucosa in Mice With Repeated Stress-Related Diarrhea Provides Novel Insight. Front. Microbiol. 2021, 12, 626691. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Xu, J.; Liu, C.L.; Shi, K.; Sun, X.J.; Song, C.C.; Xu, K.; Liu, Y.J. Atractyloside-A ameliorates spleen deficiency diarrhea by interfering with TLR4/MyD88/NF-κB signaling activation and regulating intestinal flora homeostasis. Int. Immunopharmacol. 2022, 107, 108679. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.D.; Yang, Y.; Lin, X.J.; Chen, P.W.; Ye, L.Y.; Zeng, L.Y.; Ye, Q.N.; Yang, X.N.; Ceng, J.T.; Shan, J.Y.; et al. Qiweibaizhu decoction treats diarrheal juvenile rats by modulating the gut microbiota, short-chain fatty acids, and the mucus barrier. Evid. Based Complement Alternat. Med. 2021, 2021, 8873294. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Jin, J.; Su, X.; Yin, X.; Gao, J.; Wang, X.; Zhang, S.; Bu, P.; Wang, M.; Zhang, Y.; et al. Intestinal flora modulates blood pressure by regulating the synthesis of intestinal-derived corticosterone in high salt-Induced hypertension. Circ. Res. 2020, 126, 839–853. [Google Scholar] [CrossRef]
- Ma, Z.M.; Wu, L.; Gao, L.Y. Microbial Secondary Metabolites and Their Application in Fertilizers. Fertil. Health 2022, 49, 1–6. [Google Scholar]
- Zhang, M.; Yang, L.; Zhu, M.; Yang, B.; Yang, Y.; Jia, X.; Feng, L. Moutan cortex polysaccharide ameliorates diabetic kidney disease via modulating gut microbiota dynamically in rats. Int. J. Biol. Macromol. 2022, 206, 849–860. [Google Scholar] [CrossRef]
- Deng, L.; Wojciech, L.; Png, C.W.; Koh, E.Y.; Aung, T.T.; Kioh, D.Y.Q.; Chan, E.C.Y.; Malleret, B.; Zhang, Y.; Peng, G.; et al. Experimental colonization with blastocystis ST4 is associated with protective immune responses and modulation of gut microbiome in a DSS-induced colitis mouse model. Cell Mol. Life Sci. 2022, 79, 245. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, W.L.; Pei, L.X.; Wu, X.L.; Geng, H.; Guo, J.; Sun, J.H. Effect of Tiaoshen Jianpi acupuncture therapy on gut microbiota and fecal short-chain fatty acids in patients with diarrhea type irritable bowel syndrome. Chin. Acupunct. Moxibustion 2021, 41, 137–141. [Google Scholar]
- Jacobson, A.; Lam, L.; Rajendram, M.; Tamburini, F.; Honeycutt, J.; Pham, T.; Treuren, W.V.; Pruss, K.; Stabler, S.R.; Lugo, K.; et al. Gut Commensal-Produced Metabolite Mediates Colonization Resistance to Salmonella Infection. Cell Host Microbe 2018, 24, 296–307.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Wu, L.Y.; Dou, C.Z.; Guan, X.; Wu, H.G.; Liu, H.R. Research Advance in Intestinal Mucosal Barrier and Pathogenesis of Crohn’s Disease. Gastroenterol. Res. Pract. 2016, 2016, 9686238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AldarsGarcía, L.; Marin, A.; Chaparro, M.; Gisbert, J.P. The Interplay between Immune System and Microbiota in Inflammatory Bowel Disease: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 3076. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.W.; Zhao, Q.; Huang, Q.W.; Zhao, Z.H.; Zhou, H.Y.; He, Y.Y.; Li, S.S.; Wan, S.F. Modified Renshen Wumei decoction alleviates intestinal barrier destruction in rats with diarrhoea. J. Microbiol. Biotechnol. 2021, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.C.; Ma, P. Effects and mechanism of gross saponins of tribulus terrestris on IL-1β, IL-6, TNF-α, IL-2 and NO secreted by LPS-induced macrophages. Chin. J. Immunol. 2021, 37, 1958–1963. [Google Scholar]
- Dai, S.J.; Shi, X.K.; Tong, L.; Zhang, J.; Shi, J.Y.; Wang, Y. The effect of Carthamus tinctorius polysaccharides on intestinal mucosa sIgA, plasma endotoxin and intestinal flora in mice with intestinal microecological disorders. Chin. J. Microecol. 2022, 34, 12–17. [Google Scholar]
- Han, F.; Wang, X.; Guo, J.; Qi, C.; Xu, C.; Luo, Y.; Li, E.; Qin, J.G.; Chen, L. Effects of glycinin and β-conglycinin on growth performance and intestinal health in juvenile Chinese mitten crabs (Eriocheir sinensis). Fish Shellfish. Immunol. 2019, 84, 269–279. [Google Scholar] [CrossRef]
- Busbee, P.B.; Menzel, L.; Alrafas, H.R.; Dopkins, N.; Becker, W.; Miranda, K.; Tang, C.; Chatterjee, S.; Singh, U.; Nagarkatti, M.; et al. Indole-3-carbinol prevents colitis and associated microbial dysbiosis in an IL-22–dependent manner. JCI Insight 2020, 5, e127551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, D.; Stecher, B.; Schintlmeister, A.; Reichert, J.; Brugiroux, S.; Wild, B.; Wanek, W.; Richter, A.; Rauch, I.; Decker, T.; et al. Host-compound foraging by intestinal microbiota revealed by single-cell stable isotope probing. Proc. Natl. Acad. Sci. USA 2013, 110, 4720–4725. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Wang, L.; Qian, H.Y.; Zhang, F.; Dong, B.R.; Shu, W. Research progress of creatinine as biomarkers in disease diagnosis and drug evaluation. Drug Eva. Res. 2021, 44, 2007–2012. [Google Scholar]
- Ma, Y.; Bao, G.X. Serum urea nitrogen, creatinine, β2- microglobulin combined detection in diagnostic value of renal injury in elderly patients with essential hypertension. Mod. Med. Health Res. Electron. J. 2021, 5, 26–28. [Google Scholar]
- Devlin, A.S.; Marcobal, A.; Dodd, D.; Nayfach, S.; Plummer, N.; Meyer, T.; Pollard, K.S.; Sonnenburg, J.L.; Fischbach, M.A. Modulation of a Circulating Uremic Solute via Rational Genetic Manipulation of the Gut Microbiota. Cell Host Microbe 2016, 20, 709–715. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Steps | Conditions |
|---|---|
| Column temperature requirement | 100 °C (5 min)-5 °C/min-150 °C (0 min)-30 °C/min-240 °C (30 min) |
| Flow rate requirements | 1 mL/min |
| Shunt ratio | 75:1 |
| Carrier gas | Helium |
| Chromatographic column | TG WAX 30 m × 0.25 mm × 0.25 μm |
| Injector | 240 °C |
| Mass spectrometry EI source, bombardment voltage | 70 eV |
| Single ion scan mode | Quantitative ion 63, 70 |
| Ion source temperature | 200 °C |
| Connection line temperature | 250 °C |
| Group | The 1st Day of Modeling | The 5th Day of Modeling | The 9th Day of Modeling | The 13th Day of Modeling |
|---|---|---|---|---|
| CC | 24.73 ± 0.49 | 31.31 ± 1.34 | 35.25 ± 2.21 | 38.95 ± 2.18 |
| CM | 26.10 ± 0.81 | 31.61 ± 0.92 | 34.10 ± 1.00 ** | 34.03 ± 0.55 ** |
| Group | Glomerular Pathology Score | Tubular and Interstitial Renal Pathology Score |
|---|---|---|
| CC | 0.67 ± 0.47 | 0.67 ± 0.47 |
| CM | 4.67 ± 0.47 * | 3.67 ± 0.47 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Peng, X.; Qiao, B.; Peng, M.; Deng, N.; Yu, R.; Tan, Z. Gut-Kidney Impairment Process of Adenine Combined with Folium sennae-Induced Diarrhea: Association with Interactions between Lactobacillus intestinalis, Bacteroides acidifaciens and Acetic Acid, Inflammation, and Kidney Function. Cells 2022, 11, 3261. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11203261
Li X, Peng X, Qiao B, Peng M, Deng N, Yu R, Tan Z. Gut-Kidney Impairment Process of Adenine Combined with Folium sennae-Induced Diarrhea: Association with Interactions between Lactobacillus intestinalis, Bacteroides acidifaciens and Acetic Acid, Inflammation, and Kidney Function. Cells. 2022; 11(20):3261. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11203261
Chicago/Turabian StyleLi, Xiaoya, Xinxin Peng, Bo Qiao, Maijiao Peng, Na Deng, Rong Yu, and Zhoujin Tan. 2022. "Gut-Kidney Impairment Process of Adenine Combined with Folium sennae-Induced Diarrhea: Association with Interactions between Lactobacillus intestinalis, Bacteroides acidifaciens and Acetic Acid, Inflammation, and Kidney Function" Cells 11, no. 20: 3261. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11203261