The Inhibition on MDFIC and PI3K/AKT Pathway Caused by miR-146b-3p Triggers Suppression of Myoblast Proliferation and Differentiation and Promotion of Apoptosis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cell Culture
2.3. RNA Isolation, Complementary DNA (cDNA) Synthesis, and Quantitative Real-Time PCR (q-PCR)
2.4. RNA Oligonucleotides and Plasmids Construction
2.5. MiRNA Targets Prediction and RNA Hybrid Detection
2.6. Cell Transfection
2.7. Dual-Luciferase Reporter Assay
2.8. Immunofluorescence
2.9. 5-Ethynyl-2′-deoxyuridine (EdU) Assay
2.10. Flow Cytometric Analysis of Cell Cycle
2.11. Flow Cytometric Analysis of Cell Apoptosis
2.12. Western Blot Assay
2.13. Statistical Analysis
3. Results
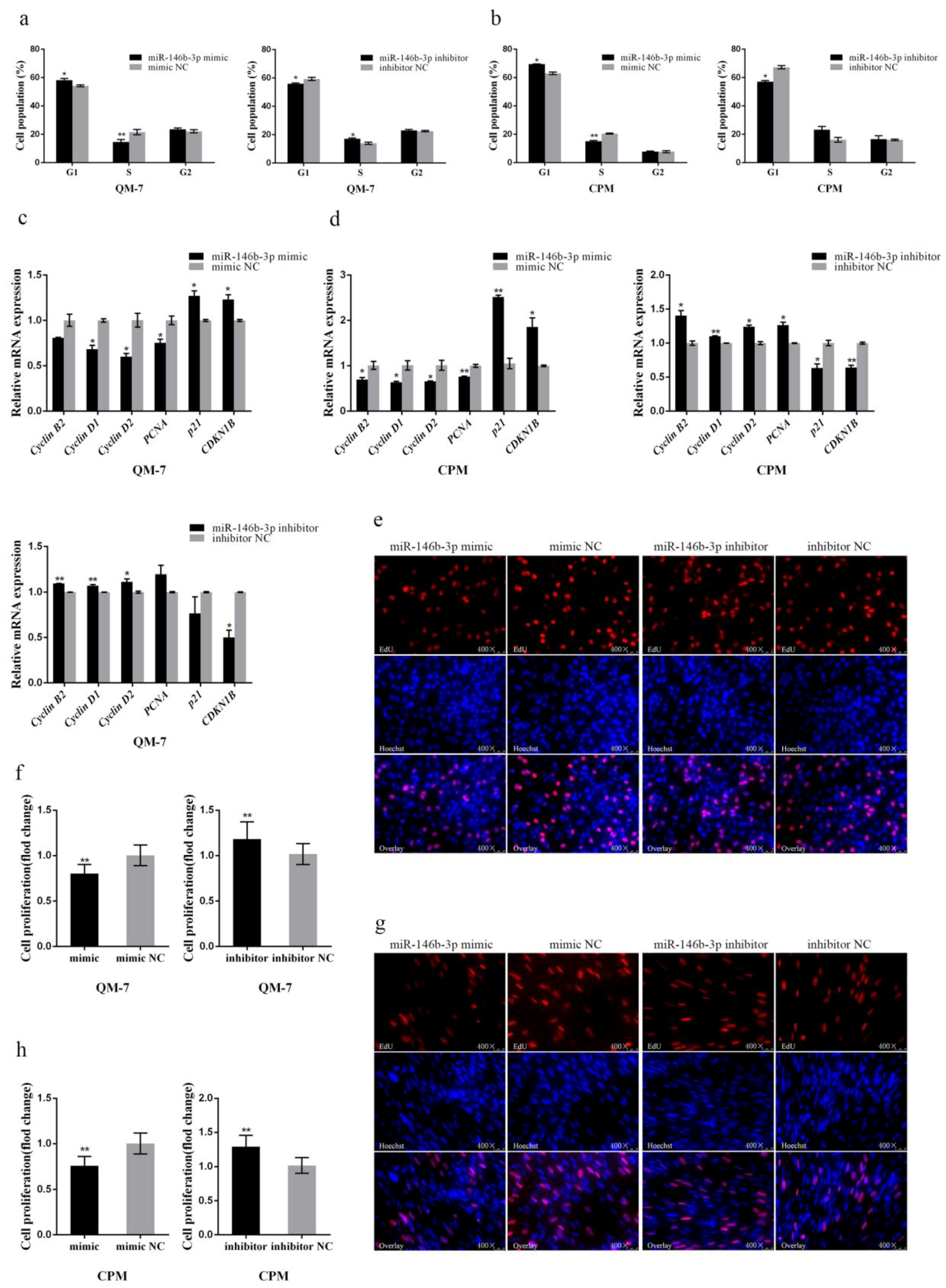
3.1. miR-146b-3p Contributes to Cell Cycle Arrest in Myoblast and Suppresses Cell Proliferation
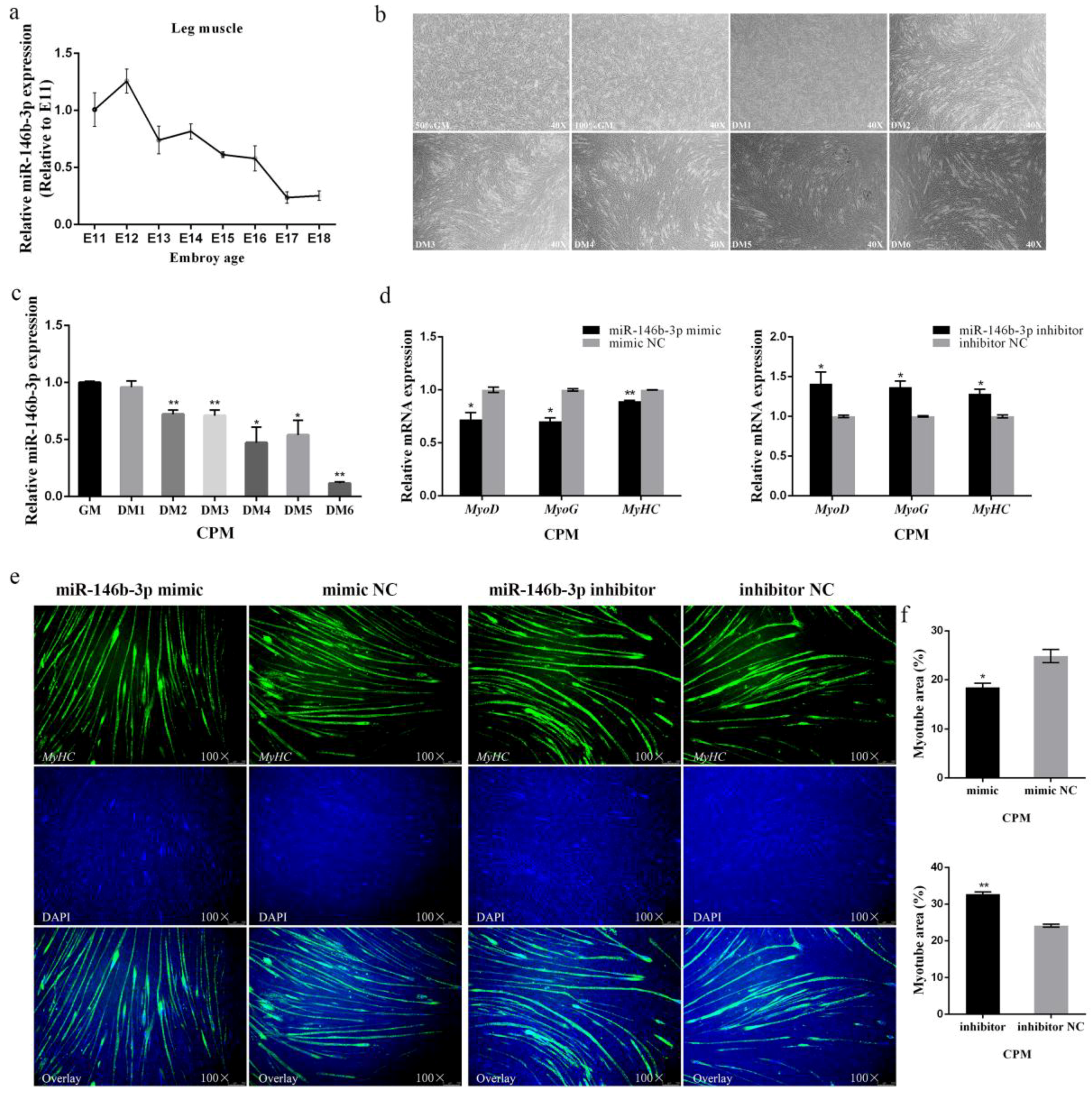
3.2. miR-146b-3p Inhibits Myoblast Differentiation
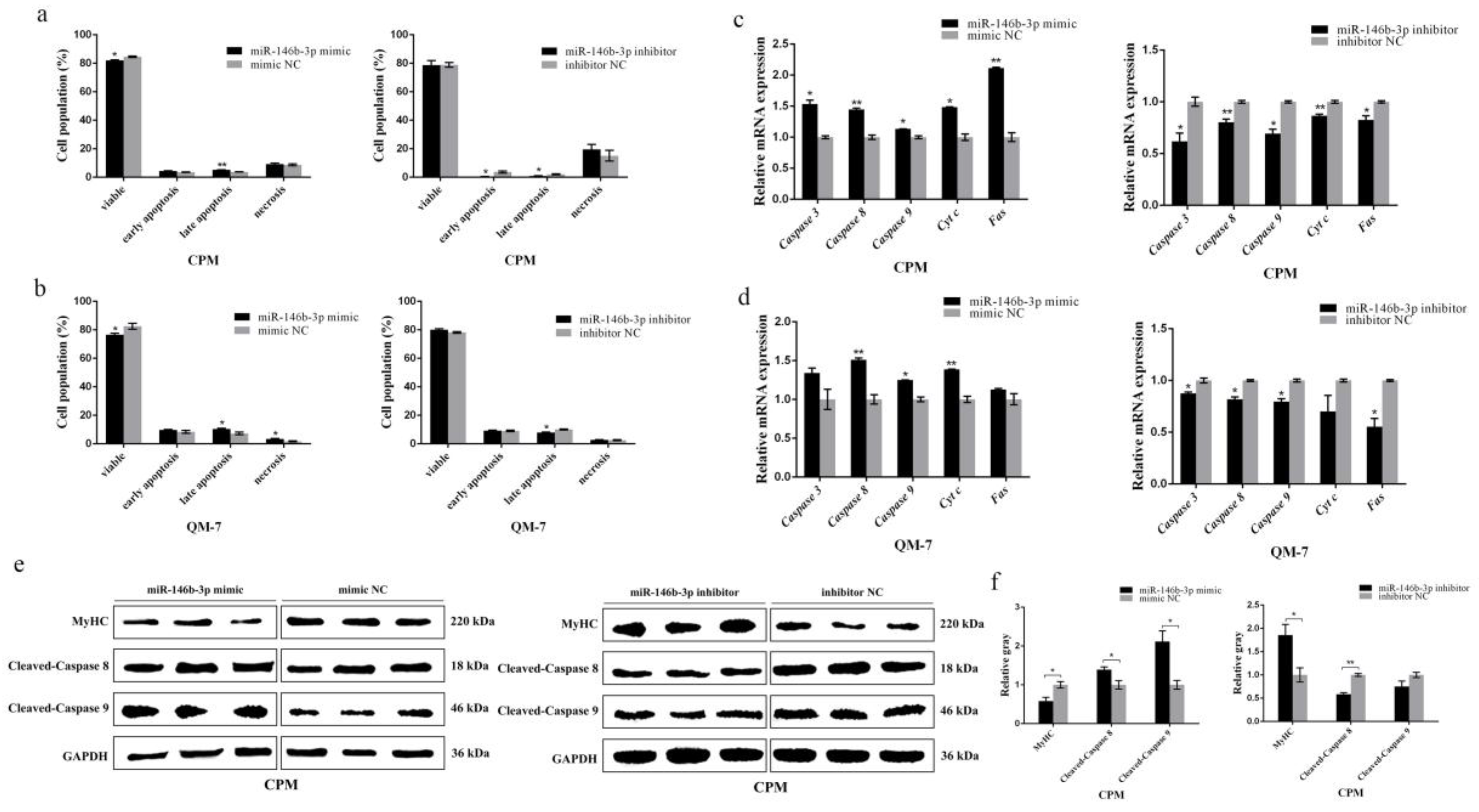
3.3. miR-146b-3p Promotes Myoblast Apoptosis
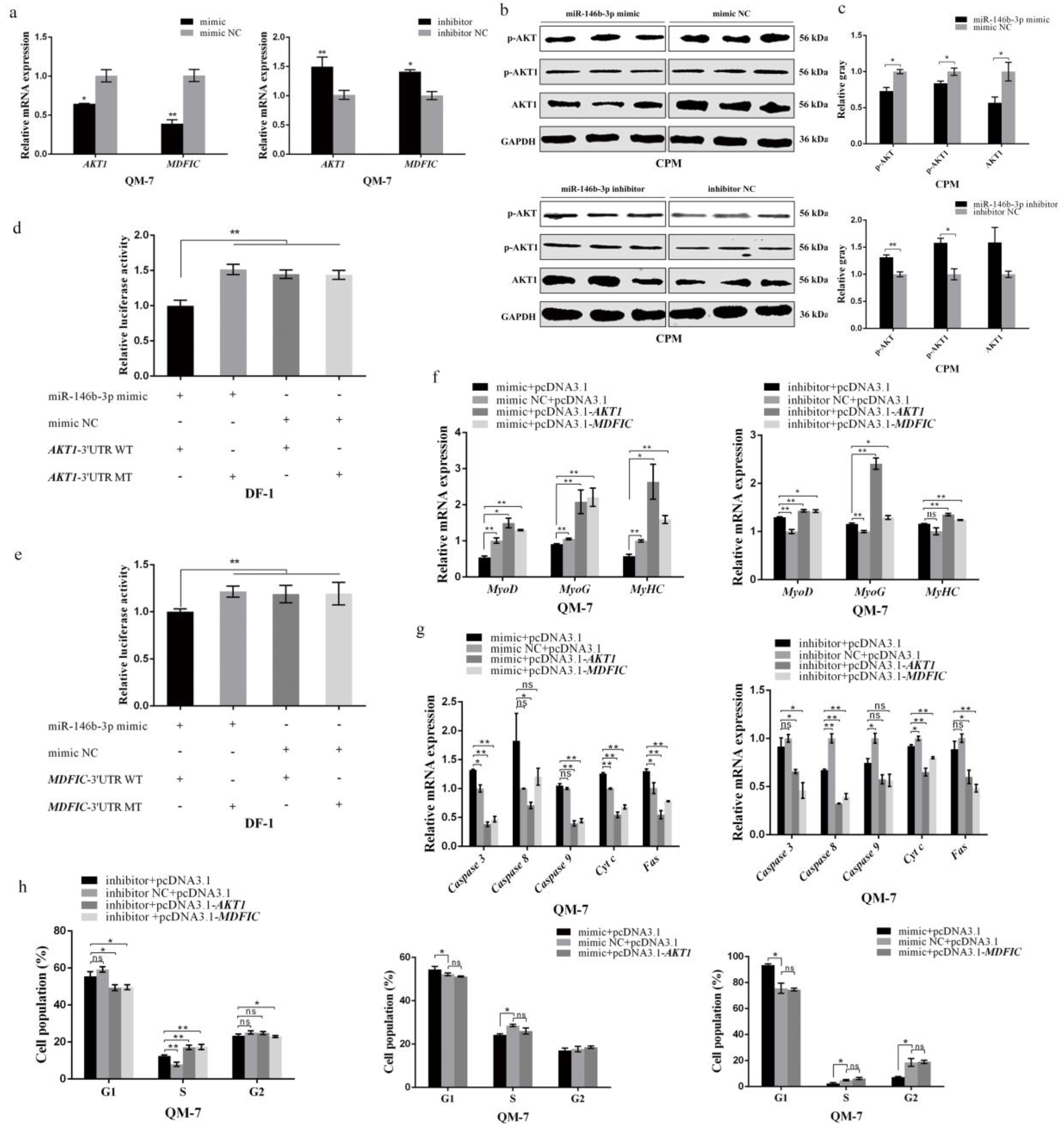
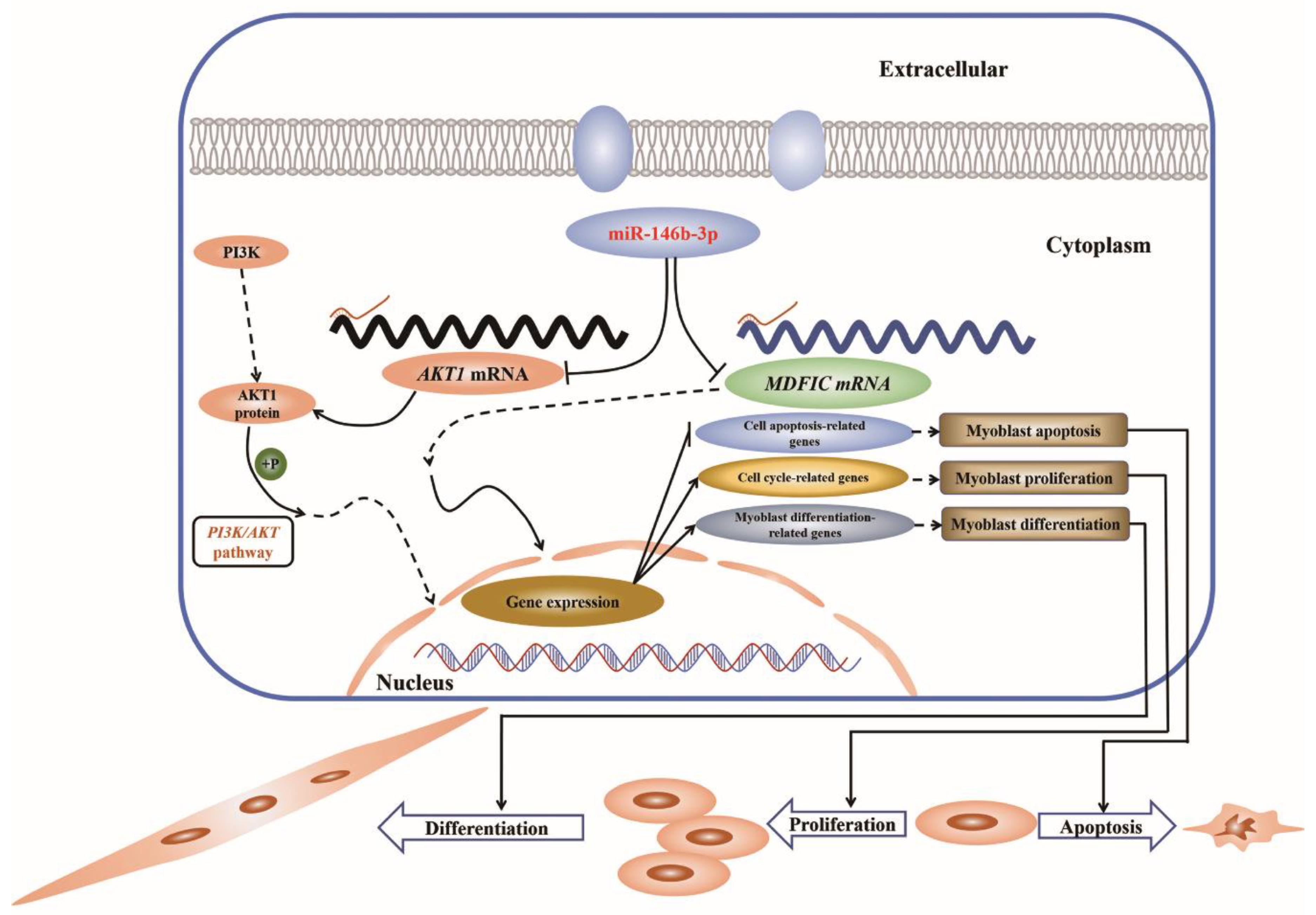
3.4. miR-146b-3p Targets AKT1 and MDFIC and Downregulates PI3K/AKT Pathway Activity
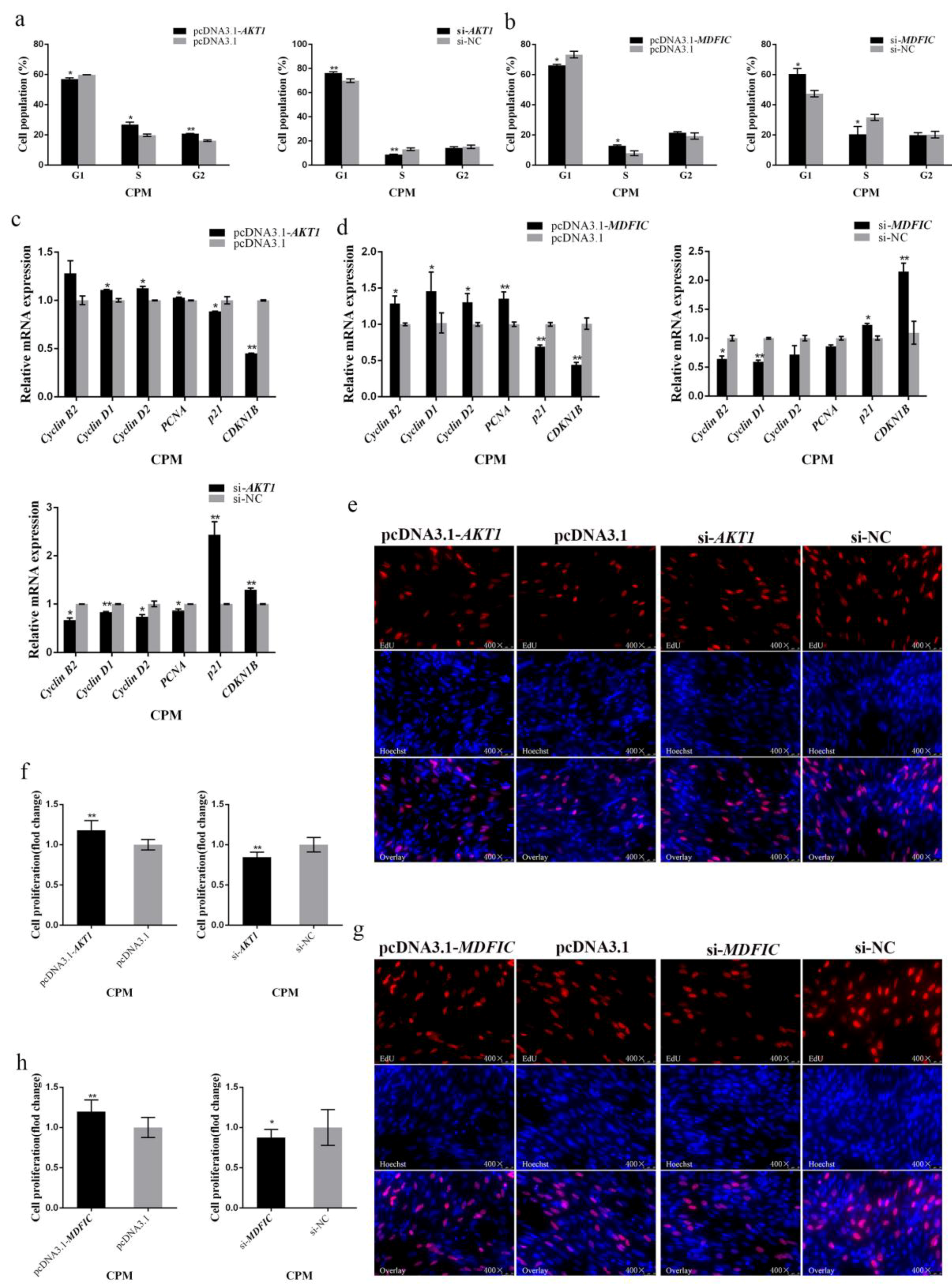
3.5. Both AKT1 and MDFIC Can Facilitate the Proliferation of Myoblast
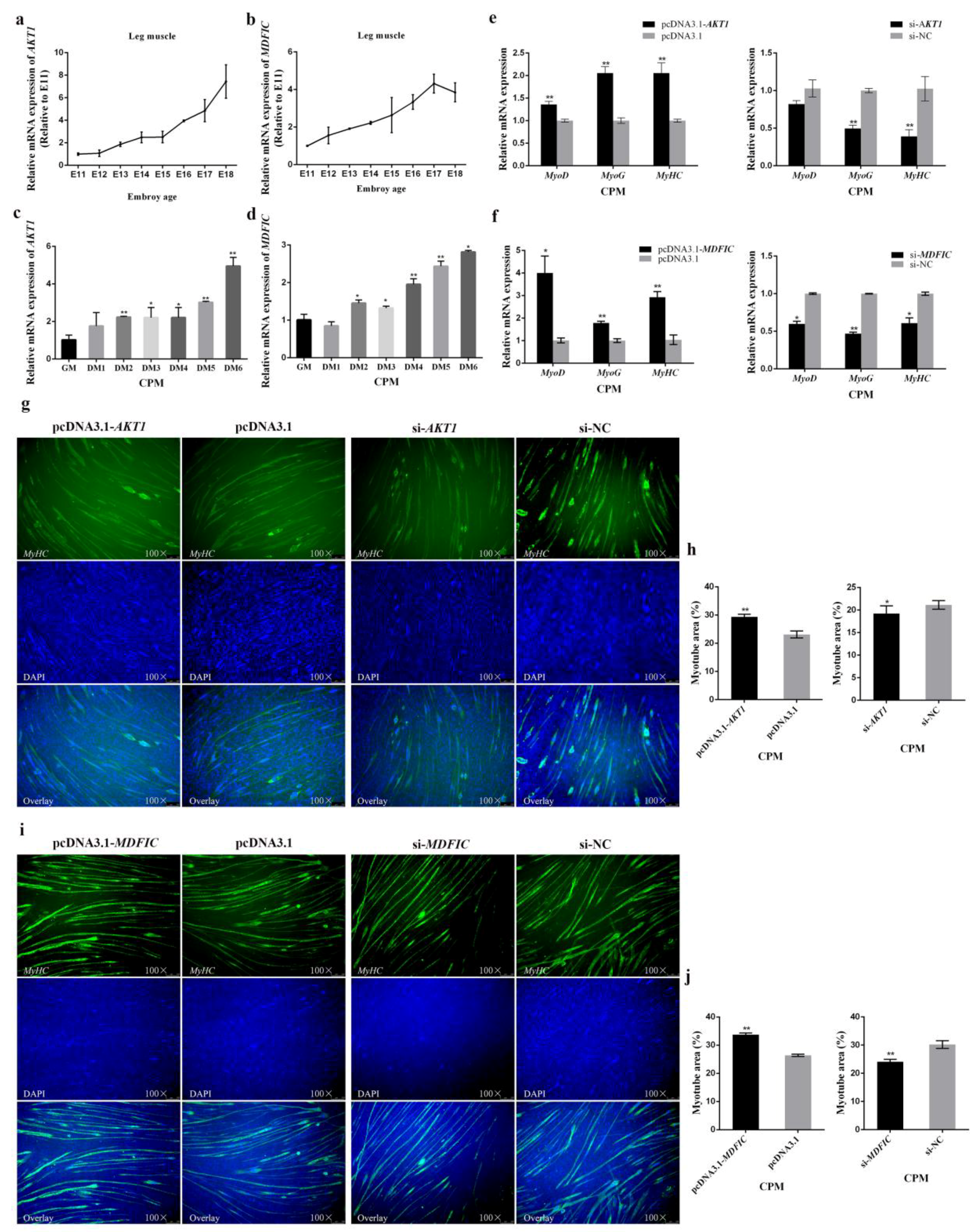
3.6. In Terms of Myoblast Differentiation, Both AKT1 and MDFIC Exhibit Positive Impacts
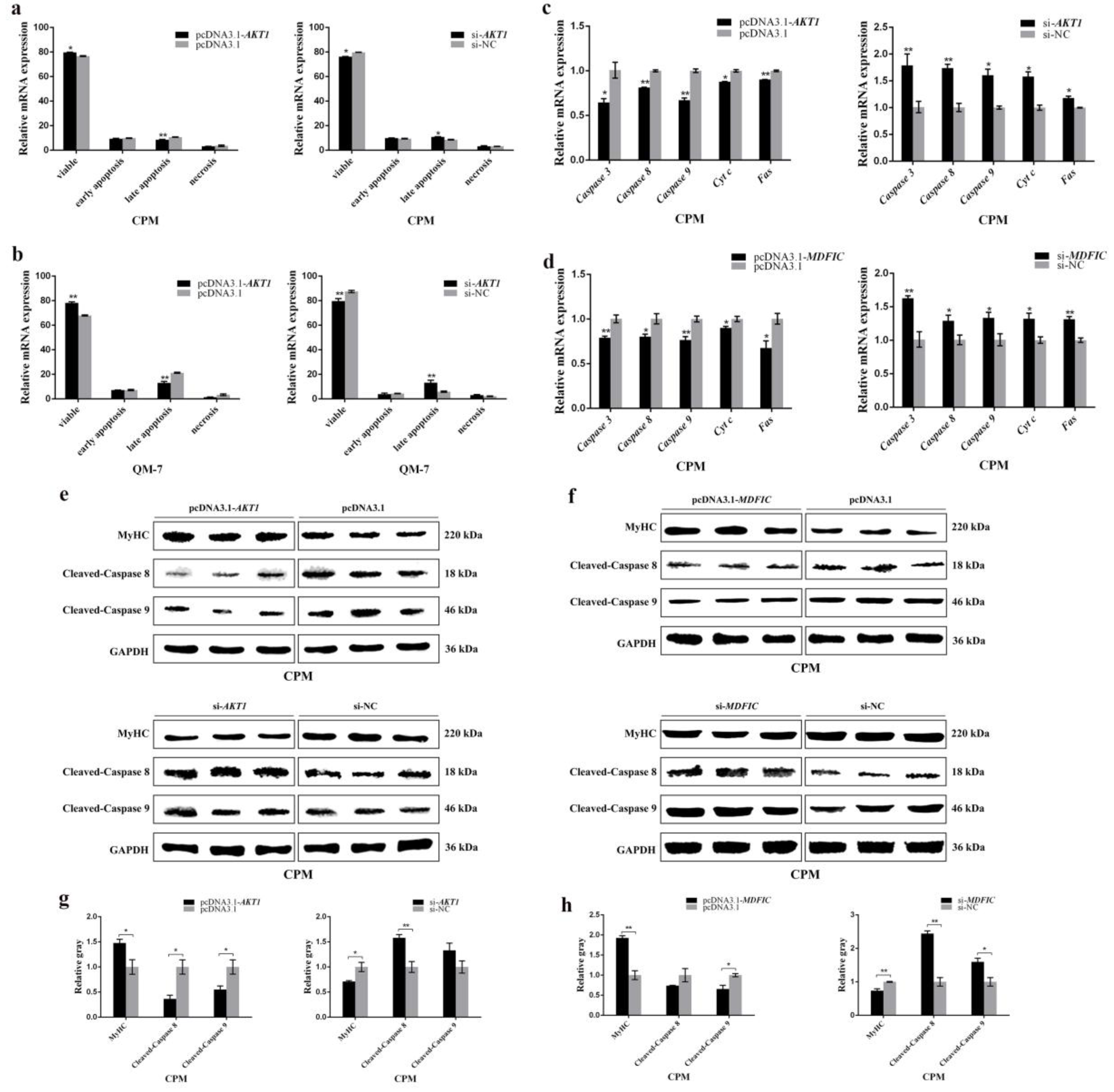
3.7. AKT1 Shows an Inhibitory Effect on Myoblast Apoptosis and so Does MDFIC
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Buckingham, M. Skeletal muscle formation in vertebrates. Curr. Opin. Genet. Dev. 2001, 11, 440–448. [Google Scholar] [CrossRef]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, Y.; Steller, H. Programmed cell death in animal development and disease. Cell 2011, 147, 742–758. [Google Scholar] [CrossRef] [PubMed]
- Kaczanowski, S. Apoptosis: Its origin, history, maintenance and the medical implications for cancer and aging. Phys. Biol. 2016, 13, 31001. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. Tumor resistance to apoptosis. Int. J. Cancer 2009, 124, 511–515. [Google Scholar] [CrossRef]
- Dahmane, G.R.; Erzen, I.; Holcman, A.; Skorjanc, D. Effects of divergent selection for 8-week body weight on postnatal enzyme activity pattern of 3 fiber types in fast muscles of male broilers (Gallus gallus domesticus). Poult. Sci. 2010, 89, 2651–2659. [Google Scholar] [CrossRef]
- Perry, R.L.; Rudnick, M.A. Molecular mechanisms regulating myogenic determination and differentiation. Front. Biosci. 2000, 5, D750–D767. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Lai, E.C. Micro RNAs are complementary to 3’ UTR sequence motifs that mediate negative post-transcriptional regulation. Nat. Genet. 2002, 30, 363–364. [Google Scholar] [CrossRef]
- Lewis, B.P.; Shih, I.H.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of mammalian microRNA targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef]
- Stefani, G.; Slack, F.J. Small non-coding RNAs in animal development. Nat. Rev. Mol. Cell Biol. 2008, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Felekkis, K.; Touvana, E.; Stefanou, C.; Deltas, C. microRNAs: A newly described class of encoded molecules that play a role in health and disease. Hippokratia 2010, 14, 236–240. [Google Scholar] [PubMed]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Lee, Y.S.; Sivaprasad, U.; Malhotra, A.; Dutta, A. Muscle-specific microRNA miR-206 promotes muscle differentiation. J. Cell Biol. 2006, 174, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Tao, Y.; Li, J.; Deng, Z.; Yan, Z.; Xiao, X.; Wang, D.Z. microRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J. Cell Biol. 2010, 190, 867–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Y.; Chen, J. MicroRNAs in skeletal myogenesis. Cell Cycle 2011, 10, 441–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Wu, H.; Ye, Y.; Li, Z.; Hao, S.; Kong, L.; Zheng, X.; Lin, S.; Nie, Q.; Zhang, X. The transient expression of miR-203 and its inhibiting effects on skeletal muscle cell proliferation and differentiation. Cell Death Dis. 2014, 5, e1347. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Ouyang, H.; Abdalla, B.A.; Xu, H.; Nie, Q.; Zhang, X. miR-16 controls myoblast proliferation and apoptosis through directly suppressing Bcl2 and FOXO1 activities. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 674–684. [Google Scholar] [CrossRef]
- Cai, B.; Ma, M.; Chen, B.; Li, Z.; Abdalla, B.A.; Nie, Q.; Zhang, X. MiR-16-5p targets SESN1 to regulate the p53 signaling pathway, affecting myoblast proliferation and apoptosis, and is involved in myoblast differentiation. Cell Death Dis. 2018, 9, 367. [Google Scholar] [CrossRef]
- Chen, B.; Yu, J.; Guo, L.; Byers, M.; Wang, Z.; Chen, X.; Xu, H.; Nie, Q. Circular RNA circHIPK3 Promotes the Proliferation and Differentiation of Chicken Myoblast Cells by Sponging miR-30a-3p. Cells 2019, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.; Li, Z.; Abdalla, B.A.; Chen, Y.; Nie, Q. MiR-34b-5p Mediates the Proliferation and Differentiation of Myoblasts by Targeting IGFBP2. Cells 2019, 8, 360. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ouyang, H.; Wang, Z.; Chen, B.; Nie, Q. A Novel Circular RNA Generated by FGFR2 Gene Promotes Myoblast Proliferation and Differentiation by Sponging miR-133a-5p and miR-29b-1-5p. Cells 2018, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Stahle, M.; Pivarcsi, A. MicroRNAs and immunity: Novel players in the regulation of normal immune function and inflammation. Semin. Cancer Biol. 2008, 18, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Hurst, D.R.; Edmonds, M.D.; Scott, G.K.; Benz, C.C.; Vaidya, K.S.; Welch, D.R. Breast Cancer Metastasis Suppressor 1 Up-regulates miR-146, Which Suppresses Breast Cancer Metastasis. Cancer Res. 2009, 69, 1279–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katakowski, M.; Buller, B.; Zheng, X.; Lu, Y.; Rogers, T.; Osobamiro, O.; Shu, W.; Jiang, F.; Chopp, M. Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett. 2013, 335, 201–204. [Google Scholar] [CrossRef] [Green Version]
- Jebessa, E.; Ouyang, H.; Abdalla, B.A.; Li, Z.; Abdullahi, A.Y.; Liu, Q.; Nie, Q.; Zhang, X. Characterization of miRNA and their target gene during chicken embryo skeletal muscle development. Oncotarget 2018, 9, 17309–17324. [Google Scholar] [CrossRef]
- Ouyang, H.; He, X.; Li, G.; Xu, H.; Jia, X.; Nie, Q.; Zhang, X. Deep Sequencing Analysis of miRNA Expression in Breast Muscle of Fast-Growing and Slow-Growing Broilers. Int. J. Mol. Sci. 2015, 16, 16242–16262. [Google Scholar] [CrossRef] [Green Version]
- Brazil, D.P.; Yang, Z.Z.; Hemmings, B.A. Advances in protein kinase B signalling: AKTion on multiple fronts. Trends Biochem. Sci. 2004, 29, 233–242. [Google Scholar] [CrossRef]
- Wu, M.; Falasca, M.; Blough, E.R. Akt/protein kinase B in skeletal muscle physiology and pathology. J. Cell. Physiol. 2011, 226, 29–36. [Google Scholar] [CrossRef]
- Wilson, E.M.; Rotwein, P. Selective control of skeletal muscle differentiation by Akt1. J. Biol. Chem. 2007, 282, 5106–5110. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.S.; Xu, P.Z.; Gottlob, K.; Chen, M.L.; Sokol, K.; Shiyanova, T.; Roninson, I.; Weng, W.; Suzuki, R.; Tobe, K.; et al. Growth retardation and increased apoptosis in mice with homozygous disruption of the Akt1 gene. Genes Dev. 2001, 15, 2203–2208. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.G.; Wagner, A.J.; Conzen, S.D.; Jordan, J.; Bellacosa, A.; Tsichlis, P.N.; Hay, N. The PI 3-kinase/Akt signaling pathway delivers an anti-apoptotic signal. Genes Dev. 1997, 11, 701–713. [Google Scholar] [CrossRef]
- Romashkova, J.A.; Makarov, S.S. NF-kappaB is a target of AKT in anti-apoptotic PDGF signalling. Nature 1999, 401, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.; Lee, J.T.; Navolanic, P.M.; Steelman, L.S.; Shelton, J.G.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: A target for cancer chemotherapy. Leukemia 2003, 17, 590–603. [Google Scholar] [CrossRef]
- Chen, C.M.; Kraut, N.; Groudine, M.; Weintraub, H. I-mf, a novel myogenic repressor, interacts with members of the MyoD family. Cell 1996, 86, 731–741. [Google Scholar] [CrossRef]
- Chou, C.; Chen, R.; Chou, F.; Chang, H.; Chen, Y.; Lee, Y.; Yang, K.D.; Cheng, J.; Huang, C.; Liu, R. miR-146b is Highly Expressed in Adult Papillary Thyroid Carcinomas with High Risk Features Including Extrathyroidal Invasion and the BRAFV600E Mutation. Thyroid 2010, 20, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zhou, P.; von Gise, A.; Gu, F.; Ma, Q.; Chen, J.; Guo, H.; van Gorp, P.R.; Wang, D.Z.; Pu, W.T. Pi3kcb links Hippo-YAP and PI3K-AKT signaling pathways to promote cardiomyocyte proliferation and survival. Circ. Res. 2015, 116, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Carpten, J.D.; Faber, A.L.; Horn, C.; Donoho, G.P.; Briggs, S.L.; Robbins, C.M.; Hostetter, G.; Boguslawski, S.; Moses, T.Y.; Savage, S.; et al. A transforming mutation in the pleckstrin homology domain of AKT1 in cancer. Nature 2007, 448, 439–444. [Google Scholar] [CrossRef]
- Vivanco, I.; Sawyers, C.L. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat. Rev. Cancer 2002, 2, 489–501. [Google Scholar] [CrossRef]
- Larson-Casey, J.L.; Deshane, J.S.; Ryan, A.J.; Thannickal, V.J.; Carter, A.B. Macrophage Akt1 Kinase-Mediated Mitophagy Modulates Apoptosis Resistance and Pulmonary Fibrosis. Immunity 2016, 44, 582–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsaves, V.; Zhang, R.; Ruder, D.; Pan, Y.; Leventaki, V.; Rassidakis, G.Z.; Claret, F.X. Constitutive control of AKT1 gene expression by JUNB/CJUN in ALK+ anaplastic large-cell lymphoma: A novel crosstalk mechanism. Leukemia 2015, 29, 2162–2172. [Google Scholar] [CrossRef] [PubMed]
- Leszczynska, K.B.; Foskolou, I.P.; Abraham, A.G.; Anbalagan, S.; Tellier, C.; Haider, S.; Span, P.N.; O’Neill, E.E.; Buffa, F.M.; Hammond, E.M. Hypoxia-induced p53 modulates both apoptosis and radiosensitivity via AKT. J. Clin. Investig. 2015, 125, 2385–2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, T.P.; Suman, S.; Alatassi, H.; Ankem, M.K.; Damodaran, C. Inhibition of AKT promotes FOXO3a-dependent apoptosis in prostate cancer. Cell Death Dis. 2016, 7, e2111. [Google Scholar] [CrossRef] [PubMed]
- Rana, C.; Piplani, H.; Vaish, V.; Nehru, B.; Sanyal, S.N. Downregulation of PI3-K/Akt/PTEN pathway and activation of mitochondrial intrinsic apoptosis by Diclofenac and Curcumin in colon cancer. Mol. Cell. Biochem. 2015, 402, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Berkes, C.A.; Tapscott, S.J. MyoD and the transcriptional control of myogenesis. Semin. Cell Dev. Biol. 2005, 16, 585–595. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequences (5′–3′) | Size (bp) | Annealing Temperature (°C) | Accession Number |
|---|---|---|---|---|
| AKT1 | F: CACACGCTGACAGAAAACCG | 128 | 60 | NM_205055.1 |
| R: AACAACTCCCCTCCGTTAGC | ||||
| MDFIC | F: CAATGGCAGCAAGAAGA | 126 | 52 | XM_416018.5 |
| R: AGAACAATGTTACAGAGGGT | ||||
| Cyclin B2 | F: CAGTAAAGGCTACGAAAG | 133 | 58 | NM_001004369.1 |
| R: ACATCCATAGGGACAGG | ||||
| Cyclin D1 | F: CAGAAGTGCGAAGAGGAAGT | 188 | 58 | NM_205381.1 |
| R: CTGATGGAGTTGTCGGTGTA | ||||
| Cyclin D2 | F: AACTTGCTCTACGACGACC | 150 | 58 | NM_204213 |
| R: TTCACAGACCTCCAACATC | ||||
| PCNA | F: GTGCTGGGACCTGGGTT | 217 | 58 | NM_204170.2 |
| R: CGTATCCGCATTGTCTTCT | ||||
| p21 | F: GAAGAGTTGTCCACGATAAGC | 247 | 58 | NM_204170.2 |
| R: TTCCAGTCCTCCTCAGTCC | ||||
| CDKN1B | F: GCTGTGCTGGGCTGAA | 207 | 58 | NM_204256.2 |
| R: GGACGAAAGGATGTGGG | ||||
| Caspase-3 | F: TGGCCCTCTTGAACTGAAAG | 106 | 61 | NM_204725.1 |
| R: TCCACTGTCTGCTTCAATACC | ||||
| Caspase-8 | F: CCCTGAAGACAGTGCCATTT | 207 | 61 | NM_204592.2 |
| R: GGGTCGGCTGGTCATTTTAT | ||||
| Caspase-9 | F: TCCCGGGCTGTTTCAACTT | 270 | 61 | XM_424580.5 |
| R: CCTCATCTTGCAGCTTGTGC | ||||
| Fas | F: TCCACCTGCTCCTCGTCATT | 78 | 61 | NM_001199487.1 |
| R: GTGCAGTGTGTGTGGGAACT | ||||
| Cyt c | F: TGTCCAGAAATGTTCCCAGTGC | 138 | 61 | NM_001079478.1 |
| R: CCTTTGTTCTTATTGGCATCTGTG | ||||
| MYOD | F: GCTACTACACGGAATCACCAAAT | 200 | 58 | NM_204214.2 |
| R: CTGGGCTCCACTGTCACTCA | ||||
| MYOG | F: CGGAGGCTGAAGAAGGTGAA | 320 | 58 | NM_204184.1 |
| R: CGGTCCTCTGCCTGGTCAT | ||||
| MYHC | F: CTCCTCACGCTTTGGTAA | 213 | 58 | NM_001319304.1 |
| R: TGATAGTCGTATGGGTTGGT | ||||
| GAPDH | F: TCCTCCACCTTTGATGCG | 146 | 50–62 | NM_204305.1 |
| R: GTGCCTGGCTCACTCCTT |
| Sequence Name | Sequences (5′–3′) |
|---|---|
| gga-miR-146b-3p mimic | GGGAUACCUAAGUCAAGACG |
| gga-miR-146b-3p inhibitor | CGUCUUGACUUAGGUAUCCC |
| si-AKT1 | GCTGAAGAAATGGAAGTTT |
| si-MDFIC | ATGGAAGTGGAATGCACAA |
| Primer Name | Primer Sequences (5′–3′) | Size (bp) | Annealing Temperature (°C) |
|---|---|---|---|
| pcDNA3.1-AKT1 | F: CCCAAGCTTGAGACATTCCCGCCATTA | 1612 | 61 |
| R: CCGCTCGAGAGAATCTGTGAGGCTTCCTA | |||
| pcDNA3.1-MDFIC | F: CCCAAGCTTGTTAGTTCGGCTGGAGGGAG | 1064 | 60 |
| R: CCGCTCGAGCAGCAGGCCGATAGTTCAGT | |||
| pmirGLO-AKT1-3′UTR-WT | F: CCCAAGCTTAAAAAAATTAAAAAAGCC | 100 | 55 |
| R: CCGCTCGAGCAAAAGGAAGGAGGAACC | |||
| pmirGLO-AKT1-3′UTR-MT | F: AGAACTGAACATTCCCTAATAAGCGGGAGCAAA | 2771 | 68 |
| R: TCTTGACTTGTAAGGGATTATTCGCCCTCGTTT | |||
| pmirGLO-MDFIC-3′UTR-WT | F: CCCAAGCTTGGACTTTCTTTCTGTATTTAT | 100 | 52 |
| R: TTGCGGCCGCAATTCTGCTTGTTTTAATTACA | |||
| pmirGLO-MDFIC-3′UTR-MT | F: GTAATTTTCACTCCATAGGCAATCTTATAAGC | 2771 | 68 |
| R: GCTTATAAGATTGAACGCTTAGTGAAAATTAC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.; Guo, L.; Zhao, M.; Zhang, D.; Xu, H.; Nie, Q. The Inhibition on MDFIC and PI3K/AKT Pathway Caused by miR-146b-3p Triggers Suppression of Myoblast Proliferation and Differentiation and Promotion of Apoptosis. Cells 2019, 8, 656. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8070656
Huang W, Guo L, Zhao M, Zhang D, Xu H, Nie Q. The Inhibition on MDFIC and PI3K/AKT Pathway Caused by miR-146b-3p Triggers Suppression of Myoblast Proliferation and Differentiation and Promotion of Apoptosis. Cells. 2019; 8(7):656. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8070656
Chicago/Turabian StyleHuang, Weiling, Lijin Guo, Minxing Zhao, Dexiang Zhang, Haiping Xu, and Qinghua Nie. 2019. "The Inhibition on MDFIC and PI3K/AKT Pathway Caused by miR-146b-3p Triggers Suppression of Myoblast Proliferation and Differentiation and Promotion of Apoptosis" Cells 8, no. 7: 656. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8070656