Cdc42 Couples T Cell Receptor Endocytosis to GRAF1-Mediated Tubular Invaginations of the Plasma Membrane


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression Constructs
2.2. Cell Culture and Sample Preparation
2.3. Flow Cytometry
2.4. Microscopy
2.5. Image Analysis
2.6. Data Analysis
3. Results and Discussion
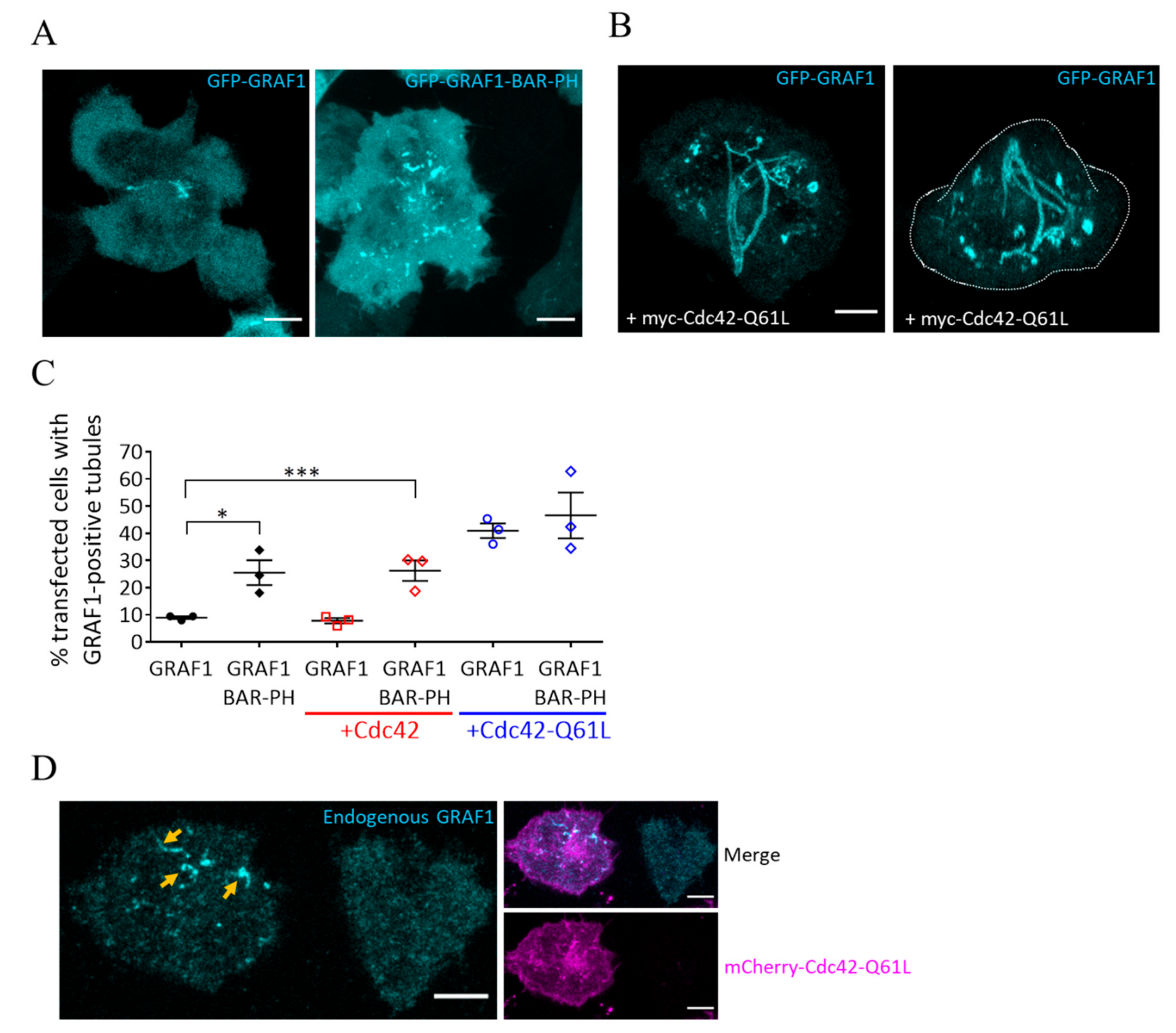
3.1. Cdc42 and GRAF1 Regulate the Formation of Tubular Structures in Activated T Cells
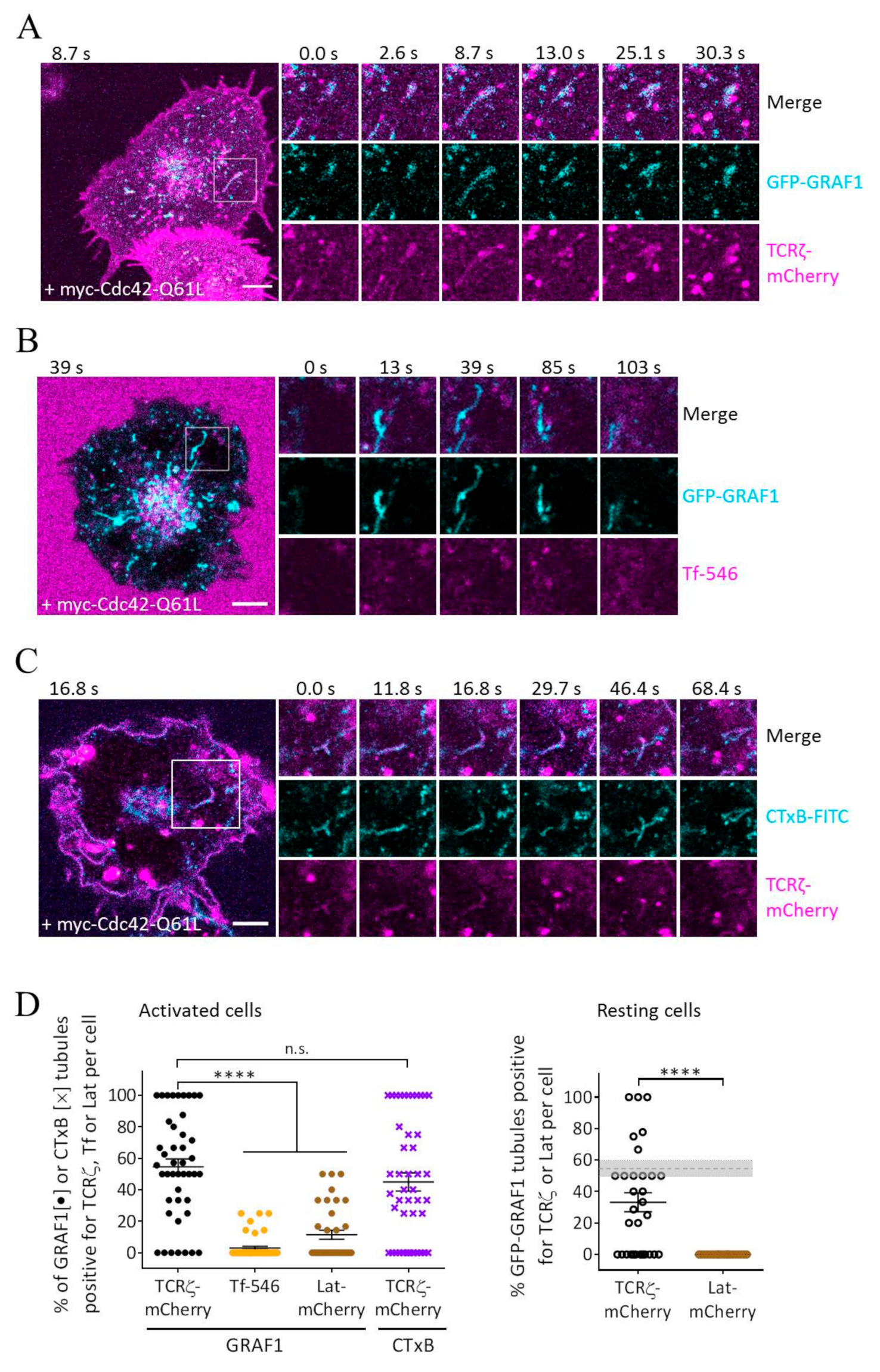
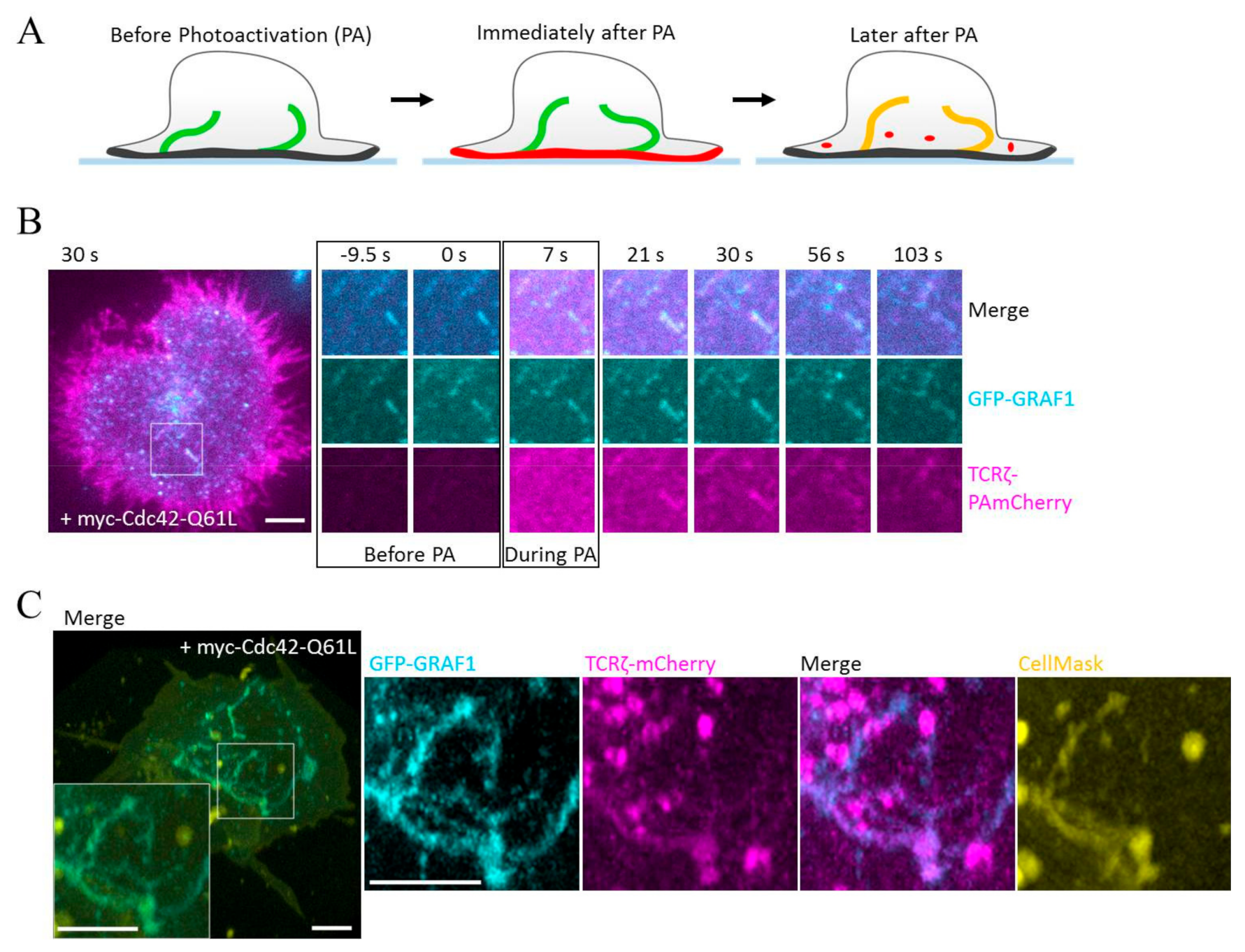
3.2. GRAF1-Positive Tubules Contain Internalized TCRζ but Not Transferrin
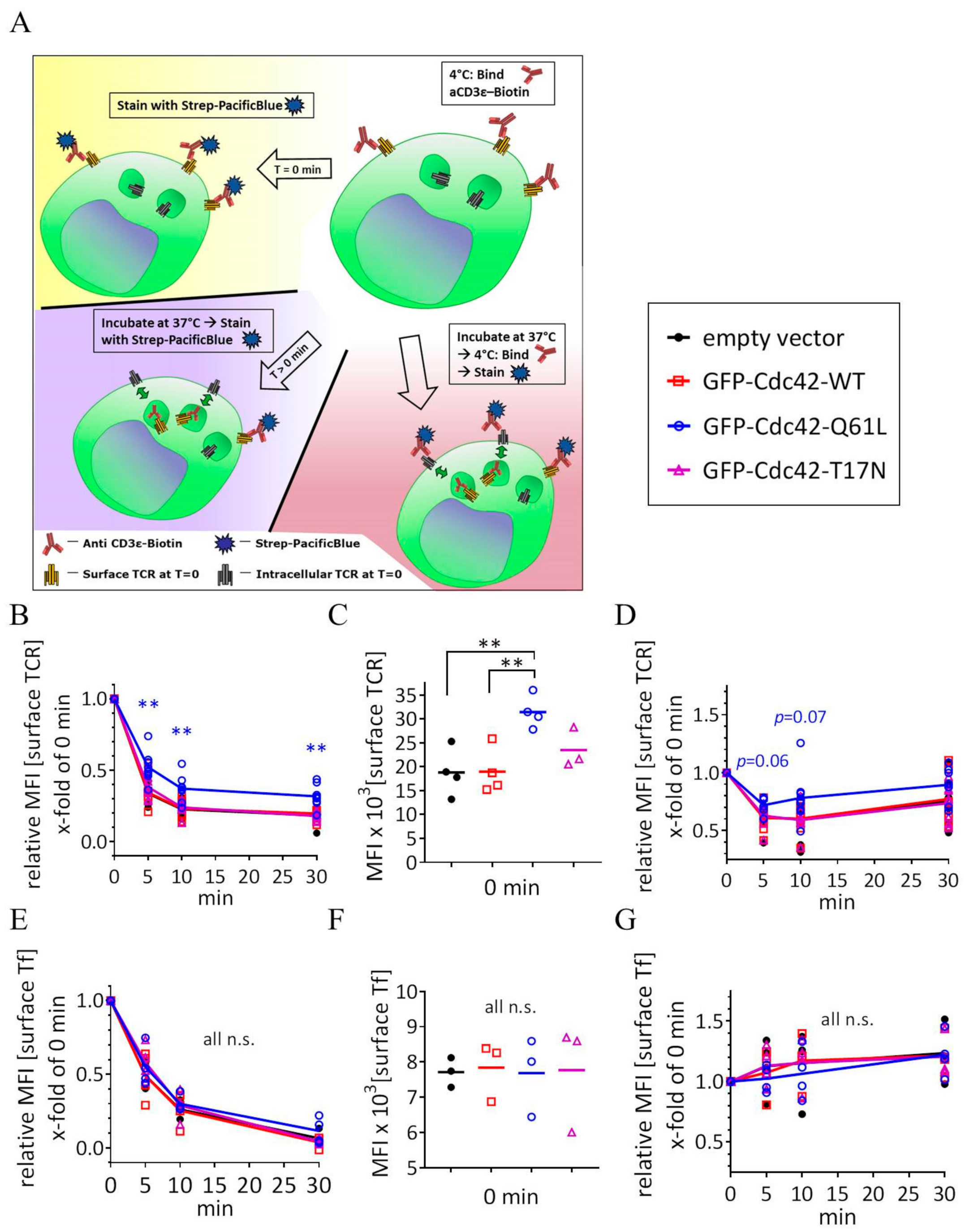
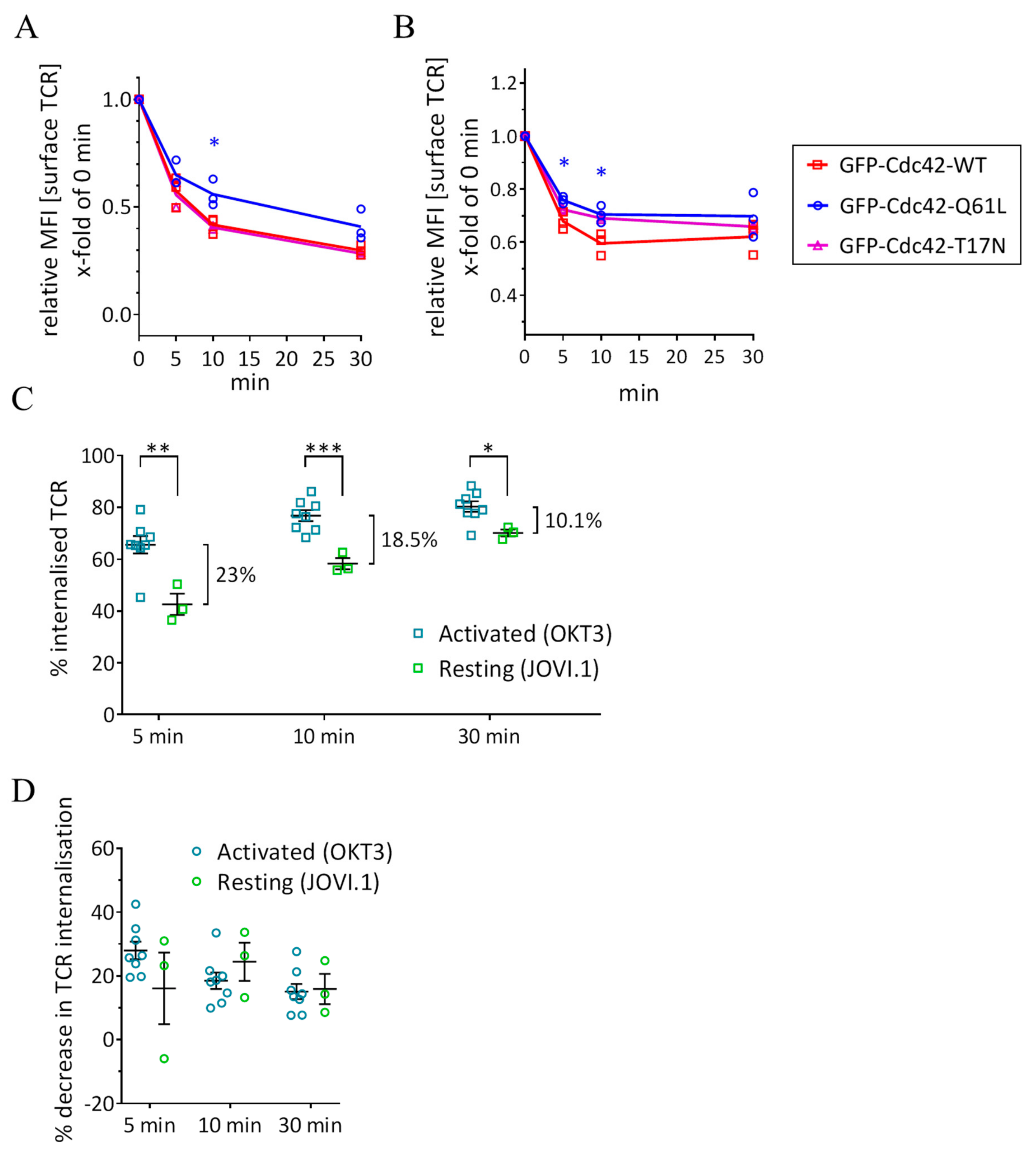
3.3. Expression of Cdc42-Q61L Selectively Impairs Internalization of the TCR-CD3 Complex
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Onnis, A.; Baldari, C.T. Orchestration of Immunological Synapse Assembly by Vesicular Trafficking. Front. Cell Dev. Biol. 2019, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Essen, M.; Bonefeld, C.M.; Siersma, V.; Rasmussen, A.B.; Lauritsen, J.P.H.; Nielsen, B.L.; Geisler, C. Constitutive and ligand-induced TCR degradation. J. Immunol. 2004, 173, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Valitutti, S.; Müller, S.; Salio, M.; Lanzavecchia, A. Degradation of T Cell Receptor (TCR)–CD3-ζ Complexes after Antigenic Stimulation. J. Exp. Med. 1997, 185, 1859–1864. [Google Scholar] [CrossRef] [PubMed]
- Mariathasan, S.; Bachmann, M.F.; Bouchard, D.; Ohteki, T.; Ohashi, P.S. Degree of TCR internalization and Ca2+ flux correlates with thymocyte selection. J. Immunol. 1998, 161, 6030–6037. [Google Scholar]
- André, P.; Boretto, J.; Hueber, A.O.; Régnier-Vigouroux, A.; Gorvel, J.P.; Ferrier, P.; Chavrier, P. A dominant-negative mutant of the Rab5 GTPase enhances T cell signaling by interfering with TCR down-modulation in transgenic mice. J. Immunol. 1997, 159, 5253–5263. [Google Scholar]
- Liu, H.; Rhodes, M.; Wiest, D.L.; Vignali, D.A. On the Dynamics of TCR:CD3 Complex Cell Surface Expression and Downmodulation. Immunity 2000, 13, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Soares, H.; Henriques, R.; Sachse, M.; Ventimiglia, L.; Alonso, M.A.; Zimmer, C.; Thoulouze, M.-I.; Alcover, A. Regulated vesicle fusion generates signaling nanoterritories that control T cell activation at the immunological synapse. J. Exp. Med. 2013, 210, 2415–2433. [Google Scholar] [CrossRef] [Green Version]
- Finetti, F.; Paccani, S.R.; Riparbelli, M.G.; Giacomello, E.; Perinetti, G.; Pazour, G.J.; Rosenbaum, J.L.; Baldari, C.T. Intraflagellar transport is required for polarized recycling of the TCR/CD3 complex to the immune synapse. Nat. Cell Biol. 2009, 11, 1332–1339. [Google Scholar] [CrossRef] [Green Version]
- Osborne, D.G.; Piotrowski, J.T.; Dick, C.J.; Zhang, J.-S.; Billadeau, D.D. SNX17 Affects T Cell Activation by Regulating TCR and Integrin Recycling. J. Immunol. 2015, 194, 4555–4566. [Google Scholar] [CrossRef]
- Compeer, E.B.; Kraus, F.; Ecker, M.; Redpath, G.; Amiezer, M.; Rother, N.; Nicovich, P.R.; Kapoor-Kaushik, N.; Deng, Q.; Samson, G.P.B.; et al. A mobile endocytic network connects clathrin-independent receptor endocytosis to recycling and promotes T cell activation. Nat. Commun. 2018, 9, 1597. [Google Scholar] [CrossRef]
- Fernandez-Arenas, E.; Calleja, E.; Martinez-Martin, N.; Gharbi, S.I.; Navajas, R.; Garcia-Medel, N.; Penela, P.; Alcami, A.; Mayor, F.; Albar, J.P.; et al. β-arrestin-1 mediates the TCR-triggered re-routing of distal receptors to the immunological synapse by a PKC-mediated mechanism. EMBO J. 2014, 33, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Monjas, A.; Alcover, A.; Alarcón, B. Engaged and Bystander T Cell Receptors Are Down-modulated by Different Endocytotic Pathways. J. Biol. Chem. 2004, 279, 55376–55384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, J.; Kastrup, J.; Nielsen, B.L.; Odum, N.; Geisler, C. Regulation and function of the CD3gamma DxxxLL motif: A binding site for adaptor protein-1 and adaptor protein-2 in vitro. J. Cell Biol. 1997, 138, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Barr, V.A.; Balagopalan, L.; Barda-Saad, M.; Polishchuk, R.; Boukari, H.; Bunnell, S.C.; Bernot, K.M.; Toda, Y.; Nossal, R.; Samelson, L.E. T-cell antigen receptor-induced signaling complexes: Internalization via a cholesterol-dependent endocytic pathway. Traffic 2006, 7, 1143–1162. [Google Scholar] [CrossRef]
- Martínez-Martín, N.; Fernández-Arenas, E.; Cemerski, S.; Delgado, P.; Turner, M.; Heuser, J.; Irvine, D.J.; Huang, B.; Bustelo, X.R.; Shaw, A.; et al. T cell receptor internalization from the immunological synapse is mediated by TC21 and RachoG GTPase-dependent phagocytosis. Immunity 2011, 35, 208–222. [Google Scholar] [CrossRef]
- Johannes, L.; Parton, R.G.; Bassereau, P.; Mayor, S. Building endocytic pits without clathrin. Nat. Rev. Mol. Cell Biol. 2015, 16, 311–321. [Google Scholar] [CrossRef]
- Sabharanjak, S.; Sharma, P.; Parton, R.G.; Mayor, S. GPI-anchored proteins are delivered to recycling endosomes via a distinct cdc42-regulated, clathrin-independent pinocytic pathway. Dev. Cell 2002, 2, 411–423. [Google Scholar] [CrossRef]
- Chadda, R.; Howes, M.T.; Plowman, S.J.; Hancock, J.F.; Parton, R.G.; Mayor, S. Cholesterol-Sensitive Cdc42 Activation Regulates Actin Polymerization for Endocytosis via the GEEC Pathway. Traffic 2007, 8, 702–717. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, N.C.; Monzo, P.; Kaddai, V.; Doye, A.; Ricci, V.; Boquet, P. Helicobacter pylori VacA Cytotoxin: A Probe for a Clathrin-independent and Cdc42-dependent Pinocytic Pathway Routed to Late Endosomes. Mol. Biol. Cell 2005, 16, 4852–4866. [Google Scholar] [CrossRef]
- Cannon, J.L.; Labno, C.M.; Bosco, G.; Seth, A.; McGavin, M.H.; Siminovitch, K.A.; Rosen, M.K.; Burkhardt, J.K. Wasp recruitment to the T cell:APC contact site occurs independently of Cdc42 activation. Immunity 2001, 15, 249–259. [Google Scholar] [CrossRef]
- Labno, C.M.; Lewis, C.M.; You, D.; Leung, D.W.; Takesono, A.; Kamberos, N.; Seth, A.; Finkelstein, L.D.; Rosen, M.K.; Schwartzberg, P.L.; et al. Itk Functions to Control Actin Polymerization at the Immune Synapse through Localized Activation of Cdc42 and WASP. Curr. Biol. 2003, 13, 1619–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, J.A.; Wang, J.; Saeed, M.; Pedrosa, A. Origin, Organization, Dynamics, and Function of Actin and Actomyosin Networks at the T Cell Immunological Synapse. Annu. Rev. Immunol. 2018, 37, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shehabeldin, A.; da Cruz, L.A.; Butler, J.; Somani, A.K.; McGavin, M.; Kozieradzki, I.; dos Santos, A.O.; Nagy, A.; Grinstein, S.; et al. Antigen receptor-induced activation and cytoskeletal rearrangement are impaired in Wiskott-Aldrich syndrome protein-deficient lymphocytes. J. Exp. Med. 1999, 190, 1329–1342. [Google Scholar] [CrossRef] [PubMed]
- Doherty, G.J.; Lundmark, R. GRAF1-dependent endocytosis. Biochem. Soc. Trans. 2009, 37, 1061–1065. [Google Scholar] [CrossRef]
- Francis, M.K.; Holst, M.R.; Vidal-Quadras, M.; Henriksson, S.; Santarella-Mellwig, R.; Sandblad, L.; Lundmark, R. Endocytic membrane turnover at the leading edge is driven by a transient interaction between Cdc42 and GRAF1. J. Cell Sci. 2015, 128, 4183–4195. [Google Scholar] [CrossRef] [Green Version]
- Lundmark, R.; Doherty, G.J.; Howes, M.T.; Cortese, K.; Vallis, Y.; Parton, R.G.; McMahon, H.T. The GTPase-activating protein GRAF1 regulates the CLIC/GEEC endocytic pathway. Curr. Biol. 2008, 18, 1802–1808. [Google Scholar] [CrossRef]
- Vidal-Quadras, M.; Holst, M.R.; Francis, M.K.; Larsson, E.; Hachimi, M.; Yau, W.-L.; Peränen, J.; Martín-Belmonte, F.; Lundmark, R. Endocytic turnover of Rab8 controls cell polarization. J. Cell Sci. 2017, 130, 1147–1157. [Google Scholar] [CrossRef] [Green Version]
- Bretou, M.; Jouannot, O.; Fanget, I.; Pierobon, P.; Larochette, N.; Gestraud, P.; Guillon, M.; Emiliani, V.; Gasman, S.; Desnos, C.; et al. Cdc42 controls the dilation of the exocytotic fusion pore by regulating membrane tension. Mol. Biol. Cell 2014, 25, 3195–3209. [Google Scholar] [CrossRef]
- Ory, S.; Gasman, S. Rho GTPases and Exocytosis: What Are the Molecular Links? Semin. Cell Dev. Biol. 2011, 22, 27–32. [Google Scholar] [CrossRef]
- Wen, P.J.; Osborne, S.L.; Zanin, M.; Low, P.C.; Wang, H.-T.A.; Schoenwaelder, S.M.; Jackson, S.P.; Wedlich-Söldner, R.; Vanhaesebroeck, B.; Keating, D.J.; et al. Phosphatidylinositol(4,5)bisphosphate coordinates actin-mediated mobilization and translocation of secretory vesicles to the plasma membrane of chromaffin cells. Nat. Commun. 2011, 2, 491. [Google Scholar] [CrossRef] [Green Version]
- Lakshminarayan, R.; Wunder, C.; Becken, U.; Howes, M.T.; Benzing, C.; Arumugam, S.; Sales, S.; Ariotti, N.; Chambon, V.; Lamaze, C.; et al. Galectin-3 drives glycosphingolipid-dependent biogenesis of clathrin-independent carriers. Nat. Cell Biol. 2014, 16, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Day, C.A.; Baetz, N.W.; Copeland, C.A.; Kraft, L.J.; Han, B.; Tiwari, A.; Drake, K.R.; De Luca, H.; Chinnapen, D.J.-F.; Davidson, M.W.; et al. Microtubule motors power plasma membrane tubulation in clathrin-independent endocytosis. Traffic 2015, 16, 572–590. [Google Scholar] [CrossRef] [PubMed]
- Redpath, G.M.I.; Ecker, M.; Kapoor-Kaushik, N.; Vartoukian, H.; Carnell, M.; Kempe, D.; Biro, M.; Ariotti, N.; Rossy, J. Flotillins promote T cell receptor sorting through a fast Rab5–Rab11 endocytic recycling axis. Nat. Commun. 2019, 10, 4392. [Google Scholar] [CrossRef] [PubMed]
- Batista, A.; Millán, J.; Mittelbrunn, M.; Sánchez-Madrid, F.; Alonso, M.A. Recruitment of Transferrin Receptor to Immunological Synapse in Response to TCR Engagement. J. Immunol. 2004, 172, 6709–6714. [Google Scholar] [CrossRef] [Green Version]
- Boucrot, E.; Ferreira, A.P.A.; Almeida-Souza, L.; Debard, S.; Vallis, Y.; Howard, G.; Bertot, L.; Sauvonnet, N.; McMahon, H.T. Endophilin marks and controls a clathrin-independent endocytic pathway. Nature 2015, 517, 460–465. [Google Scholar] [CrossRef]
- Shitara, A.; Malec, L.; Ebrahim, S.; Chen, D.; Bleck, C.; Hoffman, M.P.; Weigert, R. Cdc42 negatively regulates endocytosis during apical membrane maintenance in live animals. Mol. Biol. Cell 2019, 30, 324–332. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossatti, P.; Ziegler, L.; Schregle, R.; Betzler, V.M.; Ecker, M.; Rossy, J. Cdc42 Couples T Cell Receptor Endocytosis to GRAF1-Mediated Tubular Invaginations of the Plasma Membrane. Cells 2019, 8, 1388. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8111388
Rossatti P, Ziegler L, Schregle R, Betzler VM, Ecker M, Rossy J. Cdc42 Couples T Cell Receptor Endocytosis to GRAF1-Mediated Tubular Invaginations of the Plasma Membrane. Cells. 2019; 8(11):1388. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8111388
Chicago/Turabian StyleRossatti, Pascal, Luca Ziegler, Richard Schregle, Verena M. Betzler, Manuela Ecker, and Jérémie Rossy. 2019. "Cdc42 Couples T Cell Receptor Endocytosis to GRAF1-Mediated Tubular Invaginations of the Plasma Membrane" Cells 8, no. 11: 1388. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8111388