
On the Calculation of System Entropy in Nonlinear Stochastic Biological Networks

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. On the Measurement of System Entropy in Linear Stochastic Biological Networks

- (i)
- The LMI in Equation (4) is equivalent to the following Riccati-like equation through the Schur complement [15]:
- (ii)
- Obtaining the system randomness r by solving the LMI-constrained optimization problem in Equation (5), the Riccati-like equation in Equation (6) could be replaced by:From the Riccati-like inequality in Equation (7), it is clear that the system randomness r and also the system entropy s are all dependent on the system parameters A, B and C. Obviously, the system randomness r or entropy s is a measurement of systematic characteristic. Since the last term in the right-hand side of the inequality Equation (7) is positive, it is seen that if the eigenvalues of A are all located at the farther left-hand side of s-domain (i.e., the real parts of eigenvalues are more negative or the loops of biological system are with more strength from the systematic perspective) (Figure 2), the biological network in Equation (1) is with less system randomness r and lower system entropy s. If the eigenvalues of A are all located near the imaginary axis (i.e., less negative), in order to maintain the inequality in Equation (7), the system randomness r or system entropy s must be large enough. In summary, biological systems with more stability are with more ability to maintain its system structure (or phenotype) and therefore is with less system randomness or entropy, and vice versa.
- (iii)
- From the Riccati-like inequality in Equaiton (7), if system matrix A is fixed, in order to make system randomness r and system entropy s smaller, input coupling matrix B and output coupling matrix C in Equation (1) must be smaller. This is why there are so many membranes and semi-transparent membranes isolating biological systems from the environment with only some receptors or sensors remained to interact with the outside environment (i.e., make B and C in Equation (1) as small as possible to protect the biological system from the environment).

- (i)
- If the eigenvalues of A in Equation (11) are more negative (or the system loops are with more strong strength), then the term −(AP + ATP) in Equation (14) becomes larger due to P > 0 and the biological system could tolerate more the term due to intrinsic fluctuation in Equation (11) and could decrease the system randomness r (or system entropy s).
- (ii)
- In order to attenuate the effect of intrinsic parameter fluctuations on the increase of system randomness or system entropy (i.e., let in Equation (14) as small as possible), there exist so many redundant and module structures in biological systems to attenuate the phenotypic variations in Ai due to genetic variations and epigenetic alterations in the evolutionary process.
- (iii)
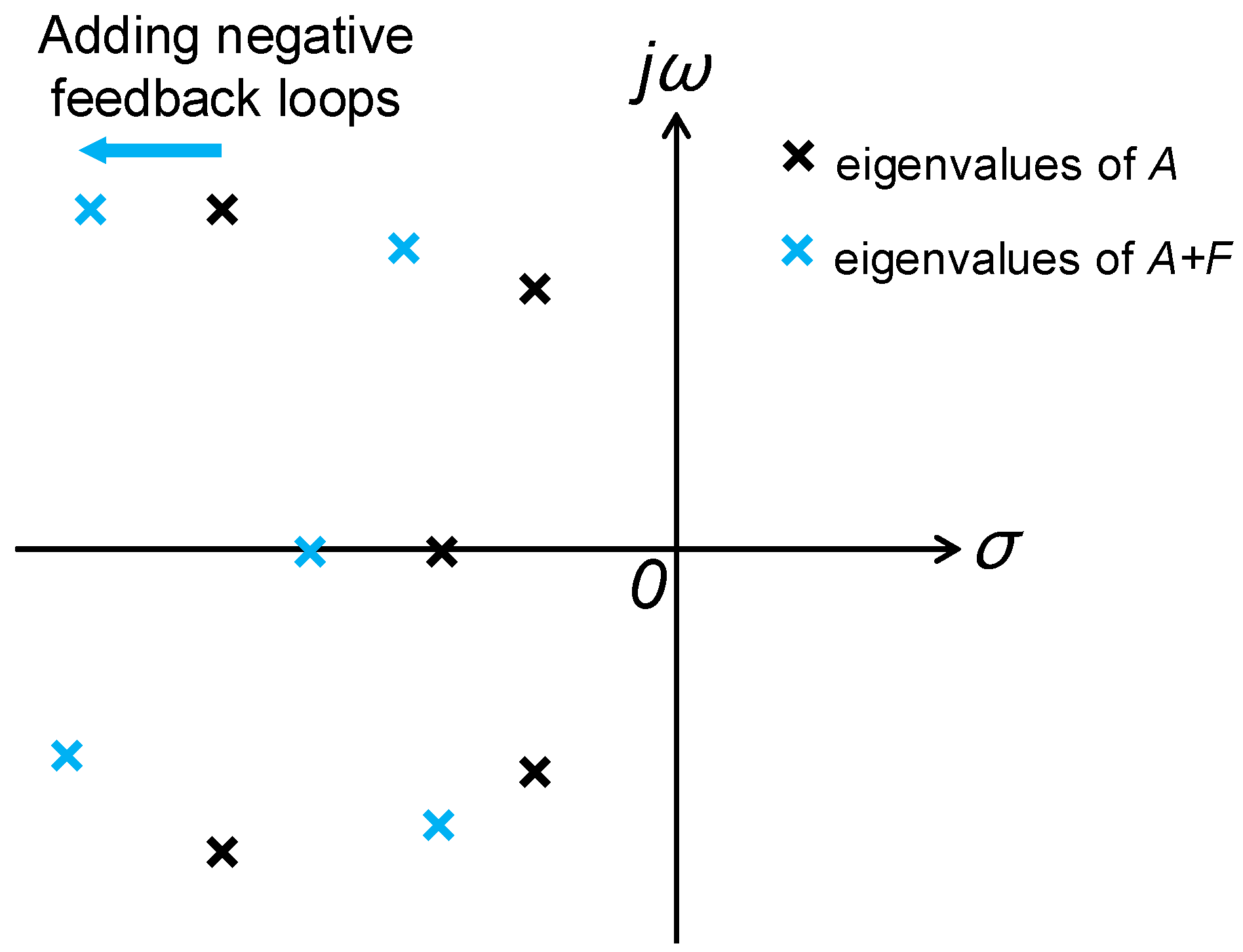
- If the state feedback loops FX(t) with all eigenvalues in the left complex plane of the s-domain are developed for the stochastic biological network Equation (9) in the evolutionary process as follows:In this situation, the Riccati-like inequality in Equation (14) should be changed as:Therefore, the biological network with feedback loops F to let all eigenvalues of A + F in the farther left complex plane of the s-domain will be with more stability robustness to tolerate more network random fluctuations and will decrease the network randomness and entropy to maintain its phenotype, i.e., a larger stability robustness in the right-hand side of Equation (16) will lead to less system randomness r or system entropy s tolerating larger random fluctuation term due to in Equation (15). However, if the added feedback loops F make the eigenvalues of A + F closer to the jω-axis than the eigenvalues of A, the feedback loops will increase the network randomness and entropy of the biological network.
- (iv)
- Dissipativity, all biological networks are open irreversible systems and the entropy production rate of biological system in Equation (16) is associated with the following dissipation rate η [26]:By the following derivative procedure with V(X(t)) = XT(t)PX(t) ≥ 0 and P > 0:
2.2. On the Measurement of System Entropy in Nonlinear Stochastic Biological Network
- (i)
- Comparing the HJI in Equation (18) with the HJI in Equation (22) when is replaced by r, it is seen that the positive term in Equation (22) due to the random intrinsic fluctuations will cause a larger system randomness r and system entropy s for the nonlinear stochastic network in Equation (20).
- (ii)
- If some nonlinear feedback loops F(X) are developed for the nonlinear stochastic biological network Equation (20) in the evolutionary process as follows:then the HJI in Equation (22) is modified as the following HJI:
2.3. The Calculation of System Entropy in Nonlinear Stochastic Biological Networks by the Global Linearization Method
- (i)
- From Equation (30), it is seen that if the eigenvalues of local linearized system matrices Ai are all with more negative real part (more stable), then the nonlinear biological network in Equations (17) or (26) will be with less system randomness r and less system entropy s, and vice versa.
- (ii)
- The LMIs-constrained optimization problem for system randomness r in Equation (29) can be easily solved by decreasing until no positive definite solution P > 0 exists in Equations (27) or (28) with the help of the LMI toolbox included in the Matlab software package.
- (i)
- Comparing Equation (30) with Equation (37), because of the positive term due to intrinsic random fluctuations in the nonlinear stochastic biological network in Equation (20), system randomness r in Equation (37) must be larger than system randomness r in Equation (30) of the nonlinear biological network Equation (17) without intrinsic random fluctuations. Obviously, the intrinsic random fluctuations can increase system randomness r and system entropy s of the nonlinear biological networks.
- (ii)
- If the nonlinear stochastic biological network in Equation (20) has developed new feedback loops F(X) to tolerate network fluctuations as follows:which could be approximated by the following global linearization system:where F(X) is approximated by , then the Riccati-like inequality Equation (37) is replaced by Equation (39):
- (i)
- Construct nonlinear dynamic equations of biological networks as Equations (17) or (20).
- (ii)
- Use global linearization technique in Equations (24) or (31) to approximate the nonlinear biological network as Equations (26) or (33).
- (iii)
- Solve the LMIs-constrained optimization in Equations (29), (36) or (40) for the system randomness r and the entropy of the nonlinear biological network in Equations (17) or (20).
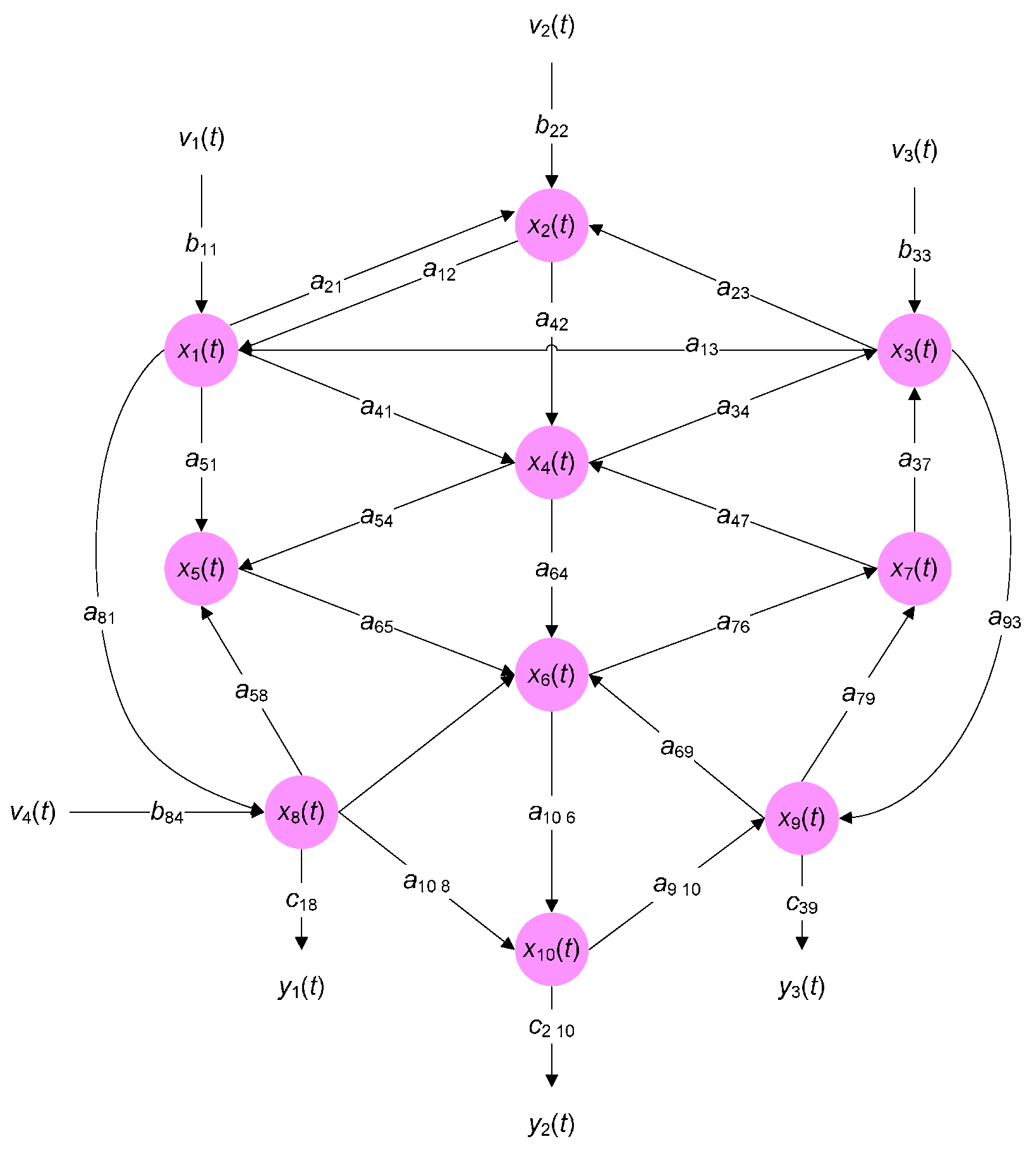
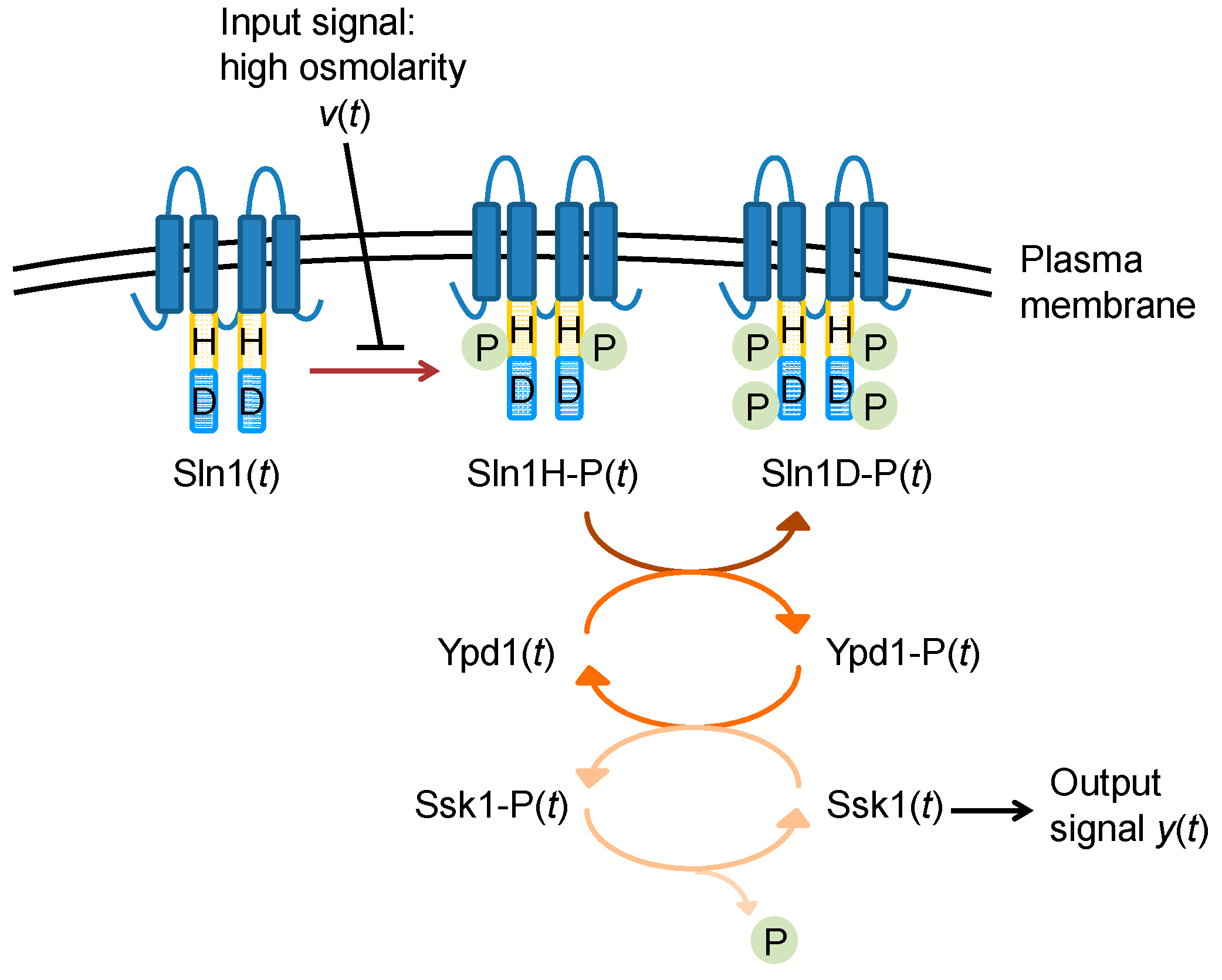
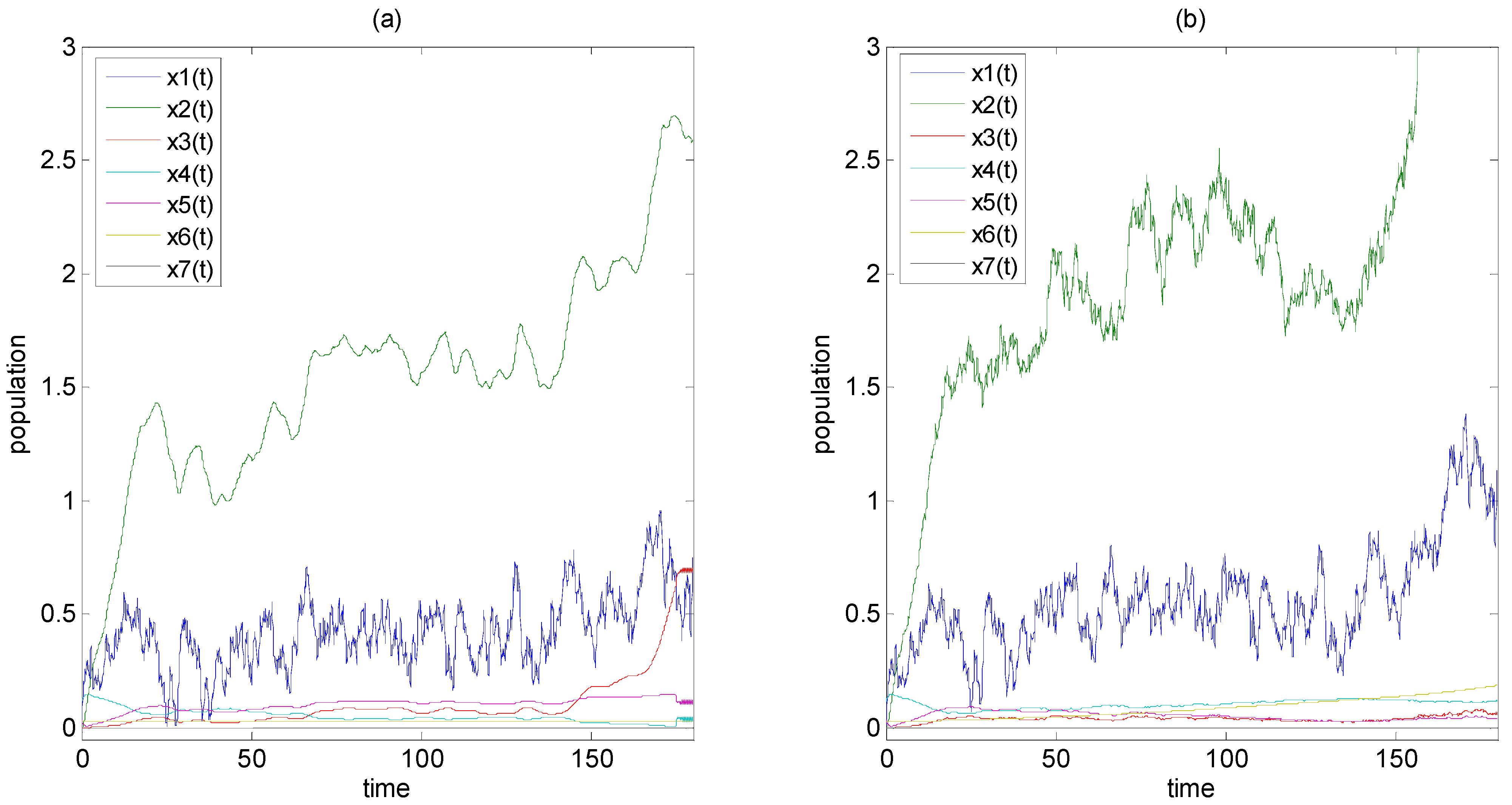
3. Example of Calculating System Entropy of Biological Networks
3.1. Example 1: The System Entropy of a Phosphorelay System


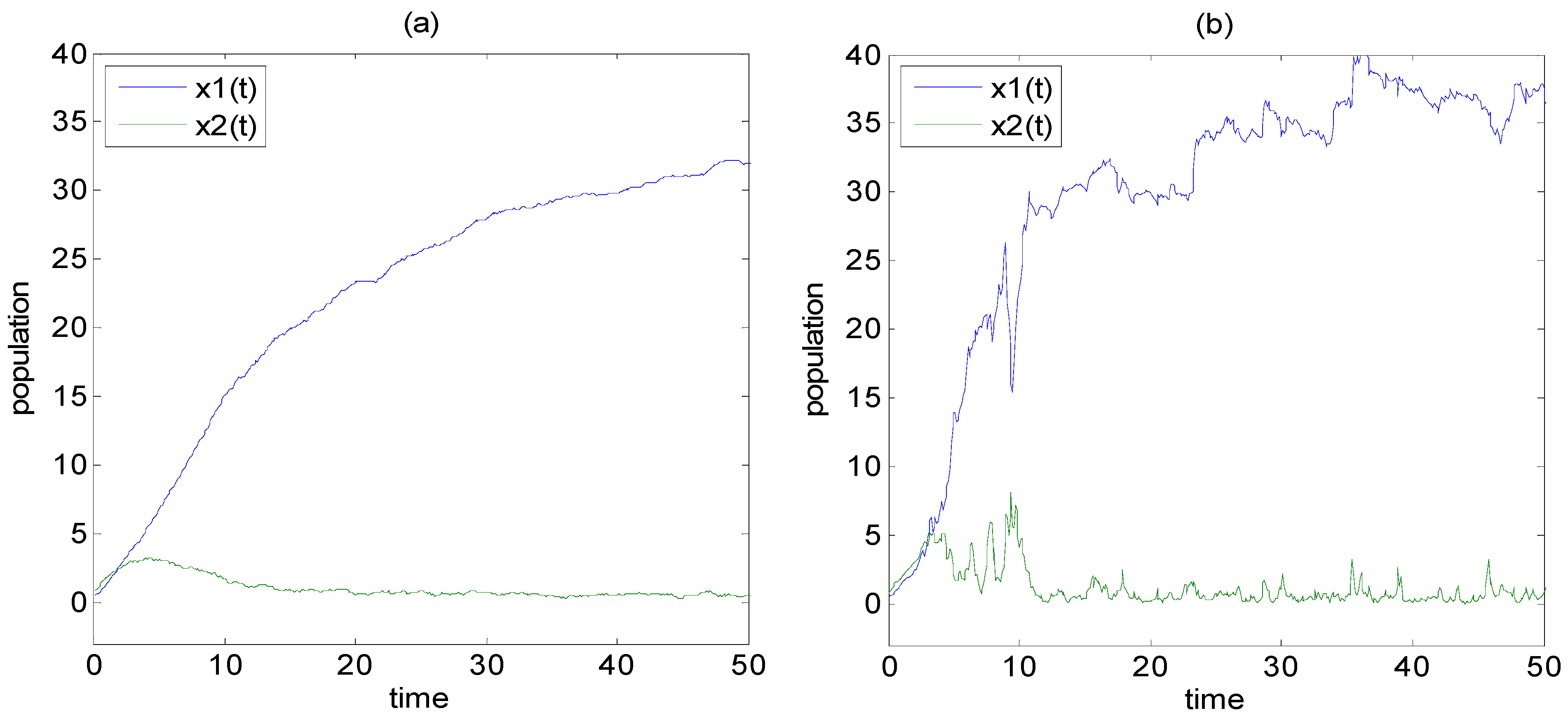
3.2. Example 2: The System Entropy of Predator-Prey Ecological System

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix
Appendix A: Proof of Proposition 1
Appendix B: Proof of Proposition 2
Appendix C: Proof of Proposition 3
Appendix D: Proof of Proposition 4
Appendix E: Proof of Proposition 5
Appendix F: Proof of Proposition 6
Appendix G:
Appendix H:
References
- Mettetal, J.T.; van Oudenaarden, A. Microbiology. Necessary noise. Science 2007, 317, 463–464. [Google Scholar] [CrossRef] [PubMed]
- Pedraza, J.M.; van Oudenaarden, A. Noise propagation in gene networks. Science 2005, 307, 1965–1969. [Google Scholar] [CrossRef] [PubMed]
- Mettetal, J.T.; Muzzey, D.; Pedraza, J.M.; Ozbudak, E.M.; van Oudenaarden, A. Predicting stochastic gene expression dynamics in single cells. Proc. Natl. Acad. Sci. USA 2006, 103, 7304–7309. [Google Scholar] [CrossRef] [PubMed]
- Krawitz, P.; Shmulevich, I. Basin Entropy in Boolean Network Ensembles. Phys. Rev. Lett. 2007, 98, 158701. [Google Scholar] [CrossRef] [PubMed]
- Krawitz, P.; Shmulevich, I. Entropy of complex relevant components of Boolean networks. Phys. Rev. E 2007, 76, 036115. [Google Scholar] [CrossRef]
- Johansson, R. System Modeling and Identification; Springer: London, UK, 1993. [Google Scholar]
- Demetrius, L. Thermodynamics and evolution. J. Theor. Biol. 2000, 206, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-S.; Li, C.-W. On the Interplay between Entropy and Robustness of Gene Regulatory Networks. Entropy 2010, 12, 1071–1101. [Google Scholar] [CrossRef]
- Schrödinger, E. What is Life? 1944. Available online: http://159.226.251.229/videoplayer/What-is-Life.pdf?ich_u_r_i=79fe139669467dd24cdc11542b4e002f&ich_s_t_a_r_t=0&ich_e_n_d=0&ich_k_e_y=1545108906750663172441&ich_t_y_p_e=1&ich_d_i_s_k_i_d=3&ich_u_n_i_t=1 (accessed on 6 October 2015).
- Lucia, U. Irreversible entropy variation and the problem of the trend to equilibrium. Phys. A 2007, 376, 289–292. [Google Scholar] [CrossRef]
- Lucia, U. Irreversibility, entropy and incomplete information. Phys. A 2009, 388, 4025–4033. [Google Scholar] [CrossRef]
- Lucia, U. Maximum entropy generation and kappa-exponential model. Phys. A 2010, 389, 4558–4563. [Google Scholar] [CrossRef]
- Lucia, U. The Gouy-Stodola Theorem in Bioenergetic Analysis of Living Systems (Irreversibility in Bioenergetics of Living Systems). Energies 2014, 7, 5717–5739. [Google Scholar] [CrossRef]
- Lucia, U.; Ponzetto, A.; Deisboeck, T.S. A thermodynamic approach to the ‘mitosis/apoptosis’ ratio in cancer. Phys. A 2015, 436, 246–255. [Google Scholar] [CrossRef]
- Boyd, S.P.; Barratt, C.H.; Boyd, S.P.; Boyd, S.P. Linear Controller Design: Limits of Performance; Prentice-Hall: Upper Saddle River, NJ, USA, 1991. [Google Scholar]
- Chen, B.-S.; Chang, Y.-T.; Wang, Y.-C. Robust H∞ stabilization design in gene networks under stochastic molecular noises: Fuzzy-interpolation approach. IEEE Trans. Syst. Man Cybern B Cybern. 2008, 38, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Díaz, J.; Alvarez-Buylla, E.R. Information flow during gene activation by signaling molecules: Ethylene transduction in Arabidopsis cells as a study system. BMC Syst. Biol. 2009, 3. [Google Scholar] [CrossRef] [PubMed]
- Lezon, T.R.; Banavar, J.R.; Cieplak, M.; Maritan, A.; Fedoroff, N.V. Using the principle of entropy maximization to infer genetic interaction networks from gene expression patterns. Proc. Natl. Acad. Sci. USA 2006, 103, 19033–19038. [Google Scholar] [CrossRef] [PubMed]
- Manke, T.; Demetrius, L.; Vingron, M. An entropic characterization of protein interaction networks and cellular robustness. J. R. Soc. Interface 2006, 3, 843–850. [Google Scholar] [CrossRef]
- McAdams, H.H.; Arkin, A. It’s a noisy business! Genetic regulation at the nanomolar scale. Trends Genet. 1999, 15, 65–69. [Google Scholar] [CrossRef]
- Nagarajan, R.; Aubin, J.E.; Peterson, C.A. Robust dependencies and structures in stem cell differentiation. Int. J. Bifurc. Chaos 2005, 15, 1503–1514. [Google Scholar] [CrossRef]
- Stoll, G.; Rougemont, J.; Naef, F. Representing perturbed dynamics in biological network models. Phys. Rev. E Stat. Nonlin. Soft Matter. Phys. 2007, 76, 011917. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-L.; Yuan, Z.-F.; Guo, M.-C.; Song, S.-D.; Zhang, Q.-Q.; Bao, Z. Maximum entropy principle and population genetic equilibrium. Acta. Genet. Sin. 2002, 29, 562–564. [Google Scholar] [PubMed]
- Wlaschin, A.P.; Trinh, C.T.; Carlson, R.; Srienc, F. The fractional contributions of elementary modes to the metabolism of Escherichia coli and their estimation from reaction entropies. Metab. Eng. 2006, 8, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, N.; Mackey, M.C. Feedback regulation in the lactose operon: A mathematical modeling study and comparison with experimental data. Biophys. J. 2003, 84, 2841–2851. [Google Scholar] [CrossRef]
- Boyd, S.P.; El Ghaoui, L.; Feron, E.; Balakrishnan, V. Linear Matrix Inequalities in System and Control Theory; SIAM: Philadelphia, PA, USA, 1994. [Google Scholar]
- Kitano, H. Biological robustness. Nat. Rev. Genet. 2004, 5, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, L.; Wang, E. Potential landscape and flux framework of nonequilibrium networks: Robustness, dissipation, and coherence of biochemical oscillations. Proc. Natl. Acad. Sci. USA 2008, 105, 12271–12276. [Google Scholar] [CrossRef] [PubMed]
- Krantz, M.; Ahmadpour, D.; Ottosson, L.G.; Warringer, J.; Waltermann, C.; Nordlander, B.; Klipp, E.; Blomberg, A.; Hohmann, S.; Kitano, H. Robustness and fragility in the yeast high osmolarity glycerol (HOG) signal-transduction pathway. Mol. Syst. Biol. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Lenz, P.; Swain, P.S. An entropic mechanism to generate highly cooperative and specific binding from protein phosphorylations. Curr. Biol. 2006, 16, 2150–2155. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-S.; Chang, Y.-T. A systematic molecular circuit design method for gene networks under biochemical time delays and molecular noises. BMC Syst. Biol. 2008, 2. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.S.; Chen, P.W. Robust Engineered Circuit Design Principles for Stochastic Biochemical Networks With Parameter Uncertainties and Disturbances. IEEE Trans. Biomed. Circuits Syst. 2008, 2, 114–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-S.; Chen, P.-W. On the estimation of robustness and filtering ability of dynamic biochemical networks under process delays, internal parametric perturbations and external disturbances. Math. Biosci. 2009, 222, 92–108. [Google Scholar] [CrossRef] [PubMed]
- Hasty, J.; McMillen, D.; Collins, J.J. Engineered gene circuits. Nature 2002, 420, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Lapidus, S.; Han, B.; Wang, J. Intrinsic noise, dissipation cost, and robustness of cellular networks: The underlying energy landscape of MAPK signal transduction. Proc. Natl. Acad. Sci. USA 2008, 105, 6039–6044. [Google Scholar] [CrossRef] [PubMed]
- Batt, G.; Yordanov, B.; Weiss, R.; Belta, C. Robustness analysis and tuning of synthetic gene networks. Bioinformatics 2007, 23, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- Kærn, M.; Elston, T.C.; Blake, W.J.; Collins, J.J. Stochasticity in gene expression: From theories to phenotypes. Nat. Rev. Genet. 2005, 6, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Voit, E.O. Computational Analysis of Biochemical Systems: A Practical Guide for Biochemists and Molecular Biologists; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Chen, B.S.; Li, C.W. On the noise enhancing of stochastic Hodgkin-Hurley neuron systems. Neural Comput. 2012, 22, 1737–1763. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.S.; Wu, W.S.; Wang, Y.C.; Li, W.H. On the Robust Circuit Design Schemes of Biochemical Networks: Steady-State Approach. IEEE Trans. Biomed. Circuits Syst. 2007, 1, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.S.; Li, C.-W. Stochastic Spatio-Temporal Dynamic Model for Gene/Protein Interaction Network in Early Drosophila Development. Gene Regul Syst. Biol. 2009, 3, 191–210. [Google Scholar]
- Chen, B.S.; Wu, W.S. Underlying Principles of Natural Selection in Network Evolution: Systems Biology Approach. Evol. Bioinfom. 2007, 3, 245–262. [Google Scholar]
- Chen, B.S.; Wu, W.S.; Wu, W.S.; Li, W.H. On the Adaptive Design Rules of Biochemical Networks in Evolution. Evol. Bioinfom. 2007, 3, 27–39. [Google Scholar]
- Chen, B.S.; Ho, S.J. The stochastic evolutionary game for a population of biological networks under natural selection. Evol. Bioinform. Online 2014, 10, 17–38. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.S.; Zhang, W.H. Stochastic H2/H∞ Control With State-Dependent Noise. IEEE Trans. Auto. Cont. 2004, 49, 45–57. [Google Scholar] [CrossRef]
- Blake, W.J.; Kærn, M.; Cantor, C.R.; Collins, J.J. Noise in eukaryotic gene expression. Nature 2003, 422, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Arkin, A.; McAdams, H.H. Stochastic mechanisms in gene expression. Proc. Natl. Acad. Sci. USA 1997, 94, 814–819. [Google Scholar]
- Freeman, S.; Herron, J.C. Evolutionary Analysis, 3rd ed; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Freeman, M. Feedback control of intercellular signalling in development. Nature 2000, 408, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Walsh, B. Genetics and Analysis of Quantitative Traits, 1st ed.; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Chen, B.-S.; Lin, Y.-P. On the Interplay between the Evolvability and Network Robustness in an Evolutionary Biological Network: A Systems Biology Approach. Evol. Bioinform. Online 2011, 7, 201–233. [Google Scholar] [CrossRef] [PubMed]
- Popkov, Y.; Popkov, A. New Methods of Entropy-Robust Estimation for Randomized Models under Limited Data. Entropy 2014, 16, 675–698. [Google Scholar] [CrossRef]
- Mall, R.; Langone, R.; Suykens, J.A. Kernel Spectral Clustering for Big Data Networks. Entropy 2013, 15, 1567–1586. [Google Scholar] [CrossRef]
- Chen, B.-S.; Wang, Y.-C. On the attenuation and amplification of molecular noise in genetic regulatory networks. BMC Bioinform. 2006, 7. [Google Scholar] [CrossRef]
- Chen, B.-S.; Wang, Y.-C.; Wu, W.-S.; Li, W.-H. A new measure of the robustness of biochemical networks. Bioinformatics 2005, 21, 2698–2705. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-S.; Wu, W.-S. Robust filtering circuit design for stochastic gene networks under intrinsic and extrinsic molecular noises. Math. Biosci. 2008, 211, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, B.-S. State Feedback H∞ Control for a Class of Nonlinear Stochastic Systems. SIAM J. Control Optim. 2006, 44, 1973–1991. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, B.-S. On stabilizability and exact observability of stochastic systems with their applications. Automatica 2004, 40, 87–94. [Google Scholar] [CrossRef]
- Chen, B.-S.; Tseng, C.-S.; Uang, H.-J. Robustness Design of Nonlinear Dynamic Systems via Fuzzy Linear Control. IEEE Fuzzy Syst. 1999, 7, 571–585. [Google Scholar] [CrossRef]
- Chen, B.-S.; Tseng, C.-S.; Uang, H.-J. Mixed H2/H∞ Fuzzy Output Feedback Control Design for Nonlinear Dynamic Systems: An LMI Approach. IEEE Trans. Fuzzy Syst. 2000, 8, 249–265. [Google Scholar] [CrossRef]
- Chen, B.-S.; Lin, Y.-P. A Unifying Mathematical Framework for Genetic Robustness, Environmental Robustness, Network Robustness and their Trade-off on Phenotype Robustness in Biological Networks Part I: Gene Regulatory Networks in Systems and Evolutionary Biology. Evol. Bioinform. Online 2013, 9, 43–68. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-S.; Lin, Y.-P. A Unifying Mathematical Framework for Genetic Robustness, Environmental Robustness, Network Robustness and their Trade-offs on Phenotype Robustness in Biological Networks. Part II: Ecological networks. Evol. Bioinform. Online 2013, 9, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-S.; Lin, Y.-P. A Unifying Mathematical Framework for Genetic Robustness, Environmental Robustness, Network Robustness and their Trade-offs on Phenotype Robustness in Biological Networks. Part III: Synthetic Gene Networks in Synthetic Biology. Evol. Bioinform. Online 2013, 9, 87–109. [Google Scholar] [CrossRef] [PubMed]
- Klipp, E.; Herwig, R.; Kowald, A.; Wierling, C.; Lehrach, H. Systems Biology in Practice: Concepts, Implementation and Applcation; Wiley: New York, NY, USA, 2005. [Google Scholar]
- Murray, J.D. Mathematical Biology, 3rd ed; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Zhang, W.; Chen, B.-S.; Tseng, C.-S. Robust H∞ Filtering for Nonlinear Stochastic Systems. IEEE Trans. Sig. Process 2005, 53, 589–598. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, H.; Chen, B.-S. Stochastic H2/H∞ control with (x, u, v)-dependent noise: Finite horizon case. Automatica 2006, 42, 1891–1898. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, B.-S. H-Representation and Applications to Generalized Lyapunov Equations and Linear Stochastic Systems. IEEE Trans. Auto. Cont. 2012, 57, 3009–3022. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, B.-S.; Sheng, L.; Gao, M. Robust H2/H∞ Filter Design for a Class of Nonlinear Stochastic Systems with State-Dependent Noise. Math. Probl. Eng. 2012, 2012, 1–16. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.-S.; Wong, S.-W.; Li, C.-W. On the Calculation of System Entropy in Nonlinear Stochastic Biological Networks. Entropy 2015, 17, 6801-6833. https://0-doi-org.brum.beds.ac.uk/10.3390/e17106801
Chen B-S, Wong S-W, Li C-W. On the Calculation of System Entropy in Nonlinear Stochastic Biological Networks. Entropy. 2015; 17(10):6801-6833. https://0-doi-org.brum.beds.ac.uk/10.3390/e17106801
Chicago/Turabian StyleChen, Bor-Sen, Shang-Wen Wong, and Cheng-Wei Li. 2015. "On the Calculation of System Entropy in Nonlinear Stochastic Biological Networks" Entropy 17, no. 10: 6801-6833. https://0-doi-org.brum.beds.ac.uk/10.3390/e17106801