
Molecular Identification of Phytoplasmas Infecting Diseased Pine Trees in the UNESCO-Protected Curonian Spit of Lithuania


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Samples and DNA Extraction
2.2. Amplification of 16S rRNA and secA Gene Sequences
2.3. Cloning and Nucleotide Sequences

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytoplasma strain and RFLP based classification | Abbreviation | Plant host | GenBank accession no. |
|---|---|---|---|
| Pine dwarf yellow needle 16SrXXI-A | PineDYN27 | Pinus mugo Turra | KR051474 * |
| Pine dwarf red needle 16SeXXI-A | PineDRN12 | Pinus mugo | KR051470 * |
| Pine red short needle 16SrXXI-A | PineRShN | Pinus mugo | KF801674 * KF791911 ** |
| Pine bunchy top 16SrXXI-A | PineBT | Pinus sylvestris L. | GU289676 * KF791913 ** KF791912 *** |
| Pine dwarf yellow needle 16SrXXI-A | PineDYN12 | Pinus sylvestris | KR051471 * |
| Pine ball-like 16SrXXI-A | PineBL | Pinus mugo | KR051475 * KR051469 ** |
| Pine proliferation decline 16SrXXI-A | PinePD13 | Pinus mugo | KR051473 * |
| Pine sessile needle 16SrXXI-A | PineSN13 | Pinus mugo | KR051472 * |
| Pine dwarf necrotic needle 16SrI-A | PineDNN14 | Pinus sylvestris | KR054620 * |
| Pine decline 16SrI-A | PineD14 | Pinus mugo | KR054619 * |
| Pine yellow short needle 16SrXXI-A | PineYShN | Pinus sylvestris | KF791914 ** |
| 16SrXXI-A | PsylLap2 | Pinus sylvestris var. lapponica | KF791910 ** |
2.4. RFLP Analysis and 16Sr Group/Subgroup Classification
2.5. Phylogenetic Analysis
3. Results
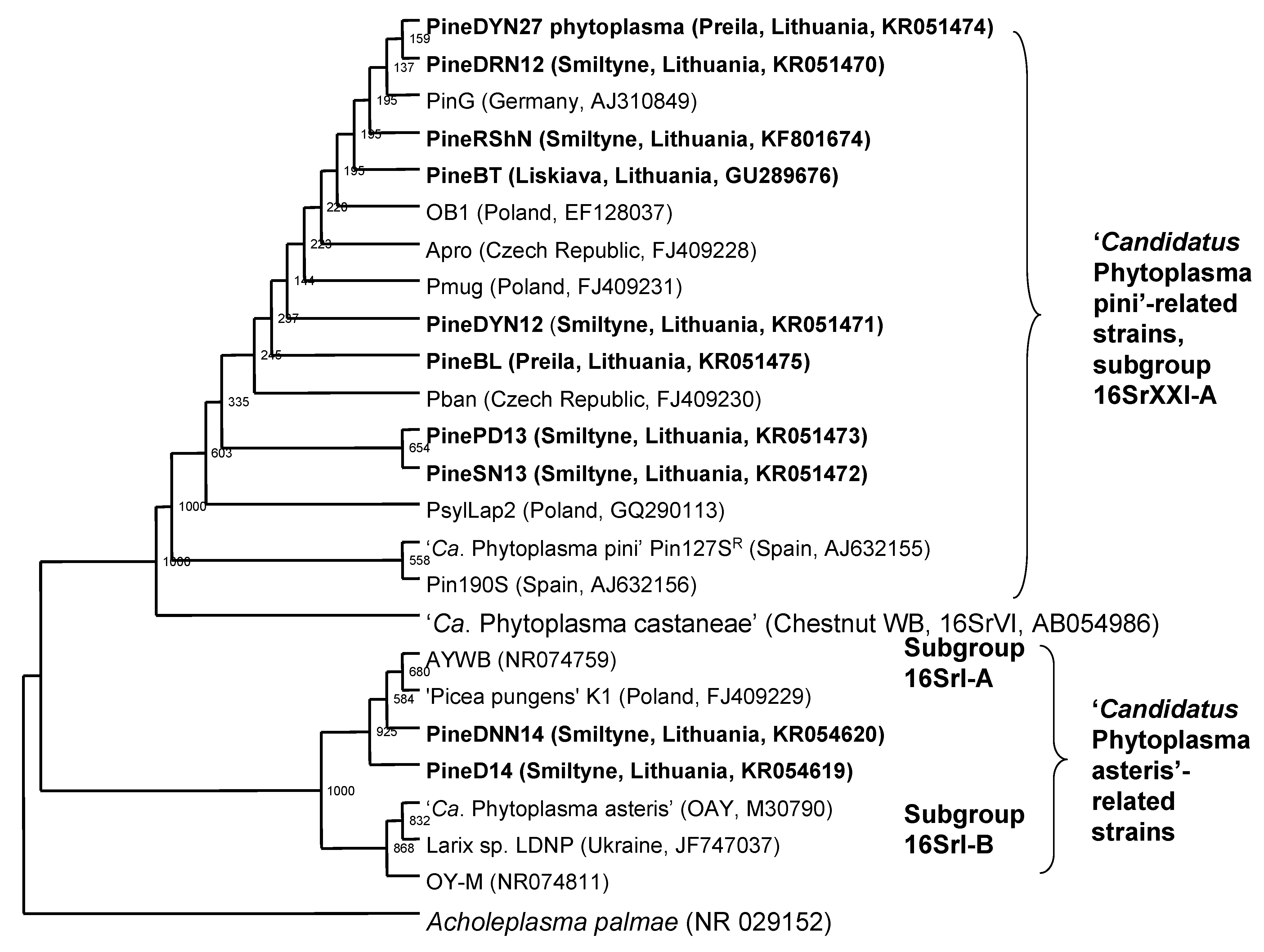
3.1. Detection and Classification of the Ca. Phytoplasma pini- and Ca. Phytoplasma asteris-Related Phytoplasma Strains



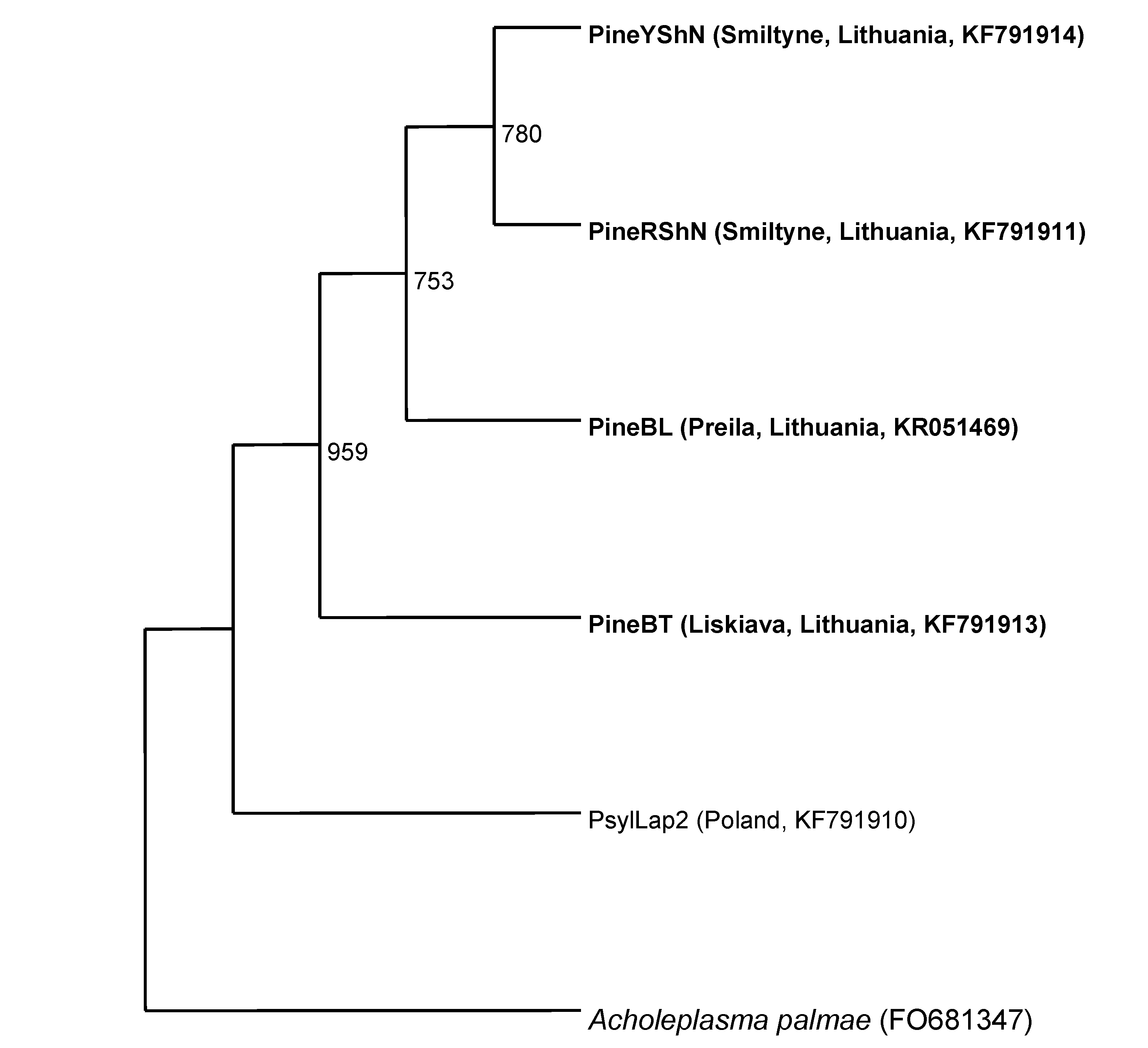
3.2. SecA Gene from the PineBT Phytoplasma
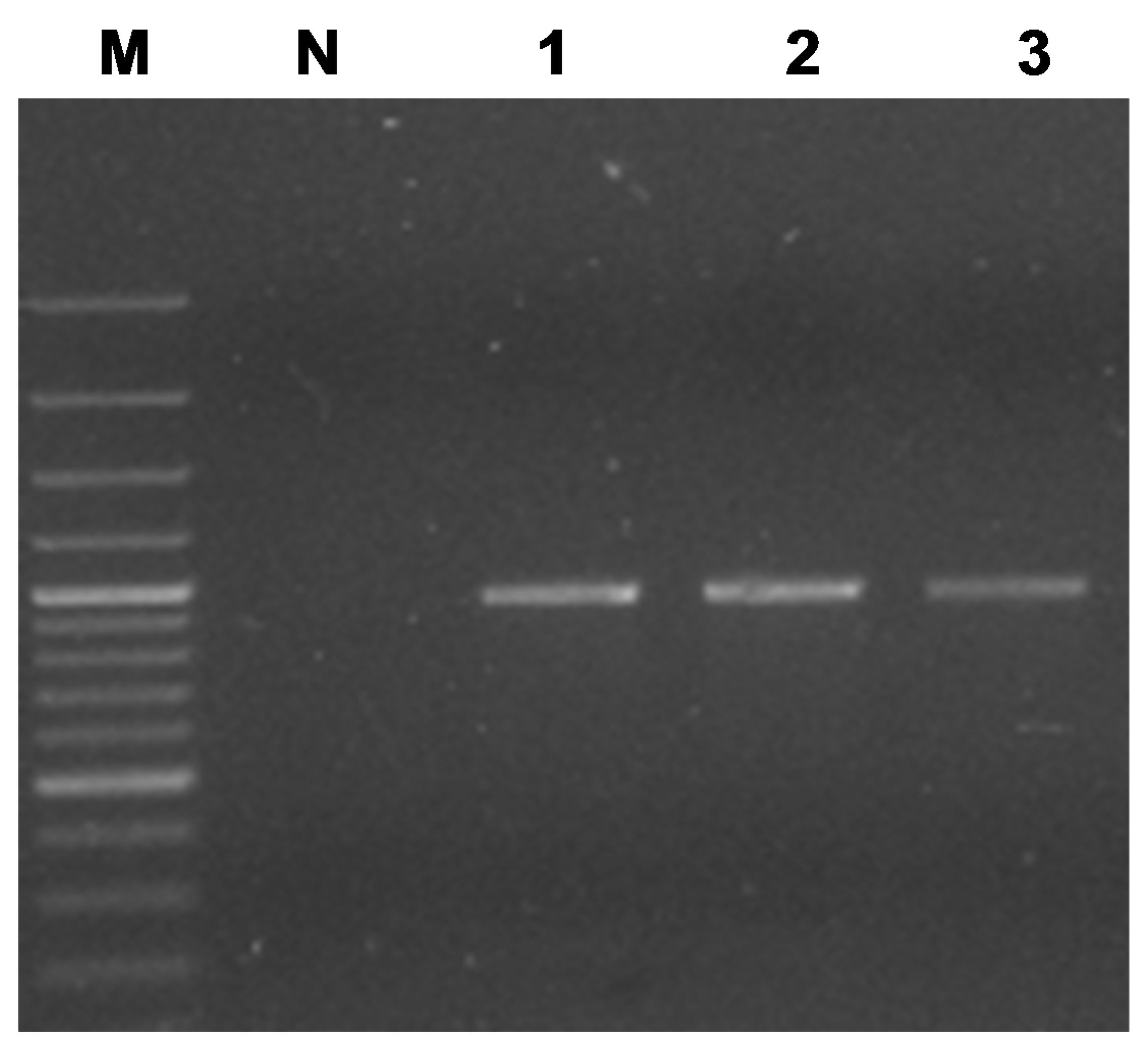
3.3. Primers for Nested or Direct PCR for secA-Based Detection of Ca. Phytoplasma pini

3.4. Phylogenetic Analysis and Alignments of 16S rDNA and secA Gene Sequences


4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mizaras, S.; Mizaraite, D.; Lebedys, A.; Pivoriunas, A.; Belova, O. Lithuania. Acta Silv. Lign. Hung. 2005, 1, 437–466, Special Edition. [Google Scholar]
- Jomantiene, R.; Davis, R.E.; Alminaite, A.; Staniulis, J.; Valiunas, D. Ribosomal RNA interoperon sequence heterogeneity in new phytoplasma lineages infecting oak, campion, thistle, and dandelion. In Proceedings of the American Phytopathological Society, Potomac Division Meeting Abstracts, Williamsburg, VA, USA, 4–6 March 2002; Available online: http://eurekamag.com/research/035/670/035670482.php (accessed on 4 March 2002).
- Valiunas, D.; Alminaite, A.; Staniulis, J.; Jomantiene, R.; Davis, R.E. First report of alder yellows phytoplasma in the Eastern Baltic Region. Plant Dis. 2001, 85, 1120. [Google Scholar] [CrossRef]
- Valiunas, D. Identification of Phytoplasmas in Lithuania and Estimation of Their Biodiversity and Molecular Evolutionary Relationships. Ph.D. Thesis, Institute of Botany and Vilnius University, Vilnius, Lithuania, 2003. [Google Scholar]
- Aučina, A.; Rudawska, M.; Leski, T.; Ryliškis, D.; Pietras, M.; Riepšas, E. Ectomycorrhizal fungal communities on seedlings and conspecific trees of Pinus mugo grown on the coastal dunes of the Curonian Spit in Lithuania. Mycorrhiza 2011, 21, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, P.G.; Beanland, L. Insect vectors of phytoplasma. Annu. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.E.; Lee, I.M. Phytoplasma. In Encyclopedia of Microbiology, 2nd ed.; Lederberg, J., Ed.; Academic Press Inc.: San Diego, CA, USA, 2000; pp. 640–646. [Google Scholar]
- Lee, I.M.; Gundersen-Rindal, D.E.; Davis, R.E.; Bartoszyk, I. Revised classification scheme of phytoplasmas based on RFLP analyses of 16S rRNA and ribosomal protein gene sequences. Int. J. Syst. Bacteriol. 1998, 48, 1153–1169. [Google Scholar] [CrossRef]
- Marcone, C.; Lee, I.M.; Davis, R.E.; Ragozzino, A.; Seemüller, E. Classification of aster yellows-group phytoplasmas based on combined analyses of rRNA and tuf gene sequences. Int. J. Syst. Evol. Microbiol. 2000, 50, 1703–1713. [Google Scholar] [PubMed]
- Davis, R.E.; Dally, E.L. Revised subgroup classification of group 16SrV phytoplasmas and placement of flavescence dorée-associated phytoplasmas in two distinct subgroups. Plant Dis. 2001, 85, 790–797. [Google Scholar] [CrossRef]
- Jomantiene, R.; Davis, R.E.; Valiunas, D.; Alminaite, A. New group 16SrIII phytoplasma lineages in Lithuania exhibit rRNA interoperon sequence heterogeneity. Eur. J. Plant Pathol. 2002, 108, 507–517. [Google Scholar] [CrossRef]
- Jomantiene, R.; Zhao, Y.; Lee, I.M.; Davis, R.E. Phytoplasmas infecting sour cherry and lilac represent two distinct lineages having close evolutionary affinities with clover phyllody phytoplasma. Eur. J. Plant Pathol. 2011, 130, 97–107. [Google Scholar] [CrossRef]
- Valiunas, D.; Jomantiene, R.; Davis, R.E. Establishment of a new phytoplasma subgroup, 16SrI-Q, to accomodate a previously undescribed phytoplasma found in diseased cherry in Lithuania. J. Plant Pathol. 2009, 91, 71–75. [Google Scholar]
- Valiunas, D.; Jomantiene, R.; Ivanauskas, A.; Abraitis, R.; Staniene, G.; Zhao, Y.; Davis, R.E. First report of a new phytoplasma subgroup, 16SrIII-T, associated with decline disease affecting sweet and sour cherry trees in Lithuania. Plant Dis. 2009, 93, 550. [Google Scholar] [CrossRef]
- Wei, W.; Davis, R.E.; Lee, I.M.; Zhao, Y. Computer-simulated RFLP analysis of 16S rRNA genes: Identification of ten new phytoplasma groups. Int. J. Syst. Evol. Microbiol. 2007, 57, 1855–1867. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sun, Q.; Wei, W.; Davis, R.E.; Wu, Q.; Liu, Q. Candidatus Phytoplasma tamaricis, a novel taxon discovered in witches’-broom-diseased salt cedar (Tamarix chinensis Lour.). Int. J. Syst. Evol. Microbiol. 2009, 59, 2496–2504. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.E.; Zhao, Y.; Dally, E.L.; Lee, I.M.; Jomantiene, R.; Douglas, S.M. Candidatus Phytoplasma pruni, a novel taxon associated with X-disease of stone fruits, Prunus spp.: Multilocus characterization based on 16S rRNA, secY, and ribosomal protein genes. Int. J. Syst. Evol. Microbiol. 2013, 63, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.E.; Dally, E.; Zhao, Y.; Lee, I.M.; Jomantiene, R.; Detweiler, A.J.; Putnam, M.L. First report of a new subgroup 16SrIX-E (Candidatus Phytoplasma phoenicium-related) phytoplasma associated with juniper witches’ broom disease in Oregon, USA. Plant Pathol. 2010, 59, 1161. [Google Scholar] [CrossRef]
- Jomantiene, R.; Valiunas, D.; Ivanauskas, A.; Urbanaviciene, L.; Staniulis, J.; Davis, R.E. Larch is a new host for a group 16SrI, subgroup B phytoplasma in Ukraine. B. Insectol. 2011, 64, S101–S102. [Google Scholar]
- Kaminska, M.; Berniak, H.; Obdrzalek, J. New natural host plants of Candidatus Phytoplasma pini in Poland and the Czech Republic. Plant Pathol. 2011, 60, 1023–1029. [Google Scholar] [CrossRef]
- Schneider, B.; Torres, E.; Martin, M.P.; Schröder, M.; Behnke, H.D.; Seemüller, E. Candidatus Phytoplasma pini, a novel taxon from Pinus silvestris and Pinus halepensis. Int. J. Syst. Evol. Microbiol. 2005, 55, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Śliwa, H.; Kaminska, M.; Korszun, S.; Adler, P. Detection of Candidatus Phytoplasma pini in Pinus sylvestris trees in Poland. J. Phytopathol. 2008, 156, 88–92. [Google Scholar] [CrossRef]
- Valiunas, D.; Jomantiene, R.; Ivanauskas, A.; Sneideris, D.; Staniulis, J.; Davis, R.E. A possible threat to the timber industry: Candidatus Phytoplasma pini in Scots pine (Pinus sylvestris L.) in Lithuania. In Current Status and Perspectives of Phytoplasma Disease Research and Management, Proceedings of the Abstract book of the combined meeting of Work Groups 1–4, COST Action FA0807, Sitges, Spain, 1–2 February 2010; Bertaccini, A., Lavifia, A., Torres, E., Eds.; p. 38.
- Valiunas, D.; Jomantiene, R.; Ivanauskas, A.; Urbonaite, I.; Davis, R.E. Molecular identification of pine tree-infecting phytoplasmas from the UNESCO-protected Curonian Spit in Lithuania. In Proceedings of the Joint IUFRO Working Party Meetings, Uppsala, Sweden, 7–12 June 2015; p. 89.
- Lee, I.M.; Martini, M.; Bottner, K.D.; Dane, R.A.; Black, M.C.; Troxclair, N. Ecological implications from a molecular analysis of phytoplasmas involved in an aster yellows epidemic in various crops in Texas. Phytopathology 2003, 93, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.M.; Martini, M.; Marcone, C.; Zhu, S.F. Classification of phytoplasma strains in the elm yellows group (16SrV) and proposal of Candidatus Phytoplasma ulmi for the phytoplasma associated with elm yellows. Int. J. Syst. Evol. Microbiol. 2004, 54, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Hiruki, C. Amplification of 16S rRNA genes from culturable and non-culturable mollicutes. J. Microbiol. Meth. 1991, 14, 53–61. [Google Scholar] [CrossRef]
- Schneider, B.; Seemüller, E.; Smart, C.D.; Kirkpatrick, B.C. Phylogenetic classification of plant pathogenic mycoplasma-like organisms or phytoplasmas. In Molecular and Diagnostic Procedures in Mycoplasmology; Razin, S., Tully, J.G., Eds.; Academic Press: San Diego, CA, USA, 1995; Volume 1, pp. 369–380. [Google Scholar]
- Gundersen, D.E.; Lee, I.M. Ultrasensitive detection of phytoplasmas by nested-PCR assays using two universal primer pairs. Phytopathol. Mediterr. 1996, 35, 144–151. [Google Scholar]
- Lee, I.-M.; Bottner, K.D.; Munyaneza, J.E.; Davis, R.E.; Crosslin, J.M.; du Toit, L.J. Carrot purple leaf: A new spiroplasma disease associated with carrots in Washington state. Plant Dis. 2006, 90, 989–993. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, W.; Lee, I.M.; Shao, J.; Suo, X.; Davis, R.E. Construction of an interactive online phytoplasma classification tool, iPhyClassifier, and its application in analysis of the peach X-disease phytoplasma group (16SrIII). Int. J. Syst. Evol. Microbiol. 2009, 59, 2582–2593. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar] [PubMed]
- Paltrinieri, S.; Martini, M.; Pondrelli, M.; Bertaccini, A. X-disease related phytoplasmas in ornamental trees and shrubs with witches’-broom and malformation symptoms. J. Plant Pathol. 1998, 80, 261. [Google Scholar]
- Ježić, M.; Poljak, I.; Šafarić, B.; Idžojtić, M.; Ćurković-Perica, M. Candidatus Phytoplasma pini in pine species in Croatia. J. Plant Dis. Prot. 2013, 120, 160–163. [Google Scholar]
- Kaminska, M.; Berniak, H. Detection and identification of three Candidatus Phytoplasma species in Picea spp. trees in Poland. J. Phytopathol. 2011, 159, 796–798. [Google Scholar] [CrossRef]
- Gupta, M.K.; Samad, A.; Shasany, A.K.; Ajayakumar, P.V.; Alam, M. First report of a 16SrVI Candidatus Phytoplasma trifolii isolate infecting Norfolk Island pine (Araucaria heterophylla) in India. Plant Pathol. 2009, 59, 399. [Google Scholar] [CrossRef]
- Huang, S.; Tiwari, A.K.; Rao, G.P. Candidatus Phytoplasma pini affecting Taxodium distichum var. imbricarium in China. Phytopathogenic Mollicutes 2011, 1, 91–94. [Google Scholar] [CrossRef]
- Hodgetts, J.; Boonham, N.; Mumford, R.; Harrison, N.; Dickinson, M. Phytoplasma phylogenetics based on analysis of secA and and 23S rRNA gene sequences for improved resolution of candidate species of Candidatus Phytoplasma. Int. J. Syst. Evol. Microbiol. 2008, 58, 1826–1837. [Google Scholar] [CrossRef] [PubMed]
- Makarova, O.; Contaldo, N.; Paltrinieri, S.; Bertaccini, A.; Nyskjold, H.; Nicolaisen, M. DNA bar-coding for phytoplasma identification. Methods Mol. Biol. 2013, 938, 301–317. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valiunas, D.; Jomantiene, R.; Ivanauskas, A.; Urbonaite, I.; Sneideris, D.; Davis, R.E. Molecular Identification of Phytoplasmas Infecting Diseased Pine Trees in the UNESCO-Protected Curonian Spit of Lithuania. Forests 2015, 6, 2469-2483. https://0-doi-org.brum.beds.ac.uk/10.3390/f6072469
Valiunas D, Jomantiene R, Ivanauskas A, Urbonaite I, Sneideris D, Davis RE. Molecular Identification of Phytoplasmas Infecting Diseased Pine Trees in the UNESCO-Protected Curonian Spit of Lithuania. Forests. 2015; 6(7):2469-2483. https://0-doi-org.brum.beds.ac.uk/10.3390/f6072469
Chicago/Turabian StyleValiunas, Deividas, Rasa Jomantiene, Algirdas Ivanauskas, Indre Urbonaite, Donatas Sneideris, and Robert E. Davis. 2015. "Molecular Identification of Phytoplasmas Infecting Diseased Pine Trees in the UNESCO-Protected Curonian Spit of Lithuania" Forests 6, no. 7: 2469-2483. https://0-doi-org.brum.beds.ac.uk/10.3390/f6072469