A Novel Insight into Functional Divergence of the MST Gene Family in Rice Based on Comprehensive Expression Patterns


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Identification of the MST Genes
2.3. Phylogenetic Analysis, Gene Structure, and Conserved Motifs
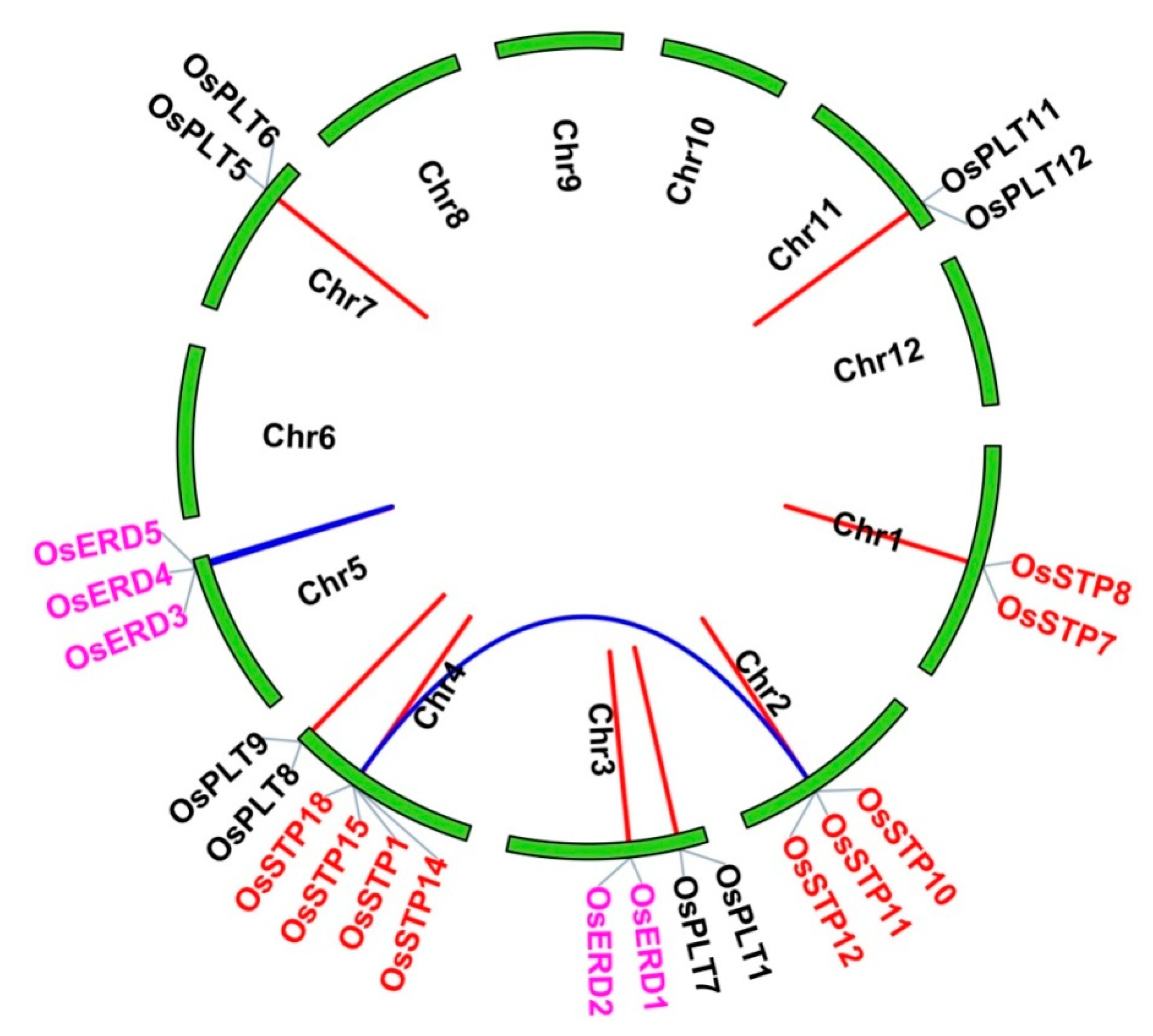
2.4. Chromosomal Locations, Gene Duplication Events, and Ka/Ks Values
2.5. Expression Analysis of MST Genes by qRT-PCR
2.6. Expression Correlation between MST Paralogs under Hormone and Abiotic Stress Treatments
3. Results
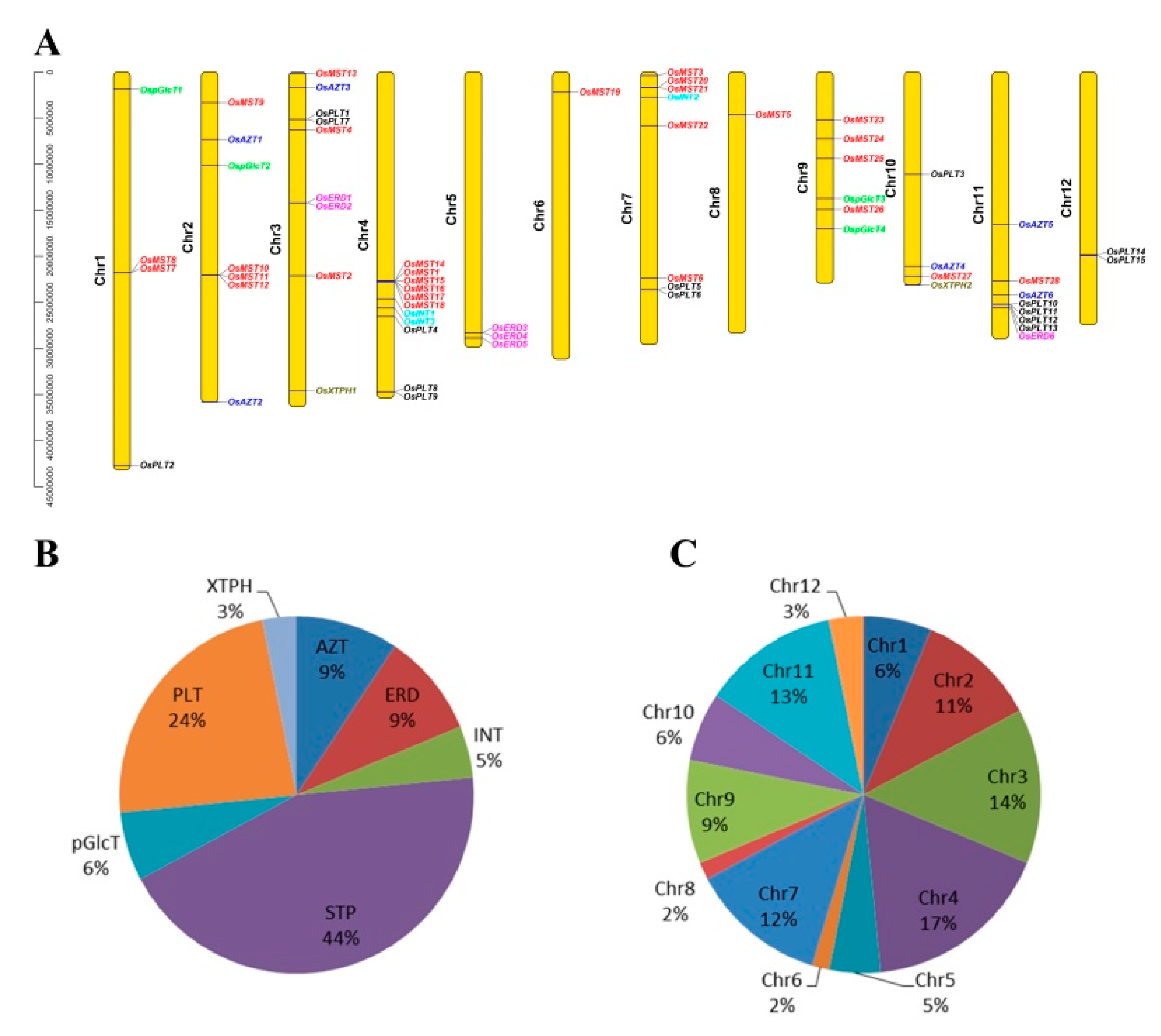
3.1. Identification, Classification, Chromosome Locations, Duplication Events, and Selection Pressure of the MST Genes
3.2. Gene Structure and Conserved Motifs
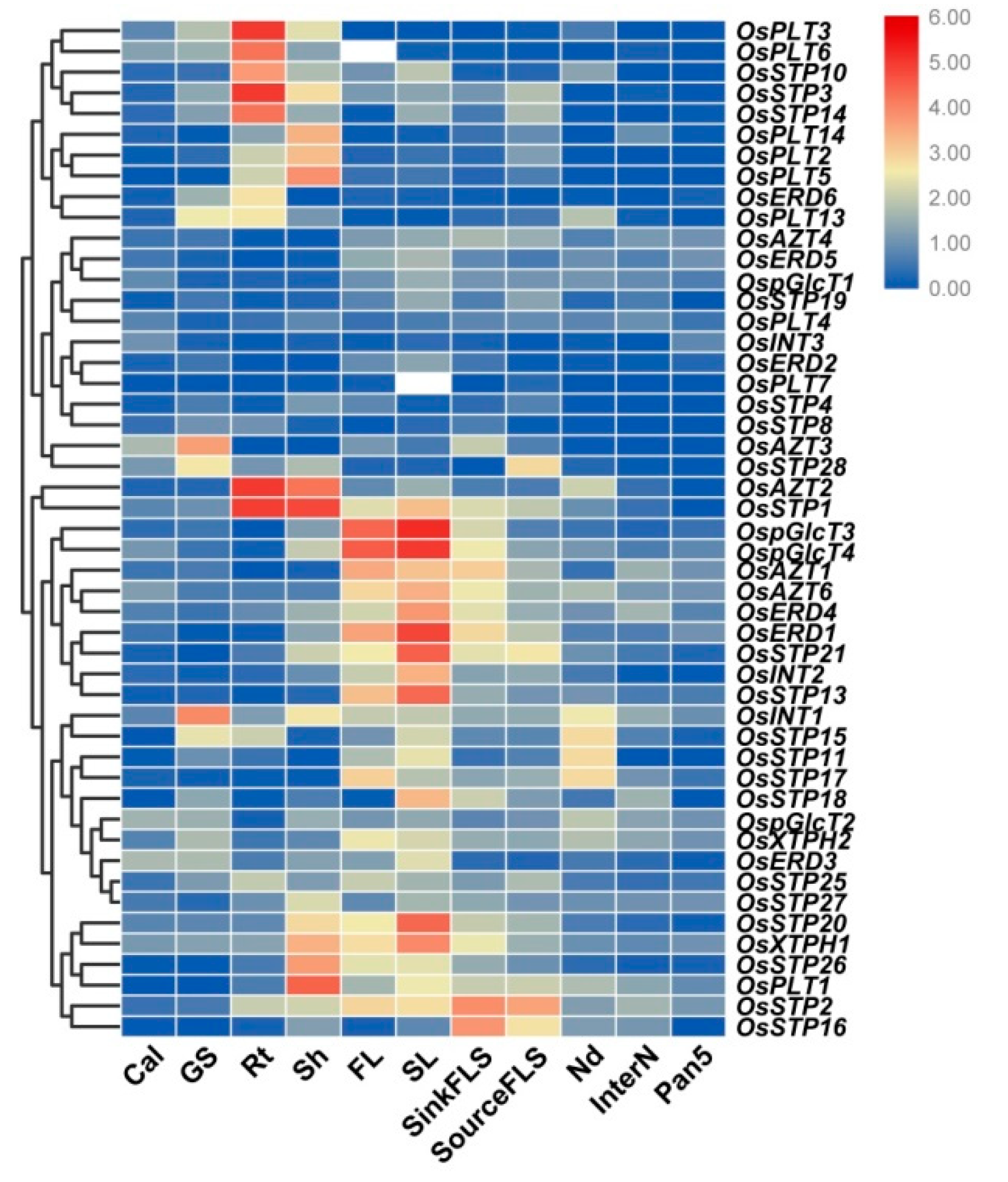
3.3. Expression Analysis of MST Genes in Various Tissues
3.4. Expression Analysis of MST Genes under Abiotic Stresses
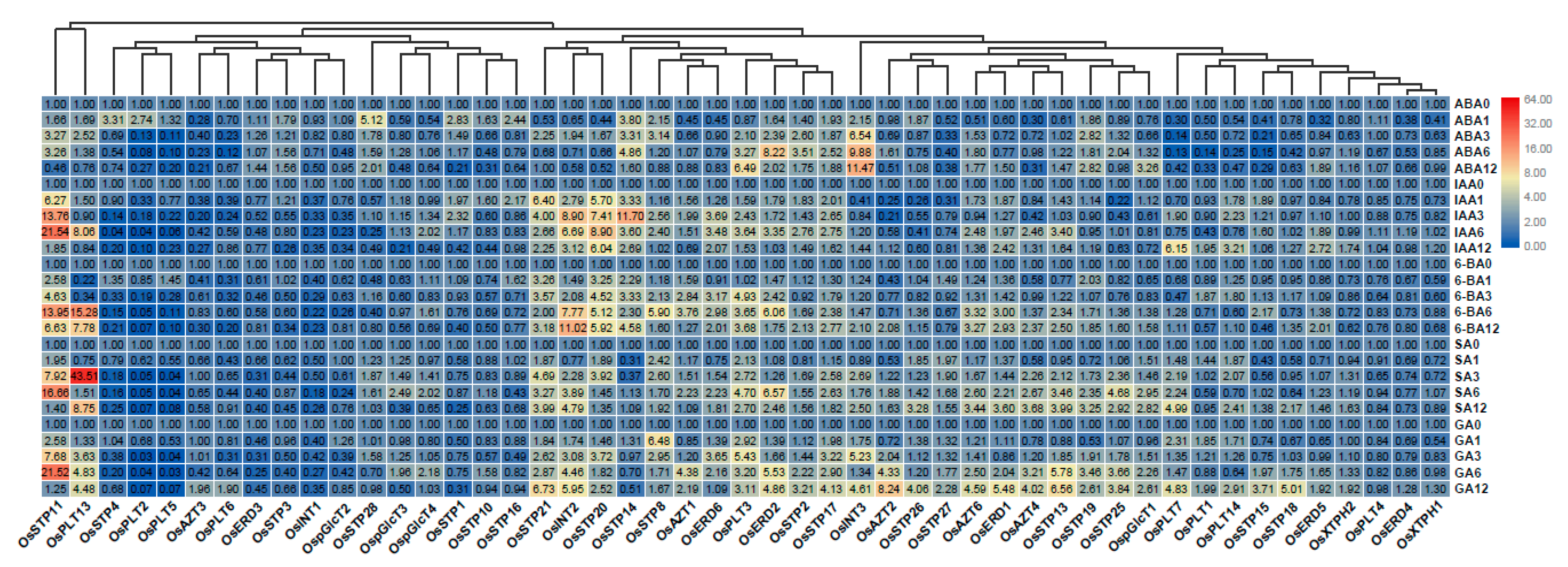
3.5. Expression Analysis of MST Genes under Hormone Treatments
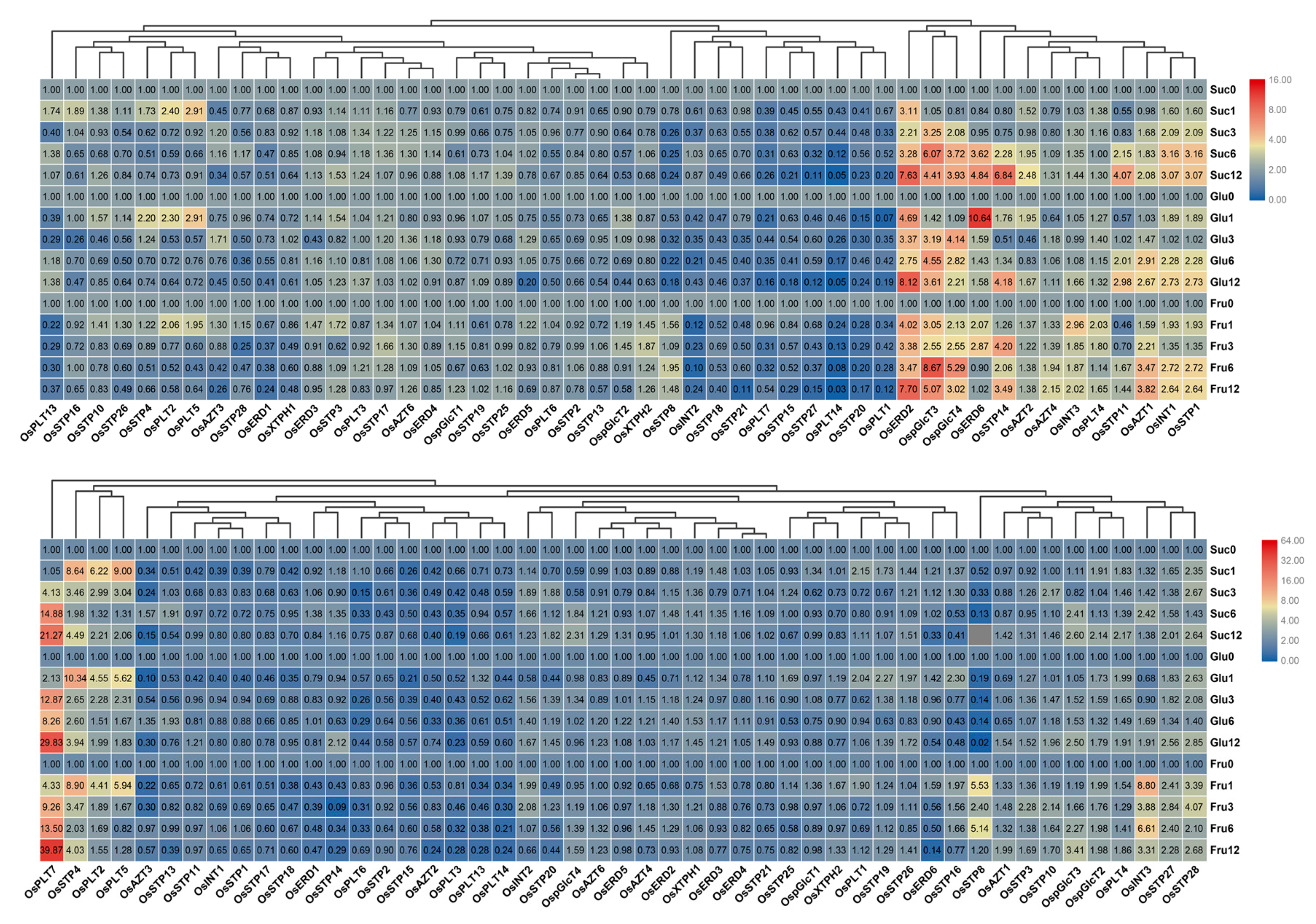
3.6. Expression Analysis of MST Genes under Sugar Treatments
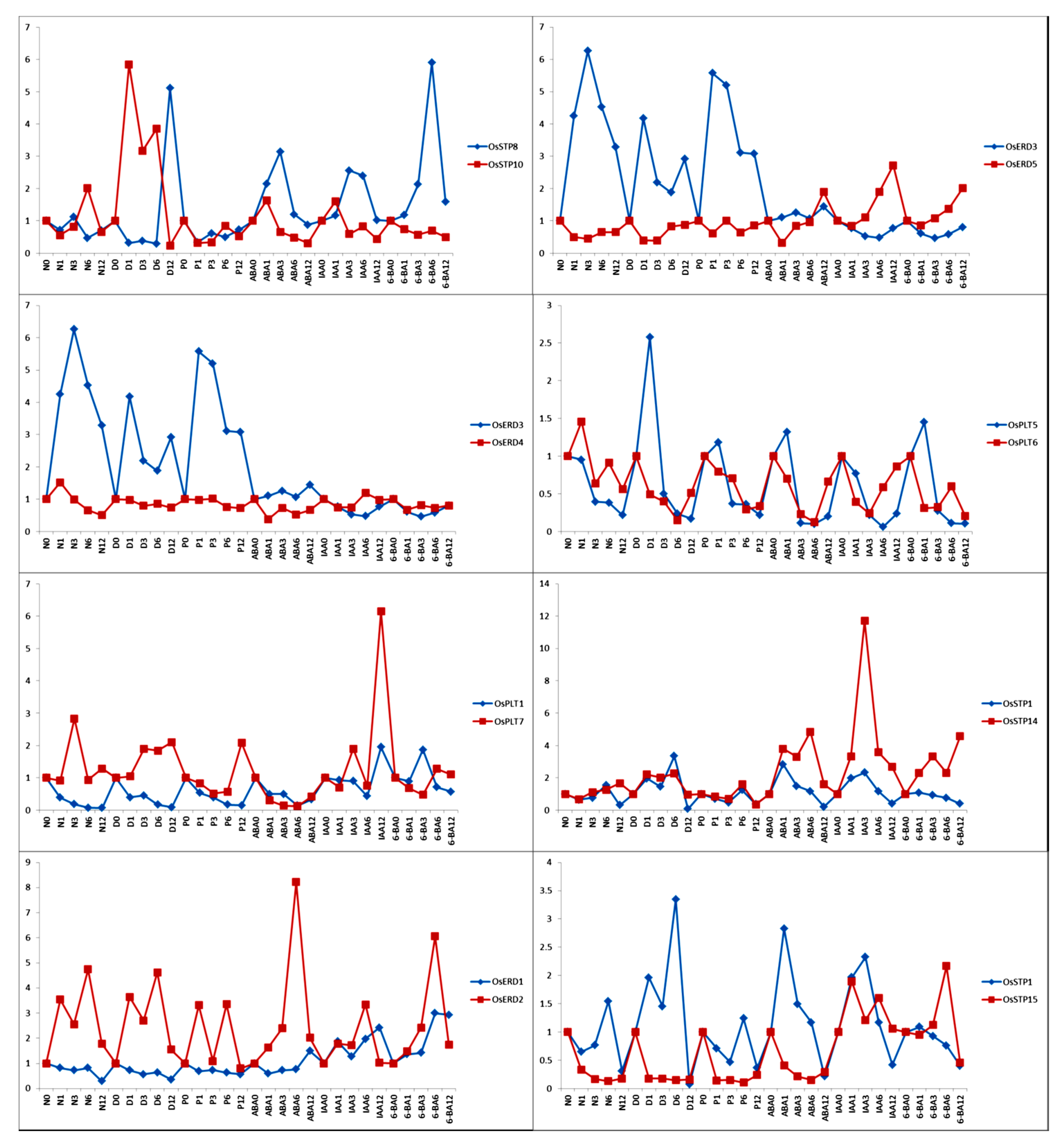
3.7. Functional Differentiation of MST Duplication Genes under Hormone Treatments and Abiotic Stress Treatments
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, Q.; Dang, H.; Chen, Z.; Wu, J.; Chen, Y.; Chen, S.; Luo, L. Genome-wide identification, expression, and functional analysis of the sugar transporter gene family in cassava (Manihot esculenta). Int. J. Mol. Sci. 2018, 19, 987. [Google Scholar] [CrossRef] [PubMed]
- Jung-Il, C.; Bo, B.; Dae-Woo, L.; Nayeon, R.; Soon-Kwan, H.; Hyun-Bi, K.; Joon-Seob, E.; Sang-Bong, C.; Man-Ho, C.; Seong Hee, B. Expression analysis and functional characterization of the monosaccharide transporters, OsTMTs, involving vacuolar sugar transport in rice (Oryza sativa). New Phytol. 2010, 186, 657–668. [Google Scholar]
- Tian, L.; Liu, L.; Yin, Y.; Huang, M.; Chen, Y.; Xu, X.; Wu, P.; Li, M.; Wu, G.; Jiang, H. Heterogeneity in the expression and subcellular localization of POLYOL/MONOSACCHARIDE Transporter genes in Lotus japonicus. PLoS ONE 2017, 12, e0185269. [Google Scholar] [CrossRef] [PubMed]
- Rottmann, T.M.; Klebl, F.; Schneider, S.; Kischka, D.; Rüscher, D.; Sauer, N.; Stadler, R. Characterization of three sugar transporters, STP7, STP8 and STP12. Plant Physiol. 2018, 176, 2330–2350. [Google Scholar] [CrossRef]
- Julius, B.T.; Leach, K.A.; Tran, T.M.; Mertz, R.A.; Braun, D.M. Sugar transporters in plants: New insights and discoveries. Plant Cell Physiol. 2017, 58, 1442. [Google Scholar] [CrossRef]
- Zheng, Q.M.; Tang, Z.; Xu, Q.; Deng, X.X. Isolation, phylogenetic relationship and expression profiling of sugar transporter genes in sweet orange (Citrus sinensis). Plant Cell Tissue Org. 2014, 119, 609–624. [Google Scholar] [CrossRef]
- Klemens, P.A.W.; Kathrin, P.; Joachim, D.; Lara, S.; Rozenn, L.H.; Catherine, B.; Magali, B.; Fabien, C.; Anne, K.; Ekkehard, N. Overexpression of the vacuolar sugar carrier AtSWEET16 modifies germination, growth, and stress tolerance in Arabidopsis. Plant Physiol. 2013, 163, 1338–1352. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, S.; Yu, F.; Tang, J.; Yu, L.; Wang, H.; Li, J. Genome-wide identification and expression profiling of sugar transporter protein (STP) family genes in cabbage (Brassica oleracea var. capitata L.) reveals their involvement in clubroot disease responses. Genes 2019, 10, 71. [Google Scholar] [CrossRef]
- Buttner, M. The monosaccharide transporter(-like) gene family in Arabidopsis. FEBS Lett. 2007, 581, 2318–2324. [Google Scholar] [CrossRef]
- Buttner, M. The Arabidopsis sugar transporter (AtSTP) family: An update. Plant Biol. 2010, 12, 35–41. [Google Scholar] [CrossRef]
- Alexandra, W.; Oliver, T.; Ingmar, F.; Christian, L.; Joachim, T.; Stefan, M.; Ulrike, S.; Enrico, M.; H Ekkehard, N. Molecular identification and physiological characterization of a novel monosaccharide transporter from Arabidopsis involved in vacuolar sugar transport. Plant Cell 2006, 18, 3476–3490. [Google Scholar]
- Afoufa-Bastien, D.; Medici, A.; Jeauffre, J.; Coutos-Thévenot, P.; Lemoine, R.; Atanassova, R.; Laloi, M. The Vitis vinifera sugar transporter gene family: Phylogenetic overview and macroarray expression profiling. BMC Plant Biol. 2010, 10, 245. [Google Scholar] [CrossRef] [PubMed]
- Emiko, O.K.; Takumi, H.; Yukio, K.; Natsumaro, K.; Junji, Y.; Seiichiro, H. Sucrose transporter NtSUT4 from tobacco BY-2 involved in plant cell shape during miniprotoplast culture. J. Plant Res. 2011, 124, 395–403. [Google Scholar]
- Mccurdy, D.W.; Dibley, S.; Cahyanegara, R.; Martin, A.; Patrick, J.W. Functional characterization and RNAi-mediated suppression reveals roles for hexose transporters in sugar accumulation by tomato fruit. Mol. Plant 2010, 3, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Aleksandra, H.; Nicolas, S.; Fernando, C.; Fernie, A.R.; Bernhard, G.; Christina, K. Sucrose transporter LeSUT1 and LeSUT2 inhibition affects tomato fruit development in different ways. Plant J. 2006, 45, 180–192. [Google Scholar]
- Doidy, J.; van Tuinen, D.; Lamotte, O.; Corneillat, M.; Alcaraz, G.; Wipf, D. The Medicago truncatula sucrose transporter family: Characterization and implication of key members in carbon partitioning towards arbuscular mycorrhizal fungi. Mol. Plant 2012, 5, 1346–1358. [Google Scholar] [CrossRef] [PubMed]
- Laloi, M. Regulation of RhSUC2, a sucrose transporter, is correlated with the light control of bud burst in Rosa sp. Plant Cell Environ. 2011, 34, 1776–1789. [Google Scholar]
- Johnson, D.A.; Hill, J.P.; Thomas, M.A. The monosaccharide transporter gene family in land plants is ancient and shows differential subfamily expression and expansion across lineages. BMC Evol. Biol. 2006, 6, 64. [Google Scholar]
- Johnson, D.A.; Thomas, M.A. The monosaccharide transporter gene family in Arabidopsis and rice: A history of duplications, adaptive evolution, and functional divergence. Mol. Biol. Evol. 2007, 24, 2412–2423. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, H.; Wei, X.; Chai, C.; Xiao, Y.; Zhang, Y.; Chen, B.; Xiao, G.; Ouwerkerk, P.B.; Wang, M.; et al. Molecular cloning and expression analysis of a monosaccharide transporter gene OsMST4 from rice (Oryza sativa L.). Plant Mol. Biol. 2007, 65, 439–451. [Google Scholar] [CrossRef]
- Schneider, S.; Beyhl, D.; Hedrich, R.N. Functional and physiological characterization of Arabidopsis INOSITOL TRANSPORTER1, a novel tonoplast-localized transporter for myo-inositol. Plant Cell 2008, 20, 1073–1087. [Google Scholar] [CrossRef]
- Poschet, G.; Hannich, B.; Büttner, M. Identification and characterization of AtSTP14, a novel galactose transporter from Arabidopsis. Plant Cell Physiol. 2010, 51, 1571. [Google Scholar] [CrossRef]
- Yvonne-Simone, K.; Dietmar, G.; Ruth, S.; Franz, K.; Lucie, L.A.; Rémi, L.; Rainer, H.; Norbert, S. Arabidopsis POLYOL TRANSPORTER5, a new member of the monosaccharide transporter-like superfamily, mediates H+-Symport of numerous substrates, including myo-inositol, glycerol, and ribose. Plant Cell 2005, 17, 204–218. [Google Scholar]
- Klepek, Y.S.; Volke, M.; Kai, R.K.; Wippel, K.; Hoth, S.; Hedrich, R.; Sauer, N. Arabidopsis thaliana POLYOL/MONOSACCHARIDE TRANSPORTERS 1 and 2: Fructose and xylitol/H+ symporters in pollen and young xylem cells. J. Exp. Bot. 2010, 61, 537–550. [Google Scholar] [CrossRef]
- Schulz, A.; Beyhl, D.; Marten, I.; Wormit, A.; Neuhaus, E.; Poschet, G.; Büttner, M.; Schneider, S.; Sauer, N.; Hedrich, R. Proton-driven sucrose symport and antiport are provided by the vacuolar transporters SUC4 and TMT1/2. Plant J. 2011, 68, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Man-Ho, C.; Hyemin, L.; Ho, S.D.; Jong-Seong, J.; Seong Hee, B.; Youn-Il, P.; Tae-Ryong, H. Role of the plastidic glucose translocator in the export of starch degradation products from the chloroplasts in Arabidopsis thaliana. New Phytol. 2011, 190, 101–112. [Google Scholar]
- Quirino, B.F.; Reiter, W.D.; Amasino, R.D. One of two tandem Arabidopsis genes homologous to monosaccharide transporters is senescence-associated. Plant Mol. Biol. 2001, 46, 447–457. [Google Scholar] [CrossRef]
- Kohji, Y.; Yuriko, O.; Junya, M.; Kazuo, N.; Yasunari, F.; Kazuo, S.; Kazuko, Y.S. Functional analysis of an Arabidopsis thaliana abiotic stress-inducible facilitated diffusion transporter for monosaccharides. J. Biol. Chem. 2010, 285, 1138–1146. [Google Scholar]
- Toyofuku, K.; Kasahara, M.; Yamaguchi, J. Characterization and expression of monosaccharide transporters (OsMSTs) in rice. Plant Cell Physiol. 2000, 41, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Budsaraporn, N.; Anna, S.; Taito, T.; Kyoko, T.; Jarunya, N.; Akira, I.; Junji, Y. Characterization of rice functional monosaccharide transporter, OsMST5. Biosci. Biotech. Biochem. 2003, 67, 556–562. [Google Scholar]
- Yongqin, W.; Yuguo, X.; Yu, Z.; Chenglin, C.; Gang, W.; Xiaoli, W.; Honglin, X.; Mei, W.; Ouwerkerk, P.B.F.; Zhen, Z. Molecular cloning, functional characterization and expression analysis of a novel monosaccharide transporter gene OsMST6 from rice (Oryza sativa L.). Planta 2008, 228, 525–535. [Google Scholar]
- Cao, H.; Guo, S.; Xu, Y.; Jiang, K.; Jones, A.M.; Chong, K. Reduced expression of a gene encoding a Golgi localized monosaccharide transporter (OsGMST1) confers hypersensitivity to salt in rice (Oryza sativa). J. Exp. Bot. 2011, 62, 4595–4604. [Google Scholar] [CrossRef]
- An, B.; Lan, J.; Deng, X.; Chen, S.; Ouyang, C.; Shi, H.; Yang, J.; Li, Y. Silencing of D-Lactate Dehydrogenase Impedes Glyoxalase System and Leads to Methylglyoxal Accumulation and Growth Inhibition in Rice. Front. Plant Sci. 2017, 8, 2071. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Yang, S.; Wang, Y.; Bendahmane, M.; Fu, X. Genome-wide identification and characterization of aquaporin gene family in Beta vulgaris. PeerJ 2017, 5, e3747. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Bendahmane, M.; Fu, X. Genome-wide identification and characterization of aquaporins and their role in the flower opening processes in carnation (Dianthus caryophyllus). Molecules 2018, 23, 1895. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Jakobsen, I.B.; Nei, M. MEGA2: Molecular evolutionary genetics analysis software. Bioinformatics 2001, 19, 1244–1245. [Google Scholar] [CrossRef]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a Toolkit for Biologists integrating various HTS-data handling tools with a user-friendly interface. BioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Kong, W.; Zhong, H.; Deng, X.; Gautam, M.; Gong, Z.; Zhang, Y.; Zhao, G.; Liu, C.; Li, Y. Evolutionary analysis of GH3 genes in six Oryza species/subspecies and their expression under salinity stress in Oryza sativa ssp. japonica. Plants 2019, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.I. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 1451–1452. [Google Scholar] [CrossRef]
- Kong, W.; Zhong, H.; Gong, Z.; Fang, X.; Sun, T.; Deng, X.; Li, Y. Meta-analysis of salt stress transcriptome responses in different rice genotypes at the seedling stage. Plants 2019, 8, 64. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 2004, 16, 1667–1678. [Google Scholar] [CrossRef]
- Yim, W.C.; Lee, B.M.; Jang, C.S. Expression diversity and evolutionary dynamics of rice duplicate genes. Mol. Genet. Genom. 2009, 281, 483–493. [Google Scholar] [CrossRef]
- Li, J.-M.; Zheng, D.-M.; Li, L.-T.; Qiao, X.; Wei, S.-W.; Bai, B.; Zhang, S.-L.; Wu, J. Genome-wide function, evolutionary characterization and expression analysis of sugar transporter family genes in pear (Pyrus bretschneideri Rehd). Plant Cell Physiol. 2015, 56, 1721–1737. [Google Scholar] [CrossRef]
- Yupeng, W.; Haibao, T.; Debarry, J.D.; Xu, T.; Jingping, L.; Xiyin, W.; Tae-Ho, L.; Huizhe, J.; Barry, M.; Hui, G. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar]
- Qihui, Z.; Song, G. Phylogenetic relationships among A-genome species of the genus Oryza revealed by intron sequences of four nuclear genes. New Phytol. 2005, 167, 249–265. [Google Scholar]
- Tang, L.; Zou, X.H.; Achoundong, G.; Potgieter, C.; Second, G.; Zhang, D.Y.; Ge, S. Phylogeny and biogeography of the rice tribe (Oryzeae): Evidence from combined analysis of 20 chloroplast fragments. Mol. Phylogenet. Evol. 2010, 54, 266–277. [Google Scholar] [CrossRef]
- Xionglei, H.; Jianzhi, Z. Rapid subfunctionalization accompanied by prolonged and substantial neofunctionalization in duplicate gene evolution. Genetics 2005, 169, 1157–1164. [Google Scholar]
- Hui, Z.; Wanqi, L.; Xijia, Y.; Xue, L.; Ning, J.; Hong, M.; Dabing, Z. Carbon starved anther encodes a MYB domain protein that regulates sugar partitioning required for rice pollen development. Plant Cell 2010, 22, 672–689. [Google Scholar]
- Mamun, E.A.; Alfred, S.; Cantrill, L.C.; Overall, R.L.; Sutton, B.G. Effects of chilling on male gametophyte development in rice. Cell Biol. Int. 2006, 30, 583–591. [Google Scholar] [CrossRef]
- Buttner, M.E.; Baier, K.; Scholz, S.J.; Sontheim, M.; Lauterbach, C.; Var, H.; Sauer, N. AtSTP3, a green leaf-specific, low affinity monosaccharide-H+ symporter of Arabidopsis thaliana. Plant Cell Environ. 2010, 23, 175–184. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | TIGR Locus Tag | RAP-DB | MST Subfamily |
|---|---|---|---|
| OsAZT1 | LOC_Os02g13560.1 | Os02g0229400 | AZT |
| OsAZT2 | LOC_Os02g58530.1 | Os02g0832100 | AZT |
| OsAZT3 | LOC_Os03g03680.1 | Os03g0128900 | AZT |
| OsAZT4 | LOC_Os10g39440.2 | Os10g0539900 | AZT |
| OsAZT5 | LOC_Os11g28610.1 | Os11g0475600 | AZT |
| OsAZT6 | LOC_Os11g40540.2 | Os11g0620400 | AZT |
| OsERD1 | LOC_Os03g24860.1 | Os03g0363500 | ERD |
| OsERD2 | LOC_Os03g24870.1 | Os03g0363600 | ERD |
| OsERD3 | LOC_Os05g49260.1 | Os05g0567700 | ERD |
| OsERD4 | LOC_Os05g49270.1 | Os05g0567800 | ERD |
| OsERD5 | LOC_Os05g50280.1 | Os05g0579000 | ERD |
| OsERD6 | LOC_Os11g42430.1 | Os11g0643800 | ERD |
| OsINT1 | LOC_Os04g41460.1 | Os04g0491700 | INT |
| OsINT2 | LOC_Os07g05640.1 | Os07g0151200 | INT |
| OsINT3 | LOC_Os04g43210.1 | Os04g0511400 | INT |
| OsSTP1 | LOC_Os04g37980.1 | Os04g0452700 | STP |
| OsSTP10 | LOC_Os02g36414.1 | Os02g0573500 | STP |
| OsSTP11 | LOC_Os02g36440.1 | Os02g0574000 | STP |
| OsSTP12 | LOC_Os02g36450.1 | Os02g0574100 | STP |
| OsSTP13 | LOC_Os03g01170.1 | Os03g0101300 | STP |
| OsSTP14 | LOC_Os04g37970.1 | Os04g0452600 | STP |
| OsSTP15 | LOC_Os04g37990.1 | Os04g0453200 | STP |
| OsSTP16 | LOC_Os04g38010.1 | Os04g0453300 | STP |
| OsSTP17 | LOC_Os04g38026.1 | Os04g0453400 | STP |
| OsSTP18 | LOC_Os04g38220.1 | Os04g0454200 | STP |
| OsSTP19 | LOC_Os06g04900.1 | Os06g0141100 | STP |
| OsSTP2 | LOC_Os03g39710.1 | Os03g0594400 | STP |
| OsSTP20 | LOC_Os07g03910.1 | Os07g0131200 | STP |
| OsSTP21 | LOC_Os07g03960.1 | Os07g0131600 | STP |
| OsSTP22 | LOC_Os07g10590.1 | Os07g0206600 | STP |
| OsSTP23 | LOC_Os09g09520.1 | Os09g0268300 | STP |
| OsSTP24 | LOC_Os09g12590.1 | Os09g0297300 | STP |
| OsSTP25 | LOC_Os09g15330.1 | Os09g0322000 | STP |
| OsSTP26 | LOC_Os09g24924.1 | Os09g0416200 | STP |
| OsSTP27 | LOC_Os10g41190.1 | Os10g0561300 | STP |
| OsSTP28 | LOC_Os11g38160.1 | Os11g0594000 | STP |
| OsSTP3 | LOC_Os07g01560.1 | Os07g0106200 | STP |
| OsSTP4 | LOC_Os03g11900.1 | Os03g0218400 | STP |
| OsSTP5 | LOC_Os08g08070.1 | Os08g0178200 | STP |
| OsSTP6 | LOC_Os07g37320.1 | Os07g0559700 | STP |
| OsSTP7 | LOC_Os01g38680.1 | Os01g0567600 | STP |
| OsSTP8 | LOC_Os01g38670.1 | Os01g0567500 | STP |
| OsSTP9 | LOC_Os02g06540.1 | Os02g0160400 | STP |
| OspGlcT1 | LOC_Os01g04190.2 | Os01g0133400 | pGlcT |
| OspGlcT2 | LOC_Os02g17500.1 | Os02g0274900 | pGlcT |
| OspGlcT3 | LOC_Os09g23110.1 | Os09g0394500 | pGlcT |
| OspGlcT4 | LOC_Os09g27900.1 | Os09g0452300 | pGlcT |
| OsPLT1 | LOC_Os03g10090.1 | Os03g0197100 | PLT |
| OsPLT10 | LOC_Os11g41830.1 | Os11g0637000 | PLT |
| OsPLT11 | LOC_Os11g41840.1 | Os11g0637100 | PLT |
| OsPLT12 | LOC_Os11g41850.1 | Os11g0637200 | PLT |
| OsPLT13 | LOC_Os11g41870.1 | Os11g0637400 | PLT |
| OsPLT14 | LOC_Os12g32760.1 | Os12g0512100 | PLT |
| OsPLT15 | LOC_Os12g32940.1 | Os12g0514000 | PLT |
| OsPLT2 | LOC_Os01g73590.1 | Os01g0966900 | PLT |
| OsPLT3 | LOC_Os10g21590.1 | Os10g0360100 | PLT |
| OsPLT4 | LOC_Os04g44750.1 | Os04g0529800 | PLT |
| OsPLT5 | LOC_Os07g39350.1 | Os07g0582400 | PLT |
| OsPLT6 | LOC_Os07g39360.1 | Os07g0582500 | PLT |
| OsPLT7 | LOC_Os03g10100.1 | Os03g0197200 | PLT |
| OsPLT8 | LOC_Os04g58220.1 | Os04g0678900 | PLT |
| OsPLT9 | LOC_Os04g58230.1 | Os04g0679000 | PLT |
| OsXTPH1 | LOC_Os03g60820.1 | Os03g0823100 | XTPH |
| OsXTPH2 | LOC_Os10g42830.1 | Os10g0579200 | XTPH |
| Seq_1 | Seq_2 | Ka | Ks | Ka/Ks | Date (MYA) | Duplication Type |
|---|---|---|---|---|---|---|
| OsSTP10 | OsSTP18 | 0.3290306 | 0.5779125 | 0.5693432 | 18.078602 | Segmental duplication |
| OsERD3 | OsERD5 | 0.1951074 | 1.1448128 | 0.1704273 | 10.720185 | Segmental duplication |
| OsSTP11 | OsSTP12 | 0.0328413 | 0.1013588 | 0.3240102 | 1.8044666 | Tandem duplication |
| OsSTP8 | OsSTP7 | 0.0615979 | 0.1135565 | 0.542443 | 3.3845026 | Tandem duplication |
| OsPLT11 | OsPLT12 | 0.0777016 | 0.1552719 | 0.5004226 | 4.2693165 | Tandem duplication |
| OsERD3 | OsERD4 | 0.0837405 | 0.5136912 | 0.1630172 | 4.6011267 | Tandem duplication |
| OsPLT8 | OsPLT9 | 0.142104 | 0.3454965 | 0.4113038 | 7.807912 | Tandem duplication |
| OsPLT5 | OsPLT6 | 0.1481074 | 0.3072356 | 0.4820647 | 8.1377715 | Tandem duplication |
| OsPLT1 | OsPLT7 | 0.1914638 | 0.3353385 | 0.5709568 | 10.51999 | Tandem duplication |
| OsSTP14 | OsSTP1 | 0.1975753 | 0.3867837 | 0.5108159 | 10.855785 | Tandem duplication |
| OsERD1 | OsERD2 | 0.2101177 | 0.9200798 | 0.228369 | 11.54493 | Tandem duplication |
| OsSTP1 | OsSTP15 | 0.3065527 | 0.4261786 | 0.7193058 | 16.843556 | Tandem duplication |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, X.; An, B.; Zhong, H.; Yang, J.; Kong, W.; Li, Y. A Novel Insight into Functional Divergence of the MST Gene Family in Rice Based on Comprehensive Expression Patterns. Genes 2019, 10, 239. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10030239
Deng X, An B, Zhong H, Yang J, Kong W, Li Y. A Novel Insight into Functional Divergence of the MST Gene Family in Rice Based on Comprehensive Expression Patterns. Genes. 2019; 10(3):239. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10030239
Chicago/Turabian StyleDeng, Xiaolong, Baoguang An, Hua Zhong, Jing Yang, Weilong Kong, and Yangsheng Li. 2019. "A Novel Insight into Functional Divergence of the MST Gene Family in Rice Based on Comprehensive Expression Patterns" Genes 10, no. 3: 239. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10030239