The Role of Plasticity and Adaptation in the Incipient Speciation of a Fire Salamander Population

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
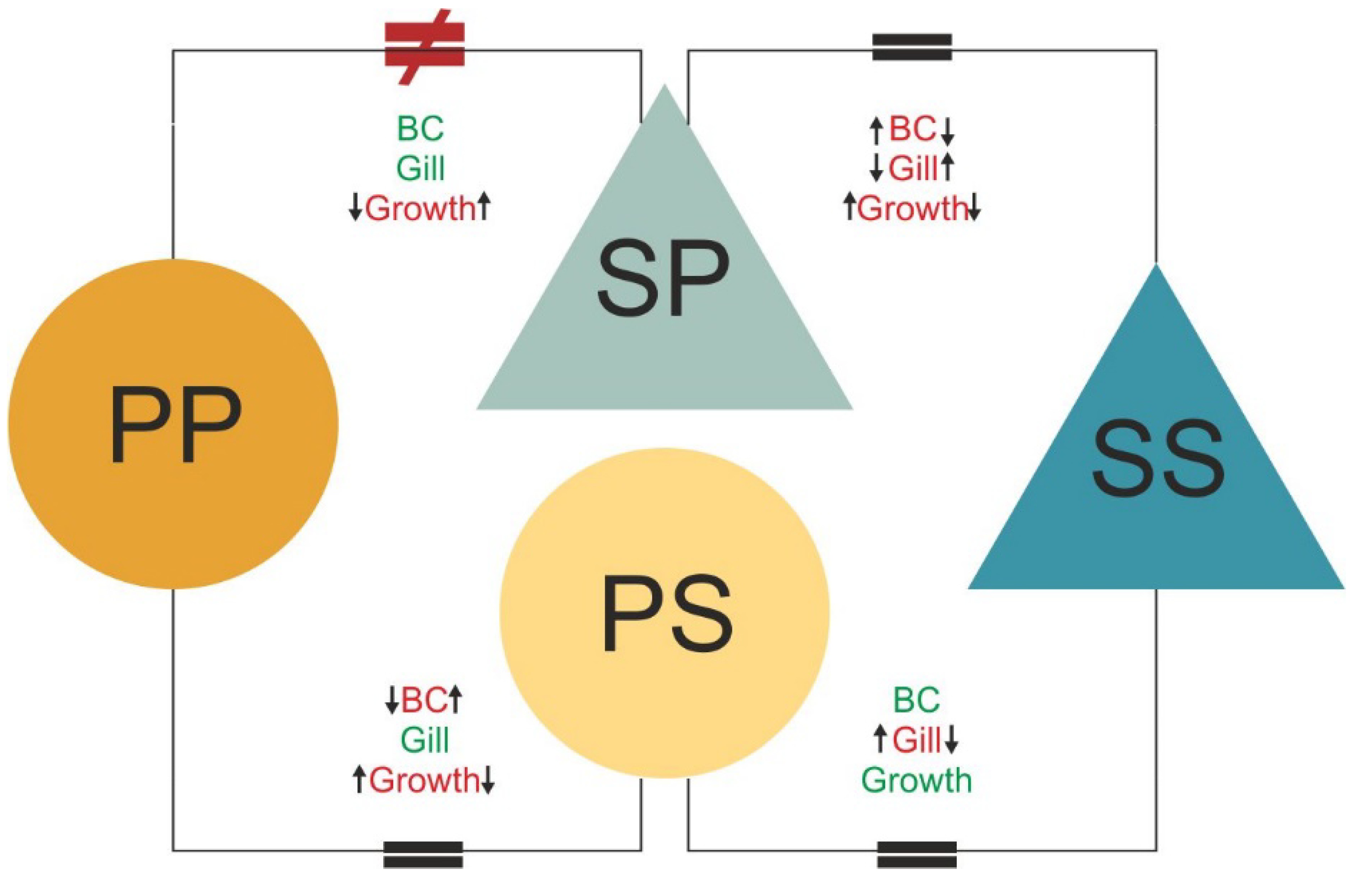
Abstract
:1. Introduction
2. Methods
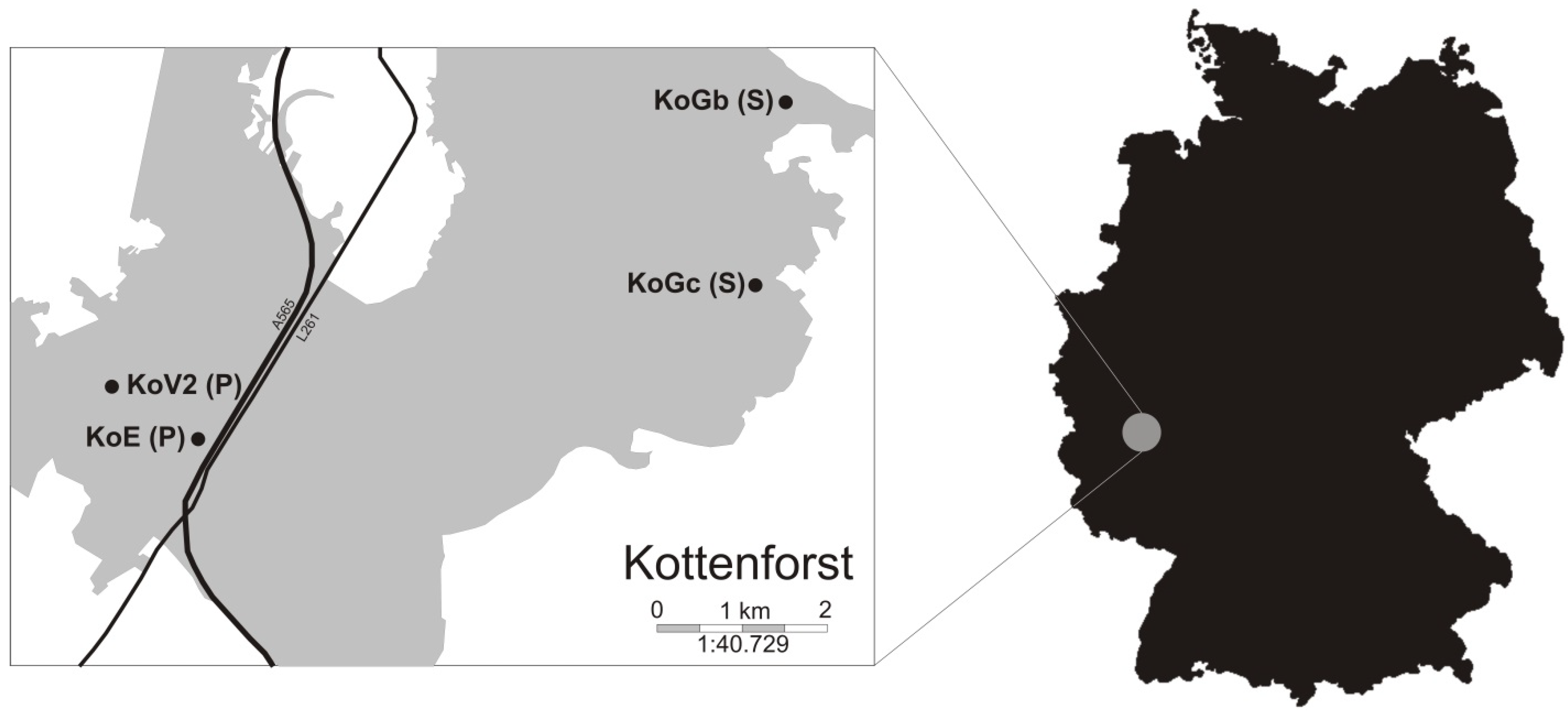
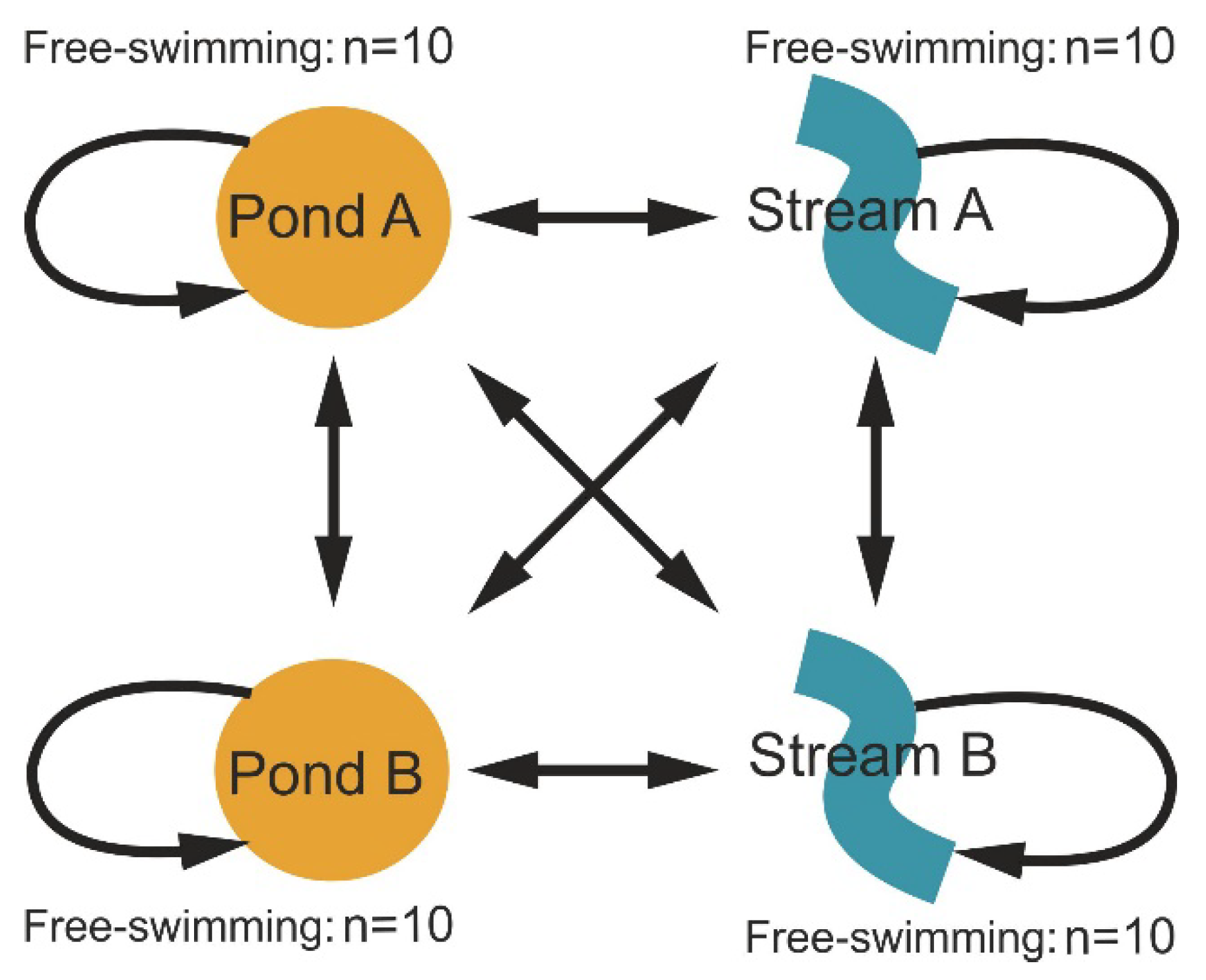
2.1. Experimental Design
2.2. Sampling and Morphology Measurements
2.3. Morphological Data Analysis
2.4. Gene Expression Analysis
3. Results
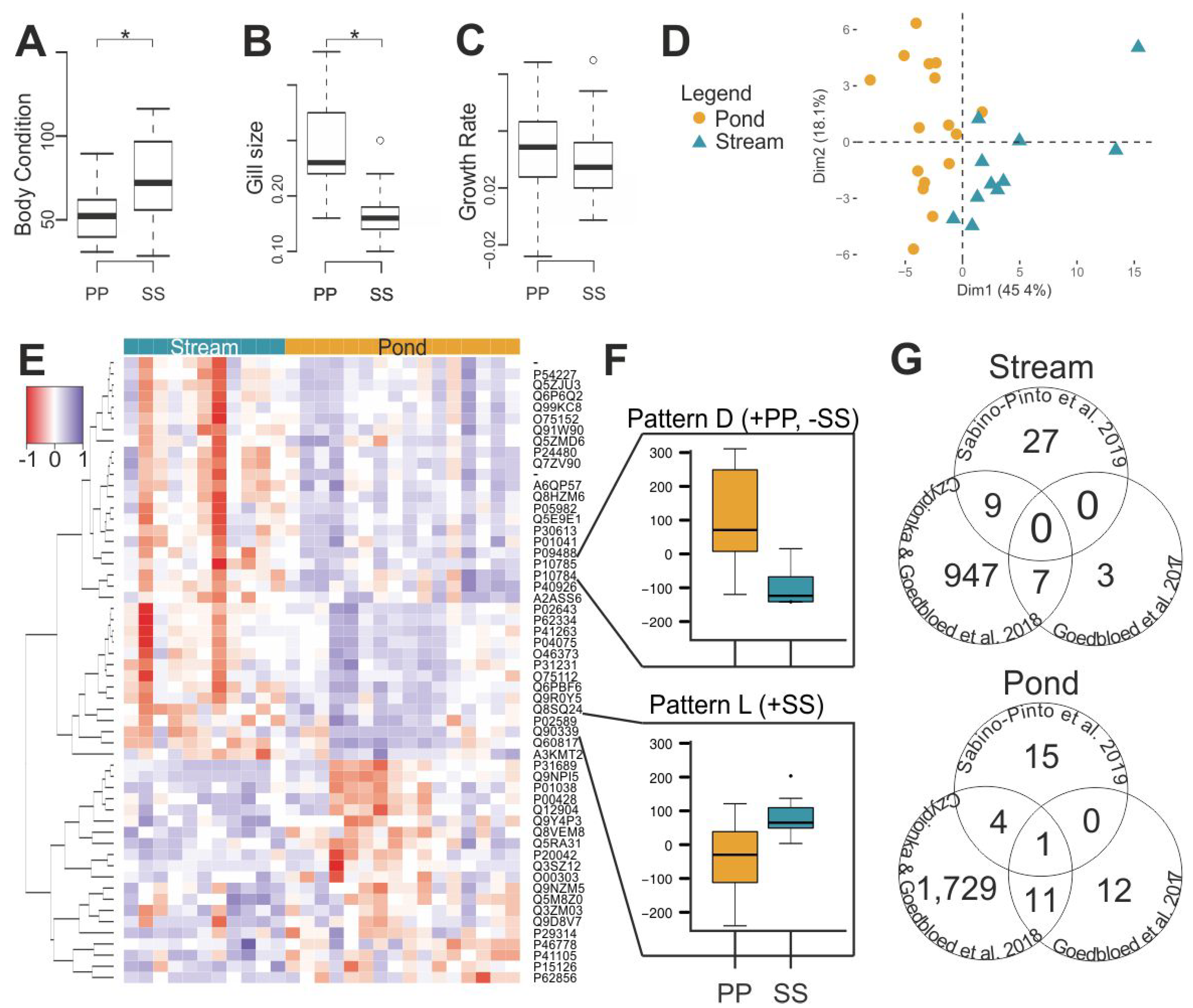
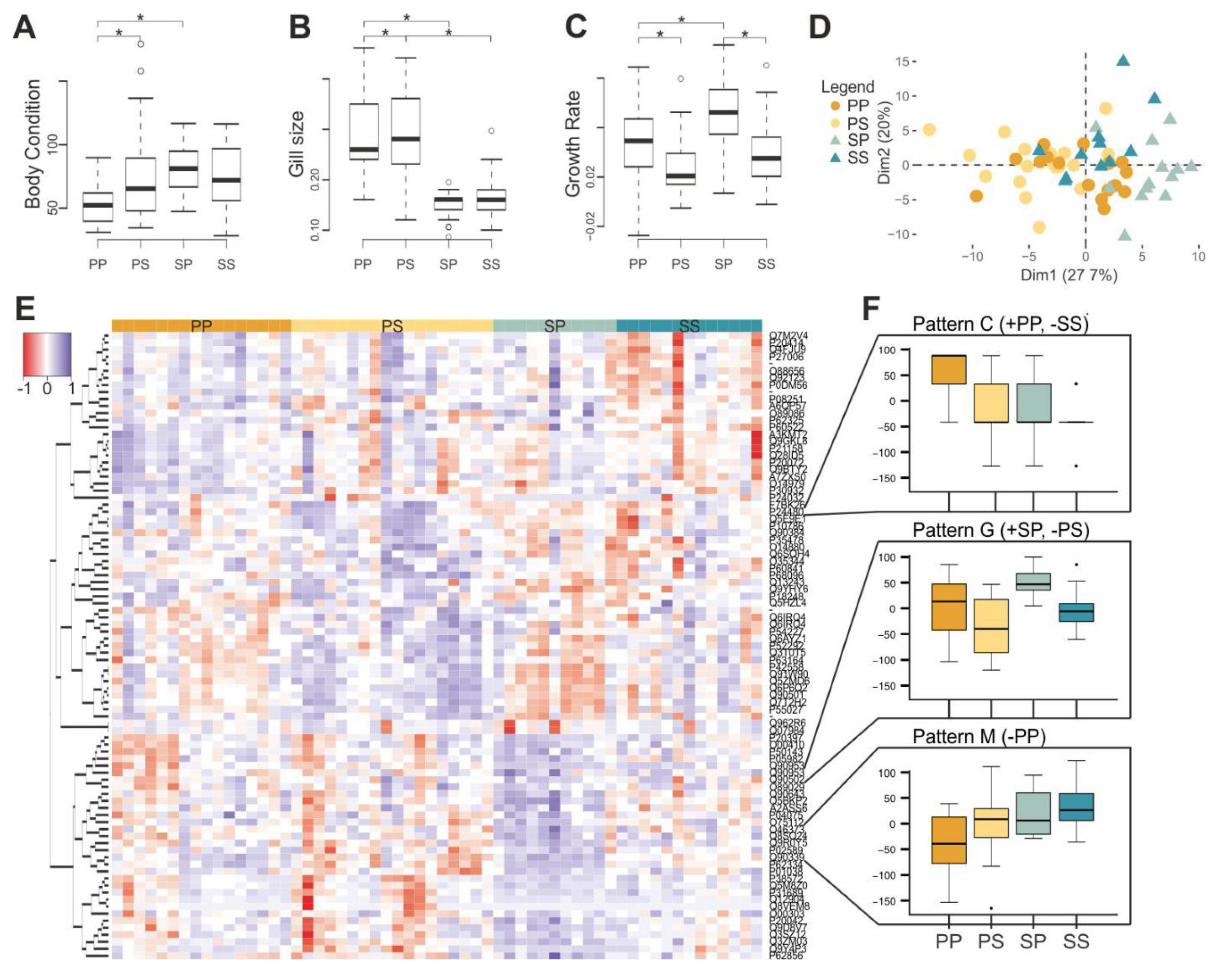
3.1. Habitat Differences
3.2. Evolutionary Divergence
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Via, S.; Gomulkiewicz, R.; De Jong, G.; Scheiner, S.M.; Schlichting, C.D.; Van Tienderen, P.H. Adaptive phenotypic plasticity: Consensus and controversy. Trends Ecol. Evol. 1995, 10, 212–217. [Google Scholar] [CrossRef]
- Pigliucci, M. Phenotypic Plasticity: Beyond Nature and Nurture; The Johns Hopkins University Press: Baltimore, MD, USA, 2001. [Google Scholar]
- West-Eberhard, M.J. Developmental Plasticity and Evolution; Oxford University Press: New York, NU, USA, 2003. [Google Scholar]
- Chevin, L.-M.; Hoffmann, A.A. Evolution of phenotypic plasticity in extreme environments. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160138. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Hansen, M.E.B.; Lo, Y.; Tishkoff, S.A. Going global by adapting local: A review of recent human adaptation. Science 2016, 354, 54–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.C. Adaptation and Natural Selection; Princeton University Press: Princeton, NJ, USA, 1966. [Google Scholar]
- Coyne, J.A.; Orr, H.A. Patterns of speciation in Drosophila. Evolution (N. Y.) 1989, 43, 362–381. [Google Scholar]
- Van Doorn, G.; Sander, G.; Edelaar, P.; Weissing, F.J. On the origin of species by natural and sexual selection. Science 2009, 326, 1704–1707. [Google Scholar] [CrossRef]
- Via, S. Reproductive isolation between sympatric races of pea aphids. I. Gene flow restriction and habitat choice. Evolution (N. Y.) 1999, 53, 1446–1457. [Google Scholar]
- Dieckmann, U.; Doebeli, M.; Metz, J.A.J.; Tautz, D. Adaptive Speciation; Cambridge University Press: Cambridge, UK, 2004; ISBN 0521828422. [Google Scholar]
- Schluter, D. Evidence for ecological speciation and its alternative. Science 2009, 323, 737–741. [Google Scholar] [CrossRef]
- Hendry, A.P. Ecological speciation! Or the lack thereof? Can. J. Fish. Aquat. Sci. 2009, 66, 1383–1398. [Google Scholar] [CrossRef]
- Nosil, P. Ecological Speciation; Oxford University Press: New York, NY, USA, 2012. [Google Scholar]
- Thibert-Plante, X.; Hendry, A.P. The consequences of phenotypic plasticity for ecological speciation. J. Evol. Biol. 2011, 24, 326–342. [Google Scholar] [CrossRef]
- Wund, M.A. Assessing the impacts of phenotypic plasticity on evolution. Integr. Comp. Biol. 2012, 52, 5–15. [Google Scholar] [CrossRef] [PubMed]
- West-Eberhard, M.J. Phenotypic Plasticity and the Origins of Diversity. Annu. Rev. Ecol. Syst. 1989, 20, 249–278. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Huey, R.B. Evolutionary analyses of morphological and physiological plasticity in thermally variable environments. Am. Zool. 1998, 38, 1–15. [Google Scholar] [CrossRef]
- Schluter, D. The Ecology of Adaptive Radiation, 2nd ed.; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Lande, R. Adaptation to an extraordinary environment by evolution of phenotypic plasticity and genetic assimilation. J. Evol. Biol. 2009, 22, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Nussey, E.D.; Postma, E.; Gienapp, P.; Visser, M.E. Evolutionary genetics of life-history traits in a structured environment. Understanding variation in clutch size and laying date in great tits (Parus major). Science 2005, 310, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Scheiner, S.M.; Gomulkiewicz, R.; Holt, R.D. The genetics of phenotypic plasticity I. Heritability. J. Evol. Biol. 1989, 2, 95–107. [Google Scholar] [CrossRef]
- Crispo, E. The Baldwin effect and genetic assimilation: Revisiting two mechanisms of evolutionary change mediated by phenotypic plasticity. Evol. Int. J. Org. Evol. 2007, 61, 2469–2479. [Google Scholar] [CrossRef]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; McGraw-Hill: New York, NY, USA, 1994. [Google Scholar]
- Newman, R.A. Ecological constraints on amphibian metamorphosis: Interactions of temperature and larval density with responses to changing food level. Oecologia 1998, 115, 9–16. [Google Scholar] [CrossRef]
- Denver, R.J. Proximate mechanisms of phenotypic plasticity in amphibian metamorphosis. Am. Zool. 1997, 37, 172–184. [Google Scholar] [CrossRef]
- Denver, R.J. Environmental stress as a developmental cue: Corticotropin-releasing hormone is a proximate mediator of adaptive phenotypic plasticity in amphibian metamorphosis. Horm. Behav. 1997, 31, 169–179. [Google Scholar] [CrossRef]
- Brockes, J.P.; Kumar, A. Plasticity and reprogramming of differentiated cells in amphibian regeneration. Nat. Rev. Mol. Cell Biol. 2002, 3, 566–574. [Google Scholar] [CrossRef] [PubMed]
- Pfennig, D.W.; Wund, M.A.; Snell-Rood, E.C.; Cruickshank, T.; Schlichting, C.D.; Moczek, A.P. Phenotypic plasticity’s impacts on diversification and speciation. Trends Ecol. Evol. 2010, 25, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Weitere, M.; Tautz, D.; Neumann, D.; Steinfartz, S. Adaptive divergence vs. environmental plasticity: Tracing local genetic adaptation of metamorphosis traits in salamanders. Mol. Ecol. 2004, 13, 1665–1677. [Google Scholar] [CrossRef] [PubMed]
- Denver, R.J.; Mirhadi, N.; Phillips, M. Adaptive plasticity in amphibian metamorphosis: Response of Scaphiopus hammondii tadpoles to habitat desiccation. Ecology 1998, 79, 1859–1872. [Google Scholar] [CrossRef]
- Levis, N.A.; Serrato-Capuchina, A.; Pfennig, D.W. Genetic accommodation in the wild: Evolution of gene expression plasticity during character displacement. J. Evol. Biol. 2017, 30, 1712–1723. [Google Scholar] [CrossRef]
- Alcobendas, M.; Castanet, J. Bone growth plasticity among populations of Salamandra salamandra: Interactions between internal and external factors. Herpetologica 2000, 56, 14–26. [Google Scholar]
- Alcobendas, M.; Buckley, D.; Tejedo, M. Variability in survival, growth and metamorphosis in the larval Fire Salamander (Salamandra Salamandra): Effects of larval birth size, sibship and environment. Herpetologica 2004, 60, 232–245. [Google Scholar] [CrossRef]
- Manenti, R.; Denoël, M.; Ficetola, G.F. Foraging plasticity favours adaptation to new habitats in fire salamanders. Anim. Behav. 2013, 86, 375–382. [Google Scholar] [CrossRef]
- Manenti, R.; Ficetola, G.F.; Bianchi, B.; de Bernardi, F. Habitat features and distribution of Salamandra salamandra in underground springs. Acta Herpetol. 2009, 4, 143–151. [Google Scholar]
- Seifert, D. Untersuchungen an einer ostthüringischen Population des Feuersalamanders (Salamandra salamandra). Artenschutzreport 1991, 1, 1–16. [Google Scholar]
- Reinhardt, T.; Steinfartz, S.; Paetzold, A.; Weitere, M. Linking the evolution of habitat choice to ecosystem functioning: Direct and indirect effects of pond-reproducing fire salamanders on aquatic-terrestrial subsidies. Oecologia 2013, 173, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.; Whitfield, M.; Biggs, J.; Bray, S.; Fox, G.; Nicolet, P.; Sear, D. Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biol. Conserv. 2004, 115, 329–341. [Google Scholar] [CrossRef]
- Sneddon, L.U.; Yerbury, J. Differences in response to hypoxia in the three-spined stickleback from lotic and lentic localities: Dominance and an anaerobic metabolite. J. Fish Biol. 2004, 64, 799–804. [Google Scholar] [CrossRef]
- Steinfartz, S.; Weitere, M.; Tautz, D. Tracing the first step to speciation: Ecological and genetic differentiation of a salamander population in a small forest. Mol. Ecol. 2007, 16, 4550–4561. [Google Scholar] [CrossRef]
- Goedbloed, D.J.; Czypionka, T.; Altmüller, J.; Rodrıguez, A.; Küpfer, E.; Segev, O.; Blaustein, L.; Templeton, A.R.; Nolte, A.W.; Steinfartz, S. Parallel habitat acclimatization is realized through different gene expression mechanisms in two closely related salamander species (genus Salamandra). Heredity (Edinb.) 2017, 119, 429–437. [Google Scholar] [CrossRef]
- Czypionka, T.; Goedbloed, D.J.; Steinfartz, S.; Nolte, A.W. Plasticity and evolutionary divergence in gene expression associated with alternative habitat use in larvae of the European Fire Salamander. Mol. Ecol. 2018, 27, 2698–2713. [Google Scholar] [CrossRef]
- Klappert, K.; Reinhold, K. Local adaptation and sexual selection: A reciprocal transfer experiment with the grasshopper Chorthippus biguttulus. Behav. Ecol. Sociobiol. 2005, 58, 36–43. [Google Scholar] [CrossRef]
- Hairston, N.G.; Olds, E.J. Population differences in the timing of diapause: Adaptation in a spatially heterogeneous environment. Oecologia 1984, 61, 42–48. [Google Scholar] [CrossRef]
- Bush, S.E.; Clayton, D.H. The role of body size in host specificity: Reciprocal transfer experiments with feather lice. Evolution (N. Y.) 2006, 60, 2158–2167. [Google Scholar]
- Bletz, M.C.; Goedbloed, D.J.; Sanchez, E.; Reinhardt, T.; Tebbe, C.C.; Bhuju, S.; Geffers, R.; Jarek, M.; Vences, M.; Steinfartz, S. Amphibian gut microbiota shifts differentially in community structure but converges on habitat-specific predicted functions. Nat. Commun. 2016, 7, 13699. [Google Scholar] [CrossRef]
- Sanchez, E.; Küpfer, E.; Goedbloed, D.J.; Nolte, A.W.; Lüddecke, T.; Schulz, S.; Vences, M.; Steinfartz, S. Morphological and transcriptomic analyses reveal three discrete primary stages of postembryonic development in the common fire salamander, Salamandra salamandra. J. Exp. Zool. Part B Mol. Dev. Evol. 2018, 330, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Segev, O.; Polevikove, A.; Blank, L.; Goedbloed, D.; Küpfer, E.; Gershberg, A.; Koplovich, A.; Blaustein, L. Effects of tail clipping on larval performance and tail regeneration rates in the near eastern fire salamander, Salamandra infraimmaculata. PLoS ONE 2015, 10, 1–12. [Google Scholar]
- Czypionka, T.; Krugman, T.; Altmüller, J.; Blaustein, L.; Steinfartz, S.; Templeton, A.R.; Nolte, A.W. Ecological transcriptomics—A non-lethal sampling approach for endangered fire salamanders. Methods Ecol. Evol. 2015, 6, 1417–1425. [Google Scholar] [CrossRef]
- Team, R.C. R: A language and environment for statistical computing 2017.
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biometrical J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Harrell, F.E., Jr.; Dupont, M.C. The Hmisc Package. R Package. Version 4.2-0. 2006. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 15 October 2019).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A. Practical Guide To Principal Component Methods in R: PCA, M (CA), FAMD, MFA, HCPC, Factoextra; STHDA. 2017. Available online: http://www.sthda.com/ (accessed on 15 October 2019).
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 October 2019).
- Segata, N.; Izard, J.; Waldron, L. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Löffler-Wirth, H.; Kalcher, M.; Binder, H. OposSOM: R-package for high-dimensional portraying of genome-wide expression landscapes on bioconductor. Bioinformatics 2015, 31, 3225–3227. [Google Scholar] [CrossRef]
- Alexa, A.; Rahnenfuhrer, J. topGO: Enrichment Analysis for Gene Ontology. 2019. Available online: https://bioconductor.org/packages/release/bioc/html/topGO.html (accessed on 15 October 2019).
- Pörtner, H.O.; Knust, R. Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 2007, 315, 95–97. [Google Scholar] [CrossRef]
- Pörtner, H.O. Oxygen-and capacity-limitation of thermal tolerance: A matrix for integrating climate-related stressor effects in marine ecosystems. J. Exp. Biol. 2010, 213, 881–893. [Google Scholar] [CrossRef]
- Yamauchi, M.; Woodley, D.T.; Mechanic, G.L. Aging and cross-linking of skin collagen. Biochem. Biophys. Res. Commun. 1988, 152, 898–903. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A. Metamorphosis and the amphibian immune system. Immunol. Rev. 1998, 166, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Levenson, S.M.; Gruber, C.A.; Rettura, G.; Gruber, D.K.; Demetriou, A.A.; Seifter, E. Supplemental vitamin A prevents the acute radiation-induced defect in wound healing. Ann. Surg. 1984, 200, 494–512. [Google Scholar] [CrossRef] [PubMed]
- Liselle, D.; Schteingart, D. Effect of obesity and starvation on thyroid hormone, growth hormone, and cortisol secretion. Endocrinol. Metab. Clin. N. Am. 2002, 31, 173–189. [Google Scholar]
- Robinson, A.M.; Girard, J.R.; Williamson, D.H. Evidence for a role of insulin in the regulation of lipogenesis in lactating rat mammary gland. Measurements of lipogenesis in vivo and plasma hormone concentrations in response to starvation and refeeding. Biochem. J. 1978, 176, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Potthoff, M.J.; Inagaki, T.; Satapati, S.; Ding, X.; He, T.; Goetz, R.; Mohammadi, M.; Finck, B.N.; Mangelsdorf, D.J.; Kliewer, S.A.; et al. FGF21 induces PGC-1 and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response. Proc. Natl. Acad. Sci. USA 2009, 106, 10853–10858. [Google Scholar] [CrossRef]
- Crump, M.L. Effect of habitat drying on developmental time and size at metamorphosis in Hyla pseudopuma. Copeia 1989, 1989, 794–797. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Wilbur, H.M. Effects of pond drying time on metamorphosis and survival in the salamander Ambystoma talpoideum. Copeia 1988, 1988, 978. [Google Scholar] [CrossRef]
- Caspers, B.A.; Junge, C.; Weitere, M.; Steinfartz, S. Habitat adaptation rather than genetic distance correlates with female preference in fire salamanders (Salamandra salamandra). Front. Zool. 2009, 6, 13. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabino-Pinto, J.; Goedbloed, D.J.; Sanchez, E.; Czypionka, T.; Nolte, A.W.; Steinfartz, S. The Role of Plasticity and Adaptation in the Incipient Speciation of a Fire Salamander Population. Genes 2019, 10, 875. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110875
Sabino-Pinto J, Goedbloed DJ, Sanchez E, Czypionka T, Nolte AW, Steinfartz S. The Role of Plasticity and Adaptation in the Incipient Speciation of a Fire Salamander Population. Genes. 2019; 10(11):875. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110875
Chicago/Turabian StyleSabino-Pinto, Joana, Daniel J. Goedbloed, Eugenia Sanchez, Till Czypionka, Arne W. Nolte, and Sebastian Steinfartz. 2019. "The Role of Plasticity and Adaptation in the Incipient Speciation of a Fire Salamander Population" Genes 10, no. 11: 875. https://0-doi-org.brum.beds.ac.uk/10.3390/genes10110875