Genome-Wide Identification, Expression Profile of the TIFY Gene Family in Brassica oleracea var. capitata, and Their Divergent Response to Various Pathogen Infections and Phytohormone Treatments


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of the B. oleracea TIFY Family Genes
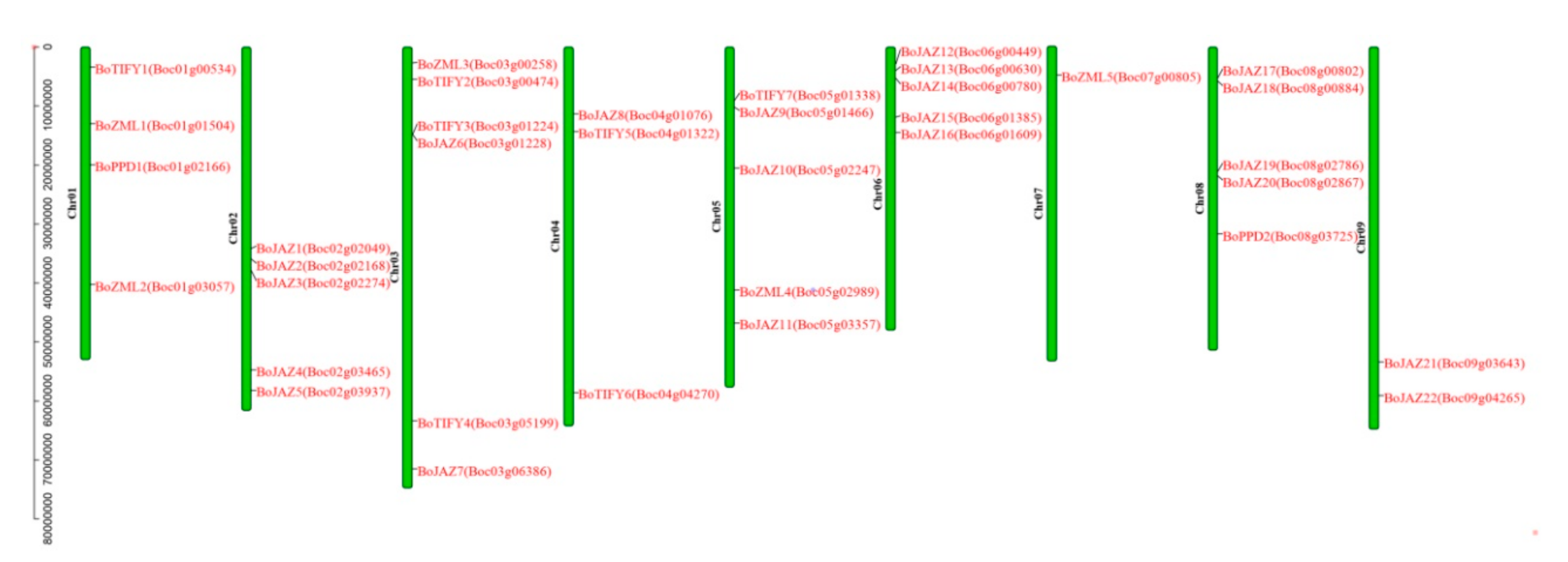
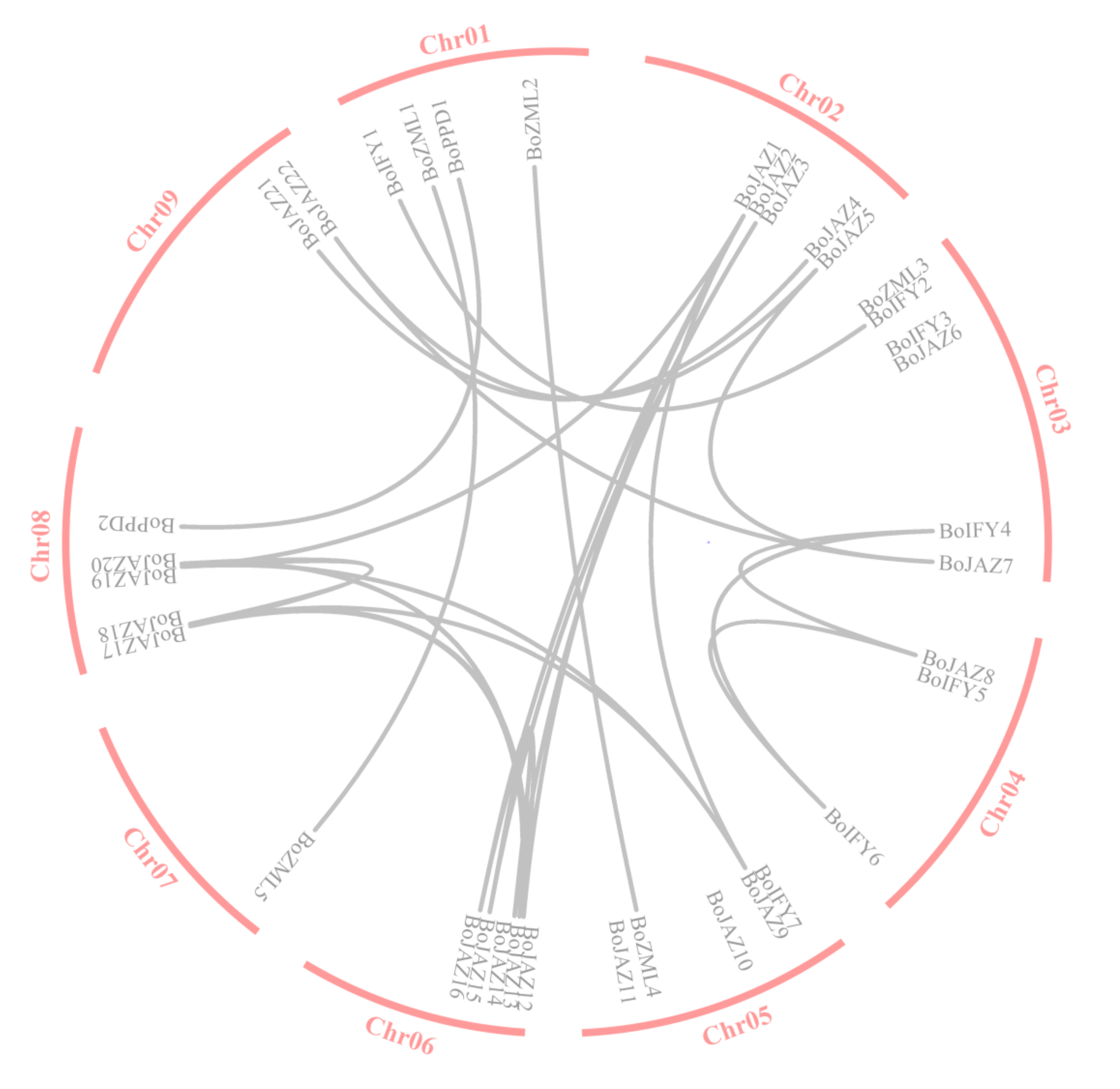
2.2. Chromosomal Location and Tandem Duplication Analysis
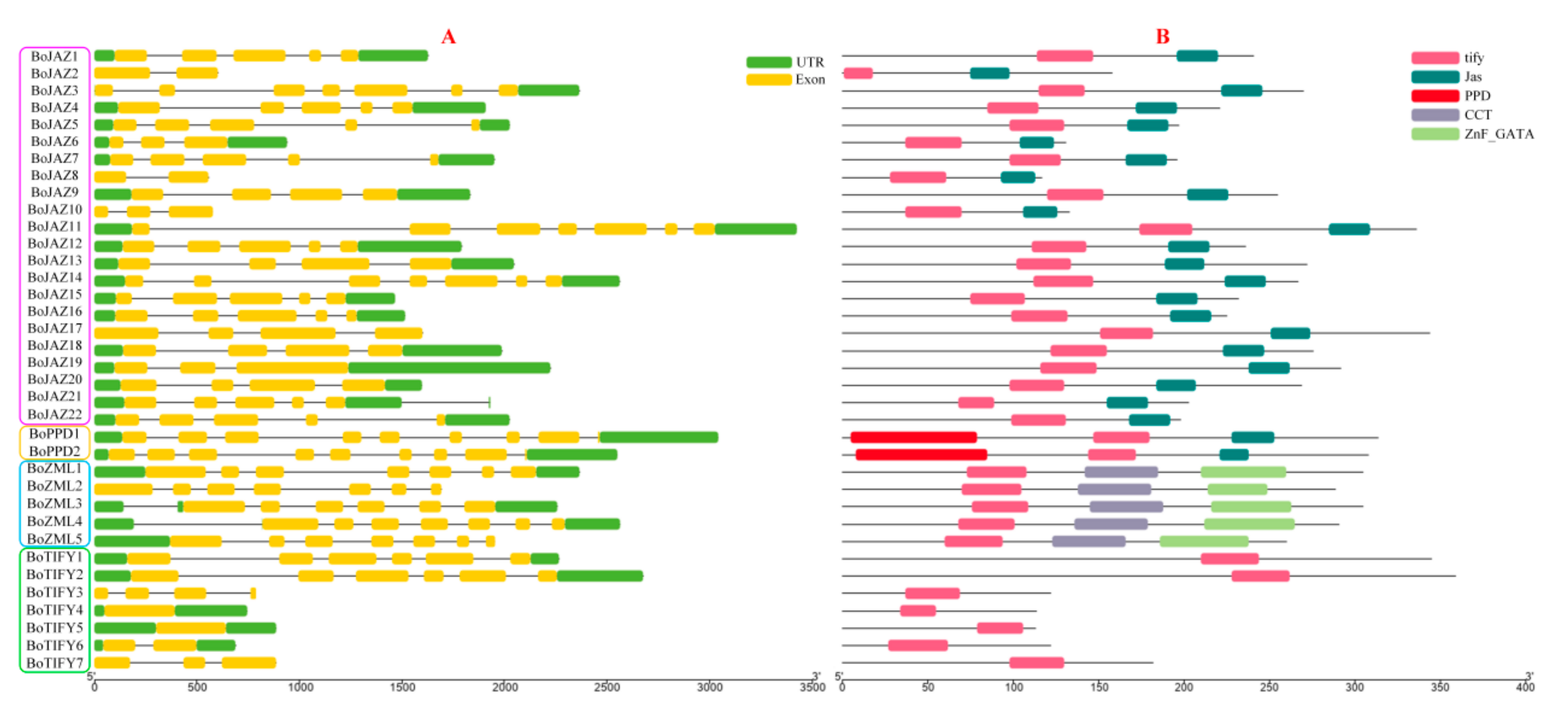
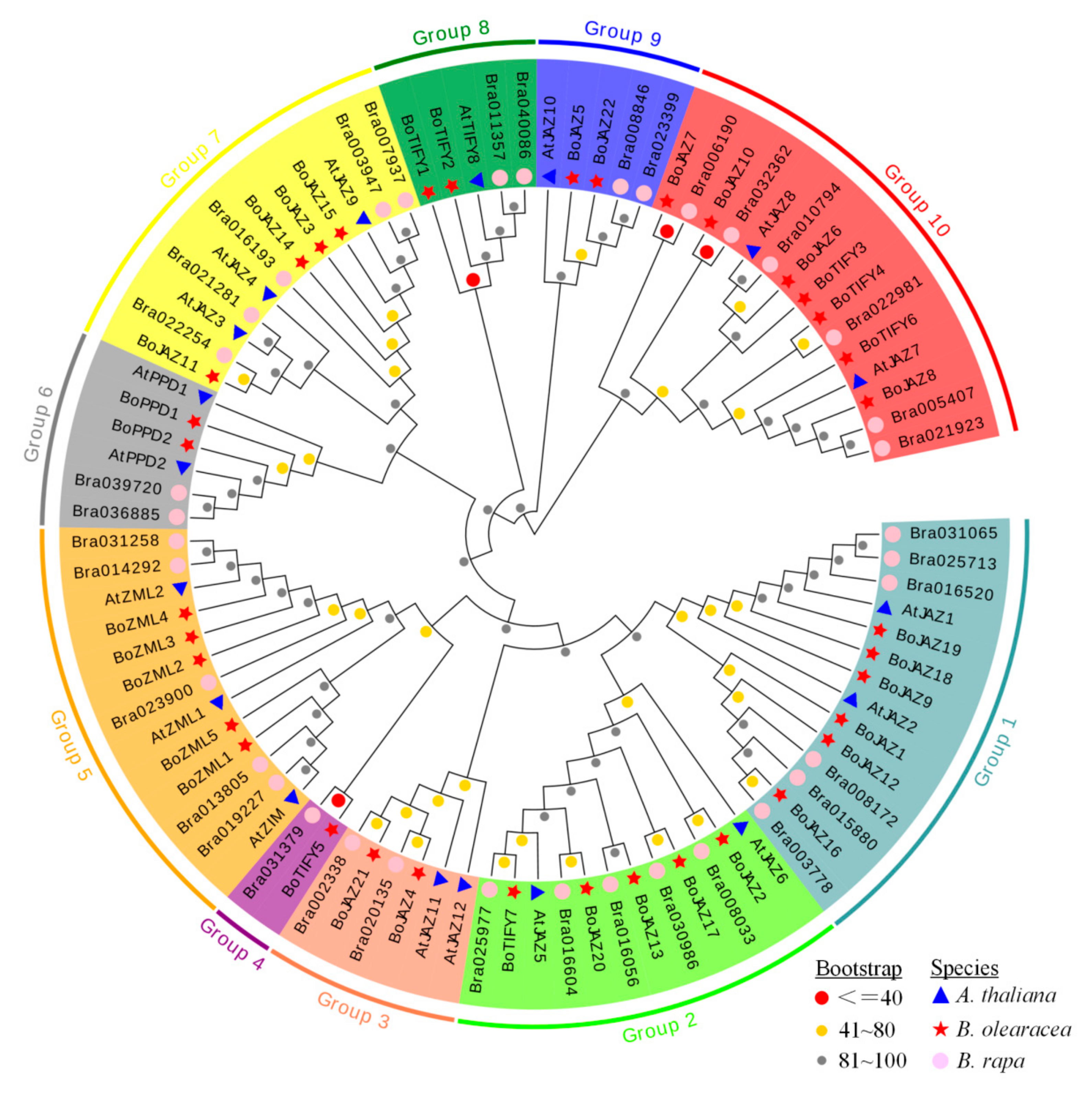
2.3. Gene Structure, Conserved Motif, and Phylogenetic Analyses
2.4. Expression Pattern Analysis of Cabbage TIFY Genes Using RNA-Seq Data
2.5. Phytohormone Treatment and Quantitative Real-Time PCR Analysis
2.6. Subcellular Localization
3. Results
3.1. Genome-Wide Identification of the TIFY Family Genes in Cabbage
3.2. Chromosomal Location and Gene Duplication Analysis of the Cabbage TIFY Genes
3.3. Gene Structure, Conserved Motif, and Phylogenetic Analysis of the Cabbage TIFY Genes
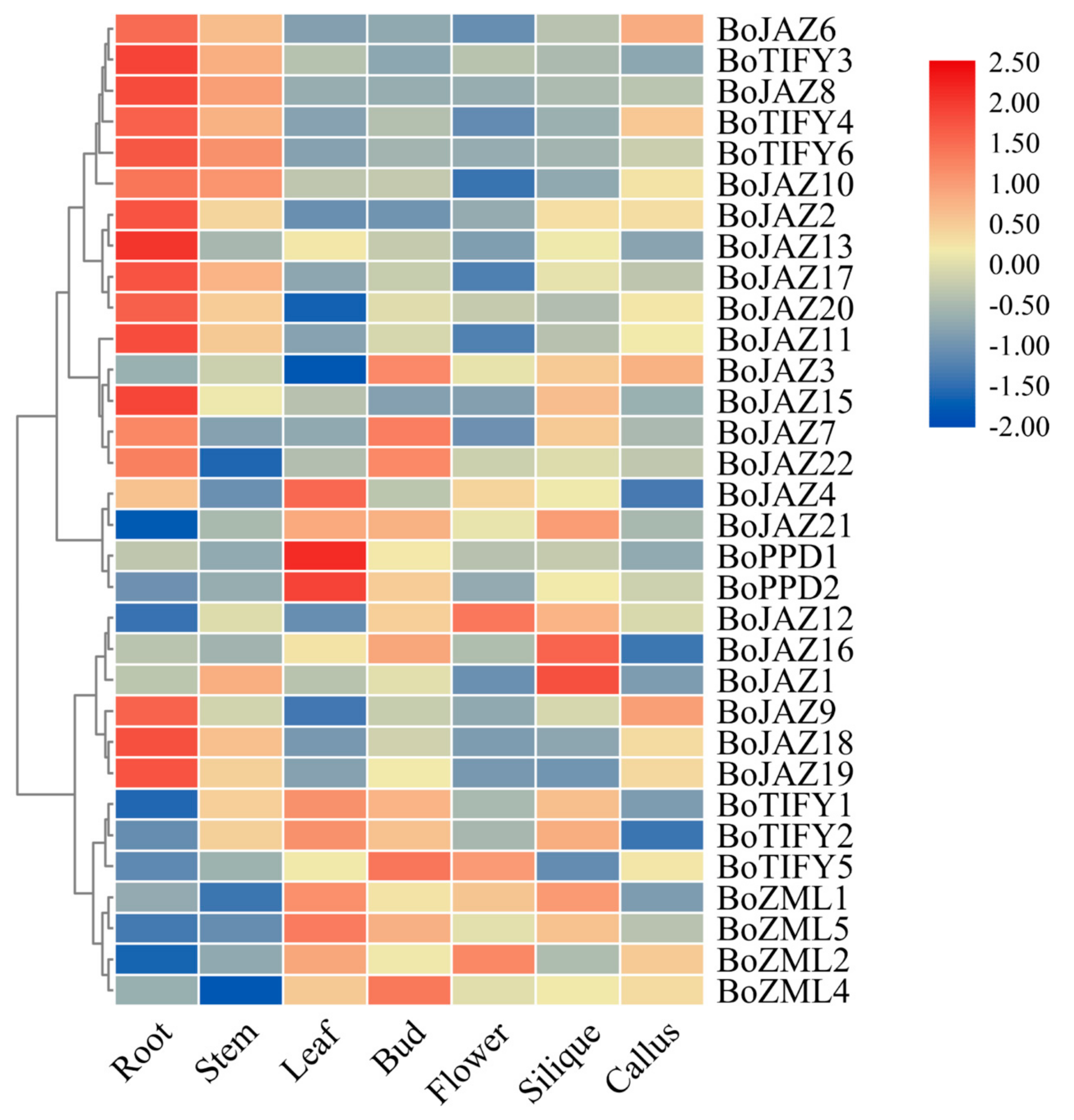
3.4. Expression Patterns of the Cabbage TIFY Family Genes in Various Tissues
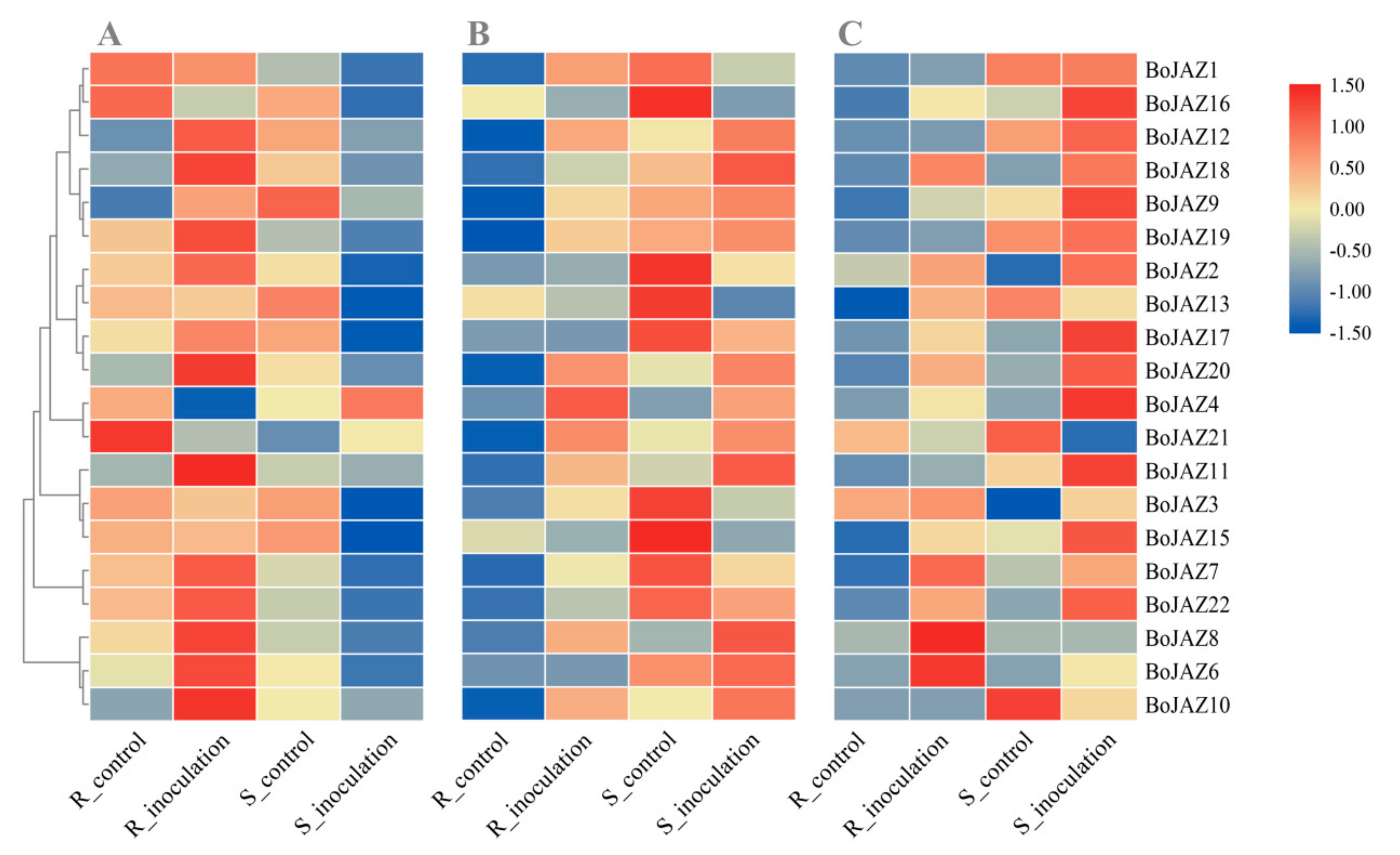
3.5. Expression Profiles of the Cabbage JAZ Genes induced by Different Pathogen Infection
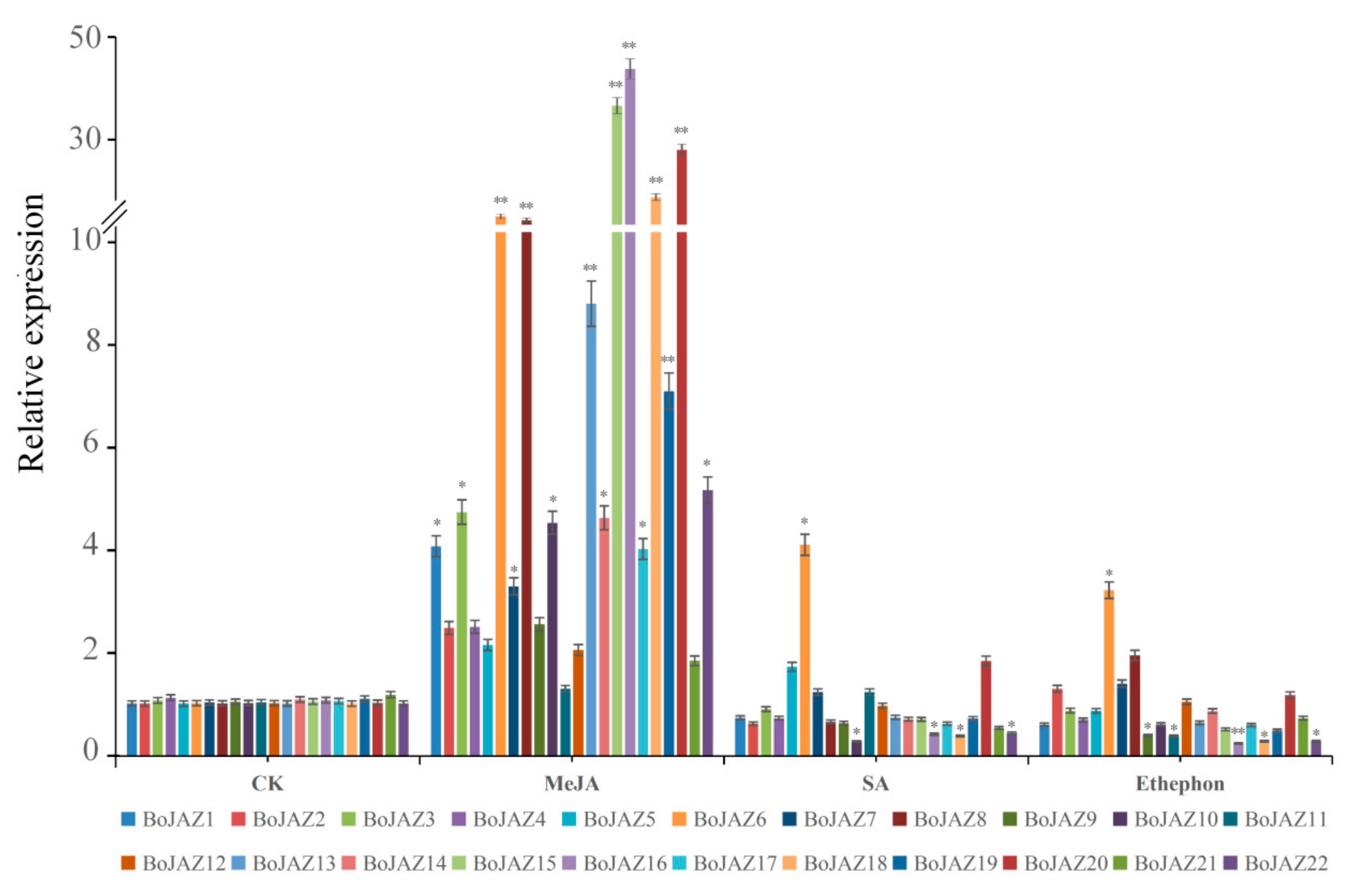
3.6. Expression Patterns of the Cabbage JAZ Genes after Exogenous Phytohormone Treatment
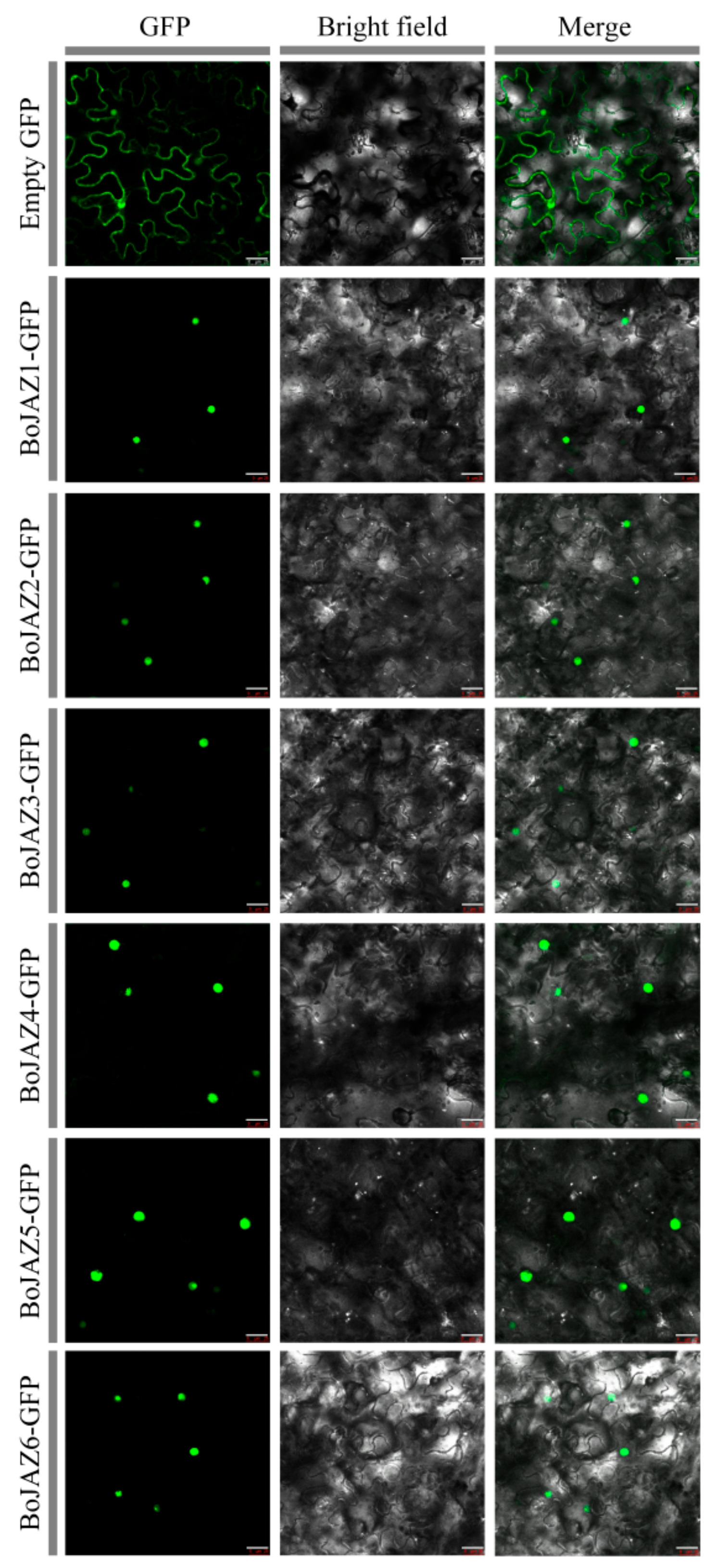
3.7. Subcellular Localization of Cabbage JAZ Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Vanholme, B.; Grunewald, W.; Bateman, A.; Kohchi, T.; Gheysen, G. The tify family previously known as ZIM. Trends Plant Sci. 2007, 12, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Nishii, A.; Takemura, M.; Fujita, H.; Shikata, M.; Yokota, A.; Kohchi, T. Characterization of a novel gene encoding a putative single zinc-finger protein, ZIM, expressed during the reproductive phase in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2000, 64, 1402–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, J.C.; Muro-Pastor, M.I.; Florencio, F.J. The GATA family of transcription factors in Arabidopsis and rice. Plant Physiol. 2004, 134, 1718–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikata, M.; Matsuda, Y.; Ando, K.; Nishii, A.; Takemura, M.; Yokota, A.; Kohchi, T. Characterization of Arabidopsis ZIM, a member of a novel plant-specific GATA factor gene family. J. Exp. Bot. 2004, 55, 631–639. [Google Scholar] [CrossRef]
- Bateman, A. The Pfam protein families database. Nucleic Acids Res. 2002, 30, 276–280. [Google Scholar] [CrossRef] [Green Version]
- Mulder, N.J.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Binns, D.; Bradley, P.; Bork, P.; Bucher, P.; Cerutti, L.; et al. InterPro, progress and status in 2005. Nucleic Acids Res. 2005, 33, D201–D205. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.S.; Howe, G.A. A critical role for the TIFY motif in repression of jasmonate signaling by a stabilized splice variant of the JASMONATE ZIM-domain protein JAZ10 in Arabidopsis. Plant Cell 2009, 21, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Meng, Y.; Huang, D.; Qi, Y.; Chen, M. Origin and evolutionary analysis of the plant-specific TIFY transcription factor family. Genomics 2011, 98, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Ebel, C.; BenFeki, A.; Hanin, M.; Solano, R.; Chini, A. Characterization of wheat (Triticum aestivum) TIFY family and role of Triticum durum TdTIFY11a in salt stress tolerance. PLoS ONE 2018, 13, e0200566. [Google Scholar] [CrossRef]
- Sirhindi, G.; Sharma, P.; Arya, P.; Goel, P.; Kumar, G.; Acharya, V.; Singh, A.K. Genome-wide characterization and expression profiling of TIFY gene family in pigeonpea (Cajanus cajan (L.) Millsp.) under copper stress. J. Plant Biochem. Biotechnol. 2016, 25, 301–310. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, F.; Chen, D.; Chu, W.; Liu, H.; Xiang, Y. Genome-wide identification and analysis of the Populus trichocarpa TIFY gene family. Plant Physiol. Biochem. 2017, 115, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Yin, X.J.; Wang, H.; Li, J.; Guo, C.L.; Gao, H.; Zheng, Y.; Fan, C.H.; Wang, X.P. Genome-wide identification and analysis of the apple (Malus × domestica Borkh.) TIFY gene family. Tree Genet. Genomes 2015, 11, 808. [Google Scholar] [CrossRef]
- Velez-Bermudez, I.C.; Salazar-Henao, J.E.; Fornalé, S.; López-Vidriero, I.; Franco-Zorrilla, J.M.; Grotewold, E.; Gray, J.; Solano, R.; Schmidt, W.; Pagés, M.; et al. A MYB/ZML complex regulates wound-induced lignin genes in Maize. Plant Cell 2015, 27, 3245–3259. [Google Scholar] [CrossRef] [Green Version]
- White, D.W. PEAPOD regulates lamina size and curvature in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 13238–13243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wager, A.; Browse, J. Social Network: JAZ protein interactions expand our knowledge of jasmonate signaling. Front. Plant Sci. 2012, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Manners, J.M. JAZ repressors and the orchestration of phytohormone crosstalk. Trends Plant Sci. 2012, 17, 22–31. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, C.; Gu, M.; Bai, Z.; Zhang, W.; Qi, T.; Cheng, Z.; Peng, W.; Luo, H.; Nan, F.; et al. The Arabidopsis CORONATINE INSENSITIVE1 protein is a jasmonate receptor. Plant Cell 2009, 21, 2220–2236. [Google Scholar] [CrossRef] [Green Version]
- Katsir, L.; Schilmiller, A.L.; Staswick, P.E.; He, S.Y.; Howe, G.A. COI1 is a critical component of a receptor for jasmonate and the bacterial virulence factor coronatine. Proc. Natl. Acad. Sci. USA 2008, 105, 7100–7105. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, O.; Chico, J.M.; Sanchez-Serrano, J.J.; Solano, R. JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef] [Green Version]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Fernández, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; García-Casado, G.; López-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Melotto, M.; Mecey, C.; Niu, Y.; Chung, H.S.; Katsir, L.; Yao, J.; Zeng, W.; Thines, B.; Staswick, P.; Browse, J.; et al. A critical role of two positively charged amino acids in the Jas motif of Arabidopsis JAZ proteins in mediating coronatine- and jasmonoyl isoleucine-dependent interactions with the COI1 F-box protein. Plant J. 2008, 55, 979–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Calvo, P.; Chini, A.; Fernández-Barbero, G.; Chico, J.M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M.; et al. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, S.; Qi, T.; Huang, H.; Ren, Q.; Wu, D.; Chang, C.; Peng, W.; Liu, Y.; Peng, J.; Xie, D. The Jasmonate-ZIM domain proteins interact with the R2R3-MYB transcription factors MYB21 and MYB24 to affect Jasmonate-regulated stamen development in Arabidopsis. Plant Cell 2011, 23, 1000–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, G.; Park, J.I.; Kayum, M.A.; Nou, I.S. A genome-wide analysis reveals stress and hormone responsive patterns of TIFY family genes in Brassica rapa. Front. Plant Sci. 2016, 7, 936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, Y.; Wang, Y.; Fang, Z.; Zhuang, M.; Zhang, Y.; Lv, H.; Liu, Y.; Li, Z.; Yang, L. Comparative transcriptome analysis of cabbage (Brassica oleracea var. capitata) infected by Plasmodiophora brassicae reveals drastic defense response at secondary infection stage. Plant Soil 2019, 443, 167–183. [Google Scholar]
- Liu, X.; Xing, M.; Kong, C.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Y.; Ling, J.; Yang, Y.; Lv, H. Genetic diversity, virulence, race profiling, and comparative genomic analysis of the Fusarium oxysporum f. sp. conglutinans strains infecting cabbages in China. Front. Microbiol. 2019, 10, 1373. [Google Scholar]
- Iglesias-Bernabé, L.; Madloo, P.; Rodríguez, V.M.; Francisco, M.; Soengas, P. Dissecting quantitative resistance to Xanthomonas campestris pv. campestris in leaves of Brassica oleracea by QTL analysis. Sci. Rep. 2019, 9, 2015. [Google Scholar]
- Carlsson, M.; Von Bothmer, R.; Merker, A. Screening and evaluation of resistance to downy mildew (Peronospora parasitica) and clubroot (Plasmodiophora brassicae) in genetic resources of Brassica oleracea. Hereditas 2013, 141, 293–300. [Google Scholar] [CrossRef]
- Zhang, L.; Zheng, L.; Hsiang, T.; Lv, R.; Huang, J. An outbreak of head rot of cabbage caused by Rhizoctonia solani AG2-1 in central China. Plant Dis. 2009, 93, 109. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.F.; Wei, J.H. First report of bacterial soft rot of konnyaku caused by Dickeya dadantii in China. Plant Dis. 2014, 98, 682. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Lv, H.; Ma, J.; Xu, D.; Li, H.; Yang, L.; Kang, J.; Wang, X.; Fang, Z. Transcriptome profiling of resistance to Fusarium oxysporum f. sp. conglutinans in cabbage (Brassica oleracea) roots. PLoS ONE 2016, 11, e0148048. [Google Scholar]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 23, 3930. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Izzah, N.K.; Choi, B.S.; Joh, H.J.; Lee, S.C.; Perumal, S.; Seo, J.; Ahn, K.; Jo, E.J.; Choi, G.J.; et al. Genotyping-by-sequencing map permits identification of clubroot resistance QTLs and revision of the reference genome assembly in cabbage (Brassica oleracea L.). DNA Res. 2015, 23, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Gao, B.; Han, F.; Fang, Z.; Yang, L.; Zhuang, M.; Lv, H.; Liu, Y.; Li, Z.; Cai, C.; et al. Genetics and fine mapping of a purple leaf gene, BoPr, in ornamental kale (Brassica oleracea L. var. acephala). BMC Genom. 2017, 18, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Yu, H.; Han, F.; Li, Z.; Fang, Z.; Yang, L.; Zhuang, M.; Lv, H.; Liu, Y.; Li, Z.; et al. Differentially expressed genes associated with the cabbage yellow-green-leaf mutant in the ygl-1 mapping interval with recombination suppression. Int. J. Mol. Sci. 2018, 19, 2936. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kong, C.; Yu, H.; Liu, X.; Fang, Z.; Liu, Y.; Yang, L.; Zhuang, M.; Wang, Y.; Lv, H.; et al. Identification of a major QTL for seed number per silique in cabbage (Brassica oleracea L. var. capitata) using genotyping by sequencing. Euphytica 2019, 215, 133. [Google Scholar]
- Chen, C.; Xia, R.; Chen, H.; He, Y. TBtools, a toolkit for biologists integrating various HTS-data handling tools with a user-friendly interface. BioRxiv 2018, 289660. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rieu, I.; Powers, S.J. Real-time quantitative RT-PCR: Design, calculations, and statistics. Plant Cell 2009, 21, 1031–1033. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O.; Lederer, C.; Baulcombe, D.C. A viral movement protein prevents systemic spread of the gene silencing signal in Nicotiana benthamiana. Cell 2000, 103, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Lysak, M.A.; Cheung, K.; Kitschke, M.; Bures, P. Ancestral chromosomal blocks are triplicated in Brassiceae species with varying chromosome number and genome size. Plant Physiol. 2007, 145, 402–410. [Google Scholar] [CrossRef]
- Thatcher, L.F.; Cevik, V.; Grant, M.; Zhai, B.; Jones, J.D.; Manners, J.M.; Kazan, K. Characterization of a JAZ7 activation-tagged Arabidopsis mutant with increased susceptibility to the fungal pathogen Fusarium oxysporum. J. Exp. Bot. 2016, 67, 2367–2386. [Google Scholar] [CrossRef] [Green Version]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D. Hormone crosstalk in plant disease and defense: More than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate action in plant growth and development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murat, F.; Louis, A.; Maumus, F.; Armero, A.; Cooke, R.; Quesneville, H.; Crollius, H.R.; Salse, J. Erratum to: Understanding Brassicaceae evolution through ancestral genome reconstruction. Genome Biol. 2016, 17, 64. [Google Scholar] [CrossRef] [Green Version]
- Warwick, S.I.; Mummenhoff, F.; Sauder, C.A.; Koch, M.A.; Al-Shehbaz, I.A. Closing the gaps: Phylogenetic relationships in the Brassicaceae based on DNA sequence data of nuclear ribosomal ITS region. Plant Systemat. Evol. 2010, 285, 209–232. [Google Scholar] [CrossRef]
- Koenig, D.; Weigel, D. Beyond the thale: Comparative genomics and genetics of Arabidopsis relatives. Nat. Rev. Genet. 2015, 16, 285–298. [Google Scholar] [CrossRef]
- Zhang, L.; Cai, X.; Wu, J.; Liu, M.; Grob, S.; Cheng, F.; Liang, J.; Cai, C.; Liu, Z.; Liu, B.; et al. Improved Brassica rapa reference genome by single-molecule sequencing and chromosome conformation capture technologies. Hortic. Res. 2018, 5, 50. [Google Scholar] [CrossRef] [Green Version]
- Lysak, M.A.; Koch, M.A.; Beaulieu, J.M.; Meister, A.; Leitch, I.J. The dynamic ups and downs of genome size evolution in Brassicaceae. Mol. Biol. Evol. 2009, 26, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Demianski, A.J.; Chung, K.M.; Kunkel, B.N. Analysis of Arabidopsis JAZ gene expression during Pseudomonas syringae pathogenesis. Mol. Plant Pathol. 2012, 13, 46–57. [Google Scholar] [CrossRef]
- Ye, H.; Du, H.; Tang, N.; Li, X.; Xiong, L. Identification and expression profiling analysis of TIFY family genes involved in stress and phytohormone responses in rice. Plant Mol. Biol. 2009, 71, 291–305. [Google Scholar] [CrossRef]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xiang, Y.; Fang, L.; Wang, Y.; Xin, H.; Li, S. Patterns of gene duplication and their contribution to expansion of gene families in grapevine. Plant Mol. Biol. Rep. 2013, 31, 852–861. [Google Scholar] [CrossRef]
- Jiao, Y.; Leebens-Mack, J.; Ayyampalayam, S.; Bowers, J.E.; McKain, M.R.; McNeal, J.; Rolf, M.; Ruzicka, D.R.; Wafula, E.; Wickett, N.J.; et al. Depamphilis, A genome triplication associated with early diversification of the core eudicots. Genome Biol. 2012, 13, R3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, W.; Yu, H.; Cao, P.; Luo, J.; Wang, N. Identification of TIFY family genes and analysis of their expression profiles in response to phytohormone treatments and Melampsora larici-populina infection in poplar. Front. Plant Sci. 2017, 8, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozo, M.J.; Van Loon, L.C.; Pieterse, C.M.J. Jasmonates-Signals in plant-microbe interactions. J. Plant Growth Regul. 2004, 23, 211–222. [Google Scholar]
- Balbi, V.; Devoto, A. Jasmonate signalling network in Arabidopsis thaliana: Crucial regulatory nodes and new physiological scenarios. New Phytol. 2008, 177, 301–318. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Chico, J.M.; Fernández-Calvo, P.; Solano, R. The ZIM domain mediates homo- and heteromeric interactions between Arabidopsis JAZ proteins. Plant J. 2009, 59, 77–87. [Google Scholar] [CrossRef]
- Thireault, C.; Shyu, C.; Yoshida, Y.; St Aubin, B.; Campos, M.L.; Howe, G.A. Repression of jasmonate signaling by a non-TIFY JAZ protein in Arabidopsis. Plant J. 2015, 82, 669–679. [Google Scholar] [CrossRef]
- Staswick, P.E.; Su, W.; Howell, S.H. Methyl jasmonate inhibition of root growth and induction of a leaf protein are decreased in an Arabidopsis thaliana mutant. Proc. Natl. Acad. Sci. USA 1992, 89, 6837–6840. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Dicke, M. Plant interactions with microbes and insects: From molecular mechanisms to ecology. Trends Plant Sci. 2007, 12, 564–569. [Google Scholar] [CrossRef] [Green Version]
- Mur, L.A.; Laarhoven, L.J.; Harren, F.J.; Hall, M.A.; Smith, A.R. Nitric oxide interacts with salicylate to regulate biphasic ethylene production during the hypersensitive response. Plant Physiol. 2008, 148, 1537–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.F.; Zhong, S.L.; Grierson, D. Recent advances in ethylene research. J. Exp. Bot. 2009, 60, 3311–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gfeller, A.; Liechti, R.; Farmer, E.E. Arabidopsis jasmonate signaling pathway. Sci. Signal. 2006, 3, cm4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mur, L.A.; Prats, E.; Pierre, S.; Hall, M.A.; Hebelstrup, K.H. Integrating nitric oxide into salicylic acid and jasmonic acid/ethylene plant defense pathways. Front. Plant Sci. 2013, 4, 215. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; An, F.; Feng, Y.; Li, P.; Xue, L.; Abu-El-Haija, A.; Jiang, Z.; Kim, J.M.; To, T.K.; Li, W.; et al. De-repression of ethylene-stabilized transcription factors (EIN3/EIL1) mediates jasmonate and ethylene signaling synergy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 12539–12544. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Xue, L.; Chintamanani, S.; Germain, H.; Lin, H.; Cui, H.; Cai, R.; Zuo, J.; Tang, X.; Li, X.; et al. ETHYLENE INSENSITIVE3 and ETHYLENE INSENSITIVE3-LIKE1 repress SALICYLIC ACID INDUCTION DEFICIENT2 expression to negatively regulate plant innate immunity in Arabidopsis. Plant Cell 2009, 21, 2527–2540. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Song, S.; Xiao, L.; Soo, H.M.; Cheng, Z.; Xie, D.; Peng, J. Gibberellin acts through jasmonate to control the expression of MYB21, MYB24, and MYB57 to promote stamen filament growth in Arabidopsis. PLoS Genet. 2009, 5, e1000440. [Google Scholar] [CrossRef] [Green Version]
- Pauwels, L.; Inzé, D.; Goossens, A. Jasmonate-inducible gene: What does it mean? Trends Plant Sci. 2009, 14, 87–91. [Google Scholar] [CrossRef]
- Hou, X.; Lee, L.Y.; Xia, K.; Yan, Y.; Yu, H. DELLAs modulate jasmonate signaling via competitive binding to JAZs. Dev. Cell 2011, 19, 884–894. [Google Scholar] [CrossRef] [Green Version]
- Navarro, L.; Bari, R.; Achard, P.; Lisón, P.; Nemri, A.; Harberd, N.P.; Jones, J.D. DELLAs control plant immune responses by modulating the balance of jasmonic acid and salicylic acid signaling. Curr. Biol. 2008, 18, 650–655. [Google Scholar] [CrossRef]
- Lemarie, S.; Robert-Seilaniantz, A.; Lariagon, C.; Lemoine, J.; Marnet, N.; Jubault, M.; Manzanares-Dauleux, M.J.; Gravot, A. Both the jasmonic acid and the salicylic acid pathways contribute to resistance to the biotrophic clubroot agent Plasmodiophora brassicae in Arabidopsis. Plant Cell Physiol. 2015, 56, 2158–2168. [Google Scholar] [PubMed] [Green Version]
- Liu, T.; Zhang, X.; Yang, H.; Agerbirk, N.; Qiu, Y.; Wang, H.; Shen, D.; Song, J.; Li, X. Aromatic glucosinolate biosynthesis pathway in Barbarea vulgaris and its response to Plutella xylostella infestation. Front. Plant Sci. 2016, 7, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Yang, H.; Ren, L.; Chen, W.; Liu, L.; Liu, F.; Zeng, L.; Yan, R.; Chen, K.; Fang, X. Jasmonic acid-mediated aliphatic glucosinolate metabolism is involved in clubroot disease development in Brassica napus L. Front. Plant Sci. 2018, 9, 750. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Accession No. a | Homologous Loci b | Chromosome No. | Length (aa) | pI | MW (Da) | Localization Predicted | |
|---|---|---|---|---|---|---|---|---|
| 1 | BoJAZ1 | Boc02g02049 | Bol039351 | Chr02 | 241 | 9.30 | 26238.56 | N c |
| 2 | BoJAZ2 | Boc02g02168 | Bol034876 | Chr02 | 158 | 6.16 | 17527.33 | N |
| 3 | BoJAZ3 | Boc02g02274 | Bol041431 | Chr02 | 270 | 9.96 | 28913.75 | N |
| 4 | BoJAZ4 | Boc02g03465 | Bol036100 | Chr02 | 221 | 4.97 | 23089.94 | N |
| 5 | BoJAZ5 | Boc02g03937 | - | Chr02 | 197 | 9.88 | 21860.19 | N |
| 6 | BoJAZ6 | Boc03g01228 | Bol008534 | Chr03 | 131 | 9.85 | 14971.78 | N |
| 7 | BoJAZ7 | Boc03g06386 | Bol034224 | Chr03 | 196 | 9.95 | 21688.00 | N |
| 8 | BoJAZ8 | Boc04g01076 | Bol027372 | Chr04 | 117 | 9.15 | 13127.05 | N |
| 9 | BoJAZ9 | Boc05g01466 | Bol026828 | Chr05 | 255 | 9.71 | 27255.92 | N |
| 10 | BoJAZ10 | Boc05g02247 | Bol022524 | Chr05 | 133 | 9.62 | 15274.14 | N |
| 11 | BoJAZ11 | Boc05g03357 | Bol013829 | Chr05 | 336 | 9.47 | 35762.95 | N |
| 12 | BoJAZ12 | Boc06g00449 | Bol026137 | Chr06 | 236 | 9.30 | 26035.25 | N |
| 13 | BoJAZ13 | Boc06g00630 | Bol026339 | Chr06 | 272 | 9.47 | 30322.98 | N |
| 14 | BoJAZ14 | Boc06g00780 | - | Chr06 | 267 | 9.71 | 28723.60 | N |
| 15 | BoJAZ15 | Boc06g01385 | Bol017418 | Chr06 | 232 | 9.03 | 25342.54 | N |
| 16 | BoJAZ16 | Boc06g01609 | Bol039922 | Chr06 | 225 | 9.23 | 24461.58 | N |
| 17 | BoJAZ17 | Boc08g00802 | Bol009774 | Chr08 | 344 | 9.43 | 38025.74 | N |
| 18 | BoJAZ18 | Boc08g00884 | Bol044840 | Chr08 | 276 | 9.43 | 30024.88 | N |
| 19 | BoJAZ19 | Boc08g02786 | Bol013163 | Chr08 | 292 | 9.51 | 31837.28 | N |
| 20 | BoJAZ20 | Boc08g02867 | Bol029321 | Chr08 | 269 | 8.84 | 29748.45 | N |
| 21 | BoJAZ21 | Boc09g03643 | Bol035782 | Chr09 | 203 | 6.85 | 21420.92 | N |
| 22 | BoJAZ22 | Boc09g04265 | Bol043451 | Chr09 | 198 | 10.07 | 21998.47 | N |
| 23 | BoPPD1 | Boc01g02166 | Bol014725 | Chr01 | 318 | 8.64 | 34439.52 | N |
| 24 | BoPPD2 | Boc08g03725 | Bol006854 | Chr08 | 308 | 8.78 | 33583.44 | N |
| 25 | BoZML1 | Boc01g01504 | Bol009539 | Chr01 | 305 | 6.05 | 33079.78 | N |
| 26 | BoZML2 | Boc01g03057 | Bol018802 | Chr01 | 289 | 6.26 | 31531.93 | N |
| 27 | BoZML3 | Boc03g00258 | - | Chr03 | 305 | 6.11 | 33290.86 | N |
| 28 | BoZML4 | Boc05g02989 | Bol038395 | Chr05 | 291 | 6.13 | 31792.40 | N |
| 29 | BoZML5 | Boc07g00805 | Bol042168 | Chr07 | 260 | 5.74 | 28003.17 | N |
| 30 | BoTIFY1 | Boc01g00534 | Bol017893 | Chr01 | 345 | 9.51 | 37246.04 | N |
| 31 | BoTIFY2 | Boc03g00474 | Bol017492 | Chr03 | 359 | 8.49 | 38823.97 | N |
| 32 | BoTIFY3 | Boc03g01224 | Bol016130 | Chr03 | 122 | 5.06 | 13390.66 | N |
| 33 | BoTIFY4 | Boc03g05199 | Bol036968 | Chr03 | 114 | 8.97 | 13033.89 | N |
| 34 | BoTIFY5 | Boc04g01322 | Bol014138 | Chr04 | 113 | 4.68 | 11939.03 | N |
| 35 | BoTIFY6 | Boc04g04270 | Bol037853 | Chr04 | 122 | 9.10 | 13653.55 | N |
| 36 | BoTIFY7 | Boc05g01338 | - | Chr05 | 182 | 8.59 | 20042.45 | N |
| No. | Paralogous Pairs | Ka a | Ks b | Ka/Ks | Effective Length (bp) | Average S-Sites c | Average N-Sites d |
|---|---|---|---|---|---|---|---|
| 1 | BoJAZ1/BoJAZ12 | 0.136239503 | 0.466279810 | 0.292184007 | 681 | 157.92 | 523.08 |
| 2 | BoJAZ1/BoJAZ9 | 0.288117541 | 0.801855456 | 0.359313564 | 705 | 167.17 | 537.83 |
| 3 | BoJAZ1/BoJAZ19 | 0.289411753 | 0.901233386 | 0.321128531 | 702 | 166.83 | 535.17 |
| 4 | BoJAZ3/BoJAZ14 | 0.090640706 | 0.237924642 | 0.380963929 | 786 | 184.75 | 601.25 |
| 5 | BoJAZ3/BoJAZ15 | 0.129965792 | 0.277005584 | 0.469181128 | 669 | 152.92 | 516.08 |
| 6 | BoJAZ4/BoJAZ21 | 0.116023822 | 0.322633719 | 0.359614682 | 600 | 151.92 | 448.08 |
| 7 | BoJAZ5/BoJAZ7 | 0.116977545 | 0.248486377 | 0.470760395 | 585 | 141.83 | 443.17 |
| 8 | BoJAZ5/BoJAZ22 | 0.077915489 | 0.387530995 | 0.201056147 | 588 | 142.08 | 445.92 |
| 9 | BoTIFY4/BoTIFY6 | 0.125567214 | 0.275793585 | 0.455294180 | 318 | 71.50 | 246.50 |
| 10 | BoTIFY4/BoJAZ8 | 0.128368343 | 0.326326040 | 0.393374500 | 303 | 69.92 | 233.08 |
| 11 | BoJAZ7/BoJAZ22 | 0.120085286 | 0.276642241 | 0.434081524 | 585 | 141.92 | 443.08 |
| 12 | BoJAZ8/BoTIFY6 | 0.116309928 | 0.170137651 | 0.683622511 | 348 | 78.83 | 269.17 |
| 13 | BoJAZ9/BoJAZ12 | 0.291213635 | 0.704555605 | 0.413329527 | 684 | 157.42 | 526.58 |
| 14 | BoJAZ9/BoJAZ18 | 0.096000993 | 0.259398924 | 0.370090173 | 756 | 177.08 | 578.92 |
| 15 | BoJAZ9/BoJAZ19 | 0.084821096 | 0.284963797 | 0.297655691 | 741 | 175.75 | 565.25 |
| 16 | BoJAZ12/BoJAZ16 | 0.185531899 | 0.474852727 | 0.390714611 | 621 | 138.33 | 482.67 |
| 17 | BoJAZ12/BoJAZ18 | 0.269450506 | 0.922697085 | 0.292024881 | 693 | 158.08 | 534.92 |
| 18 | BoJAZ13/BoJAZ17 | 0.270636401 | 1.000601128 | 0.270473812 | 795 | 179.50 | 615.50 |
| 19 | BoJAZ13/BoJAZ20 | 0.254382445 | 1.085309439 | 0.234387020 | 774 | 173.33 | 600.67 |
| 20 | BoJAZ14/BoJAZ15 | 0.157843966 | 0.338195612 | 0.466723875 | 657 | 150.00 | 507.00 |
| 21 | BoJAZ17/BoJAZ20 | 0.130211313 | 0.385659245 | 0.337633065 | 804 | 180.75 | 623.25 |
| 22 | BoJAZ18/BoJAZ19 | 0.082922850 | 0.304787220 | 0.272068001 | 774 | 181.67 | 592.33 |
| 23 | BoTIFY1/BoTIFY2 | 0.094462148 | 0.272162609 | 0.347079816 | 1014 | 236.58 | 777.42 |
| 24 | BoZML1/BoZML5 | 0.112145386 | 0.405464237 | 0.276585148 | 774 | 183.58 | 590.42 |
| 25 | BoZML2/BoZML4 | 0.059319442 | 0.309010440 | 0.191965817 | 849 | 191.50 | 657.50 |
| 26 | BoPPD1/BoPPD2 | 0.081285177 | 0.335838405 | 0.242036575 | 897 | 211.17 | 685.83 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhao, C.; Yang, L.; Zhang, Y.; Wang, Y.; Fang, Z.; Lv, H. Genome-Wide Identification, Expression Profile of the TIFY Gene Family in Brassica oleracea var. capitata, and Their Divergent Response to Various Pathogen Infections and Phytohormone Treatments. Genes 2020, 11, 127. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020127
Liu X, Zhao C, Yang L, Zhang Y, Wang Y, Fang Z, Lv H. Genome-Wide Identification, Expression Profile of the TIFY Gene Family in Brassica oleracea var. capitata, and Their Divergent Response to Various Pathogen Infections and Phytohormone Treatments. Genes. 2020; 11(2):127. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020127
Chicago/Turabian StyleLiu, Xing, Cunbao Zhao, Limei Yang, Yangyong Zhang, Yong Wang, Zhiyuan Fang, and Honghao Lv. 2020. "Genome-Wide Identification, Expression Profile of the TIFY Gene Family in Brassica oleracea var. capitata, and Their Divergent Response to Various Pathogen Infections and Phytohormone Treatments" Genes 11, no. 2: 127. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11020127