Comparative Analysis of miRNA Expression Profiles between Heat-Tolerant and Heat-Sensitive Genotypes of Flowering Chinese Cabbage Under Heat Stress Using High-Throughput Sequencing

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Total RNA Isolation
2.2. Construction and Sequencing sRNA Libraries
2.3. Identification of Conserved and Novel miRNAs
2.4. Analysis of Differentially Expressed miRNAs
2.5. Prediction of miRNA Secondary Structure
2.6. Target Prediction of Differentially Expressed miRNAs
2.7. GO and KEGG Prediction of miRNA-Related Regulatory Pathways
2.8. Validation of miRNAs Using RT-qPCR
2.9. Statistical Analysis
3. Results
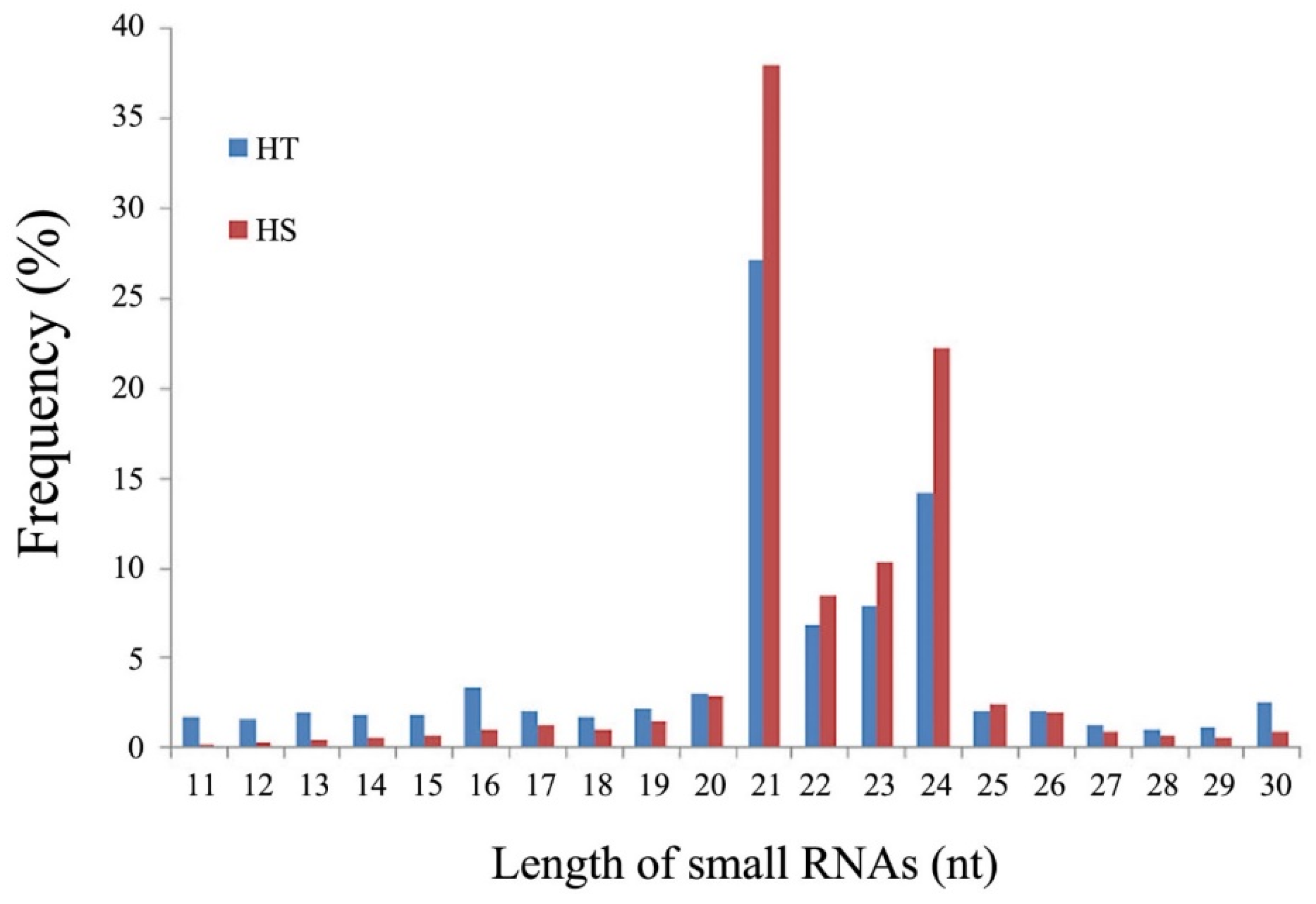
3.1. Analysis and Classification of sRNAs Sequence
3.2. Known miRNAs from Flowering Chinese Cabbage
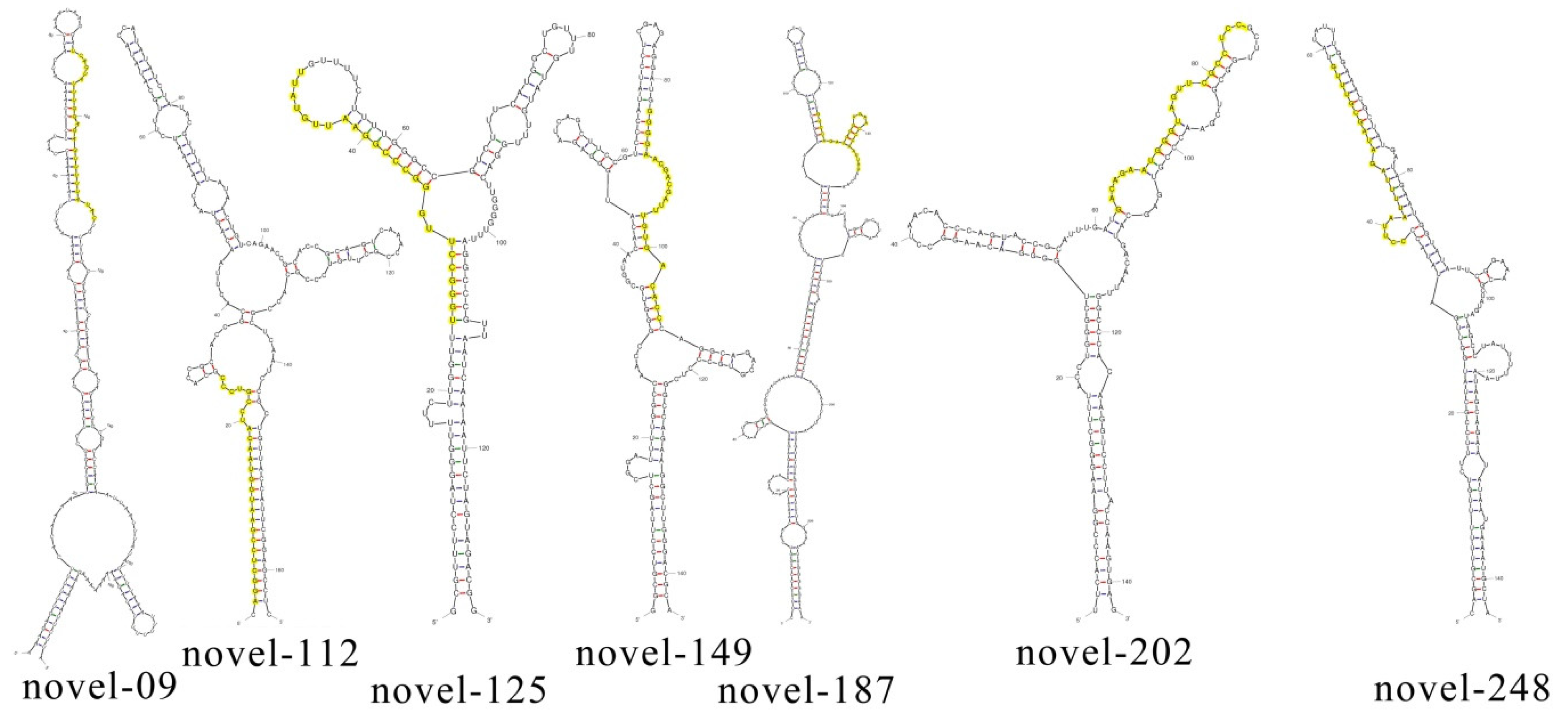
3.3. Identification of Novel miRNAs
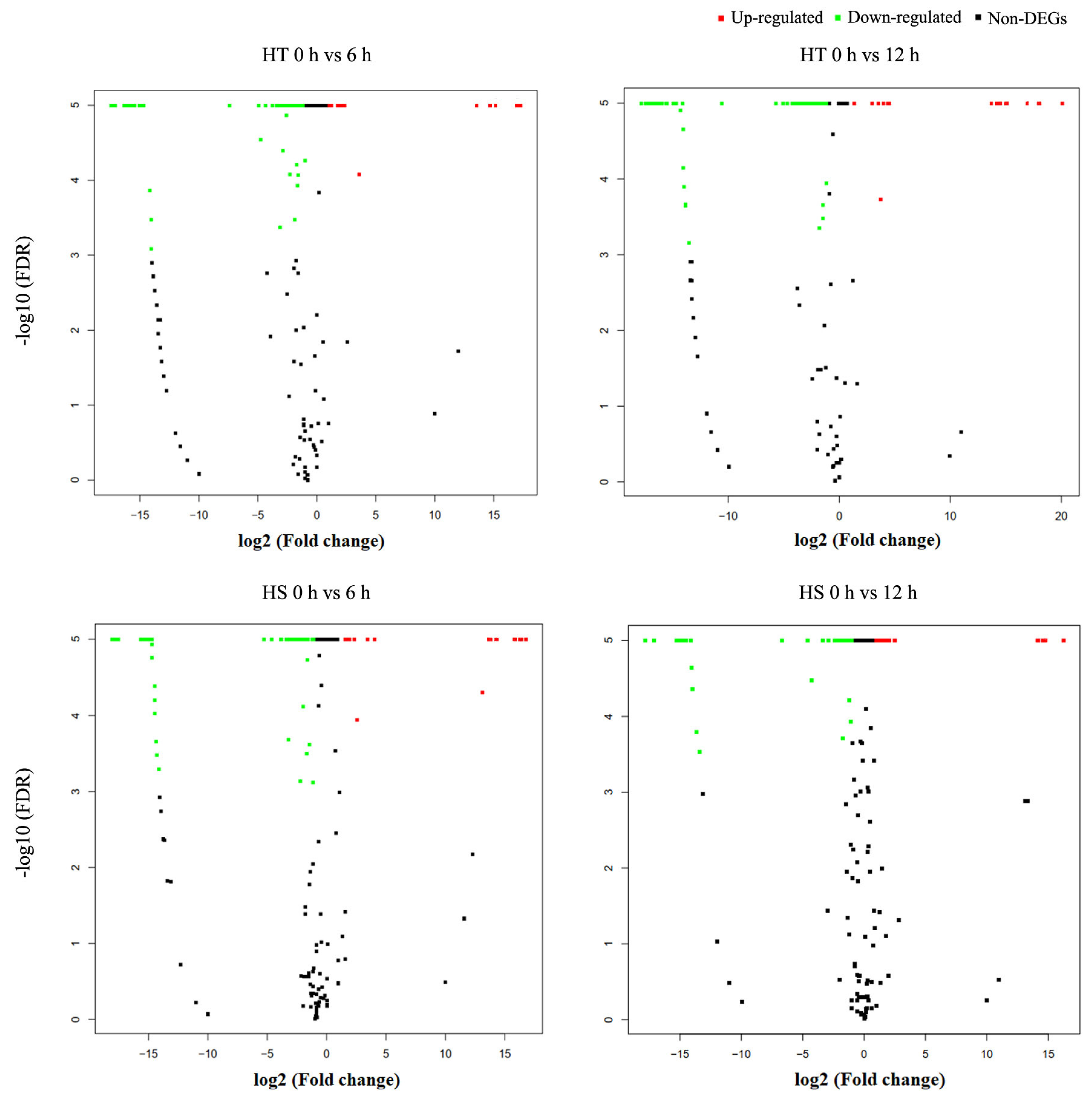
3.4. Differential Expression Profiling of Known and Novel miRNAs
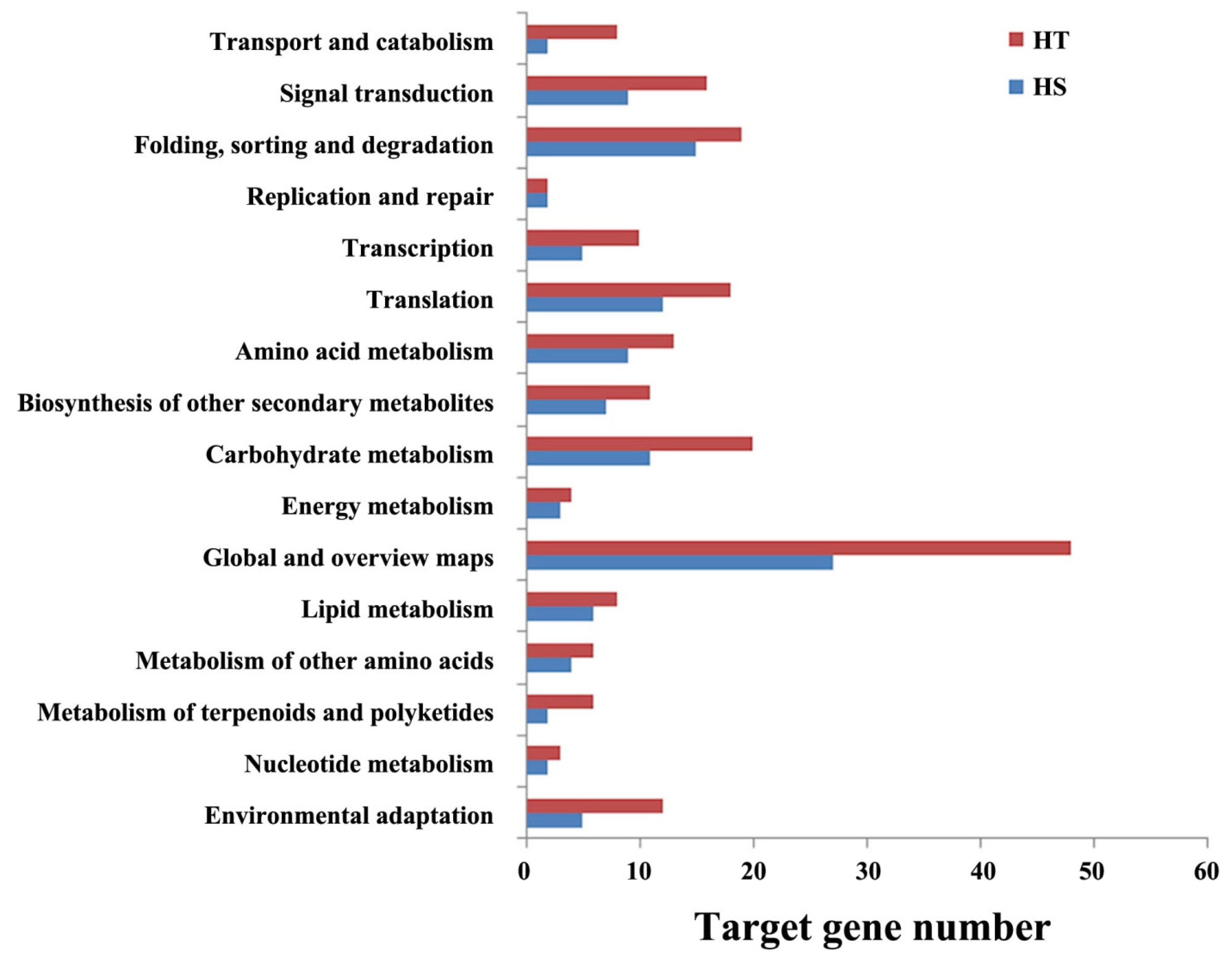
3.5. Putative Target Genes of Differentially Expressed miRNAs in HT and HS Genotypes
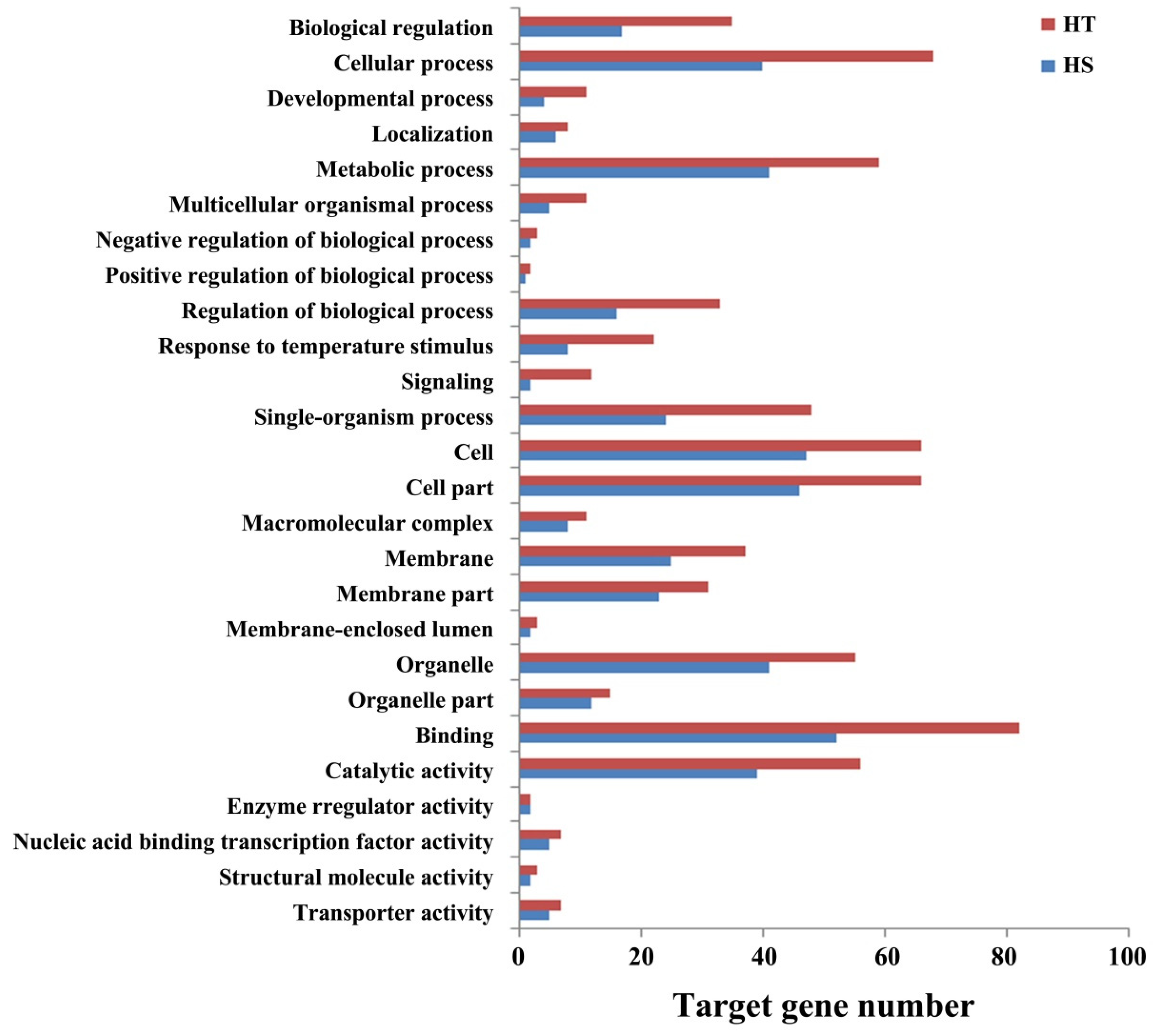
3.6. Functional Annotation of miRNA Target Genes
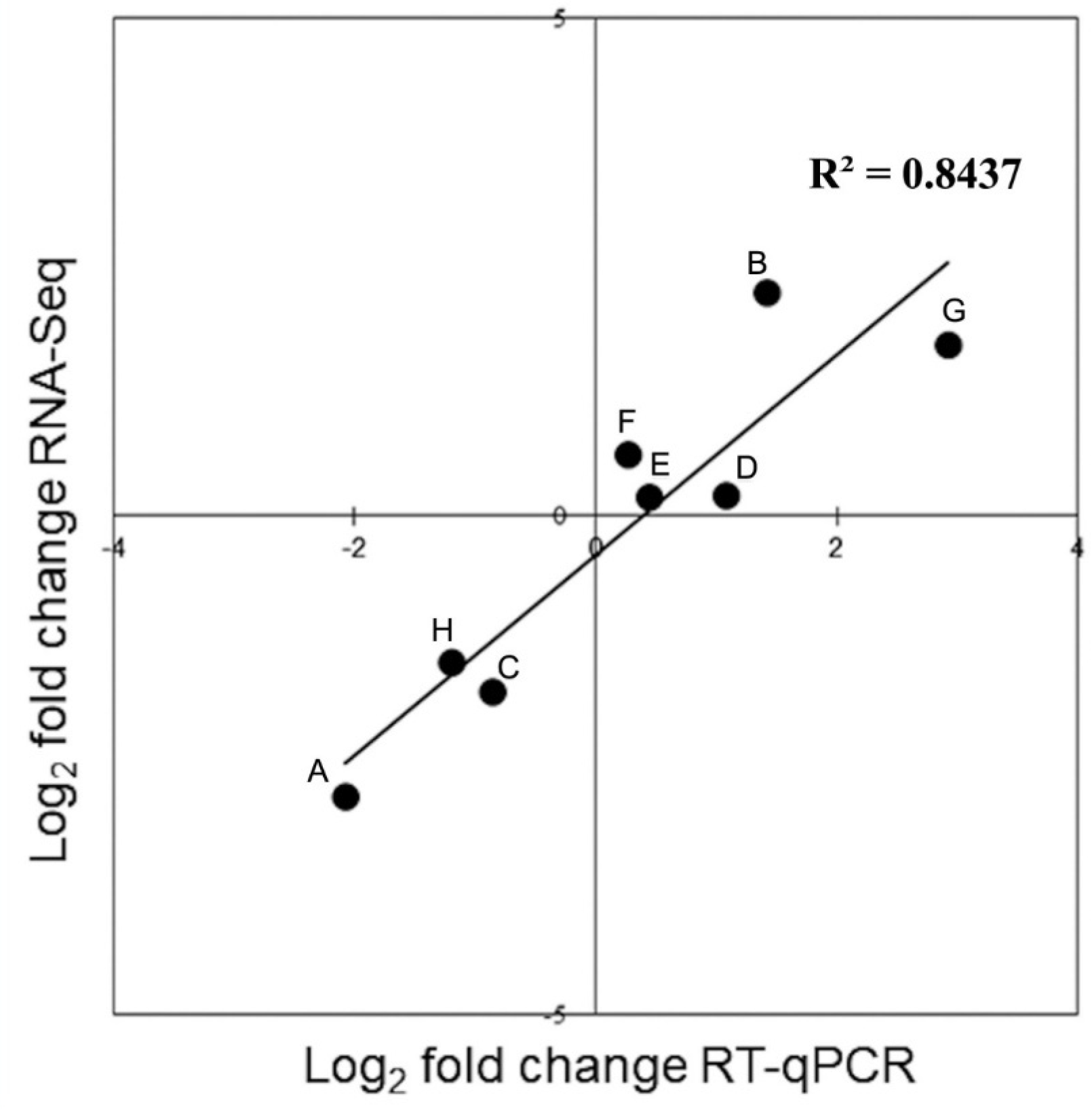
3.7. Validation of Differentially Expressed miRNAs by RT-qPCR
4. Discussion
4.1. miRNA in Flowering Chinese Cabbage
4.2. miRNAs Are Involved in Heat Stress Responses
4.3. miRNA–Targeted Gene-Networks Involved in Response of Flowering Chinese Cabbage to Heat Stress
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, J.; Li, R.; Xia, Y.; Bai, G.; Guo, P.; Wang, Z.; Zhang, H.; Siddique, K.H. Development of EST-SSR markers in flowering Chinese cabbage (Brassica campestris L. ssp. chinensis var. utilis Tsen et Lee) based on de novo transcriptomic assemblies. PLoS ONE 2017, 12, e0184736. [Google Scholar]
- Ahmed, W.; Xia, Y.; Li, R.; Bai, G.; Siddique, K.H.; Guo, P. Non-coding RNAs: Functional roles in the regulation of stress response in Brassica crops. Genomics 2020, 112, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Dresselhaus, T.; Hückelhoven, R. Biotic and Abiotic Stress Responses in Crop Plants; Multidisciplinary Digital Publishing Institute: Basel, Switzerland, 2018. [Google Scholar]
- Young, L.W.; Wilen, R.W.; Bonham-Smith, P.C. High temperature stress of Brassica napus during flowering reduces micro-and megagametophyte fertility, induces fruit abortion, and disrupts seed production. J. Exp. Bot. 2004, 55, 485–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant. Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battisti, D.S.; Naylor, R.L. Historical warnings of future food insecurity with unprecedented seasonal heat. Science 2009, 323, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trend. Plant. Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Zhai, Z.; Lin, Z.; Chen, H.; Chen, Z.; Center, G. Temporal and spatial variation of temperature suitability index for Brassica parachinesis in Guangdong. Guangdong Agric. Sci. 2016, 3, 14. [Google Scholar]
- Sato, S.; Katoh, N.; Iwai, S.; Hagimori, M. Effect of low temperature pretreatment of buds or inflorescence on isolated microspore culture in Brassica rapa (syn. B. campestris). Breed. Sci. 2002, 52, 23–26. [Google Scholar] [CrossRef] [Green Version]
- Kalisz, A.; Siwek, P. Yield and quality of spring Chinese cabbage as affected by different temperature conditions during seedling production. Folia Horticul. 2006, 18, 3–15. [Google Scholar]
- Chen, C.-C.; Fu, S.-F.; Norikazu, M.; Yang, Y.-W.; Liu, Y.-J.; Ikeo, K.; Gojobori, T.; Huang, H.-J. Comparative miRNAs analysis of two contrasting broccoli inbred lines with divergent head-forming capacity under temperature stress. BMC Genom. 2015, 16, 1026. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katiyar-Agarwal, S.; Jin, H. Role of small RNAs in host-microbe interactions. Ann. Rev. Phyto. 2010, 48, 225–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkindale, J.; Hall, J.D.; Knight, M.R.; Vierling, E. Heat stress phenotypes of Arabidopsis mutants implicate multiple signaling pathways in the acquisition of thermotolerance. Plant. Physiol. 2005, 138, 882–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, M.; Li, H.; Jin, C.; Liu, Q.; Chen, C.; Song, W.; Wang, C. Genome-wide identification and characterization of miRNAs in the hypocotyl and cotyledon of cauliflower (Brassica oleracea L. var. botrytis) seedlings. Planta 2014, 239, 341–356. [Google Scholar] [CrossRef]
- Wei, L.; Xiao, M.; Hayward, A.; Fu, D. Applications and challenges of next-generation sequencing in Brassica species. Planta 2013, 238, 1005–1024. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids. Res. 2013, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Wang, H.; Lu, Y.; De Ruiter, M.; Cariaso, M.; Prins, M.; Van Tunen, A.; He, Y. Identification of conserved and novel microRNAs that are responsive to heat stress in Brassica rapa. J. Exp. Bot. 2011, 63, 1025–1038. [Google Scholar] [CrossRef]
- Wang, A.; Hu, J.; Huang, X.; Li, X.; Zhou, G.; Yan, Z. Comparative transcriptome analysis reveals heat-responsive genes in Chinese cabbage (Brassica rapa ssp. chinensis). Front. Plant Sci. 2016, 7, 939. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, A.R.; Joshi, G.; Pandey, R.; Kukreja, B.; Goel, S.; Jagannath, A.; Kumar, A.; Katiyar-Agarwal, S.; Agarwal, M. A genome-wide perspective of miRNAome in response to high temperature, salinity and drought stresses in Brassica juncea (Czern) L. PLoS ONE 2014, 9, e92456. [Google Scholar] [CrossRef]
- Ahmed, W.; Xia, Y.; Zhang, H.; Li, R.; Bai, G.; Siddique, K.H.; Guo, P. Identification of conserved and novel miRNAs responsive to heat stress in flowering Chinese cabbage using high-throughput sequencing. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.I.; Muthusamy, M.; Nawaz, M.A.; Hong, J.K.; Lim, M.-H.; Kim, J.A.; Jeong, M.-J. Genome-wide analysis of spatiotemporal gene expression patterns during floral organ development in Brassica rapa. Mol. Genet. Genom. 2019, 294, 1403–1420. [Google Scholar] [CrossRef] [PubMed]
- Rehman, H.M.; Nawaz, M.A.; Shah, Z.H.; Ludwig-Müller, J.; Chung, G.; Ahmad, M.Q.; Yang, S.H.; Lee, S.I. Comparative genomic and transcriptomic analyses of Family-1 UDP glycosyltransferase in three Brassica species and Arabidopsis indicates stress-responsive regulation. Sci. Rep. 2018, 8, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Lv, M.; Liang, Y.; Ma, Z.; Cao, J. Identification of novel and conserved miRNAs involved in pollen development in Brassica campestris ssp. chinensis by high-throughput sequencing and degradome analysis. BMC Genom. 2014, 15, 146. [Google Scholar]
- Körbes, A.P.; Machado, R.D.; Guzman, F.; Almerao, M.P.; De Oliveira, L.F.V.; Loss-Morais, G.; Turchetto-Zolet, A.C.; Cagliari, A.; Dos Santos Maraschin, F.; Margis-Pinheiro, M. Identifying conserved and novel microRNAs in developing seeds of Brassica napus using deep sequencing. PLoS ONE 2012, 7, e50663. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Method. 2012, 9, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acid. Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Meyers, B.C.; Axtell, M.J.; Bartel, B.; Bartel, D.P.; Baulcombe, D.; Bowman, J.L.; Cao, X.; Carrington, J.C.; Chen, X.; Green, P.J. Criteria for annotation of plant MicroRNAs. Plant Cell. 2008, 20, 3186–3190. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Bonnet, E.; He, Y.; Billiau, K.; Van de Peer, Y. TAPIR, a web server for the prediction of plant microRNA targets, including target mimics. Bioinformatics 2010, 26, 1566–1568. [Google Scholar] [CrossRef]
- Srivastava, P.K.; Moturu, T.R.; Pandey, P.; Baldwin, I.T.; Pandey, S.P. A comparison of performance of plant miRNA target prediction tools and the characterization of features for genome-wide target prediction. BMC Genom. 2014, 15, 348. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acid. Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhang, Z.; Lin, H.; Liu, H.; Chen, J.; Peng, H.; Cao, M.; Rong, T.; Pan, G. Cytoplasmic male sterility-regulated novel microRNAs from maize. Funct. Integr. Genom. 2011, 11, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kulik, A.; Wawer, I.; Krzywińska, E.; Bucholc, M.; Dobrowolska, G. SnRK2 protein kinases—key regulators of plant response to abiotic stresses. OMICS J. Integr. Biol. 2011, 15, 859–872. [Google Scholar] [CrossRef]
- Tian, Y.; Tian, Y.; Luo, X.; Zhou, T.; Huang, Z.; Liu, Y.; Qiu, Y.; Hou, B.; Sun, D.; Deng, H. Identification and characterization of microRNAs related to salt stress in broccoli, using high-throughput sequencing and bioinformatics analysis. BMC Plant Biol. 2014, 14, 226. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell signaling during cold, drought, and salt stress. Plant Cell. 2002, 14, S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Sunkar, R.; Li, Y.-F.; Jagadeeswaran, G. Functions of microRNAs in plant stress responses. Trend. Plant. Sci. 2012, 17, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Lin, L.; Sui, N. Regulation mechanism of microRNA in plant response to abiotic stress and breeding. Mol. Biol. Rep. 2019, 46, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Lu, D.; Mason, A.S.; Li, B.; Xiao, M.; An, S.; Fu, D. Non-coding RNAs and transposable elements in plant genomes: Emergence, regulatory mechanisms and roles in plant development and stress responses. Planta 2019, 250, 23–40. [Google Scholar] [CrossRef]
- Santos, L.S.; Maximiano, M.R.; Megias, E.; Pappas, M.; Ribeiro, S.G.; Mehta, A. Quantitative expression of microRNAs in Brassica oleracea infected with Xanthomonas campestris pv. campestris. Mol. Biol. Rep. 2019, 46, 3523–3529. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.-Y.; Yang, C.-Y.; Ma, Q.-B.; Li, X.-P.; Dong, W.-W.; Nian, H. Identification of wild soybean miRNAs and their target genes responsive to aluminum stress. BMC Plant. Biol. 2012, 12, 182. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Tai, H.; Sun, S.; Zhang, F.; Xu, Y.; Li, W.-X. Cloning and characterization of maize miRNAs involved in responses to nitrogen deficiency. PLoS ONE 2012, 7, e29669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Qin, Y.; Duan, H.; Yin, W.; Xia, X. Genome-wide characterization of new and drought stress responsive microRNAs in Populus euphratica. J. Exp. Bot. 2011, 62, 3765–3779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heisel, S.E.; Zhang, Y.; Allen, E.; Guo, L.; Reynolds, T.L.; Yang, X.; Kovalic, D.; Roberts, J.K. Characterization of unique small RNA populations from rice grain. PLoS ONE 2008, 3, e2871. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, X.; Xu, H.; Chi, X.; Zhang, M.; Hou, X. Identification and characterization of microRNAs and their target genes in Brassica oleracea. Gene 2012, 505, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Pegler, J.L.; Oultram, J.M.; Grof, C.P.; Eamens, A.L. Profiling the abiotic stress responsive microRNA landscape of Arabidopsis thaliana. Plants 2019, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahlaut, V.; Baranwal, V.K.; Khurana, P. miRNomes involved in imparting thermotolerance to crop plants. 3 Biotech 2018, 8, 497. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; He, X.; Qian, L.; Wang, F.; Zhang, Z.; Sun, C.; Lin, L.; Guan, C. Comparative transcriptome analysis in oilseed rape (Brassica napus) reveals distinct gene expression details between nitrate and ammonium nutrition. Genes 2019, 10, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.A.; Azeem, F.; Nawaz, M.A.; Acet, T.; Abbas, A.; Imran, Q.M.; Shah, K.H.; Rehman, H.M.; Chung, G.; Yang, S.H. Transcription factors WRKY11 and WRKY17 are involved in abiotic stress responses in Arabidopsis. J. Plant. Physiol. 2018, 226, 12–21. [Google Scholar] [CrossRef]
- Song, X.; Li, Y.; Cao, X.; Qi, Y. MicroRNAs and their regulatory roles in plant–environment interactions. Ann. Rev. Plant. Biol. 2019, 70, 489–525. [Google Scholar] [CrossRef]
- Chen, Z.; Huo, Q.; Yang, H.; Jian, H.; Qu, C.; Lu, K.; Li, J. Joint RNA-Seq and miRNA profiling analyses to reveal molecular mechanisms in regulating thickness of pod canopy in Brassica napus. Genes 2019, 10, 591. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yu, X.; Wang, H.; Lu, Y.-Z.; De Ruiter, M.; Prins, M.; He, Y.-K. A novel class of heat-responsive small RNAs derived from the chloroplast genome of Chinese cabbage (Brassica rapa). BMC Genom. 2011, 12, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queval, G.; Issakidis-Bourguet, E.; Hoeberichts, F.A.; Vandorpe, M.; Gakière, B.; Vanacker, H.; Miginiac-Maslow, M.; Van Breusegem, F.; Noctor, G. Conditional oxidative stress responses in the Arabidopsis photorespiratory mutant cat2 demonstrate that redox state is a key modulator of daylength-dependent gene expression, and define photoperiod as a crucial factor in the regulation of H2O2-induced cell death. Plant J. 2007, 52, 640–657. [Google Scholar] [PubMed]
- Cheng, M.-C.; Liao, P.-M.; Kuo, W.-W.; Lin, T.-P. The Arabidopsis ETHYLENE RESPONSE FACTOR1 regulates abiotic stress-responsive gene expression by binding to different cis-acting elements in response to different stress signals. Plant Physiol. 2013, 162, 1566–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhou, H.; Zhang, Q.; Zhang, J.; Ni, F.; Liu, C.; Qi, Y. DNA methylation mediated by a microRNA pathway. Mol. Cell 2010, 38, 465–475. [Google Scholar] [CrossRef]
- Zhang, B. MicroRNA: A new target for improving plant tolerance to abiotic stress. J. Exp. Bot. 2015, 66, 1749–1761. [Google Scholar] [CrossRef]
- Fujii, H.; Chiou, T.-J.; Lin, S.-I.; Aung, K.; Zhu, J.-K. A miRNA involved in phosphate-starvation response in Arabidopsis. Curr. Biol. 2005, 15, 2038–2043. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Cai, C.; Zhai, J.; Lin, F.; Li, L.; Shreve, J.; Thimmapuram, J.; Hughes, T.J.; Meyers, B.C.; Ma, J. Coordination of microRNAs, phasiRNAs, and NB-LRR genes in response to a plant pathogen: Insights from analyses of a set of soybean Rps gene near-isogenic lines. Plant. Genom. 2015, 8. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Read Type | HT | HS | ||||
|---|---|---|---|---|---|---|
| 0 h | 6 h | 12 h | 0 h | 6 h | 12 h | |
| Total | 25,331,960 (100%) | 22,632,118 (100%) | 19,999,279 (100%) | 27,647,469 (100%) | 26,043,571 (100%) | 26,275,173 (100%) |
| Intergenic | 15,914,365 (62.82%) | 7,884,837 (34.84%) | 14,061,923 (70.31%) | 15,559,731 (56.28%) | 9,035,253 (34.69%) | 12,686,730 (48.28%) |
| Intron | 1,074,553 (4.24%) | 420,049 (1.86%) | 423,989 (2.12%) | 939,369 (3.4%) | 560,265 (2.15%) | 894,981 (3.41%) |
| Exon | 794,907 (3.14%) | 580,063 (2.56%) | 318,139 (1.59%) | 842,483 (3.05%) | 650,488 (2.5%) | 1,283,163 (4.88%) |
| Precursor | 144,519 (0.57%) | 249,867 (1.1%) | 137,914 (0.69%) | 198,968 (0.72%) | 103,036 (0.4%) | 176,153 (0.67%) |
| Mature | 4,129,468 (16.3%) | 3,858,651 (17.05%) | 2,127,450 (10.64%) | 7,304,825 (26.42%) | 3,398,482 (13.05%) | 8,806,566 (33.52%) |
| Rfam other sncRNA | 4412 (0.02%) | 15,136 (0.07%) | 81,044 (0.41%) | 5515 (0.02%) | 7588 (0.03%) | 6884 (0.03%) |
| rRNA | 98,508 (0.39%) | 112,095 (0.5%) | 252,496 (1.26%) | 148,003 (0.54%) | 83,184 (0.32%) | 180,019 (0.69%) |
| snRNA | 16,377 (0.06%) | 27,407 (0.12%) | 11,777 (0.06%) | 13,109 (0.05%) | 8068 (0.03%) | 18,387 (0.07%) |
| snoRNA | 7442 (0.03%) | 6920 (0.03%) | 6907 (0.03%) | 5794 (0.02%) | 4392 (0.02%) | 7306 (0.03%) |
| tRNA | 65,593 (0.26%) | 22,894 (0.1%) | 1,076,247 (5.38%) | 43,977 (0.16%) | 27,661 (0.11%) | 24,478 (0.09%) |
| Unmapped | 3,081,816 (12.17%) | 9,454,199 (41.77%) | 1,501,393 (7.51%) | 2,585,695 (9.35%) | 12,165,154 (46.71%) | 2,190,506 (8.34%) |
| miRNA Name | Sequence (5>3) | HT Count (0 h) | HT Count (6 h) | log2 Ratio (HT 6/HT 0) | p-Value | HT Count (0 h) | HT Count (12 h) | log2 Ratio (HT 12/HT 0) | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Novel miRNAs | |||||||||
| novel_mir27 | TCAAGCTGGTGTCTGGATGAGT | 155 | 47 | −1.71 | 3.28 × 10−5 | 155 | 0 | −17.23 | 6.96 × 10−41 |
| novel_mir95 | GGTGAGGTCGCTCTGAGAGGATAG | 29 | 0 | −14.8 | 1.89 × 10−6 | 29 | 0 | −14.77 | 3.32 × 10−8 |
| novel_mir149 | GGGGAACGACGATTTGTGACACC | 143 | 0 | −17.1 | 2.41 × 10−29 | 143 | 29 | −2.25 | 7.67 × 10−15 |
| novel_mir46 | GAAAACTATTCGATACATATGGCC | 0 | 15 | 13.66 | 5.58 × 10−6 | ||||
| novel_mir89 | AGGGACAGAGGACTGACATGTGGC | 107 | 0 | −16.62 | 1.96 × 10−28 | ||||
| novel_mir105 | ACTAAATCTACACCAATATTGAT | 85 | 0 | −16.29 | 9.93 × 10−23 | ||||
| novel_mir114 | ATTCTTGAGTCCTTAATACATATA | 20 | 0 | −14.29 | 7.16 × 10−6 | ||||
| novel_mir120 | GACTCTAAAAATACCCTTGGTACTT | 79 | 21 | −1.85 | 6.84 × 10−7 | ||||
| novel_mir177 | TATTCCCGCGAAACCCACGGC | 0 | 13 | 13.55 | 1.82 × 10−6 | ||||
| novel_mir238 | CCTGCGGCTGCGGCGATATT | 241 | 0 | −17.86 | 3.46 × 10−63 | ||||
| novel_mir240 | CAATGGGATCCGCGAACAGTGCA | 17 | 0 | −14.05 | 4.29 × 10−5 | ||||
| novel_mir243 | GCTGATGGAACACTGGCCCGGCCCA | 0 | 20 | 14.21 | 1.03 × 10−7 | ||||
| novel_mir250 | TATAGTTAGGCGTTAGGCACTATG | 104 | 0 | −16.67 | 1.17 × 10−27 | ||||
| novel_mir255 | CAAGCGGTTCAACTGCGGTGCGGT | 1494 | 494 | −1.67 | 5.79 × 10−31 | ||||
| Known miRNAs | |||||||||
| bra-miR156e-3p | TGCTCACCTCTCTTTCTGTCAGT | 1971 | 696 | −1.51 | 6.29 × 10−90 | ||||
| bra-miR824 | TAGACCATTTGTGAGAAGGGA | 1026 | 396 | −1.39 | 2.77 × 10−13 | 1026 | 261 | −2.01 | 7.86 × 10−77 |
| bra-miR1885a | CATCAATGAAAGGTATGATTCC | 1271 | 396 | −1.68 | 4.82 × 10−31 | ||||
| bra-miR1885b | TACATCTTCTCCGCGGAAGCTC | 1546 | 572 | −1.42 | 1.63 × 10−22 | ||||
| bra-miR172d-5p | GCAGCATCATTAAGATTCACA | 1 | 14 | 3.70 | 0.00011 | ||||
| bra-miR400-5p | TATGAGAGTATTATAAGTCAC | 750 | 231 | −1.7 | 1.77 × 10−19 | ||||
| bra-miR396-3p | GCTCAAGAAAGCTGTGGGAAA | 1694 | 693 | −1.29 | 2.00 × 10−16 | ||||
| bra-miR391-5p | TTCGCAGGAGAGATAGCGCCA | 335 | 110 | −1.59 | 3.48 × 10−8 | ||||
| bra-miR2111b-3p | ATCCTCGGGATACGGATTACC | 30 | 1 | −4.75 | 1.48 × 10−5 | 30 | 0 | −14.72 | 1.82 × 10−8 |
| miRNA Name | Sequence (5>3) | HT Count (6 h) | HS Count (6 h) | log2 Ratio (HT 6/HS 6) | p-Value | HT Count (12 h) | HS Count (12 h) | log2 Ratio (HT 12/HS 12) | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| bra-miR156a-3p | GCTTACTCTCTCTCTGTCACC | 39 | 92 | −1.21 | 3.5 × 10−6 | 26 | 111 | −2.09 | 3.02 × 10−9 |
| bra-miR164e-3p | CACGTGCTCCCCTCCTCCAAC | 15 | 0 | 13.66 | 1.85 × 10−5 | 14 | 0 | 13.77 | 6.52 × 10−6 |
| bra-miR164e-5p | TGGAGAAGCAGGGCACGTGCAA | 4 | 49 | −3.61 | 4.42 × 10−11 | 36 | 11 | 1.66 | 3.41 × 10−6 |
| bra-miR390-5p | AAGCTCAGGAGGGATAGCGCC | 130 | 63 | 1.02 | 1.26 × 10−7 | 577 | 164 | 1.77 | 4 × 10−83 |
| bra-miR5712 | AATATTAATATAATTGGTGAG | 18 | 71 | −1.97 | 4.17 × 10−8 | 95 | 477 | −2.40 | 1.41 × 10−41 |
| novel_mir23 | ACCCGTCCATGGGCCCCAGGCTCA | 0 | 37 | −15.13 | 2.31 × 10−11 | 26 | 182 | −2.79 | 1.98 × 10−16 |
| novel_mir112 | AGGCTCCGAATGGTAACATCCGTCCC | 97 | 0 | 16.55 | 2.93 × 10−31 | 92 | 477 | −2.37 | 1.38 × 10−30 |
| novel_mir128 | AATTAAGAAACTCCCATTGGACCGC | 0 | 16 | −13.87 | 2.56 × 10−5 | 24 | 79 | −1.72 | 3.31 × 10−5 |
| novel_mir134 | ACGTGGAACACTCTGACTAGTCTGAC | 21 | 0 | 14.35 | 2.39 × 10−7 | 0 | 97 | −16.56 | 2.1 × 10−24 |
| novel_mir225 | CCTGCGGCTGCGGCGATATT | 56 | 13 | 1.97 | 2.52 × 10−8 | 409 | 1569 | −1.96 | 9.67 × 10−98 |
| miRNA | Target Name | Target Id | E-Value | Putative Function of Target |
|---|---|---|---|---|
| Target genes of heat responsive miRNAs in the heat-tolerant genotype | ||||
| novel-mir128 | BraA09g043410.3C | XP_018458905.1 | 1.40E-77 | Dihydrolipoyllysine-residue acetyltransferase component 1 of pyruvate dehydrogenase complex |
| novel-mir243 | BraA09g015030.3C | XP_013688447.1 | 2.10E-291 | Disease resistance protein RML1A-like isoform X1 |
| novel-mir243 | BraA02g007610.3C | XP_013616826.1 | 4.5E-53 | Disease resistance protein TAO1-like |
| novel-mir243 | BraA02g036020.3C | XP_009129096.1 | 2.2E-82 | Protein PHLOEM PROTEIN 2-LIKE A5-like |
| novel-mir243 | BraA06g012120.3C | XP_009149053.1 | 1.30E-195 | Protein PHLOEM PROTEIN 2-LIKE A8-like |
| novel-mir243 | BraA09g016880.3C | XP_013659354.1 | 4.20E-87 | Putative disease resistance protein At4g11170 |
| Target genes of general heat-stress responsive miRNAs that were differentially expressed between HT and HS genotypes | ||||
| novel-mir23 | BraA01g022410.3C | XP_018508806.1 | 1.30E-183 | Protein strictosidine synthase |
| novel-mir78 | BraA05g033150.3C | XP_009146435.1 | 1.60E-77 | Uncharacterized protein LOC103870086 |
| novel-mir78 | BraA05g027310.3C | XP_009145697.1 | 8.20E-247 | Uncharacterized protein LOC103869376 |
| novel_mir99 | BraA08g020340.3C | XP_009109238.1 | 1.9E-272 | Protein NBR1 homolog |
| novel-mir125 | BraA04g007130.3C | KHN00936.1 | 1.60E-250 | SEC12-like protein 1 |
| novel-mir151 | BraA10g012290.3C | XP_013639528.1 | 7.70E-54 | Uncharacterized protein LOC106344762 |
| novel-mir187 | BraA07g017040.3C | XP_009103473.1 | 4.80E-200 | Serine/threonine-protein kinase SRK2I |
| novel-mir202 | BraA01g001310.3C | XP_013683960.1 | 7.40E-229 | Probable N-acetyltransferase HLS1 |
| novel-mir214 | BraA08g005730.3C | XP_013605618.1 | 3.20E-27 | Serine hydroxymethyltransferase 7 |
| novel-mir214 | BraA09g031950.3C | XP_009123743.1 | 1.30E-280 | Protein DETOXIFICATION 23 |
| novel-mir255 | BraA08g009450.3C | NP_198334.1 | 3.30E-260 | Purple acid phosphatase 26 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, W.; Li, R.; Xia, Y.; Bai, G.; H. M. Siddique, K.; Zhang, H.; Zheng, Y.; Yang, X.; Guo, P. Comparative Analysis of miRNA Expression Profiles between Heat-Tolerant and Heat-Sensitive Genotypes of Flowering Chinese Cabbage Under Heat Stress Using High-Throughput Sequencing. Genes 2020, 11, 264. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11030264
Ahmed W, Li R, Xia Y, Bai G, H. M. Siddique K, Zhang H, Zheng Y, Yang X, Guo P. Comparative Analysis of miRNA Expression Profiles between Heat-Tolerant and Heat-Sensitive Genotypes of Flowering Chinese Cabbage Under Heat Stress Using High-Throughput Sequencing. Genes. 2020; 11(3):264. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11030264
Chicago/Turabian StyleAhmed, Waqas, Ronghua Li, Yanshi Xia, Guihua Bai, Kadambot H. M. Siddique, Hua Zhang, Yansong Zheng, Xinquan Yang, and Peiguo Guo. 2020. "Comparative Analysis of miRNA Expression Profiles between Heat-Tolerant and Heat-Sensitive Genotypes of Flowering Chinese Cabbage Under Heat Stress Using High-Throughput Sequencing" Genes 11, no. 3: 264. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11030264