
Circadian Clock Components Offer Targets for Crop Domestication and Improvement


Department of Biological Sciences, Dartmouth College, Hanover, NH 03755, USA
Genes 2021, 12(3), 374; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030374
Submission received: 30 January 2021
/
Revised: 1 March 2021
/
Accepted: 4 March 2021
/
Published: 6 March 2021
(This article belongs to the Special Issue Molecular Mechanisms of Circadian Clock Function in Plants)
{kind=link}
{kind=link}
{kind=link}
Abstract
:During plant domestication and improvement, farmers select for alleles present in wild species that improve performance in new selective environments associated with cultivation and use. The selected alleles become enriched and other alleles depleted in elite cultivars. One important aspect of crop improvement is expansion of the geographic area suitable for cultivation; this frequently includes growth at higher or lower latitudes, requiring the plant to adapt to novel photoperiodic environments. Many crops exhibit photoperiodic control of flowering and altered photoperiodic sensitivity is commonly required for optimal performance at novel latitudes. Alleles of a number of circadian clock genes have been selected for their effects on photoperiodic flowering in multiple crops. The circadian clock coordinates many additional aspects of plant growth, metabolism and physiology, including responses to abiotic and biotic stresses. Many of these clock-regulated processes contribute to plant performance. Examples of selection for altered clock function in tomato demonstrate that with domestication, the phasing of the clock is delayed with respect to the light–dark cycle and the period is lengthened; this modified clock is associated with increased chlorophyll content in long days. These and other data suggest the circadian clock is an attractive target during breeding for crop improvement.
1. Introduction
1.1. Plant Domestication
About 10,000 years ago, shortly after the end of the most recent ice age, humans began the transition from foraging to farming with extraordinary consequences for both the human domesticators and their domesticated plants and animals, as well as for their non-domesticated counterparts [1]. Plants were domesticated in multiple regions around the globe. For example, emmer and einkorn wheat, barley, peas, lentils, chickpeas, and flax were domesticated in the so-called Fertile Crescent of southwest Asia [1]. Rice was domesticated in China [2] and teosinte was domesticated to maize in Mesoamerica [3].
Plant domestication entails the exposure of wild species to new selective environments associated with human cultivation and use [1,4,5]. Although the ways in which plants are cultivated and used (for example, plants can be grown for fruits or seeds versus vegetative organs) influences the traits selected for during domestication, it is generally recognized that a common suite of traits are associated with the domestication of seed and fruit crops [1,6]. Typically, domestication is associated with increased fruit or grain size, although often the number of fruits or seeds is reduced, an increase in overall plant robustness, more determinate growth or increased apical dominance (reduced growth of side stems in comparison to the central stem), and a retention of the seeds on the plant for easy harvest. Other commonly encountered domestication traits include a loss of seed dormancy, a decrease in bitter substances in edible structures, changes in photoperiod sensitivity, and synchronized flowering [1].
Domestication is accompanied by considerable loss of genetic diversity because the farmers typically used only a limited number of individuals of the progenitor species. Moreover, only seeds from the best plants are retained for the next generation so this loss of diversity increases with each successive generation during the domestication process resulting in a genetic bottleneck. The extent of this loss of diversity depends on the population size during the domestication period and the duration of that period [7]. However, genetic diversity is not lost equally throughout the genome [1]. For genes that influence the desirable phenotypes (like those described above) the loss of diversity is greater because plants carrying selected alleles contribute more progeny to each subsequent generation and other alleles are reduced in frequency or eliminated from the population. In contrast, the loss in diversity of genes that do not influence favored phenotypes (neutral genes) is less and simply a function of the strength of the bottleneck in terms of the population size and duration [1,8].
Here, I adopt the terminology of Meyer and Purugganan [9] and define a domestication gene as a gene for which function has been characterized and underlies a trait that has undergone positive selection, and for which the causative mutation is completely or near-completely fixed in all lineages from that single domestication event [9]. Other genes that control important traits but for which causative mutation(s) are segregating in domesticated populations are considered as diversification or improvement genes that played a lineage-specific role in the crop’s regional adaptation or subsequent improvement. Initial domestication occurs in the environment of the wild progenitor. However, following this initial domestication, there is frequently a period of range expansion in which successfully domesticated crops are moved through trade into new geographic regions where they encounter novel environmental features, both abiotic and biotic. This range expansion, therefore, can be expected to be accompanied by selection for improved performance in these novel environments. The circadian clock plays an important role in the response of plants to their environment and therefore is a likely target for selection during this second period of diversification or improvement [10]. The purpose of this review is to consider the evidence that such selection for altered clock function has occurred.
1.2. The Plant Circadian Clock
1.2.1. The Plant Circadian Clock Consists of Multiple Interlocked Feedback Loops
The rotation of the earth on its axis means that life in the biosphere is exposed to a daily cycle of light and relative warmth versus dark and relative cold. This drastic daily change in the environment occurs with a predictable 24-h period and a fitness advantage is conferred by the ability to anticipate those environmental changes and adjust physiological and metabolic states in coordination with the expected environmental conditions [11,12,13]. Thus, circadian clocks can be found in all domains of life, including Archaea, Bacteria, and Eukaryota [14]. Within the Eukaryota, circadian clocks found in animals, fungi, and plants all have a common architecture of interlocked feedback loops, although the molecular components that comprise plant clocks do not seem to be evolutionarily related to those in animal and fungal clocks [15]. However, plant clocks seem to be complex and include more feedback loops than animal and fungal clocks. This may be a consequence of a proliferation of clock genes resulting from the multiple instances of polyploidization that have occurred during plant evolutionary history [16].
In plants, the oscillator mechanism central to circadian rhythm generation consists of multiple interlocked transcription/translation feedback loops [17,18]. Proteins within the oscillator provide feedback by directly or indirectly repressing their own activity or their own transcription. One cycle of activation and repression of these proteins, followed by activation again, takes ~24 h and generates the period of the circadian rhythm. Although transcriptional activation and repression contribute critically to plant clock function, many post-transcriptional regulatory mechanisms also play integral roles. For example, alternative splicing has been shown to affect circadian timing [19,20,21] and mutations affecting spliceosomal components and their modification alter circadian period [22,23,24,25]. Protein synthesis, protein post-translational modification, protein stability, and protein subcellular localization also play important roles in clock function [26,27]. For example, after dusk ZEITLUPE (ZTL), an F-box protein with a blue light photosensing LOV (Light, Oxygen, Voltage-sensitive) domain, targets PRR5 and TOC1 for ubiquitylation and proteasomal degradation [28,29,30,31,32,33,34,35].
Most of our knowledge of the plant circadian clock has been gained through studies in Arabidopsis. Many of the components of the Arabidopsis circadian oscillator have been identified, although our understanding of the circadian oscillator remains incomplete and new components and regulatory relationships continue to be identified and characterized; e.g., [36,37]. The first identified plant circadian clock component was TIMING OF CAB EXPRESSION1 (TOC1), so-named because mutations that impair TOC1 function shorten the period of the circadian rhythm as assayed using the CAB2:LUC reporter [38]. The abundances of both TOC1 mRNA and TOC1 protein show circadian rhythms, with mRNA abundance maximal at about dusk and protein abundance maximal at night [39]. When TOC1 is expressed constantly using a heterologous promoter, all circadian rhythms tested are abolished, demonstrating that rhythmic expression of TOC1 is essential for function of the circadian clock [40]. TOC1 encodes a nuclear protein with sequence motifs similar to those found in two-component signal-transduction systems common in bacteria and functions as a transcriptional repressor [39,41].
Two other oscillator proteins are the nuclear MYB-related transcription factors LATE ELONGATED HYPOCOTYL (LHY) and CIRCADIAN CLOCK ASSOCIATED 1 (CCA1). These are closely related proteins whose abundance shows circadian rhythms, peaking at dawn. Mutations that impair the function of CCA1 or LHY cause circadian rhythms to cycle with a short period, and inactivating both genes dramatically shortens period, indicating that these two proteins function at least partially redundantly. As with TOC1, constant expression of either CCA1 or LHY causes arrhythmicity, establishing the necessity of cycling expression of CCA1 and LHY for circadian clock function [42,43,44,45].
A model was proposed in which LHY and CCA1 interact with TOC1 to create a negative autoregulatory feedback loop at the heart of the plant circadian oscillator [46]. LHY and CCA1 begin to accumulate just before dawn and repress TOC1 expression and eventually, as their proteins accumulate, repress their own expression. As the levels of LHY and CCA1 proteins fall, the expression of TOC1 mRNA rises, with a peak of expression at the end of the day. TOC1 protein then indirectly activates the expression of the LHY and CCA1 genes, thereby starting another cycle.
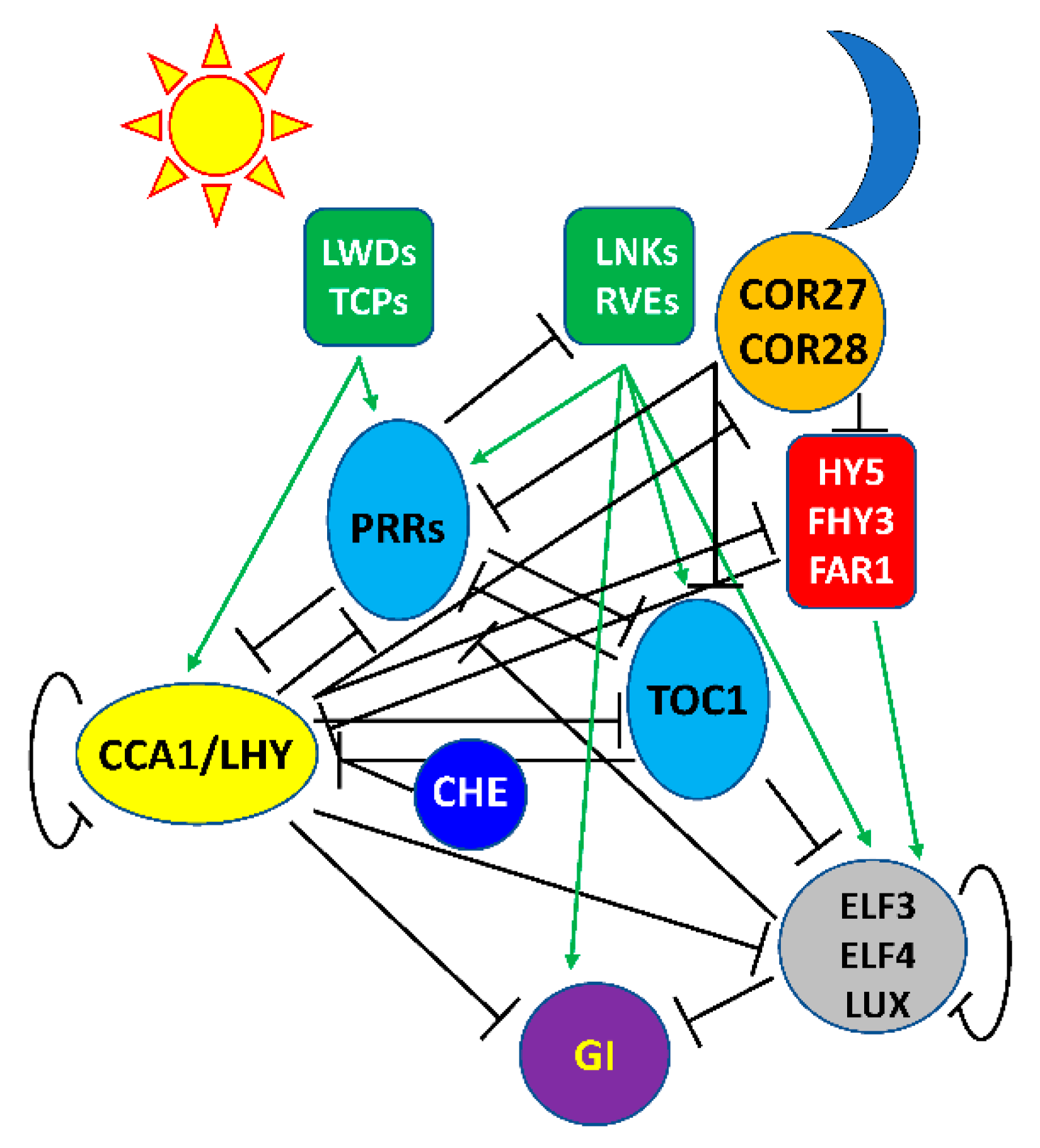
It soon became clear that this simple model was incomplete; we now know that the Arabidopsis circadian clock is much more complex, with more than 20 transcription factors assembled into multiple interlocked feedback loops (Figure 1). CCA1/LHY are the first in a progression of transcriptional repressors expressed sequentially over the day: CCA1/LHY expression is followed by that of a family of PSEUDO RESPONSE REGULATORS (PRRs) closely related to TOC1 in the sequence of PRR9, PRR7, and PRR5 [18,47,48,49] and then TOC1 at dusk. TOC1 is a transcriptional repressor and interacts with TEOSINTE BRANCHED1-CYCLOIDEA-PCF21 (TCP21, also called CHE) to repress CCA1 [50]. The other PRRs are also transcriptional repressors, interacting with the transcriptional corepressor TOPLESS (TPL) through a conserved EAR motif [51]. Recently it has also been shown that COLD-REGULATED 27 (COR27) and COR28 repress PRR5 and TOC1 expression [36,37]. COR27 and COR28 lack DNA-binding activity and likely serve as co-repressors by interacting with as-yet unidentified DNA-binding transcription factors. COR27 and COR28 expression is repressed by CCA1 [37]. The Evening Complex (EC), consisting of LUX ARRHYTHMO (LUX) or the close LUX homolog, BROTHER OF LUX ARRHYTHMO (BOA, also known as NOX), complexed with EARLY FLOWERING 3 (ELF3) and ELF4, accumulates after dusk [52,53,54]. The EC and all of these other components function as transcriptional repressors and each represses expression of the previous and subsequent components in the progression. The EC accumulates after dusk and maintains repression of CCA1 and LHY, restricting their expression to late night and early morning.
Much less is known about transcriptional activators in the circadian oscillator, likely due to functional redundancy hindering their identification via loss of function mutations. Expression of CCA1 and LHY is activated around dawn by a complex of the LIGHT-REGULATED WD1 (LWD1) and LWD2 with DNA-binding TCP20/TCP22 transcription factors [55]. Consistent with a role in the regulation of CCA1 transcription, TCP20 transcript cycles with a pre-dawn maximum [56]. The LWD/TCP complex also activates expression of the PRRs. Later in the afternoon a complex (or complexes) of REVEILLE (RVE) transcription factors (RVE8, RVE4, and RVE6) closely related to CCA1 and LHY together with members of the family of NIGHT LIGHT–INDUCIBLE AND CLOCK-REGULATED (LNK) transcriptional corepressors activate the expression of PRR5, TOC1, GIGANTEA (GI), and ELF4 [57,58,59,60], although expression of the EC remains repressed until after the degradation of PRR5 and TOC1. ELF4 transcription is also activated by FAR-RED ELONGATED HYPOCOTYL3 (FHY3), FAR-RED IMPAIRED RESPONSE1 (FAR1), and ELONGATED HYPOCOTYL5 (HY5), three transcription factors that are positive regulators of phytochrome A signaling [61]. FHY3 and FAR1 also directly bind to the CCA1 promoter to drive light-induced CCA1 expression [62]. Interestingly, CCA1 and LHY interact with and inhibit the transcriptional activation activity of FHY3, FAR1, and HY5, which contributes to the roles of CCA1 and LHY as transcriptional repressors [61]. Similarly, COR27 interacts with HY5 to repress its DNA binding activity [63].
It is not surprising that these rhythmic changes in transcription of clock genes are accompanied by changes in chromatin structure. This was first described for TOC1, where histone acetylation/deacetylation cycles with transcriptional activity [64]. The EC interacts with HISTONE DEACETYLASE9 (HDAC9) to recruit it to the TOC1 promoter to repress TOC1 expression at night [65]. The EC and HDA9 also interact with HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE15 (HOS15) to deacetylate histones at the GI promoter and transcriptionally repress GI expression at night [66]. There are multiple additional examples of epigenetic control of clock gene expression that have been recently reviewed as part of this Special Issue [67].
1.2.2. Photoperiodic Induction of Flowering
Successful reproduction is essential for fitness, so the timing of flowering to ensure maximal reproductive success is subject to natural selection in wild species and likewise has been subject to artificial selection in domesticated species. Flowering can be initiated in response to both environmental cues and endogenous pathways, although their relative importance varies among species. Here, I focus on photoperiodic flowering. Photoperiodism makes it possible for plants to infer seasonality from day length. At the equator, day length and night length are equal and remain constant throughout the year. However, as one moves from the equator towards the poles, the days become longer in summer and shorter in winter. Plants detect these seasonal changes in day length and use them to coordinate their flowering to the appropriate season.
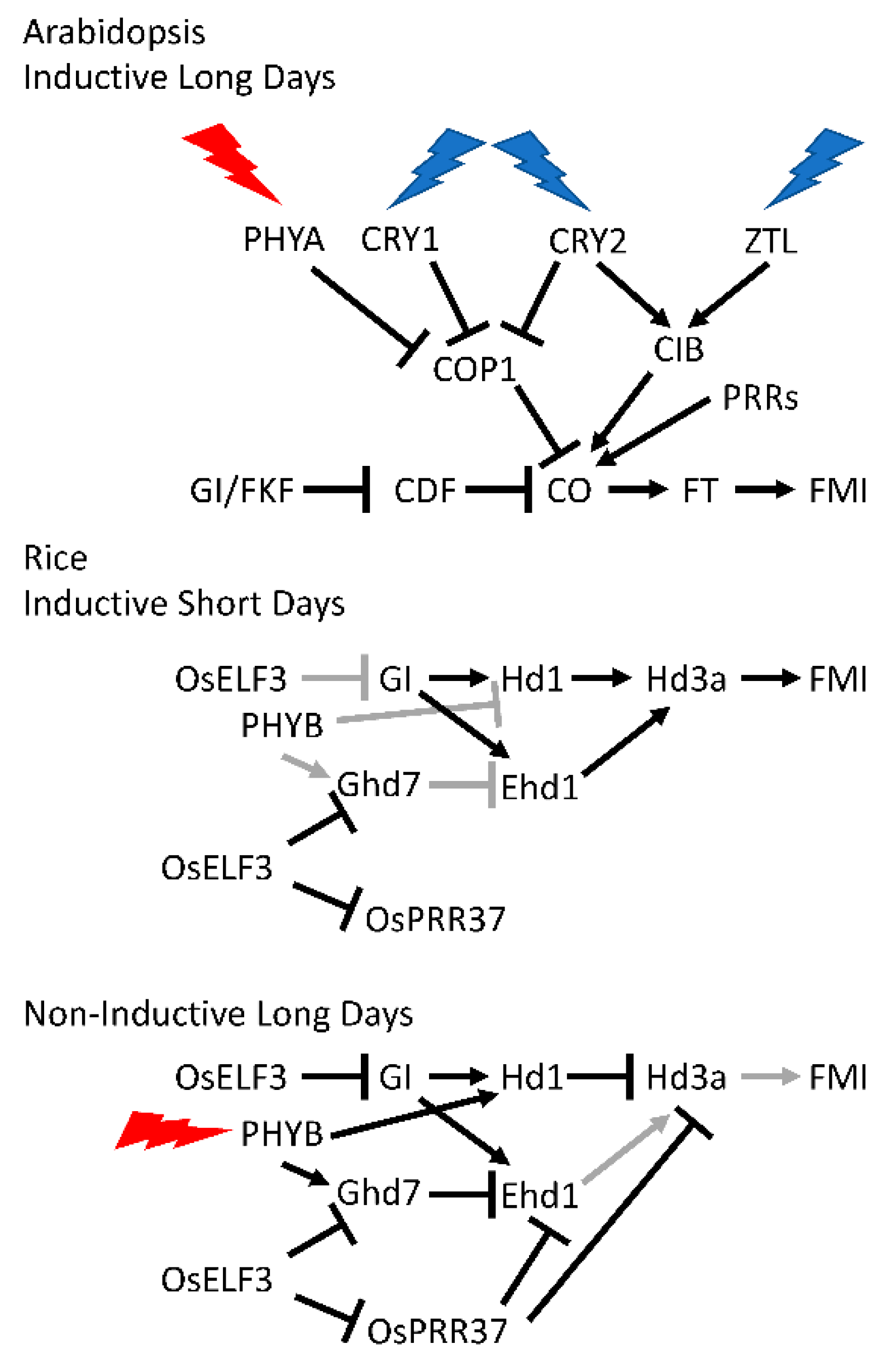
Typically, the leaf is the site of perception of the photoperiodic signal, generating a mobile flowering inducer, termed florigen, that is transmitted to the shoot apical meristem where it induces FLORAL MERISTEM IDENTITY (FMI) genes, the key regulators that initiate the genetic programs required for flower development [68]. This pathway has been worked out in considerable detail in Arabidopsis, in which flowering is accelerated in response to long days (Figure 2). In Arabidopsis, florigen is encoded by the FLOWERING LOCUS T (FT) gene. The circadian clock regulates the induction of FT and of its critical transcriptional inducer, CONSTANS (CO), via an external coincidence mechanism in which light coincides with an inductive window that is restricted (gated) by the circadian clock [69,70].
The circadian clock drives morning-specific expression of several CYCLING DOF FACTOR (CDF) genes whose protein products repress CO transcription. The CDF proteins are targeted for degradation by a SCF complex containing FLAVIN BINDING, KELCH REPEAT, F-BOX1 (FKF1) and the clock component GIGANTEA (GI), both of which cycle in protein abundance. In short days, GI protein abundance peaks at dusk while FKF1 protein peaks after dark. The FKF1-GI complex forms in the dark and only degrades the CO-repressing CDF proteins after dusk. Thus, CO transcription is repressed until about dusk and CO mRNA accumulates after dusk. CO protein is unstable in the dark so, in short days, CO protein fails to accumulate and FT transcription is not induced. However, in long days the phase of peak GI accumulation coincides with that of FKF1 before dusk. The FKF1-GI complex degrades the CDFs in the late afternoon, relieving transcriptional repression of CO. CO mRNA accumulates in the light, which permits the stabilization of nascent CO protein and activation of FT transcription. In this way FT is expressed in the vasculature of the leaf under inductive photoperiods and FT protein travels through the phloem to the shoot apical meristem where it works together with meristem expressed FD to induce FLORAL MERISTEM IDENTITY (FMI) genes and initiate flowering.
A critical element of this photoperiod pathway is the light-mediated stabilization of CO protein. CO protein is degraded via COP1 in the dark, but in the light, PHYA, CRY1, and CRY2 suppress COP1 activity to stabilize CO [70,71,72]. Similarly, the PRRs physically interact with and stabilize CO protein during the day, when they are abundant [73].
FT transcription is also induced independently of CO. Several CRY2-INTERACTING bHLH (CIB) transcription factors accumulate in long days to stimulate FT transcription. The CIBs are activated in the afternoon by blue-light-dependent interaction with CRY2. In addition, CIB protein stability is enhanced via a blue-light-dependent interaction with the FKF1 relatives, ZTL and LKP2, although not with FKF1 [70,74,75].
In crops, flowering time is an important agronomic trait that determines seasonal and regional adaptation. If a cultivar flowers too early in a specific location there will be inadequate use of light and temperature resources, and consequent lower yield. On the contrary, if a cultivar is too late in flowering it cannot complete flowering and grain development before the onset of cold, also resulting in lower yield.
Let us consider flowering time (heading date) in rice, a cereal, as an example (Figure 2). The CO-FT pathway is important in photoperiodic flowering in rice, a short-day plant, but additional pathways also play important roles [70,76]. As in Arabidopsis, florigens encoded by the rice FT homologs, Heading Date 3a (Hd3a) and RICE FLOWERING LOCUS T 1 (RFT1), are induced in the leaf vasculature under inductive (short) days and move to the shoot apical meristem to induce flowering. Hd1, the rice CO ortholog, promotes Hd3a transcription under short days. However, in long days Hd1 is converted from an activator to a repressor by light signaling from phytochrome B and inhibits Hd3a transcription [77,78]. There is also a second bifunctional transcriptional regulator of Hd3a that activates in short days and represses in long days. Thus, although there are general similarities with the Arabidopsis CO-FT pathway, there are clear differences in mechanistic detail [70].
Rice has a second pathway that regulates Hd3a expression. Early heading date 1 (Ehd1), a rice-specific B-type response regulator, upregulates Hd3a expression to promote flowering mainly in short days [79], when blue light signaling coincides with the morning phase set by the circadian clock. Ehd1 expression is inhibited by Ghd7 (a CCT-domain protein encoded by GRAIN NUMBER, PLANT HEIGHT AND HEADING DATE 7), which is regulated by the circadian clock [80,81,82]. OsELF3-1/Hd17/Early flowering7 (Ef7) participates in this repressive regulation of Ghd7 [83,84,85]. Disruption of OsELF3-1/Hd17/Early flowering7 (Ef7) function results in elevated expression of Ghd7 in both long and short days, resulting in reduced Ehd1 and Hd3a expression [84,85]. OsELF3-1/Hd17/Early flowering7 (Ef7) also negatively affects OsGI expression, which is responsible for Ehd1 expression, and OsPRR37, which suppresses expression of Hd3a but not that of RFT1 under long-day conditions [83,84,85]. OsPRR37 functions as a transcriptional repressor expression of day-phased clock genes as well as Ehd1 [86].
Photoperiodic flowering in the long-day cereals, wheat and barley, has been reviewed elsewhere [70] and will not be addressed here due to space limitations.
2. Circadian Clock Components as Domestication and Crop Improvement Loci
As mentioned above, plants were domesticated in a number of distinct geographical regions [1] and it is self-evident that initial domestication occurred in the environment of the wild progenitor. However, many domesticated crops were subsequently moved into new geographic regions where they encountered novel environmental conditions.
Environmental changes associated with range expansion include those in the annual patterns of temperature and photoperiod, both of which vary with latitude [87]. As a consequence, in a number of animals circadian clock function varies systematically with latitude. For example, in Drosophila melanogaster alternative splicing of the clock gene period (per) alters an activity rhythm under warmer temperatures to avoid desiccation [88]. Latitudinal clines are found in two predominant splice forms of per that alter temperature compensation of the clock [89,90,91]. Similarly, latitudinal clines in the frequencies of allelic variants of the circadian rhythm gene Clock are found in birds (Cyanistes caeruleus) [92] and salmon (Oncorhynchus tshawytscha) [93]. This establishes that divergent selection on circadian clock function contributes to local adaptation in animals.
Are similar latitudinal clines in clock function found in plants? Initial studies in Arabidopsis found suggestive correlations of period with latitude [94,95]. A positive correlation was observed between latitude and circadian period in the wildflower, Mimulus guttatus [96]. One might therefore expect to find alterations in clock function that accompany latitudinal expansion in crops.
2.1. Soybean
One phenotype likely to respond to latitude is photoperiodic flowering, and soybean offers a number of excellent examples. Soybean (Glycine max) was domesticated from wild soybean (Glycine soja) in East Asia 6000–9000 years ago [97,98]. Soybean is a short-day plant that flowers when daylength becomes shorter than a critical length [99]. Photoperiod-sensitivity determines the cultivation limits of soybean, making control of flowering time important for regional adaptation and range expansion [100,101,102]. Multiple maturity loci, including 11 E loci, have been identified [103,104,105]. Dominant alleles at E1, E2, E3, E4, E7, E8, and E10 confer late flowering, whereas dominant alleles at E6, E9, and E11 confer early flowering [102]. Of these E loci, E2 is GmGI [106], E3 is GmPHYA3 [107], and E4 is GmPHYA2 [108], all genes with roles in clock oscillator function or in light input to the clock in Arabidopsis. Allelic variation among these E loci is associated with differences in flowering time and adaptation to different latitudes among soybean cultivars [102,109,110,111,112,113,114,115,116,117].
There are a total of 12 FT homologs in soybean [102]. E9 is GmFT2a [118]. GmFT2a and GmFT5a, which are highly induced under inductive short-day conditions, are the most important flowering inducers [99]. These genes are not fully redundant and play distinct roles in floral induction, with loss-of-function of GmFT2a associated with delayed flowering under SD and loss-of-function of GmFT5a associated with delayed flowering under LD [119,120]. GmPHYA3 and GmPHYA2 (E3 and E4, respectively), are negative regulators of both GmFT2a and GmFT5a under non-inductive long-day conditions [99]. GmFT1a, GMFT1b, GmFT4, and GmFT6 act as floral inhibitors, like TFL1 in Arabidopsis [121,122,123,124]. E10 most likely corresponds to GmFT4 [104]. It remains to be determined the extent to which either standing or induced variation among these many FT homologs can contribute usefully to the manipulation of photoperiodic flowering towards improved field performance.
During post-domestication improvement, soybean, a short-day plant, has gained the ability to flower under long-day conditions during the growing season at higher latitudes. This has come about mainly through dysfunction of GmPHYA3 and GmPHYA2 and the floral repressor E1 [103]. Recently, photoperiod insensitivity and earlier flowering in long-day conditions in Far-Eastern Russian soybean cultivars was associated with loss of function of E1-Like b (E1Lb), a homeolog of E1 [125]. Lines carrying the loss-of-function e1lb allele exhibited elevated expression of GmFT2a and GmFT5a and flowered earlier than those carrying the functional E1Lb allele under long-day conditions. Thus, E1Lb retards flowering under long-day conditions by repressing the expression of GmFT2a and GmFT5a independently of E1 [125]. A second E1-Like gene, E1La, also functions as a floral repressor [126]. Early maturity is critical for northern expansion of soybean cultivation in the USA and Canada as well as in northeast Asia. GmFT5a has also been shown to underlie a QTL promoting flowering under long days [127].
GIGANTEA (GI) was first identified in Arabidopsis through loss of function mutations that were very late flowering, allowing prolonged vegetative growth, which explains the name of the locus [128]. In long-day plants such as Pisum sativum, Hordeum vulgare, Triticum aestivum, and Brassica rapa, as in Arabidopsis, GI acts as a flowering activator [129,130,131,132]. In contrast, in soybean and rice, both short day plants, GI acts as a floral repressor [81,106,133]. In soybean, there are three GI loci, GmGIa, GmGIb (GmGI1), and GmGIc (GmGI2), although only GmGIa, which corresponds to E2, has been established to be important in flowering and maturity [106]. However, both GmGIb (GmGI1), and GmGIc (GmGI2) bind to both GmFKF1 and GmFKF2 and to GmCDF1 and so may contribute to flowering time regulation [134]. Nonetheless, there has been no loss of nucleotide diversity of GmGIb and GmGIc in domesticated relative to wild soybean, indicating that they have not been subject to selection during domestication and improvement [135]. In contrast, there has been a great loss in diversity of GmGIa during domestication and improvement [135]. In particular, among Chinese genotypes there has been enrichment for a haplotype that encodes a truncated GI protein and presumably confers at least partial loss of function, permitting earlier flowering in long days. Interestingly, this haplotype was not found among Japanese and Korean wild soybeans [135], although another haplotype with a nonsense mutation in the second exon has been found in Korean early flowering lines [136].
In Arabidopsis it is well-established that GI regulates photoperiodic flowering through an external coincidence mechanism in which in long days the phase of peak GI expression coincides with that of FKF1 in late afternoon [70]. FKF1 is a blue-light photoreceptor, and the interaction of FKF1 with GI is enhanced by blue light [137]. The resultant FKF1-GI complex degrades the CDFs, transcriptional repressors of CO, in the late afternoon of long days [138]. Thus, CO mRNA accumulates in the light, which permits the stabilization of nascent CO protein and consequent activation of FT transcription [139]. However, GI also exerts clock-independent effects on flowering. In particular, Arabidopsis GI positively regulates transcription of miR172 [140], which promotes photoperiodic flowering through a CO-independent genetic pathway. miR172 post-transcriptionally represses a set of APETALA2 (AP2)-like genes, including TARGET OF EAT1 (TOE1), TOE2, and TOE3 that repress FT transcription [140,141]. Overexpressing TOE1 causes late flowering, whereas miR172-overexpressing plants exhibit early flowering under both long and short days [140]. As in Arabidopsis, soybean GmGIa positively regulates Gma-miR172a, although through post-transcriptional processing [142]. Gma-miR172a targets include a soybean TOE1 ortholog, Glyma03g33470. Thus, GmGIa serves as a photoperiod-independent flowering activator by increasing the expression of TOE1 targets, including FT, AP1 and LFY [142].
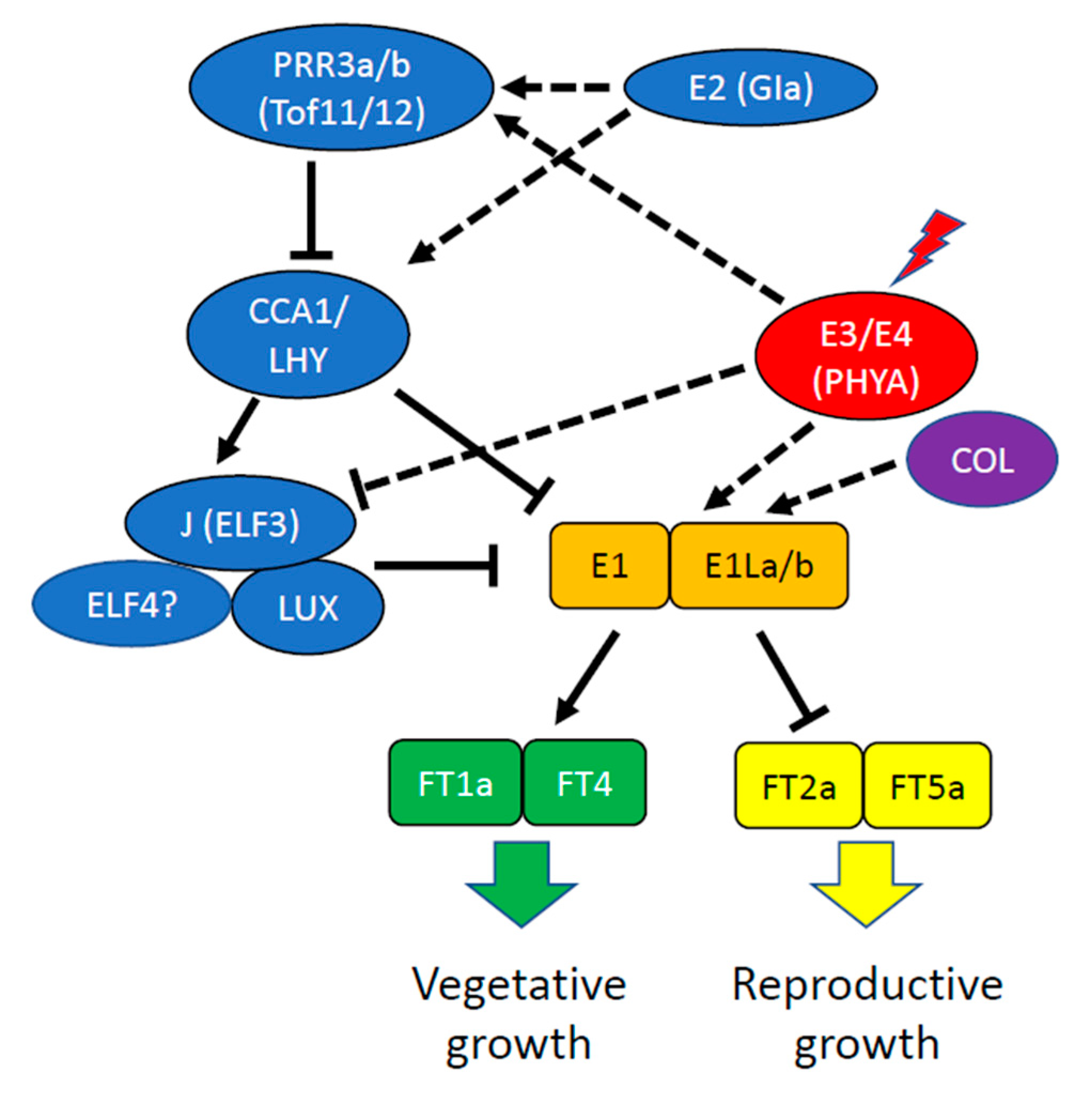
Photoperiodic responses have also hampered the expansion of soybean cultivation into the tropics because temperate varieties matured extremely early, leading to a reduced vegetative size that resulted in extremely low grain yields when temperate lines were grown below 20° latitude. Long juvenile phase (LJ) soybean lines discovered in the 1970s exhibit delayed flowering under short photoperiods, which significantly enhanced yield [143,144]. Two loci, J and E6, have been reported to control this response [145,146]. E6 has been mapped and is tightly linked to J [147]. Genetically, E6 acts as a suppressor of E1 and the function of E6 is dependent on E1. However, the molecular identify of E6 remains to be established [147].
J has been identified as the ortholog of Arabidopsis EARLY FLOWERING3 (ELF3) [148], a component of the EC (Figure 3). GmELF3 is suppressed by PHYA (encoded by E3 and E4), and the GmELF3 protein physically binds to the promoter of E1 near the LUX-binding motif to suppress E1 transcription. This relieves the E1-dependent transcriptional repression of FT2a and FT5a, thereby promoting flowering. Multiple independent loss of function alleles of GmELF3 (J) have been identified in low-latitude genotypes [148]. In LJ lines, with impaired GmELF3 function, E1 itself is released from repression and is able to repress FT2a and FT5a, resulting in later flowering. This permits increased vegetative growth and subsequently increased numbers of flowers, grain set, and yield [148]. This result is reminiscent of the observation that overexpression of an Arabidopsis thaliana B-box domain gene (AtBBX32) or its functional homologs, GmBBX52 and GmBBX53, in transgenic soybean extended the duration of the pod and seed development period and thereby significantly increased soybean grain yield [149]. In Arabidopsis, AtBBX32 is clock regulated and its overexpression in transgenic soybean alters transcript levels of the soybean clock genes GmTOC1 and LHY-CCA1-like2 (GmLCL2) around dawn (chiefly), although it has not been established that these changes in clock gene expression are either necessary or sufficient to confer the reproductive development changes leading to enhanced yield.
ELF3 homologs are important controllers of flowering time in a number of other crops. For example, in chickpea, Cicer arietinum, spring flowering arises in landraces through mutations in CaELF3a [150]. ELF3 homologs control short-day flowering in other legumes, including peas and lentils [151].
Other clock-related loci have been implicated in the control of flowering time. Elite cultivars of soybean exhibit a latitudinal cline in circadian period, with period lengthening at higher latitude [96]. Soybean has six cryptochrome-like (CRY) genes; one, GmCRY1a, is a strong promoter of flowering, and GmCRY1a exhibits a circadian rhythm in protein abundance that varies with latitude and correlates with photoperiodic flowering in long (but not in short) days [152].
In Arabidopsis, five PSEUDO-RESPONSE REGULATORS, important transcriptional repressors in the oscillator mechanism, are expressed sequentially from dawn until after dusk in the order, PRR9, PRR7, PRR5, PRR3, and PRR1 (TOC1) [47]. In soybean, GMPRR3a and GmPRR3b have been shown to underlie growth period QTL, with extended growth period associated with increased yield [153]. In each case, the domesticated allele carries a loss of function (premature stop codon) mutation resulting in loss of the CCT domain [153] that is necessary for normal nuclear localization and DNA-binding [41,154]. Such loss of function alleles were associated with elevated expression of GmFT2a and GmFT5a and earlier flowering [153]. The frequency of this mutation of GmPRR3b increased from 5.6% in wild soybeans to 78.1% in landraces to 98.6% in improved cultivars, consistent with its selection during domestication and improvement [153]. Another study identified GMPRR3a and GmPRR3b as Tof11 and Tof12 and showed that they act as repressors of GmLHY/CCA1 homologs (also called GmLCL) [155]. Soybean has four GmLHY/CCA1 homologs that are clearly important for circadian clock function because a quadruple loss-of-function mutant has an extremely short period [155]. The GmLHY/CCA1 homologs are repressors of E1. Thus, loss of GmPRR3 function leads to upregulation of GmLHY/CCA1, downregulation of E1 and, thereby, induction of GmFT2a and GmFT5a [156]. Conversely, the Gmlhy/cca1 quadruple mutant fails to repress E1 and flowers late [155]. GmPRR3b has also been associated with flowering time and maturity through a genome-wide association study [157]. One specific allele, GmPRR3bH6 is predominant among modern soybean cultivars, consistent with selection during domestication. Its overexpression increases main stem node number and grain yield, and loss of function delays growth and flowering [157]. These effects on flowering may result from the repression of other clock genes, including GmCCA1a, via its regulation of GmELF3a (J).
A second PRR locus, a PRR7 homolog, has been identified as a strong candidate for a flowering time QTL, qFT12-1, although there is no evidence establishing that different alleles of this locus alter clock function [158]. There are an additional nine PRR genes (a total of 12) in soybean, although there is to date no evidence linking these additional genes to domestication or improvement [153].
In Arabidopsis, considerable evidence links the circadian clock to both biotic and abiotic stress responses [12,159,160]. Clock function modulates responses to multiple stresses and, reciprocally, stresses modulate clock function, e.g., [161,162,163,164,165,166]. This reciprocal relationship extends to crops: the circadian clock modulates the drought response in poplar [167] and in B. rapa [168]. The soybean circadian clock has been shown to respond to multiple environmental stresses [169]. Two homologous pairs of GmLHY/CCA1 genes were shown to be negative regulators of the drought response, and quadruple Gmlhy/cca1 (more simply named quadruple lhy) loss of function mutants exhibit improved drought tolerance, likely through alteration of ABA signaling [170]. Thus, it seems possible that clock components such as LHY may provide multiple targets for improvement of crop stress responses, through either harnessing natural variation or via genome editing.
2.2. Tomato
Tomatoes (Solanum lycopersicon) offer an excellent example of clock function being subject to selection during domestication and improvement. The wild progenitor of the tomato (Solanum pimpinellifolium) originated in the Andes of Ecuador and Peru. Domestication occurred in two stages, first in South America and later in Central America [171]. The first step, in South America, was associated with the earliest domesticated Ecuadorian cherry tomatoes showing a delayed circadian phase relative to wild species [172]. The causal gene was shown to encode a phytochrome A–associated F-box protein homologous to Arabidopsis EID1, which is a negatively acting component of phytochrome A signaling [173]. The delayed phase phenotype is associated with a three bp deletion removing a conserved residue in the C terminus of the protein. Consistent with the cultivated EID1 allele and altered circadian phase conferring an adaptive advantage, plants carrying the cultivated allele of EID1 were shorter, flowered later and had higher chlorophyll content than those bearing the wild allele and the differences in chlorophyll content occurred specifically under long days [172]. The second step in domestication resulted in lengthened period resulting from a partial deletion of LNK2 that, as described above, encodes a transcriptional coactivator in the clock [172,174]. In Arabidopsis, mutational disruption of LNK2 function prevents transcriptional activation of PRR5 by RVE8 and results in long circadian period [59]. LNK2, like EID1, contributes to light signaling to the circadian clock [59,175]. Both EID1 and LNK2 are located in chromosomal regions that exhibit very low genetic diversity, consistent with positive selection during tomato domestication or improvement. It seems reasonable to hypothesize that the slower and delayed phase clock represents an adaptation to long photoperiods encountered at higher latitudes, which may in turn enhance overall crop performance [172,174].
2.3. Sugar Beet
Sugar beet (Beta vulgaris) belongs to the Amaranthaceae family, whose lineage diverged from that of Arabidopsis shortly after the monocot–dicot split ~140 million years ago. Domestication of sugar beet occurred only within the past 200 years and domestication entailed a switch from an annual to a biennial habit with a requirement for vernalization, because bolting and flowering in the first year is associated with a drastic reduction in yield. Four loci, B, B2, B3, and B5 have been identified as controlling bolting [176,177]. Of these, B has been shown to be a PRR3/PRR7 homolog, BOLTING TIME CONTROL 1 (BvBTC1) [178]. BvBTC1 is necessary for flowering and mediates the response to both long days and vernalization through regulation of BvFT genes [179] and partial loss of function alleles of BvBTC1 have been selected during domestication [178]. B2 encodes a second transcription factor, B-BOX TYPE ZINC FINGER 19 (BvBBXC19), with both proteins necessary for CO-like activity and induction of the FT genes [180,181]. However, there is no experimental evidence establishing roles for these loci in circadian clock function in sugar beet.
2.4. Monocot Clocks
The examples discussed to this point have all been from eudicots, but similar pressures have been associated with range expansion among monocots. Cultivated grasses such as rice (Oryza sativa), maize (Zea mays), wheat (T. aestivum), and barley (H. vulgare) are enormously important crops. Although monocots have long been known to exhibit circadian regulation of gene expression, e.g., [182,183], the study of the molecular basis of monocot clocks has lagged behind that of eudicots. Phylogenetic analysis of the PRR and CCA1/LHY gene families shows that circadian clocks composed of multiple interlocked feedback loops evolved prior to the divergence of monocots and eudicots [184,185,186]. Orthologs of Arabidopsis clock genes have been identified in monocots such as rice [187,188], Lemna [189,190], other duckweeds [191], and barley [192,193,194,195].
Rice has a set of highly conserved clock-associated genes, including OsCCA1, OsLUX(PCL), five OsPRR genes including OsTOC1(PRR1), OsZTLs, and OsGI [187,188]. Similarly, barley has multiple orthologs to Arabidopsis clock genes, including HvCCA1, HvRVE7 and HvRVE8, five HvPRR genes including HvTOC1(PRR1), HvGI, and HvLUX(PCL) [192,193,194,195].
In functional studies, loss of function of OsGI affected diurnal expression of 75% of all tested genes and conferred reduced seasonal adaptability in field-grown rice [196]. Rice orthologs of TOC1 and PRR7 partially complemented the corresponding Arabidopsis toc1 and prr7 mutants, which is consistent with the function of these proteins being conserved between monocots and Arabidopsis [197]. Recently it has been established that OsPRR73 is involved in a feedback loop of the rice clock and connects the circadian clock to the photoperiodic flowering pathway by binding to the Ehd1 promoter as well as to the OsLHY promoter [198]. Loss of function of OsPRR73 results in early heading under LD but not SD, whereas overexpression results in late heading under both LD and SD [198]. Misexpression of OsPRR73 perturbs the expression of a number of clock genes.
In Lemna gibba, overexpression or RNAi-mediated downregulation of several genes (LgLHYH1, LgLHYH2, LgGIH1, and LgELF3H1) tested their roles in the circadian system. Overexpression of each gene and RNAi knock-downs of each of the genes except LgLHYH2 disrupted the bioluminescence rhythms of clock reporter constructs [190].
There are several examples in which mutations of barley orthologs of Arabidopsis clock genes disrupt clock function. HvPHYC carrying a mutation in a conserved region of the GAF domain is a candidate underlying the early maturity 5 (eam5) locus in barley [199]. HvPHYC interacts with Ppd-H1 to accelerate flowering under noninductive short days. In addition, expression of a number of clock genes is perturbed in eam5 mutants. Other barley early flowering mutants (early maturity; eam), eam8 (allelic to mat-a.8) and eam10, carry mutations in HvELF3 and HvLUX1, respectively [193,194,200]. As discussed below, these mutations facilitate adaption to short growing season, extending cultivation northward. Loss of function of HvELF3 causes an up-regulation of Ppd-H1 and the downstream HvFT1 under noninductive SD conditions and results in severe perturbation of the expression of circadian clock genes [193]. Both the Hvelf3 and Hvlux1 loss of function mutants are arrhythmic and have lost circadian transcriptome oscillations under constant conditions [195]. eam7, another early flowering mutant whose identity has not yet been resolved, also exhibited severely perturbed clock function [195].
Collectively, these functional observations in Lemna, barley, and rice indicate that the structure of the circadian clock is likely to be conserved between monocots and eudicots.
2.5. Rice
Rice originated in the subtropics but is now cultivated over an expanded latitudinal range north to ~53°N and south to the tropics. Weakened photoperiod sensitivity is a critical factor for adaptation of rice to high-latitude regions. Combinations of weak alleles of Ghd7, Hd1, PhyB, as well as other important flowering determinants, act additively to reduce photoperiod sensitivity to enable rice cultivation in high latitude areas [201,202,203]. Similarly, allelic variants of flowering activation genes such as Ehd4 and RFT1 have also contributed to adaptation of rice to higher latitudes [204,205]. Given the importance of the circadian clock in the regulation of flowering time [70,76], discussed above, it seems quite possible to allelic variants of clock genes may contribute to improved performance and further range expansion.
2.6. Barley
Genetic variation in photoperiod response was also crucial for the successful expansion of barley cultivation from its origin in the Fertile Crescent to northern latitudes. A number of photoperiod insensitive eam loci, some of which correspond to clock genes, were mentioned above. However, the major determinant of the barley photoperiod response is the HvPPR37 gene, Ppd-H1. Reduced photoperiod responsiveness of the ppd-H1 mutant is highly advantageous in spring-sown varieties and results in late flowering, which can be explained by altered circadian expression of the photoperiod pathway genes HvCO1 and HvCO2, delaying their diurnal expression peaks into the dark and so preventing accumulation of CO protein. This reduced CO accumulation results in reduced expression of its downstream target, HvFT (Vrn-H3) [206].
PRR genes feature prominently among regulators of flowering time (heading date) in the grasses. However, the evolution of the PRR family has differed in the grasses from that in the eudicots [185], and at least in some cases it seems that the genes regulating flowering time are distinct from those regulating the circadian clock. The ppd-H1 mutation does not perturb circadian clock gene expression, suggesting that Ppd-H1(PRR37) does not contribute to circadian clock function [192]. This would be consistent with sub-functionalization among the barley PRR gene family in which Ppd-H1(PRR37) lost clock function, which was presumably retained by HvPRR73. Alternatively, functional redundancy between Ppd-H1(PRR37) and HvPRR73 might prevent detection of a clock defect in the ppd-H1 single mutant, although double mutant ppd-H1 Hvprr73 might exhibit a more extreme phenotypic defect than the Hvprr73 single mutant. For example, neither the fkf1 not the lkp2 single mutants of Arabidopsis exhibit clock defects, but both mutations enhance the long-period phenotype of the ztl mutant [207]. Ppd-H1(PRR37) is also a major determinant of leaf size in barley, likely via its induction of the MADS BOX genes BM3 and BM8 in the leaf [208].
3. Concluding Remarks
The importance of flowering time to crop performance has made it a critical target of efforts to expand latitudinal zones of cultivation. The centrality of the circadian clock to photoperiodic flowering has meant that allelic variation of circadian clock genes has contributed to range expansion in many crop species. However, the circadian clock contributes to plant performance in many ways beyond photoperiodic flowering. The circadian clock contributes to the regulation of circadian of growth and metabolism, as well as to abiotic and biotic stress responses [12,13,209]. As a consequence, there are many routes by which the circadian clock contributes to plant fitness and crop performance [11,210,211].
In Arabidopsis, a clock that resonates with the environmental daylength enhances photosynthesis and biomass accumulation [212]. An altered circadian clock in a new coffee clone has been correlated with higher photosynthesis efficiency and improved agronomic performance, although the mechanistic basis by which altered clock function does so remains uncertain [213]. Ni et al. (2009) showed that subtle changes in the temporal expression pattern of CCA1 contributed to growth vigor in Arabidopsis hybrids and allopolyploids [214]. Epigenetic changes in circadian-related genes, including altered methylation of the promoter of the parental CCA1 allele, have been shown to lead to biomass heterosis in Arabidopsis hybrids [215]. This has been extended from Arabidopsis to crops; early activation of CCA1-binding targets in maize hybrids promoted photosynthesis and biomass heterosis [216].
It seems likely that optimizing circadian function will continue to offer opportunities to enhance crop productivity, particularly in crops grown over broad latitudinal ranges. This need will be exacerbated by global warming which necessitates a poleward migration of zones of cultivation [217]. Efforts towards crop improvement to date have focused on standing allelic variation. However, improvements in genome editing now offer the potential to make precise targeted changes to the genome that are independent of standing variation [218]. Of course, such efforts towards genome editing need to be informed by greater understanding of the mechanistic means by which the clock enhances plant performance. The pervasive nature of clock influence mandates an informed approach to clock manipulation lest such efforts prove counterproductive. This offers a compelling rationale for the continued study of not only how the plant circadian clock keeps time, but also of how the plant uses that time information to regulate output pathways in a temporally dynamic fashion.
Funding
This review was funded by the National Science Foundation, grant number IOS 1547796 and by the Rural Development Administration, Republic of Korea Next Generation BioGreen 21, grant number SSAC PJ01327306.
Acknowledgments
I apologize to those colleagues whose work was not discussed here due to space constraints.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the writing of the manuscript.
References
- Doebley, J.F.; Gaut, B.S.; Smith, B.D. The molecular genetics of crop domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, J.; Sikora, M.; Garud, N.; Flowers, J.M.; Rubinstein, S.; Reynolds, A.; Huang, P.; Jackson, S.; Schaal, B.A.; Bustamante, C.D.; et al. Molecular evidence for a single evolutionary origin of domesticated rice. Proc. Natl. Acad. Sci. USA 2011, 108, 8351–8356. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, Y.; Vigouroux, Y.; Goodman, M.M.; Sanchez, G.J.; Buckler, E.; Doebley, J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc. Natl. Acad. Sci. USA 2002, 99, 6080–6084. [Google Scholar] [CrossRef] [Green Version]
- Purugganan, M.D.; Fuller, D.Q. The nature of selection during plant domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Purugganan, M.D. Evolutionary insights into the nature of plant domestication. Curr. Biol. 2019, 29, R705–R714. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K. Das Domestikationssyndrom. Kulturpflanze 1984, 32, 11–34. [Google Scholar] [CrossRef]
- Eyre-Walker, A.; Gaut, R.L.; Hilton, H.; Feldman, D.L.; Gaut, B.S. Investigation of the bottleneck leading to the domestication of maize. Proc. Natl. Acad. Sci. USA 1998, 95, 4441–4446. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.I.; Bi, I.V.; Schroeder, S.G.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S. The effects of artificial selection on the maize genome. Science 2005, 308, 1310–1314. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Bendix, C.; Marshall, C.M.; Harmon, F.G. Circadian clock genes universally control key agricultural traits. Mol. Plant 2015, 8, 1135–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerushalmi, S.; Green, R.M. Evidence for the adaptive significance of circadian rhythms. Ecol. Lett. 2009, 12, 970–981. [Google Scholar] [CrossRef]
- Greenham, K.; McClung, C.R. Integrating circadian dynamics with physiological processes in plants. Nat. Rev. Genet. 2015, 16, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.J. The intracellular dynamics of circadian clocks reach for the light of ecology and evolution. Annu. Rev. Plant Biol. 2016, 67, 595–618. [Google Scholar] [CrossRef]
- Edgar, R.S.; Green, E.W.; Zhao, Y.; van Ooijen, G.; Olmedo, M.; Qin, X.; Xu, Y.; Pan, M.; Valekunja, U.K.; Feeney, K.A.; et al. Peroxiredoxins are conserved markers of circadian rhythms. Nature 2012, 485, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Young, M.W.; Kay, S.A. Time zones: A comparative genetics of circadian clocks. Nat. Rev. Genet. 2001, 2, 702–715. [Google Scholar] [CrossRef]
- Lou, P.; Wu, J.; Cheng, F.; Cressman, L.G.; Wang, X.; McClung, C.R. Preferential retention of circadian clock genes during diploidization following whole genome triplication in Brassica rapa. Plant Cell 2012, 24, 2415–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClung, C.R. The plant circadian oscillator. Biology 2019, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, N. The transcriptional network in the Arabidopsis circadian clock system. Genes 2020, 11, 1284. [Google Scholar] [CrossRef]
- James, A.B.; Syed, N.H.; Bordage, S.; Marshall, J.; Nimmo, G.A.; Jenkins, G.I.; Herzy, P.; Brown, J.W.S.; Nimmo, H.G. Alternative splicing mediates responses of the Arabidopsis circadian clock to temperature changes. Plant Cell 2012, 24, 961–981. [Google Scholar] [CrossRef] [Green Version]
- Filichkin, S.A.; Priest, H.D.; Givan, S.A.; Shen, R.; Bryant, D.W.; Fox, S.E.; Wong, W.-K.; Mockler, T.C. Genome-wide mapping of alternative splicing in Arabidopsis thaliana. Genome Res. 2010, 20, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Romanowski, A.; Schlaen, R.G.; Perez-Santangelo, S.; Mancini, E.; Yanovsky, M.J. Global transcriptome analysis reveals circadian control of splicing events in Arabidopsis thaliana. Plant J. 2020, 103, 889–902. [Google Scholar] [CrossRef]
- Sanchez, S.E.; Petrillo, E.; Beckwith, E.J.; Zhang, X.; Rugnone, M.L.; Hernando, C.E.; Cuevas, J.C.; Godoy Herz, M.A.; Depetris-Chauvin, A.; Simpson, C.G.; et al. A methyl transferase links the circadian clock to the regulation of alternative splicing. Nature 2010, 468, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.; Song, H.-R.; Lutz, K.; Kerstetter, R.A.; Michael, T.P.; McClung, C.R. Type II Protein Arginine Methyltransferase PRMT5 is required for circadian period determination in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 21211–21216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.A.; Williams, B.A.; McNicol, J.; Simpson, C.G.; Brown, J.W.S.; Harmer, S.L. Mutation of Arabidopsis SPLICEOSOMAL TIMEKEEPER LOCUS1 causes circadian clock defects. Plant Cell 2012, 24, 4066–4082. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wu, F.; Xie, Q.; Wang, H.; Wang, Y.; Yue, Y.; Gahura, O.; Ma, S.; Liu, L.; Cao, Y.; et al. SKIP is a component of the spliceosome linking alternative splicing and the circadian clock in Arabidopsis. Plant Cell 2012, 24, 3278–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, P.J.; Mas, P. Multiple layers of posttranslational regulation refine circadian clock activity in Arabidopsis. Plant Cell 2014, 26, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Mateos, J.L.; de Leone, M.J.; Torchio, J.; Reichel, M.; Staiger, D. Beyond transcription—Fine tuning of circadian timekeeping by posttranscriptional regulation. Genes 2018, 9, 616. [Google Scholar] [CrossRef] [Green Version]
- Más, P.; Kim, W.-Y.; Somers, D.E.; Kay, S.A. Targeted degradation of TOC1 by ZTL modulates circadian function in Arabidopsis thaliana. Nature 2003, 426, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, S.; Wang, L.; Han, L.; Suh, S.S.; Salomé, P.A.; McClung, C.R.; Somers, D.E. Post-translational regulation of the circadian clock through selective proteolysis and phosphorylation of pseudo-response regulator proteins. J. Biol. Chem. 2008, 283, 23073–23083. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Fujiwara, S.; Somers, D.E. PRR5 regulates phosphorylation, nuclear import and subnuclear localization of TOC1 in the Arabidopsis circadian clock. EMBO J. 2010, 29, 1903–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.-s.; Kim, W.Y.; Fujiwara, S.; Kim, J.; Cha, J.-Y.; Park, J.H.; Lee, S.Y.; Somers, D.E. HSP90 functions in the circadian clock through stabilization of the client F-box protein ZEITLUPE. Proc. Natl. Acad. Sci. USA 2011, 108, 16843–16848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Geng, R.; Gallenstein, R.A.; Somers, D.E. The F-box protein ZEITLUPE controls stability and nucleocytoplasmic partitioning of GIGANTEA. Development 2013, 140, 4060–4069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Han, S.; Yeom, M.; Kim, H.; Lim, J.; Cha, J.-Y.; Kim, W.-Y.; Somers, D.E.; Putterill, J.; Nam, H.G.; et al. Balanced nucleocytosolic partitioning defines a spatial network to coordinate circadian physiology in plants. Dev. Cell 2013, 26, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.-Y.; Kim, J.; Kim, T.-S.; Zeng, Q.; Wang, L.; Lee, S.Y.; Kim, W.-Y.; Somers, D.E. GIGANTEA is a co-chaperone which facilitates maturation of ZEITLUPE in the Arabidopsis circadian clock. Nat. Commun. 2017, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Romanowski, A.; Yanovsky, M.J. Circadian rhythms and post-transcriptional regulation in higher plants. Front. Plant Sci. 2015, 6, 437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Ma, D.; Lu, S.X.; Hu, X.; Huang, R.; Liang, T.; Xu, T.; Tobin, E.M.; Liu, H. Blue light- and low temperature-regulated COR27 and COR28 play roles in the Arabidopsis circadian clock. Plant Cell 2016, 28, 2755–2769. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Cui, X.; Zhao, C.; Shi, L.; Zhang, G.; Sun, F.; Cao, X.; Yuan, L.; Xie, Q.; Xu, X. COR27 and COR28 encode nighttime repressors integrating Arabidopsis circadian clock and cold response. J. Int. Plant Biol. 2017, 59, 78–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, A.J.; Carré, I.A.; Strayer, C.A.; Chua, N.-H.; Kay, S.A. Circadian clock mutants in Arabidopsis identified by luciferase imaging. Science 1995, 267, 1161–1163. [Google Scholar] [CrossRef] [PubMed]
- Strayer, C.; Oyama, T.; Schultz, T.F.; Raman, R.; Somers, D.E.; Más, P.; Panda, S.; Kreps, J.A.; Kay, S.A. Cloning of the Arabidopsis clock gene TOC1, an autoregulatory response regulator homolog. Science 2000, 289, 768–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Más, P.; Alabadí, D.; Yanovsky, M.J.; Oyama, T.; Kay, S.A. Dual role of TOC1 in the control of circadian and photomorphogenic responses in Arabidopsis. Plant Cell 2003, 15, 223–236. [Google Scholar] [CrossRef] [Green Version]
- Gendron, J.M.; Pruneda-Paz, J.L.; Doherty, C.J.; Gross, A.M.; Kang, S.E.; Kay, S.A. Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc. Natl. Acad. Sci. USA 2012, 109, 3167–3172. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, R.; Ramsay, N.; Samach, A.; Corden, S.; Putterill, J.; Carré, I.A.; Coupland, G. The late elongated hypocotyl mutation of Arabidopsis disrupts circadian rhythms and the photoperiodic control of flowering. Cell 1998, 93, 1219–1229. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Tobin, E.M. Constitutive expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) gene disrupts circadian rhythms and suppresses its own expression. Cell 1998, 93, 1207–1217. [Google Scholar] [CrossRef] [Green Version]
- Alabadí, D.; Yanovsky, M.J.; Más, P.; Harmer, S.L.; Kay, S.A. Critical role for CCA1 and LHY in maintaining circadian rhythmicity in Arabidopsis. Curr. Biol. 2002, 12, 757–761. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, T.; Wheatley, K.; Hanzawa, Y.; Wright, L.; Mizoguchi, M.; Song, H.-R.; Carré, I.A.; Coupland, G. LHY and CCA1 are partially redundant genes required to maintain circadian rhythms in Arabidopsis. Dev. Cell 2002, 2, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Alabadí, D.; Oyama, T.; Yanovsky, M.J.; Harmon, F.G.; Más, P.; Kay, S.A. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 2001, 293, 880–883. [Google Scholar] [CrossRef] [PubMed]
- Matsushika, A.; Makino, S.; Kojima, M.; Mizuno, T. Circadian waves of expression of the APRR1/TOC1 family of pseudo-response regulators in Arabidopsis thaliana: Insight into the plant circadian clock. Plant Cell Physiol. 2000, 41, 1002–1012. [Google Scholar] [CrossRef] [Green Version]
- Nakamichi, N.; Kita, M.; Ito, S.; Sato, E.; Yamashino, T.; Mizuno, T. PSEUDO-RESPONSE REGULATORS, PRR9, PRR7 and PRR5, together play essential roles close to the circadian clock of Arabidopsis thaliana. Plant Cell Physiol. 2005, 46, 686–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, N.; Kiba, T.; Henriques, R.; Mizuno, T.; Chua, N.-H.; Sakakibara, H. PSEUDO-RESPONSE REGULATORS 9, 7 and 5 are transcriptional repressors in the Arabidopsis circadian clock. Plant Cell 2010, 22, 594–605. [Google Scholar] [CrossRef] [Green Version]
- Pruneda-Paz, J.L.; Breton, G.; Para, A.; Kay, S.A. A functional genomics approach reveals CHE as a component of the Arabidopsis circadian clock. Science 2009, 323, 1481–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Kim, J.; Somers, D.E. Transcriptional corepressor TOPLESS complexes with pseudoresponse regulator proteins and histone deacetylases to regulate circadian transcription. Proc. Natl. Acad. Sci. USA 2013, 110, 761–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nusinow, D.A.; Helfer, A.; Hamilton, E.E.; King, J.J.; Imaizumi, T.; Schultz, T.F.; Farre, E.M.; Kay, S.A. The ELF4-ELF3-LUX complex links the circadian clock to diurnal control of hypocotyl growth. Nature 2011, 475, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Herrero, E.; Kolmos, E.; Bujdoso, N.; Yuan, Y.; Wang, M.; Berns, M.C.; Uhlworm, H.; Coupland, G.; Saini, R.; Jaskolski, M.; et al. EARLY FLOWERING4 recruitment of EARLY FLOWERING3 in the nucleus sustains the Arabidopsis circadian clock. Plant Cell 2012, 24, 428–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, S.; Wei, X.; Pei, L.; Thompson, R.L.; Liu, Y.; Heard, J.E.; Ruff, T.G.; Beachy, R.N. BROTHER OF LUX ARRHYTHMO is a component of the Arabidopsis circadian clock. Plant Cell 2011, 23, 961–972. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.-F.; Tsai, H.-L.; Joanito, I.; Wu, Y.-C.; Chang, C.-W.; Li, Y.-H.; Wang, Y.; Hong, J.C.; Chu, J.-W.; Hsu, C.-P.; et al. LWD-TCP complex activates the morning gene CCA1 in Arabidopsis. Nat. Commun. 2016, 7, 13181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mockler, T.C.; Michael, T.P.; Priest, H.D.; Shen, R.; Sullivan, C.M.; Givan, S.A.; McEntee, C.; Kay, S.A.; Chory, J. The Diurnal Project: Diurnal and circadian expression profiling, model-based pattern matching, and promoter analysis. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Rawat, R.; Takahashi, N.; Hsu, P.Y.; Jones, M.A.; Schwartz, J.; Salemi, M.R.; Phinney, B.S.; Harmer, S.L. REVEILLE8 and PSEUDO-REPONSE REGULATOR5 form a negative feedback loop within the Arabidopsis circadian clock. PLoS Genet. 2011, 7, e1001350. [Google Scholar] [CrossRef]
- Hsu, P.Y.; Devisetty, U.K.; Harmer, S.L. Accurate timekeeping is controlled by a cycling activator in Arabidopsis. ELife 2013, 2, e00473. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Wang, P.; Liu, X.; Yuan, L.; Wang, L.; Zhang, C.; Li, Y.; Xing, H.; Zhi, L.; Yue, Z.; et al. LNK1 and LNK2 are transcriptional coactivators in the Arabidopsis circadian oscillator. Plant Cell 2014, 26, 2843–2857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- José de Leone, M.; Esteban, H.C.; Romanowski, A.; García-Hourquet, M.; Careno, D.; Casal, J.; Rugnone, M.; Mora-García, S.; Yanovsky, M.J. The LNK gene family: At the crossroad between light signaling and the circadian clock. Genes 2019, 10, 2. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Siddiqui, H.; Teng, Y.; Lin, R.; Wan, X.-y.; Li, J.; Lau, O.-S.; Ouyang, X.; Dai, M.; Wan, J.; et al. Coordinated transcriptional regulation underlying the circadian clock in Arabidopsis. Nat. Cell Biol. 2011, 13, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, M.; Li, G.; Yuan, L.; Xie, Y.; Wei, H.; Ma, X.; Li, Q.; Devlin, P.F.; Xu, X.; et al. Transcription factors FHY3 and FAR1 regulate light-induced CIRCADIAN CLOCK ASSOCIATED1 gene expression in Arabidopsis. Plant Cell 2020, 32, 1464–1478. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, H.; Lin, F.; Zhao, X.; Jiang, Y.; Xu, D.; Deng, X.W. COLD-REGULATED GENE27 integrates signals from light and the circadian clock to promote hypocotyl growth in Arabidopsis. Plant Cell 2020, 32, 3155–3169. [Google Scholar] [CrossRef] [PubMed]
- Perales, M.; Más, P. A functional link between rhythmic changes in chromatin structure and the Arabidopsis biological clock. Plant Cell 2007, 19, 2111–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.; Mas, P.; Seo, P.J. The EC-HDA9 complex rhythmically regulates histone acetylation at the TOC1 promoter in Arabidopsis. Commun. Biol. 2019, 2. [Google Scholar] [CrossRef]
- Park, H.J.; Baek, D.; Cha, J.-Y.; Liao, X.; Kang, S.-H.; McClung, C.R.; Lee, S.Y.; Yun, D.-J.; Kim, W.-Y. HOS15 interacts with the histone deactetylase HDA9 and the Evening Complex to epigenetically regulate the floral activator GIGANTEA. Plant Cell 2019, 31, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Maric, A.; Mas, P. Chromatin dynamics and transcriptional control of circadian rhythms in Arabidopsis. Genes 2020, 11, 1170. [Google Scholar] [CrossRef]
- Kende, H.; Zeevart, J.A.D. Anton Lang. In Biographical Memoirs; The National Academies Press: Washington, DC, USA, 1998; Volume 74, pp. 49–61. [Google Scholar]
- Romera-Branchat, M.; Andres, F.; Coupland, G. Flowering responses to seasonal cues: What’s new? Curr. Op. Plant Biol. 2014, 21, 120–127. [Google Scholar] [CrossRef]
- Song, Y.H.; Shim, J.S.; Kinmonth-Schultz, H.A.; Imaizumi, T. Photoperiodic flowering: Time measurement mechanisms in leaves. Annu. Rev. Plant Biol. 2015, 66, 441–464. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Z.; Liu, H.; Liu, B.; Liu, X.; Lin, C. Blue light-dependent interaction of CRY2 with SPA1 regulates COP1 activity and floral initiation in Arabidopsis. Curr. Biol. 2011, 21, 841–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarid-Krebs, L.; Panigrahi, K.C.S.; Fornara, F.; Takahashi, Y.; Hayama, R.; Jang, S.; Tilmes, V.; Valverde, F.; Coupland, G. Phosphorylation of CONSTANS and its COP1-dependent degradation during photoperiodic flowering of Arabidopsis. Plant J. 2015, 84, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayama, R.; Sarid-Krebs, L.; Richter, R.; Fernandez, V.; Jang, S.; Coupland, G. PSEUDO RESPONSE REGULATORs stabilize CONSTANS protein to promote flowering in response to day length. EMBO J. 2017, 36, 904–918. [Google Scholar] [CrossRef]
- Liu, H.; Yu, X.; Li, K.; Klejnot, J.; Yang, H.; Lisiero, D.; Lin, C. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science 2008, 322, 1535–1538. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, X.; Li, K.; Liu, H.; Lin, C. Multiple bHLH proteins form heterodimers to mediate CRY2-dependent regulation of flowering-time in Arabidopsis. PLoS Genet. 2013, 9, e1003861. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Zhu, S.; Cui, S.; Hou, H.; Wu, H.; Hao, B.; Cai, L.; Xu, Z.; Liu, L.; Jiang, L.; et al. Transcriptional and post-transcriptional regulation of heading date in rice. New Phytol. 2021, 229. [Google Scholar] [CrossRef]
- Izawa, T.; Oikawa, T.; Sugiyama, N.; Tanisaka, T.; Yano, M.; Shimamoto, K. Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice. Genes Dev. 2002, 16, 2006–2020. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, R.; Aoki, M.; Kurotani, K.-i.; Yokoi, S.; Shinomura, T.; Takano, M.; Shimamoto, K. Phytochrome B regulates Heading date 1 (Hd1)-mediated expression of rice florigen Hd3a and critical day length in rice. Mol. Genet. Genom. 2011, 285, 461–470. [Google Scholar] [CrossRef]
- Doi, K.; Izawa, T.; Fuse, T.; Yamanouchi, U.; Kubo, T.; Shimatani, Z.; Yano, M.; Yoshimura, A. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev. 2004, 18, 926–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Nonoue, Y.; Yano, M.; Izawa, T. A pair of floral regulators sets critical day length for Hd3a florigen expression in rice. Nat. Genet. 2010, 42, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Osugi, A.; Itoh, H.; Ikeda-Kawakatsu, K.; Takano, M.; Izawa, T. Molecular dissection of the roles of phytochrome in photoperiodic flowering in rice. Plant Physiol. 2011, 157, 1128–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, K.; Ogiso-Tanaka, E.; Hori, K.; Ebana, K.; Ando, T.; Yano, M. Natural variation in Hd17, a homolog of Arabidopsis ELF3 that is involved in rice photoperiodic flowering. Plant Cell Physiol. 2012, 53, 709–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, H.; Ogiso-Tanaka, E.; Okumoto, Y.; Yoshitake, Y.; Izumi, H.; Yokoo, T.; Matsubara, K.; Hori, K.; Yano, M.; Inoue, H.; et al. Ef7 encodes an ELF3-like protein and promotes rice flowering by negatively regulating the floral repressor gene Ghd7 under both short- and long-day conditions. Plant Cell Physiol. 2012, 53, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Huang, X.; Ouyang, X.; Chen, W.; Du, A.; Zhu, L.; Wang, S.; Deng, X.W.; Li, S. OsELF3-1, an ortholog of Arabidopsis EARLY FLOWERING 3, regulates rice circadian rhythm and photoperiodic flowering. PLoS ONE 2012, 7, e43705. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Qu, X.; Zhou, Y.; Song, G.; Abiri, N.; Xiao, Y.; Liang, F.; Jiang, D.; Hu, Z.; Yang, D. OsPRR37 confers an expanded regulation of the diurnal rhythms of the transcriptome and photoperiodic flowering pathways in rice. Plant Cell Environ. 2018, 41, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Hut, R.A.; Paolucci, S.; Dor, R.; Kyriacou, C.P.; Daan, S. Latitudinal clines: An evolutionary view on biological rhythms. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130433. [Google Scholar] [CrossRef] [Green Version]
- Majercak, J.; Sidote, D.; Hardin, P.E.; Edery, I. How a circadian clock adapts to seasonal decreases in temperature and day length. Neuron 1999, 24, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.; Kyriacou, C.P. Functional and evolutionary implications of natural variation in clock genes. Curr. Opin. Neurobiol. 1998, 8, 659–664. [Google Scholar] [CrossRef]
- Rosato, E.; Piccin, A.; Kyriacou, C.P. Circadian rhythms: From behaviour to molecules. BioEssays 1997, 19, 1075–1082. [Google Scholar] [CrossRef]
- Sawyer, L.A.; Hennessy, J.M.; Peixoto, A.A.; Rosato, E.; Parkinson, H.; Costa, R.; Kyriacou, C.P. Natural variation in a Drosophila clock gene and temperature compensation. Science 1997, 278, 2117–2120. [Google Scholar] [CrossRef]
- Johnsen, A.; Fidler, A.E.; Kuhn, S.; Carter, K.L.; Hoffmann, A.; Barr, I.R.; Biard, C.; Charmantier, A.; Eens, M.; Korsten, P.; et al. Avian Clock gene polymorphism: Evidence for a latitudinal cline in allele frequencies. Mol. Ecol. 2007, 16, 4867–4880. [Google Scholar] [CrossRef]
- O’Malley, K.G.; Banks, M.A. A latitudinal cline in the Chinook salmon (Oncorhynchus tshawytscha) Clock gene: Evidence for selection on PolyQ length variants. Proc. R. Soc. Lond. B 2008, 275, 2813–2822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, T.P.; Salomé, P.A.; Yu, H.J.; Spencer, T.R.; Sharp, E.L.; Alonso, J.M.; Ecker, J.R.; McClung, C.R. Enhanced fitness conferred by naturally occurring variation in the circadian clock. Science 2003, 302, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.D.; Lynn, J.R.; Gyula, P.; Nagy, F.; Millar, A.J. Natural allelic variation in the temperature compensation mechanisms of the Arabidopsis thaliana circadian clock. Genetics 2005, 170, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenham, K.; Lou, P.; Puzey, J.R.; Kumar, G.; Arnevik, C.; Farid, H.; Willis, J.H.; McClung, C.R. Geographic variation of plant circadian clock function in natural and agricultural settings. J. Biol. Rhythms 2017, 32, 26–34. [Google Scholar] [CrossRef]
- Kim, M.Y.; Van, K.; Kang, Y.J.; Kim, K.; Lee, S.-H. Tracing soybean domestication history: From nucleotide to genome. Breed. Sci. 2012, 61, 445–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedivy, E.J.; Wu, F.; Hanzawa, Y. Soybean domestication: The origin, genetic architecture and molecular bases. New Phytol. 2017, 214, 539–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.; Liu, B.; Xia, Z.; Sato, S.; Kim, B.M.; Watanabe, S.; Yamada, T.; Tabata, S.; Kanazawa, A.; Harada, K.; et al. Two coordinately regulated homologs of FLOWERING LOCUS T are involved in the control of photoperiodic flowering in soybean. Plant Physiol. 2010, 154, 1220–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cober, E.R.; Morrison, M.J. Regulation of seed yield and agronomic characters by photoperiod sensitivity and growth habit genes in soybean. Theor. Appl. Genet. 2010, 120, 1005–1012. [Google Scholar] [CrossRef]
- Kim, M.Y.; Shin, J.H.; Kang, Y.J.; Shim, S.R.; Lee, S.-H. Divergence of flowering genes in soybean. J. Biosci. 2012, 37, 857–870. [Google Scholar] [CrossRef]
- Lin, X.; Liu, B.; Weller, J.L.; Abe, J.; Kong, F. Molecular mechanisms for the photoperiodic regulation of flowering in soybean. J. Integr. Plant Biol. 2021. [Google Scholar] [CrossRef]
- Cao, D.; Takeshima, R.; Zhao, C.; Liu, B.; Abe, J.; Kong, F. Molecular mechanisms of flowering under long days and stem growth habit in soybean. J. Exp. Bot. 2017, 68, 1873–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanfar, B.; Molnar, S.J.; Charette, M.; Schoenrock, A.; Dehne, F.; Golshani, A.; Belzile, F.; Cober, E.R. Mapping and identification of a potential candidate gene for a novel maturity locus, E10, in soybean. Theor. Appl. Genet. 2017, 130, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Nan, H.; Chen, L.; Fang, C.; Zhang, H.; Su, T.; Li, S.; Cheng, Q.; Dong, L.; Liu, B.; et al. A new dominant locus, E11, controls early flowering time and maturity in soybean. Mol. Breed. 2019, 39, 70. [Google Scholar] [CrossRef]
- Watanabe, S.; Xia, Z.; Hideshima, R.; Tsubokura, Y.; Sato, S.; Yamanaka, N.; Takahashi, R.; Anai, T.; Tabata, S.; Kitamura, K.; et al. A map-based cloning strategy employing a residual heterozygous line reveals that the GIGANTEA gene Is Involved in soybean maturity and flowering. Genetics 2011, 188, 395–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Hideshima, R.; Xia, Z.; Tsubokura, Y.; Sato, S.; Nakamoto, Y.; Yamanaka, N.; Takahashi, R.; shimoto, M.; Anai, T.; et al. Map-based cloning of the gene associated with the soybean maturity locus E3. Genetics 2009, 182, 1251–1262. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Kanazawa, A.; Matsumura, H.; Takahashi, R.; Harada, K.; Abe, J. Genetic redundancy in soybean photoresponses associated with duplication of the phytochrome A gene. Genetics 2008, 180, 995–1007. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Kim, M.; Kang, Y.J.; Van, K.; Lee, Y.-H.; Srinives, P.; Yuan, D.; Lee, S.-H. QTL identification of flowering time at three different latitudes reveals homeologous genomic regions that control flowering in soybean. Theor. Appl. Genet. 2011, 123, 545–553. [Google Scholar] [CrossRef]
- Jiang, B.; Nan, H.; Gao, Y.; Tang, L.; Yue, Y.; Lu, S.; Ma, L.; Cao, D.; Sun, S.; Wang, J.; et al. Allelic combinations of soybean maturity loci E1, E2, E3 and E4 result in diversity of maturity and adaptation to different latitudes. PLoS ONE 2014, 9, e106042. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Jiang, B.; Wu, C.; Lu, W.; Hou, W.; Sun, S.; Yan, H.; Han, T. Maturity group classification and maturity locus genotyping of early-maturing soybean varieties from high-latitude cold regions. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Tsubokura, Y.; Watanabe, S.; Xia, Z.; Kanamori, H.; Yamagata, H.; Kaga, A.; Katayose, Y.; Abe, J.; Ishimoto, M.; Harada, K. Natural variation in the genes responsible for maturity loci E1, E2, E3 and E4 in soybean. Ann. Bot. 2014, 113, 429–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, H.; Lu, S.; Wang, Y.; Chen, X.; Ren, H.; Yang, J.; Cheng, W.; Zong, C.; Gu, H.; Qiu, H.; et al. Allelic variations at four major maturity E genes and transcriptional abundance of the E1 gene are associated with flowering time and maturity of soybean cultivars. PLoS ONE 2014, 9, e97636. [Google Scholar] [CrossRef] [Green Version]
- Zhai, H.; Lu, S.; Wu, H.; Zhang, Y.; Zhang, X.; Yang, J.; Wang, Y.; Yang, G.; Qiu, H.; Cui, T.; et al. Diurnal expression pattern, allelic variation, and association analysis reveal functional features of the E1 gene in control of photoperiodic flowering in soybean. PLoS ONE 2015, 10, e0135909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Li, Y.; Wang, J.; Srinives, P.; Nan, H.; Cao, D.; Wang, Y.; Li, J.; Li, X.; Fang, C.; et al. QTL mapping for flowering time in different latitude in soybean. Euphytica 2015, 206, 725–736. [Google Scholar] [CrossRef]
- Kurasch, A.K.; Hahn, V.; Leiser, W.L.; Vollmann, J.; Schori, A.; Bétrix, C.A.; Mayr, B.; Winkler, J.; Mechtler, K.; Aper, J.; et al. Identification of mega-environments in Europe and effect of allelic variation at maturity E loci on adaptation of European soybean. Plant Cell Environ. 2017, 40, 765–778. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Song, W.; Huang, X.; Zhou, J.; Zeng, H.; Sun, S.; Jia, H.; Li, W.; Zhou, X.; et al. Genetic variation of maturity groups and four E genes in the Chinese soybean mini core collection. PLoS ONE 2017, 12, e0172106. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Takeshima, R.; Zhu, J.; Xu, M.; Sato, M.; Watanabe, S.; Kanazawa, A.; Liu, B.; Kong, F.; Yamada, T.; et al. A recessive allele for delayed flowering at the soybean maturity locus E9 is a leaky allele of FT2a, a FLOWERING LOCUS T ortholog. BMC Plant Biol. 2016, 16, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Chen, L.; Liu, X.; Guo, C.; Sun, S.; Wu, C.; Jiang, B.; Han, T.; Hou, W. CRISPR/Cas9-mediated targeted mutagenesis of GmFT2a delays flowering time in soya bean. Plant Biotechnol. J. 2018, 16, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Wang, L.; Chen, L.; Wu, T.; Liu, L.; Sun, S.; Wu, C.; Yao, W.; Jiang, B.; Yuan, S.; et al. Mutagenesis of GmFT2a and GmFT5a mediated by CRISPR/Cas9 contributes for expanding the regional adaptability of soybean. Plant Biotechnol. J. 2020, 18, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Zhai, H.; Lü, S.; Liang, S.; Wu, H.; Zhang, X.; Liu, B.; Kong, F.; Yuan, X.; Li, J.; Xia, Z. GmFT4, a homolog of FLOWERING LOCUS T, is positively regulated by E1 and functions as a flowering repressor in soybean. PLoS ONE 2014, 9, e89030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhou, Z.; Liu, Y.; Liu, T.; Li, Q.; Ji, Y.; Li, C.; Fang, C.; Wang, M.; Wu, M.; et al. Functional evolution of phosphatidylethanolamine binding proteins in soybean and Arabidopsis. Plant Cell 2015, 27, 323–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickland, D.P.; Hanzawa, Y. The FLOWERING LOCUS T/TERMINAL FLOWER 1 gene family: Functional evolution and molecular mechanisms. Mol. Plant 2015, 8, 983–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Jiang, B.; Ma, L.; Zhang, S.; Zhai, H.; Xu, X.; Hou, W.; Xia, Z.; Wu, C.; Sun, S.; et al. Functional diversification of Flowering Locus T homologs in soybean: GmFT1a and GmFT2a/5a have opposite roles in controlling flowering and maturation. New Phytol. 2018, 217, 1335–1345. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Takeshima, R.; Harigai, K.; Xu, M.; Kong, F.; Liu, B.; Kanazawa, A.; Yamada, T.; Abe, J. Loss of function of the E1-Like-b ene associates with early flowering under long-day conditions in soybean. Front. Plant Sci. 2019, 9, 1867. [Google Scholar] [CrossRef]
- Xu, M.; Yamagishi, N.; Zhao, C.; Takeshima, R.; Kasai, M.; Watanabe, S.; Kanazawa, A.; Yoshikawa, N.; Liu, B.; Yamada, T.; et al. Soybean-specific E1 family of floral repressors controls night-break responses through down-regulation of FLOWERING LOCUS T orthologs. Plant Physiol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshima, R.; Hayashi, T.; Zhu, J.; Zhao, C.; Xu, M.; Yamaguchi, N.; Sayama, T.; Ishimoto, M.; Kong, L.; Shi, X.; et al. A soybean quantitative trait locus that promotes flowering under long days is identified as FT5a, a FLOWERING LOCUS T ortholog. J. Exp. Bot. 2016, 67, 5247–5258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koornneef, M.; Hanhart, C.J.; van der Veen, J.H. A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana. Mol. Gen. Genet. 1991, 229, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Dunford, R.P.; Griffiths, S.; Christodoulou, V.; Laurie, D.A. Characterisation of a barley (Hordeum vulgare L.) homologue of the Arabidopsis flowering time regulator GIGANTEA. Theor. Appl. Genet. 2005, 110, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Liu, M.S.; Li, J.R.; Guan, C.M.; Zhang, X.S. The wheat TaGI1, involved in photoperiodic flowering, encodes an Arabidopsis GI ortholog. Plant Mol. Biol. 2005, 58, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Knowles, C.L.; Schoor, J.K.V.; Liew, L.C.; Jones, S.E.; Lambert, M.J.M.; Weller, J.L. Pea LATE BLOOMER1 Is a GIGANTEA ortholog with roles in photoperiodic flowering, deetiolation, and transcriptional regulation of circadian clock gene homologs. Plant Physiol. 2007, 144, 648–661. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Lou, P.; Hermand, V.; Aman, R.; Park, H.J.; Yun, D.-J.; Kim, W.Y.; Salmela, M.J.; Ewers, B.E.; Weinig, C.; et al. Allelic polymorphism of GIGANTEA is responsible for naturally occurring variation in circadian period in Brassica rapa. Proc. Natl. Acad. Sci. USA 2015, 112, 3829–3834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayama, R.; Yokoi, S.; Tamaki, S.; TYano, M.; Shimamoto, K. Adaptation of photoperiodic control pathways produces short-day flowering in rice. Nature 2003, 422, 719–722. [Google Scholar] [CrossRef]
- Li, F.; Zhang, X.; Hu, R.; Wu, F.; Ma, J.; Meng, Y.; Fu, Y. Identification and molecular characterization of FKF1 and GI homologous genes in soybean. PLoS ONE 2013, 8, e79036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gu, Y.; Gao, H.; Qiu, L.; Chang, R.; Chen, S.; He, C. Molecular and geographic evolutionary support for the essential role of GIGANTEAa in soybean domestication of flowering time. BMC Evol. Biol. 2016, 16, 79. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-K.; Kim, E.-S.; Kim, K.H.; Jeong, N.; Lee, J.S.; Kang, S. Genetic variance for flowering time conferring E2 gene in photoperiod-insensitive early-maturing soybean accessions and topological distribution in Korea peninsula. Mol. Breed. 2018, 38, 148. [Google Scholar] [CrossRef]
- Sawa, M.; Nusinow, D.A.; Kay, S.A.; Imaizumi, T. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 2007, 318, 261–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.H.; Smith, R.W.; To, B.J.; Millar, A.J.; Imaizumi, T. FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering. Science 2012, 336, 1045–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaizumi, T.; Schultz, T.F.; Harmon, F.G.; Ho, L.A.; Kay, S.A. FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science 2005, 309, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-H.; Seo, Y.-H.; Seo, P.J.; Reyes, J.L.; Yun, J.; Chua, N.-H.; Park, C.-M. The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis. Plant Cell 2007, 19, 2736–2748. [Google Scholar] [CrossRef] [Green Version]
- Aukerman, M.J.; Sakai, H. Regulation of flowering time and floral organ identity by a microRNA and its APETALA2-LIKE target genes. Plant Cell 2003, 15, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Sun, M.-Y.; Wang, X.-S.; Li, W.-B.; Li, Y.-G. Over-expression of GmGIa-regulated soybean miR172a confers early flowering in transgenic Arabidopsis thaliana. Int. J. Mol. Sci. 2016, 17, 645. [Google Scholar] [CrossRef] [Green Version]
- Hartwig, E.E.; Kiihl, R.A.S. Identification and utilization of a delayed flowering character in soybeans for short-day condtions. Field Crops Res. 1979, 2, 145–151. [Google Scholar] [CrossRef]
- Destro, D.; Carpentieri-Pipolo, V.; Kiihl, R.A.S.; de Almeida, L.A. Photoperiodism and genetic control of the long juvenile period in soybean: A review. Crop Breed. Appl. Biotechnol. 2001, 1, 72–92. [Google Scholar] [CrossRef]
- Ray, J.D.; Hinson, K.; Mankono, E.B.; Malo, M.F. Genetic control of a long-juvenile trait in soybean. Crop Sci. 1995, 35, 1001–1006. [Google Scholar] [CrossRef]
- Bonato, E.R.; Vello, N.A. E6, a dominant gene conditioning early flowering and maturity in soybeans. Genet. Mol. Biol. 1999, 22, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fang, C.; Xu, M.; Zhang, F.; Lu, S.; Nan, H.; Su, T.; Li, S.; Zhao, X.; Kong, L.; et al. Quantitative Trait Locus mapping of soybean maturity gene E6. Crop Sci. 2017, 57, 2547–2554. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Zhao, X.; Hu, Y.; Liu, S.; Nan, H.; Li, X.; Fang, C.; Cao, D.; Shi, X.; Kong, L.; et al. Natural variation at the soybean J locus improves adaptation to the tropics and enhances yield. Nat. Genet. 2017, 49, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Preuss, S.B.; Meister, R.; Xu, Q.; Urwin, C.P.; Tripodi, F.A.; Screen, S.E.; Anil, V.S.; Zhu, S.; Morrell, J.A.; Liu, G.; et al. Expression of the Arabidopsis thaliana BBX32 gene in soybean increases grain yield. PLoS ONE 2012, 7, e30717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridge, S.; Deokar, A.; Lee, R.; Daba, K.; Macknight, R.C.; Weller, J.L.; Tar’an, B. The Chickpea Early Flowering 1 (Efl1) locus Is an ortholog of Arabidopsis ELF3. Plant Physiol. 2017, 175, 802–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, J.L.; Liew, L.C.; Hecht, V.F.G.; Rajandran, V.; Laurie, R.E.; Ridge, S.; Wenden, B.; Vander Schoor, J.K.; Jaminon, O.; Blassiau, C.; et al. A conserved molecular basis for photoperiod adaptation in two temperate legumes. Proc. Natl. Acad. Sci. USA 2012, 109, 21158–21163. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, H.; Li, R.; Hu, R.; Fan, C.; Chen, F.; Wang, Z.; Liu, X.; Fu, Y.; Lin, C. Association of the circadian rhythmic expression of GmCRY1a with a latitudinal cline in photoperiodic flowering of soybean. Proc. Natl. Acad. Sci. USA 2008, 105, 21028–21033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.-W.; Liu, W.; Lam, H.-M.; Gendron, J.M. Characterization of two growth period QTLs reveals modification of PRR3 genes during soybean domestication. Plant Cell Physiol. 2019, 60, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.B.; Shen, Y.; Chang, H.-C.; Hou, Y.; Harris, A.; Ma, S.F.; McPartland, M.; Hymus, G.J.; Adam, L.; Marion, C.; et al. The flowering time regulator CONSTANS is recruited to the FLOWERING LOCUS T promoter via a unique cis-element. New Phytol. 2010, 187, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yuan, L.; Su, T.; Wang, Q.; Gao, Y.; Zhang, S.; Jia, Q.; Yu, G.; Fu, Y.; Cheng, Q.; et al. Light and temperature-entrainable circadian clock in soybean development. Plant Cell Environ. 2020, 43, 637–648. [Google Scholar] [CrossRef]
- Lu, S.; Dong, L.; Fang, C.; Liu, S.; Kong, L.; Cheng, Q.; Chen, L.; Su, T.; Nan, H.; Zhang, D.; et al. Stepwise selection on homeologous PRR genes controlling flowering and maturity during soybean domestication. Nat. Genet. 2020, 52, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, Y.-h.; Li, Y.; Lu, H.; Hong, H.; Tian, Y.; Li, H.; Zhao, T.; Zhou, X.; Liu, J.; et al. A domestication-associated gene GmPRR3b regulates the circadian clock and flowering time in soybean. Mol. Plant 2020, 13, 745–759. [Google Scholar] [CrossRef]
- Li, Y.; Dong, Y.; Wu, H.; Hu, B.; Zhai, H.; Yang, J.; Xia, Z. Positional cloning of the flowering time QTL qFT12-1 reveals the link between the clock related PRR homolog with photoperiodic response in soybeans. Front. Plant Sci. 2019, 10, 1303. [Google Scholar] [CrossRef] [Green Version]
- Hotta, C.T.; Gardner, M.J.; Hubbard, K.E.; Baek, S.J.; Dalchau, N.; Suhita, D.; Dodd, A.N.; Webb, A.A.R. Modulation of environmental responses of plants by circadian clocks. Plant Cell Environ. 2007, 30, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; McClung, C.R.; Zhang, C. Tick Tock: Circadian regulation of plant innate immunity. Annu. Rev. Phytopathol. 2017, 55, 287–311. [Google Scholar] [CrossRef] [Green Version]
- Fowler, S.G.; Cook, D.; Thomashow, M.F. Low temperature induction of Arabidopsis CBF1, 2, and 3 is gated by the circadian clock. Plant Physiol. 2005, 137, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, O.; Bräutigam, K.; Campbell, M.M. Time of day shapes Arabidopsis drought transcriptomes. Plant J. 2010, 63, 715–727. [Google Scholar] [CrossRef]
- Goodspeed, D.; Chehab, E.W.; Min-Venditti, A.; Braam, J.; Covington, M.F. Arabidopsis synchronizes jasmonate-mediated defense with insect circadian behavior. Proc. Natl. Acad. Sci. USA 2012, 109, 4674–4677. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Carlsson, J.; Takeuchi, T.; Newton, L.; Farré, E.M. Direct regulation of abiotic responses by the Arabidopsis circadian clock component PRR7. Plant J. 2013, 76, 101–114. [Google Scholar] [CrossRef]
- Lei, J.; Jayaprakasha, G.K.; Singh, J.; Uckoo, R.; Borrego, E.J.; Finlayson, S.; Kolomiets, M.; Patil, B.S.; Braam, J.; Zhu-Salzman, K. CIRCADIAN CLOCK-ASSOCIATED1 controls resistance to aphids by altering indole glucosinolate production. Plant Physiol. 2019, 181, 1344–1359. [Google Scholar] [CrossRef] [Green Version]
- Mody, T.; Bonnot, T.; Nagel, D.H. Interaction between the circadian clock and regulators of heat stress responses in plants. Genes 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, O.; Waldron, L.; Nahal, H.; Provart, N.J.; Campbell, M.M. Genotype and time of day shape the Populus drought response. Plant J. 2009, 60, 703–715. [Google Scholar] [CrossRef]
- Greenham, K.; Guadagno, C.R.; Gehan, M.A.; Mockler, T.C.; Weinig, C.; Ewers, B.E.; McClung, C.R. Temporal network analysis identifies early physiological and transcriptomic indicators of mild drought in Brassica rapa. eLIFE 2017, 6, e29655. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Cao, L.; Mwimba, M.; Zhou, Y.; Li, L.; Zhou, M.; Schnable, P.S.; O’Rourke, J.A.; Dong, X.; Wang, W. Comprehensive mapping of abiotic stress inputs into the soybean circadian clock. Proc. Natl. Acad. Sci. USA 2019, 116, 23840–23849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Bu, T.; Cheng, Q.; Dong, L.; Su, T.; Chen, Z.; Kong, F.; Gong, Z.; Liu, B.; Li, M. Two homologous LHY pairs negatively control soybean drought tolerance by repressing the abscisic acid responses. New Phytol. 2020. [Google Scholar] [CrossRef]
- Blanca, J.; Cañizares, J.; Cordero, L.; Pascual, L.; Diez, M.J.; Nuez, F. Variation revealed by SNP genotyping and morphology provides insight into the origin of the tomato. PLoS ONE 2012, 7, e48198. [Google Scholar] [CrossRef] [Green Version]
- Müller, N.A.; Wijnen, C.; Srinivasan, A.; Ryngajllo, M.; Ofner, I.; Lin, T.; Ranjan, A.; West, D.; Maloof, J.N.; Sinha, N.R.; et al. Domestication selected for deceleration of the circadian clock in cultivated tomato. Nat. Genet. 2016, 48, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Dieterle, M.; Zhou, Y.-C.; Schäfer, E.; Funk, M.; Kretsch, T. EID1, an F-box protein involved in phytochrome A-specific light signaling. Genes Dev. 2001, 15, 939–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, N.A.; Zhang, L.; Koornneef, M.; Jiménez-Gómez, J.M. Mutations in EID1 and LNK2 caused light-conditional clock deceleration during tomato domestication. Proc. Natl. Acad. Sci. USA 2018, 115, 7135–7140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rugnone, M.L.; Faigón Soverna, A.; Sanchez, S.E.; Schlaen, R.G.; Hernando, C.E.; Seymour, D.K.; Mancini, E.; Chernomoretz, A.; Weigel, D.; Más, P.; et al. LNK genes integrate light and clock signaling networks at the core of the Arabidopsis oscillator. Proc. Natl. Acad. Sci. USA 2013, 110, 12120–12125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abegg, F.A. A genetic factor for the annual habit in beets and linkage relationship. J. Agric. Res. 1936, 53, 493–511. [Google Scholar]
- Büttner, B.; Abou-Elwafa, S.F.; Zhang, W.; Jung, C.; Müller, A.E. A survey of EMS-induced biennial β vulgaris mutants reveals a novel bolting locus which is unlinked to the bolting gene B. Theor. Appl. Genet. 2010, 121, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Pin, P.A.; Zhang, W.; Vogt, S.H.; Dally, N.; Büttner, B.; Schulze-Buxloh, G.; Jelly, N.S.; Chia, T.Y.; Mutasa-Göttgens, E.S.; Dohm, J.C.; et al. The role of a pseudo-response regulator gene in life cycle adaptation and domestication of beet. Curr. Biol. 2012, 22, 1095–1101. [Google Scholar] [CrossRef] [Green Version]
- Pin, P.A.; Benlloch, R.; Bonnet, D.; Wremerth-Weich, E.; Kraft, T.; Gielen, J.J.; Nilsson, O. An antagonistic pair of FT homologs mediates the control of flowering time in sugar beet. Science 2010, 330, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Dally, N.; Xiao, K.; Holtgräwe, D.; Jung, C. The B2 flowering time locus of beet encodes a zinc finger transcription factor. Proc. Natl. Acad. Sci. USA 2014, 111, 10365–10370. [Google Scholar] [CrossRef] [Green Version]
- Höft, N.; Dally, N.; Jung, C. Sequence variation in the bolting time regulator BTC1 changes the life cycle regime in sugar beet. Plant Breed. 2019, 137, 412–422. [Google Scholar] [CrossRef]
- Nagy, F.; Kay, S.A.; Chua, N.H. A circadian clock regulates transcription of the wheat Cab-1 gene. Genes Dev. 1988, 2, 376–382. [Google Scholar] [CrossRef] [Green Version]
- Redinbaugh, M.G.; Sabre, M.; Scandalios, J.G. Expression of the maize Cat3 catalase gene is under the influence of a circadian rhythm. Proc. Natl. Acad. Sci. USA 1990, 87, 6853–6857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takata, N.; Saito, S.; Saito, C.T.; Nanjo, T.; Shinohara, K.; Uemura, M. Molecular phylogeny and expression of poplar circadian clock genes,LHY1 and LHY2. New Phytol. 2009, 181, 808–819. [Google Scholar] [CrossRef]
- Takata, N.; Saito, S.; Saito, C.T.; Uemura, M. Phylogenetic footprint of the plant clock system in angiosperms: Evolutionary processes of Pseudo-Response Regulators BMC Evol. Biol. 2010, 10, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClung, C.R. A modern circadian clock in the common angiosperm ancestor of monocots and eudicots. BMC Biol. 2010, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Ashikari, M.; Miura, K.; Yamashino, T.; Mizuno, T. The evolutionarily conserved OsPRR quintet: Rice Pseudo-Response Regulators implicated in circadian rhythm. Plant Cell Physiol. 2003, 44, 1229–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, M.; Tago, Y.; Yamashino, T.; Mizuno, T. Comparative overviews of clock-associated genes of Arabidopsis thaliana and Oryza sativa. Plant Cell Physiol. 2007, 48, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Miwa, K.; Serikawa, M.; Suzuki, S.; Kondo, T.; Oyama, T. Conserved expression profiles of circadian clock-related genes in two Lemna species showing long-day and short-day photoperiodic flowering responses. Plant Cell Physiol. 2006, 47, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Serikawa, M.; Miwa, K.; Kondo, T.; Oyama, T. Functional conservation of clock-related genes in flowering plants: Overexpression and RNAi analyses of the circadian rhythm in the monocotyledon Lemna gibba. Plant Physiol. 2008, 146, 1952–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muranaka, T.; Okada, M.; Yomo, J.; Kubota, S.; Oyama, T. Characterisation of circadian rhythms of various duckweeds. Plant Biol. 2015, 17, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Campoli, C.; Shtaya, M.; Davis, S.J.; von Korff, M. Expression conservation within the circadian clock of a monocot: Natural variation at barley Ppd-H1 affects circadian expression of flowering time genes, but not clock orthologs. BMC Plant Biol. 2012, 12, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faure, S.; Turner, A.S.; Gruszka, D.; Christodoulou, V.; Davis, S.J.; von Korff, M.; Laurie, D.A. Mutation at the circadian clock gene EARLY MATURITY 8 adapts domesticated barley (Hordeum vulgare) to short growing seasons. Proc. Natl. Acad. Sci. USA 2012, 109, 8328–8333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campoli, C.; Pankin, A.; Drosse, B.; Casao, C.M.; Seth, J.; Davis, S.J.; von Korff, M. HvLUX1 is a candidate gene underlying the early maturity 10 locus in barley: Phylogeny, diversity, and interactions with the circadian clock and photoperiodic pathways. New Phytol. 2013, 199, 1045–1059. [Google Scholar] [CrossRef] [Green Version]
- Müller, L.M.; Mombaerts, L.; Pankin, A.; Davis, S.J.; Webb, A.A.R.; Goncalves, J.; von Korff, M. Differential effects of day/night cues and the circadian clock on the barley transcriptome. Plant Physiol. 2020, 183, 765–779. [Google Scholar] [CrossRef] [PubMed]
- Izawa, T.; Mihara, M.; Suzuki, Y.; Gupta, M.; Itoh, H.; Nagano, A.J.; Motoyama, R.; Sawada, Y.; Yano, M.; Hirai, M.Y.; et al. Os-GIGANTEA confers robust diurnal rhythms on the global transcriptome of rice in the field. Plant Cell 2011, 23, 1741–1755. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Tago, Y.; Yamashino, T.; Mizuno, T. Characterization of the rice circadian clock-associated Pseudo-Response Regulators in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2007, 71, 1107–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Zhang, Z.; Cheng, N.; Liu, H.; Song, S.; Hu, Y.; Zhou, X.; Zhang, J.; Xing, Y. The transcriptional repressor OsPRR73 links the circadian clock and photoperiod pathway to control heading date in rice. Plant Cell Environ. 2021, 44, 842–855. [Google Scholar] [CrossRef]
- Pankin, A.; Campoli, C.; Dong, X.; Kilian, B.; Sharma, R.; Himmelbach, A.; Saini, R.; Davis, S.J.; Stein, N.; Schneeberger, K.; et al. Mapping-by-sequencing identifies HvPHYTOCHROME C as a candidate gene for the early maturity 5 locus modulating the circadian clock and photoperiodic flowering in barley. Genetics 2014, 198, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakhrabekova, S.; Gough, S.P.; Braumann, I.; Müller, A.H.; Lundqvist, J.; Ahmann, K.; Dockter, C.; Matyszczak, I.; Kurowska, M.; Druka, A.; et al. Induced mutations in circadian clock regulator Mat-a facilitated short-season adaptation and range extension in cultivated barley. Proc. Natl. Acad. Sci. USA 2012, 109, 4326–4331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Jin, M.; Zheng, X.-M.; Chen, J.; Yuan, D.; Xin, Y.; Wang, M.; Huang, D.; Zhang, Z.; Zhou, K.; et al. Days to heading 7, a major quantitative locus determining photoperiod sensitivity and regional adaptation in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 16337–16342. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.-M.; Feng, L.; Wang, J.; Qiao, W.; Zhang, L.; Cheng, Y.; Yang, Q. Nonfunctional alleles of long-day suppressor genes independently regulate flowering time. J. Integr. Plant Biol. 2016, 58, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, J.; Feng, L.; Liu, S.; Li, J.; Qiao, W.; Song, Y.; Zhang, Z.; Cheng, Y.; Zhang, L.; et al. A combination of long-day suppressor genes contributes to the northward expansion of rice. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Gao, H.; Zheng, X.-M.; Fei, G.; Chen, J.; Jin, M.; Ren, Y.; Wu, W.; Zhou, K.; Sheng, P.; Zhou, F.; et al. Ehd4 encodes a novel and Oryza-genus-specific regulator of photoperiodic flowering in rice. PLoS Genet. 2013, 9, e1003281. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Chen, H.; Ren, D.; Tang, H.; Qiu, R.; Feng, J.; Long, Y.; Niu, B.; Chen, D.; Zhong, T.; et al. Genetic interactions between diverged alleles of Early heading date 1 (Ehd1) and Heading date 3a (Hd3a)/ RICE FLOWERING LOCUS T1 (RFT1) control differential heading and contribute to regional adaptation in rice (Oryza sativa). New Phytol. 2015, 208, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.; Beales, J.; Faure, S.; Dunford, R.P.; Laurie, D.A. The Pseudo-Response Regulator Ppd-H1 provides adaptation to photoperiod in barley. Science 2005, 310, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Ito, S.; Song, Y.H.; Strait, A.A.; Kiba, T.; Lu, S.; Henriques, R.; Pruneda-Paz, J.L.; Chua, N.-H.; Tobin, E.M.; et al. F-Box Proteins FKF1 and LKP2 act in concert with ZEITLUPE to control Arabidopsis clock progression. Plant Cell 2010, 22, 606–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Digel, B.; Tavakol, E.; Verderio, G.; Tondelli, A.; Xu, X.; Cattivelli, L.; Rossini, L.; von Korff, M. Photoperiod-H1 (Ppd-H1) controls leaf size. Plant Physiol. 2016, 172, 405–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruneda-Paz, J.L.; Kay, S.A. An expanding universe of circadian networks in higher plants. Trends Plant Sci. 2010, 15, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Green, R.M.; Tingay, S.; Wang, Z.-Y.; Tobin, E.M. Circadian rhythms confer a higher level of fitness to Arabidopsis plants. Plant Physiol. 2002, 129, 576–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerushalmi, S.; Yakir, E.; Green, R.M. Circadian clocks and adaptation in Arabidopsis. Mol. Ecol. 2011, 20, 1155–1165. [Google Scholar] [CrossRef]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kevei, E.; Toth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A.R. Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef] [Green Version]
- Toniutti, L.; Breitler, J.-C.; Guittin, C.; Doulbeau, S.; Etienne, H.; Campa, C.; Lambot, C.; Herrera Pinilla, J.-C.; Bertrand, B. An altered circadian clock coupled with a higher photosynthesis efficiency could explain the better agronomic performance of a new coffee clone when compared with a standard variety. Intl. J. Mol. Sci. 2019, 20, 736. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.; Kim, E.-D.; Ha, M.; Lackey, E.; Liu, J.; Zhang, Y.; Sun, Q.; Chen, Z.J. Altered circadian rhythms regulate growth vigour in hybrids and allopolyploids. Nature 2009, 457, 327–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, D.W.-K.; Miller, M.; Yu, H.H.; Huang, T.-Y.; Kim, E.-D.; Lu, J.; Xie, Q.; McClung, C.R.; Chen, Z.J. A role for CHH methylation in the parent-of-origin effect on altered circadian rhythms and biomass heterosis in Arabidopsis intraspecific hybrids. Plant Cell 2014, 26, 2430–2440. [Google Scholar] [CrossRef] [Green Version]
- Ko, D.K.; Rohozinski, D.; Song, Q.; Taylor, S.H.; Juenger, T.E.; Harmon, F.G.; Chen, Z.J. Temporal shift of circadian-mediated gene expression and carbon fixation contributes to biomass heterosis in maize hybrids. PLoS Genet. 2016, 12, e1006197. [Google Scholar] [CrossRef]
- Howden, S.M.; Soussana, J.-F.; Tubiello, F.N.; Chhetri, N.; Dunlop, M.; Meinke, H. Adapting agriculture to climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19691–19696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Massel, K.; Godwin, I.D.; Gao, C. Applications and potential of genome editing in crop improvement. Genome Biol. 2018, 19, 210. [Google Scholar] [CrossRef]
Figure 1.
A simplified representation of the Arabidopsis circadian clock network. The clockwork components are represented from left to right according to the time-of-day of their peak expression. CCA1 and LHY are expressed early in the morning, followed sequentially by PRR9, PRR7, PRR5, and TOC1, which peaks at dusk. The “evening complex” (EC) is formed by LUX, ELF3, and ELF4, and is expressed after dusk. These components are transcriptional repressors, with repression indicated by lines ending in perpendicular lines. TOC1 interacts with CHE (TCP21) to negatively regulate CCA1 and LHY. In the morning, the transcription factors TCP20 and TCP22 with LWD1 and LWD2 acting as coactivators activate expression of (indicated by the green arrows) CCA1, LHY, PRR9 and PRR5. In the middle of the day RVE8 (and RVE4 and RVE6), with LNK1 and LNK2 acting as coactivators, activate expression of PRR5, TOC1, GI, LUX, and ELF4. Late in the day. FHY3, FAR1, and HY5 activate ELF4 expression. Please consult the main text for further details.
Figure 1.
A simplified representation of the Arabidopsis circadian clock network. The clockwork components are represented from left to right according to the time-of-day of their peak expression. CCA1 and LHY are expressed early in the morning, followed sequentially by PRR9, PRR7, PRR5, and TOC1, which peaks at dusk. The “evening complex” (EC) is formed by LUX, ELF3, and ELF4, and is expressed after dusk. These components are transcriptional repressors, with repression indicated by lines ending in perpendicular lines. TOC1 interacts with CHE (TCP21) to negatively regulate CCA1 and LHY. In the morning, the transcription factors TCP20 and TCP22 with LWD1 and LWD2 acting as coactivators activate expression of (indicated by the green arrows) CCA1, LHY, PRR9 and PRR5. In the middle of the day RVE8 (and RVE4 and RVE6), with LNK1 and LNK2 acting as coactivators, activate expression of PRR5, TOC1, GI, LUX, and ELF4. Late in the day. FHY3, FAR1, and HY5 activate ELF4 expression. Please consult the main text for further details.

Figure 2.
Regulation of photoperiodic flowering in Arabidopsis, a long-day plant, and in rice, a short-day plant. Upper panel. In Arabidopsis, CO is a central activator of flowering time, inducing expression of FT. FT protein acts as florigen, traveling from the leaf vasculature to the shoot apical meristem where it, together with FD, induces expression of the FLORAL MERISTEM IDENTITY (FMI) genes. CO transcription is repressed by the CDF proteins, which in long days are targeted for proteolytic degradation by the LOV/KELCH/F-BOX protein FKF1 complexed with GI. CO transcription is also independently activated by light-activated CIB proteins. In long days light signaling stabilizes CO protein through inhibition of proteolytic degradation of CO, which in short days is ubiquitylated by the E3 ubiquitin ligase COP1. CO protein is also stabilized through interaction with the PRRs during the day, when they are maximally abundant. Please consult the main text for further details. Middle panel. In rice in inductive short days the OsGI/Hd1 (OsCO) pathway activates expression of Hd3a (OsFT). Hd3a expression is also activated by Ehd1 in a distinct pathway not found in Arabidopsis. Black indicates strong regulation and gray weak regulation. Please consult the main text for further details. Lower panel. In rice in non-inductive long days light signaling through PHYB converts Hd1 into a transcriptional repressor of Hd3a. Light signaling also induces Ghd7 which encodes a repressor of Ehd1 expression. Similarly, OsPRR37 represses Ehd1 and Hda3 expression. Black indicates strong regulation and gray weak regulation. Please consult the main text for further details.
Figure 2.
Regulation of photoperiodic flowering in Arabidopsis, a long-day plant, and in rice, a short-day plant. Upper panel. In Arabidopsis, CO is a central activator of flowering time, inducing expression of FT. FT protein acts as florigen, traveling from the leaf vasculature to the shoot apical meristem where it, together with FD, induces expression of the FLORAL MERISTEM IDENTITY (FMI) genes. CO transcription is repressed by the CDF proteins, which in long days are targeted for proteolytic degradation by the LOV/KELCH/F-BOX protein FKF1 complexed with GI. CO transcription is also independently activated by light-activated CIB proteins. In long days light signaling stabilizes CO protein through inhibition of proteolytic degradation of CO, which in short days is ubiquitylated by the E3 ubiquitin ligase COP1. CO protein is also stabilized through interaction with the PRRs during the day, when they are maximally abundant. Please consult the main text for further details. Middle panel. In rice in inductive short days the OsGI/Hd1 (OsCO) pathway activates expression of Hd3a (OsFT). Hd3a expression is also activated by Ehd1 in a distinct pathway not found in Arabidopsis. Black indicates strong regulation and gray weak regulation. Please consult the main text for further details. Lower panel. In rice in non-inductive long days light signaling through PHYB converts Hd1 into a transcriptional repressor of Hd3a. Light signaling also induces Ghd7 which encodes a repressor of Ehd1 expression. Similarly, OsPRR37 represses Ehd1 and Hda3 expression. Black indicates strong regulation and gray weak regulation. Please consult the main text for further details.

Figure 3.
Regulation of photoperiodic flowering in soybean. In inhibitory long days, red light signaling through PhyA, encoded by E3 and E4, induces the expression of the flowering repressors, E1, E1La and E1Lb. E1, E1La and E1Lb inhibit expression of FT2a and FT5a, which encode florigens that travel to the shoot apical meristem to induce FLORAL MERISTEM IDENTITY genes (FMI). E1, E1La and E1Lb also activate expression of FT1a and FT4, which encode flowering inhibitors. In long days, PhyA signaling also activates expression of PRR3a and PRR3b, which inhibit the expression of CCA1/LHY, which repress expression of E1. In short days, the evening complex (J/GmELF3, LUX, and likely GmELF4) inhibits the expression of the flowering repressors. Thus loss of J or LUX function delays flowering in short days, extending vegetative growth and increasing yield in the short days of the tropics. Arrows indicate positive regulation and lines with blunt ends indicate negative regulation. Please consult the main text for further details. Modified from [102].
Figure 3.
Regulation of photoperiodic flowering in soybean. In inhibitory long days, red light signaling through PhyA, encoded by E3 and E4, induces the expression of the flowering repressors, E1, E1La and E1Lb. E1, E1La and E1Lb inhibit expression of FT2a and FT5a, which encode florigens that travel to the shoot apical meristem to induce FLORAL MERISTEM IDENTITY genes (FMI). E1, E1La and E1Lb also activate expression of FT1a and FT4, which encode flowering inhibitors. In long days, PhyA signaling also activates expression of PRR3a and PRR3b, which inhibit the expression of CCA1/LHY, which repress expression of E1. In short days, the evening complex (J/GmELF3, LUX, and likely GmELF4) inhibits the expression of the flowering repressors. Thus loss of J or LUX function delays flowering in short days, extending vegetative growth and increasing yield in the short days of the tropics. Arrows indicate positive regulation and lines with blunt ends indicate negative regulation. Please consult the main text for further details. Modified from [102].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
McClung, C.R. Circadian Clock Components Offer Targets for Crop Domestication and Improvement. Genes 2021, 12, 374. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030374
AMA Style
McClung CR. Circadian Clock Components Offer Targets for Crop Domestication and Improvement. Genes. 2021; 12(3):374. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030374
Chicago/Turabian StyleMcClung, C. Robertson. 2021. "Circadian Clock Components Offer Targets for Crop Domestication and Improvement" Genes 12, no. 3: 374. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030374
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.