
Diverse Regulation but Conserved Function: SOX9 in Vertebrate Sex Determination

1
Sex Development Laboratory, Hudson Institute of Medical Research, Melbourne, VIC 3168, Australia
2
Department of Molecular and Translational Science, Monash University, Melbourne, VIC 3800, Australia
*
Author to whom correspondence should be addressed.
Genes 2021, 12(4), 486; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040486
Submission received: 5 March 2021
/
Revised: 23 March 2021
/
Accepted: 24 March 2021
/
Published: 26 March 2021
(This article belongs to the Special Issue Evolution and Development of Testis)
Abstract
:Sex determination occurs early during embryogenesis among vertebrates. It involves the differentiation of the bipotential gonad to ovaries or testes by a fascinating diversity of molecular switches. In most mammals, the switch is SRY (sex determining region Y); in other vertebrates it could be one of a variety of genes including Dmrt1 or dmy. Downstream of the switch gene, SOX9 upregulation is a central event in testes development, controlled by gonad-specific enhancers across the 2 Mb SOX9 locus. SOX9 is a ‘hub’ gene of gonadal development, regulated positively in males and negatively in females. Despite this diversity, SOX9 protein sequence and function among vertebrates remains highly conserved. This article explores the cellular, morphological, and genetic mechanisms initiated by SOX9 for male gonad differentiation.
1. Introduction
Sexual development and the mechanisms underlying sex determination have been a key interest of developmental and evolutionary biologists. The embryonic gonad has been revered as the ideal model for the investigation of organogenesis, as the choice between male and female cell lineages from the bipotential gonad provides valuable information about the molecular regulation of cell fate and pattern formation. Vertebrate sex determination during embryogenesis is usually controlled by a genetic “switch” in which the development of the testes or ovaries arises from the bipotential gonads. Using model systems such a mealworms [1], Drosophila melanogaster [2], rabbits [3], Caenorhabditis elegans [4] and Mus musculus, the understanding of sex determination and testicular development has progressed. Reptiles [5,6,7], avian species [8,9], monotremes [10,11] and fish [12,13] display various genetic and environmental triggers of sex determination and gonad differentiation. Yet, there is the possibility for cellular transdifferentiation such that the sexual fate of post-natal gonadal cells is not permanent [14,15]. SOX9 (SRY-box 9) has a conserved role in vertebrate gonadal development, underpinned by a variety of switch mechanisms that control its expression for sex determination.
The apparent conserved role of SOX9 in male sex determination and the highly conserved protein sequence suggests a conserved function for SOX9 as a transcriptional activator of male-promoting target genes. However, the sex determination switch that controls SOX9 expression is not conserved across vertebrate species, despite the importance, and conservation, of sexually dimorphic features [16]. Across taxa, crocodilians, many turtles and fish rely on environmental cues such as temperature for sex determination, indicating there is no genetic predisposition for a temperature-sensitive species to develop as female or male until the thermosensitive period of development [5,6,17,18]. Male humans, mice, and most mammals have heteromorphic XY chromosomes in which the Y chromosome harbours the male-dominant sex-determining SRY/Sry gene (Sex determining region of Y chromosome) [19,20,21], whereas females carry XX chromosomes. In avian sex determination, females are heterogametic ZW, and males are homogametic ZZ, with the avian Z chromosome harbouring the dosage-dependent sex determining gene DMRT1 (Doublesex and mab-3 related transcription factor 1) [8]. Amphibians can have either or both XY/XX or ZW/WW sex determining mechanisms [22] and fish have quite diverse chromosomal sex determining mechanisms [23]—for example, the medaka has an XY system with dmrt1 (dmy) as the sex determining gene [13]. The platypus presents a peculiar sex determination system: males have five X and five Y chromosomes, lacking an Sry gene; females have five pairs of X chromosomes, and the platypus still harbours an autosomal Sox9 gene [10,24].
Sex determination can be considered a contest between pro-testes and pro-ovarian genes; for example in most mammals, the presence of the Y chromosome tips the balance in favour of the male cell fate [25]. At the genetic level, there is molecular antagonism among the positive and negative regulators (transcription factors) and regulatory regions (enhancers and repressors) acting on SOX9. This article will review cellular and morphological changes unique to early formation of the male gonads, with SOX9 function being conserved among species yet regulated via different mechanisms, and how this information fits into the comprehensive network of gene regulation during and after sex determination.
2. Unique Cellular and Morphological Changes during Male Gonad Development
Sex determination occurs when bipotential gonads differentiate into testes or ovaries, usually due to the karyotype of the developing fetus. Sexually dimorphic species, in which there are two distinct sexes, rely on sexual reproduction to produce offspring with genetic variation, such as the case with mammals, birds, and reptiles. While vertebrates vary in the processes of sex determination and chromosomal features, they can have relatively similar structural anatomy [26].
The genital ridge is an undifferentiated region from which testes and ovaries form, comprised of a narrow band of proliferating cells, appearing in mice around embryonic day (E)10.5 [27,28] and in humans around gestational week 4 [29]. The primordial germ cells migrate to colonise the thickening genital ridge (Figure 1) [27,30,31] to associate with undifferentiated somatic cells [32,33]. After E10.5 in mice, sex differentiation diverges, whereby the bipotential gonad commits to either the testicular or ovarian pathway [33]. Supporting cells commit to develop as either Sertoli or granulosa cells, steroidogenic cells commit to either Leydig or theca cells, and germ cells prepare for spermatogenesis or oogenesis later.
Sertoli cell differentiation in humans and mice depends on the expression of a unique gene residing on the Y chromosome, and only present in males, known as the SRY/Sry gene [19,20]. SRY in turn upregulates the transcription of the highly conserved gene SOX9/Sox9. Sertoli cell proliferation in mice from E11.25–E13.5 causes the gonadal width to double every 24 h [32] and consequently Sertoli cells aggregate to form the presumptive seminiferous tubules [34] (Figure 1). Exposure of mouse XY gonads to inhibitors of proliferation during the critical window of E10.8–11.2 results in a failure of cord formation and reduced expression of male-specific genes [34]. This critical window also aligns with the onset of SRY and SOX9 expression and the onset of early male sex determination. Following differentiation, Sertoli cells direct the compartmentalisation of the testis cords and the interstitial space around E12.5 [32]. Testis cords encapsulate the germ cells, and Sertoli cells work in conjunction with peritubular myoid cells to deposit a basal lamina (Figure 1). This deposition is critical to provide structural integrity to the testis cords and separate them from the interstitial space to provide the right conditions for spermatogenesis later.
In summary, Sertoli cells are a key component of testis differentiation, driven by the expression of SRY and SOX9. It is remarkable that three unique cells lineages in the bipotential fetal gonad (supporting cells, steroidogenic cells, and germ cells) have the capacity to differentiate into testicular or ovarian lineages. This fate commitment, once thought to be permanent, requires constant reinforcement to prevent transdifferentiation [14,15]. Understanding the molecular mechanisms that drive gonadal differentiation can facilitate the understanding of the genes and pathways related to disorders of sex development (DSD) and provide understanding of differentiation and development of other organs during embryogenesis.
3. SRY and DMRT1: Key Switches in Sex Determination
Historically, the sex determining region of the Y chromosome in humans and mice was given the acronyms TDF (Testis-determining factor) and Tdy (testis-determining region on the Y chromosome), respectively [35]. Now, the gene determining male sex has been identified and named SRY in humans and Sry in mice [19,20]. This gene has been previously well reviewed [36,37,38,39,40], and will not be comprehensively covered in this article. To fully understand the development of the testis it is important to briefly discuss SRY/Sry—the gene essential for male sex determination.
SRY/Sry is expressed exclusively and transiently in the supporting cells of the genital ridge to direct cells to develop as Sertoli cells [41,42]. In mice, Sry gene expression begins at E10.5, reaching peak expression at E11.5 then declining rapidly to an undetectable level by E12.5 [43]; in humans, SRY gene expression is initiated around day 40 post conception, plateauing around day 48 [42]. The expression of the mouse SRY protein first appears centrally in the developing gonad, further moving towards the posterior and anterior poles [44]. The resulting Sertoli cells continually express SOX9 during testicular development. SOX9 acts as a part of a cascade, initiating a positive feedback loops for SOX9 expression, and upregulating expression of anti-Müllerian hormone (AMH), Desert Hedgehog (DHH), Peptidyl arginine deaminase 2 (PADI2), and Prostaglandin D2 synthase (PTGDS) [45,46]. Both SRY and SOX9 contain a sequence-specific DNA HMG-box capable of regulating transcription. Interestingly, while it has been shown that Sox9/SOX9 is regulated by SRY expression in mice [47] and humans [48], it remains less clear how regulation of Sry/SRY occurs [40,49].
SRY is not the key sex determining switch in all species as it is only present in mammals. As previously mentioned, DMRT1 plays an integral role in avian sex determination in a dosage-dependent manner [8]. Expression of Dmrt1 in the mouse gonad is not observed in a testis-specific manner until E12.5, as expression is observed in somatic cells and germ cells of both sexes until this point [50]. The Japanese rice fish Oryzias latipes (Medaka) has an XY system with dmy as the sex determining gene [13,51], with dmy considered to have appeared via gene duplication of DMRT1 [52]. Further review of DNA-binding DM domains and the role of Dmrt1 in many vertebrates has been previously reviewed [53] and will not be covered comprehensively in this review.
4. Identification of Testis-Specific Enhancers of SOX9/Sox9
Within the unusually large topologically associating domain (TAD) spanning 68.67 to 70.45 Mb on Chromosome 17 in humans (17q24.3), SOX9 is the only protein coding gene [54,55,56], and tissue and temporal-specific regulation is achieved through complex mechanisms [55]. The specific mechanism by which SRY activates SOX9/Sox9 has only begun to be elucidated, with previously poor understanding in humans and partial understanding in mice. SOX9 tissue specific expression is driven by long-range regulatory elements, such as enhancers, within the 2 Mb region upstream of the TSS [48]. Initially, it was proposed that SRY and Steroidogenic factor 1 (SF1, encoded by NR5A1) act synergistically in mice to activate a Sox9 enhancer known as the Testis Specific Enhancer of Sox9 core (Tesco) [47,48,57,58]. This testis-specific enhancer was discovered starting with a bacterial artificial chromosome harbouring a 120 kb genomic fragment with regulatory regions up and downstream of the Sox9 transcription start site (TSS), and in which the Sox9 gene was replaced by a lac-Z reporter gene [58]. Within a 3.2 kb genomic fragment (TES), a 1.4 kb enhancer (TESCO) mirrored endogenous Sox9 expression, including onset at E10.5, increased expression at E11.5, and expression restricted to the testis only by E12.5 [58]. After SOX9 expression is initiated, SOX9 creates a positive feedback loop in which it auto-regulates its own transcription via TESCO [58], and Sox9 expression is maintained via the action of Fibroblast growth factor 9 (FGF9)-fibroblast growth factor receptor 2 (FGFR2) and prostaglandin D2 synthase (PTGDS)-prostaglandin D2 (PGD2) positive feedback loops [59]. However, although deletion of TESCO in mice results in reduced Sox9 expression in the testis, it is insufficient to cause sex reversal [47]. This implied that Tesco is not the sole enhancer required for Sox9 expression in mice. Human equivalents of mouse enhancers are not always active in mice [60]; the human TES sequence fails to direct testes-specific expression in transgenic mice [60] and no mutations in TESCO have been identified in DSD patients [61].
Within TESCO, an evolutionary conserved region (ECR) of 180 bp exists in mammals, reptiles, birds and amphibians [62]. Within this ECR, highly conserved modules indicate predicted regulatory roles for SOX, DMRT and GATA proteins; this conserved sequence supports the notion that vertebrates might share common aspects of Sox9 transcriptional regulation despite the diversity of sex determination switches [62].
Through understanding the varied biological mechanisms causing disorders of sex development, the understanding of SOX9 gene regulation has subsequently expanded, which has led to the identification of several SOX9/Sox9 testis-specific enhancers. This further highlights SOX9 as a ‘hub’ gene of gonadal development. XYSR is a regulatory region approximately 500 kb upstream of the SOX9 TSS, in which 46,XY sex reversal occurs with deletion of the region [63,64,65] (Figure 2). This region was narrowed to 5.2 kb and hypothesised to include a core gonadal enhancer for SOX9 involved in 46,XY and 46,XX disorders of sex development (DSD) [48].
RevSex (Reversal of Sex), a 24 kb sex determining region 517 kb upstream of SOX9 was identified through its duplication in patients with isolated DSDs [48,64]. The region was further explored to identify a putative sex reversal enhancer (eSR-B) within the RevSex region. Enhancer ability of eSR-B in a luciferase assay was repressed by the pro-ovarian transcription factor FOXL2 (Forkhead Box L2), despite the stimulatory effect of SOX9 in the same assay [48]. Interestingly, CRISPR/Cas9 deletion of the mouse eSR-B region showed no obvious gonadal or sex reversal phenotype at either embryonic or adult stages, and no significant changes in Sox9, Wnt4, Foxl2 or Amh mRNA expression levels were observed [48], implying that this particular enhancer may be human specific. An additional SRY-responsive enhancer eALDI (Alternate Long-Distance Initiator) identified in humans shows high resemblance to the functional characteristics of mouse TESCO: it is strongly activated by SOX9+SF1, and deletion of the corresponding region in mice demonstrated that this enhancer, like TESCO, is important for Sox9 expression levels yet not crucial for male gonad differentiation [48].
More recently, a novel gonadal regulatory element upstream of murine Sox9 has been identified and named enhancer 13 (Enh13) [48,67]. The 25.7 kb sequence in mice contains an orthologous enhancer to eSR-A of human SOX9 [48], and homozygous deletion of Enh13 lead to complete XY sex reversal in mice. Most importantly, the conserved region between mice and humans indicates that Enh13 may also have a critical role in human SOX9 expression and gonadal differentiation. In light of the new regions identified, such as Enh13 and eSR-A, more work is required to understand the role of antagonism and synergistic regulation through these enhancers.
5. Vertebrate SOX9 Proteins
SOX9, the direct target of SRY, is important during embryogenesis for cellular differentiation of many organs and tissues [68,69,70,71,72]. SOX9 protein is expressed in a variety of tissues, with key functions in cartilage, testis, heart, glial cells, inner ear and neural crest development [56,70,73]. SOX9 is a member of the SRY-associated HMG-box (SOX) family of transcription factors, specifically subgroup SOXE, that can act upon other genes to regulate or modulate expression for cellular differentiation in a tissue specific manner. SOX-family proteins are defined by their 70 amino acid high mobility group (HMG)-type domain which is at least 50% identical to that of SRY. HMG domains of SRY/SOX proteins are evolutionarily conserved and preferentially bind double-stranded DNA with the AACAAT motif [74]. Upon binding, a DNA bend is induced [75] hence SRY/SOX are considered architectural transcription factors. The molecular basis for DNA recognition and DNA target sites identification of SOX proteins has been recently reviewed elsewhere [76]. Analysis of DNA-binding specificity of SOX9 in vitro by SELEX assay showed that the optimal SOX9 binding sequence, AGAACAATGG, includes a core DNA-binding element AACAAT, flanked by 5′ AG and 3′ GG nucleotides [77].
The HMG-box contains nuclear localisation sequences (NLS) that bind to calmodulin [78] and importin-β [79]. Defects in the calmodulin-binding NLS can prevent nuclear transportation of SRY, leading to XY sex reversal and ectopic expression of ovarian markers, as the repressed transportation of SRY means it cannot act as a transcriptional regulator of SOX9 [80,81]. The human SOX9 protein is 509 amino acids with several distinct domains: the defining high-mobility group box, a dimerization domain, a predicted transactivating domain in the middle (TAM), a weak transactivation domain rich in prolines/glutamines/alanines (PQA) and a strong transactivation domain rich in prolines/glutamines/serines (PQS, also referred to as the TAC, or transactivating domain at the C-terminus) [60] (Figure 3). The unique TAC may enhance and mediate transactivation activity in specific contexts through the PQA-rich domain [82,83,84]. SOX9 proteins can homodimerize or heterodimerize with other SOXE proteins via the DIM-HMG interactions (Figure 3) to cooperatively regulate their target genes [85]. Additionally, partner factors cooperate with SOX9 to change genome engagement and target gene expression. For example, in the testis, SOX9 and SF1 recruit each other to the testis-specific enhancers of SOX9/Sox9 to maintain SOX9 protein expression [48,58]. During chondrogenesis, SOX9 together with SOX5 and SOX6 induce chondrocyte-specific gene expression [86]. In addition, SOX9 functions as a pioneer factor in hair follicle stem cells, capable of binding condensed chromatin, to promote and maintain cell fate [87].
SOX9 is also subject to post-translational modifications: phosphorylation and acetylation to modify nuclear import, and ubiquitination and SUMOylation (small ubiquitin-like modifier) for rate of degradation [88]. Post-translational modifications of SOX9 are observed in many species, such as phosphorylation of Ser64 and Ser181 in mouse, chicken and human SOX9 orthologues [89]. Phosphorylation of SOX9 by protein kinase A (PKA) in response to BMP/TGF-β signaling occurs at Ser64 and Ser181 for chondrocyte differentiation, and SUMOylation through lysine, analogous to ubiquitination, usually occurs to regulate transcriptional repression [89]. The regions in which post-translational modifications for chondrocyte differentiation occur are also within the highly conserved sequence near the N-terminus of the orthologs—further highlighting the notion that in many species, the role of SOX9 shows higher conservation during chondrogenesis compared to sex determination. CARM1 methylation of SOX9 near the HMG-box at arginine residues (R74 and R152) is also observed in chondrocytes, driving cell cycle progression [89].
SOX9 protein sequences from 28 vertebrates were aligned (Figure 4). The multiple sequence alignment (Figure 4) highlights the high degree to which the amino acid sequence is conserved across all 28 species examined, and Table 1 lists the identity concordance, ranging from 100% between human and rhesus monkey, through to the lowest level of 70.34% with the zebrafish (58 mismatches, 47 residue difference in length). This table also indicates the specific number of residues different between various species and human SOX9.
SOX9 proteins vary in amino acid length across vertebrate species as shown in Table 1; for example, from 462 amino acids in the zebrafish to 529 in the leopard. It is apparent in the multiple sequence alignment (Figure 4) that there are very little differences in the HMG box across the 28 species examined. Little difference also exists in the DIM, and variations in sequence are most abundant towards the C-terminus. Amino acid conservation of SOX9 relative to human varies from 99.61% (baboon) to 100% (rhesus monkey) concordance in primates. In rodents, this varies from 96.27% (rat) to 96.86% (hamster). Of the avian species included, the kiwi and the chicken show 82.73% and 83.52% concordance, respectively. However, the turkey shows a low level of concordance to human (71.64%); as observed in the phylogenetic tree (Figure 5), the turkey SOX9 sequence also varies to that of the phylogenetically related chicken (as evident from the extended branch). The high variation in the C-terminus region in these vertebrate species, specifically in the PQA and PQS transactivating domains, indicate that the transcriptional activation functions have evolved.
In zebrafish (Danio rerio), Japanese medaka (Oryzias latipes) and platyfish (Xiphosphorus maculatus), gene duplication has resulted in the occurrence of two orthologs: sox9a and sox9b [91]. Through analysis using the dN/dS ratio, a measure of evolutionary pressure on protein-coding regions, it becomes apparent that the sox9 paralogs have similar coding sequence divergence and higher dN/dS ratio than non-teleost orthologs—indicating that there may have been relaxed negative selection on both sox9a and sox9b after gene duplication [91]. Retention of the two copies may occur as a result of advantageous mutations leading to new functions. This may explain why sox9a and sox9b are both expressed in the eyes and brain of zebrafish, Japanese medaka, and platyfish, yet sox9a is expressed in the testis of only the zebrafish; and why in the Japanese medaka and the playtfish, sox9a is expressed in the ovary [91]. Conversely, in zebrafish sox9b is expressed in the ovary, whereas in Medaka sox9b is initially expressed in the gonads of both sexes, but later becomes testis-specific at the time of testicular tubule development [91,92,93,94].
Comparing SOX9 orthologs indicates that while in humans the PQA comprises a 35–45 amino acid region rich in proline (42%), glutamine (39%) and alanines (18%); in lower vertebrates it has only 5–14 residues, with only a few glutamines in ancient fish [83]. Comparison of amino acid sequences of SOX9 from various vertebrate species highlights the high degree that the entire sequence is conserved across species (Figure 4), particularly around the HMG-box and near the N-terminus. The sequence similarity depicted in Figure 3 indicates the lower degree to which fish (Japanese medaka, platyfish and zebrafish) SOX9 sequences are conserved against human SOX9, especially near the C-terminus. The regions that are more highly conserved (near the N-terminus) are also more commonly associated with regulation during chondrogenesis, implying that the SOX9 transcriptional function during chondrogenesis is more highly conserved than during sex determination.
The neighbour-joining phylogenetic tree (Figure 5) indicates high conservation of SOX9 between supraprimate species, rodents, reptiles and birds. The mechanism of sex determination varies between these species, yet the degree to which SOX9 protein sequences vary appears minor. Monotreme species, such as the platypus, present an interesting evolutionary link between sex determination pathways. One of the five platypus X chromosomes confers homology to the avian Z chromosome [10,11]. In platypus, reverse gene dosage compared to birds is observed for DMRT1, the avian Z-linked sex gene. Male birds (ZZ) express higher levels of DMRT1, whereas in the platypus Dmrt1 has been mapped to the X5 chromosome, of which females have two [10,95]. While the platypus SOX9 protein is orthologous to that of other species, it has been ruled out as the sex determining switch [24].
6. Conserved Function of Vertebrate SOX9 Protein
A pair of XY chromosomes with SRY initiating the gene cascade for male sex determination evolved between 166 and 148 million years ago, and has remained stable in most mammals [96]. SOX9, a target of SRY, is arguably the most critical gene in the sex determination cascade of many vertebrates since it is highly conserved across species. However, the role of SOX9 in the male sex determination cascade across vertebrate species is not as conserved as to be expected. The understanding of the fundamental role that SOX9 plays in male sex determination has continued to expand since 1996, when the sexually dimorphic expression of Sox9/SOX9 became evident in mouse and chicken embryos [97] —two phylogenetically distant species, with different sex determination switch mechanisms. Expression of SOX9 in the genital ridge driving Sertoli cell differentiation is observed in mouse, chicken, turtle, [38], as well as a similar expression pattern of SOX9 in chicken and mouse skeletal systems [97]. Thus, the role that SOX9 plays within vertebrates appears to be pivotal, regardless of the sex determining switch or downstream mechanism.
Figure 5.
The evolutionary history of SOX9 was inferred using the Neighbour-Joining method [98]. The optimal tree is shown. The tree is drawn to scale, with branch lengths in the units of the number of amino acid substitutions per site to infer evolutionary distance, computed using the Poisson correction method [99]. This analysis involved the amino acid sequences of the SOX9 protein from 28 species aligned using NCBI COBALT [90]. All ambiguous positions were removed for each sequence pair (pairwise deletion option). Phylogenetic tree generated using MEGA X software [100].
Figure 5.
The evolutionary history of SOX9 was inferred using the Neighbour-Joining method [98]. The optimal tree is shown. The tree is drawn to scale, with branch lengths in the units of the number of amino acid substitutions per site to infer evolutionary distance, computed using the Poisson correction method [99]. This analysis involved the amino acid sequences of the SOX9 protein from 28 species aligned using NCBI COBALT [90]. All ambiguous positions were removed for each sequence pair (pairwise deletion option). Phylogenetic tree generated using MEGA X software [100].

SOX9 is a central ‘hub’ gene of gonadal development, with a conserved role in testis development across many vertebrates—including at the time of sex determination, and shortly after during gonad differentiation. In mammals, SOX9 is expressed in pre-Sertoli cells at the time of male sex determination and in mammals, SOX9 is both necessary and sufficient for testis development. In both mice and humans, loss of SOX9 in XY gonads leads to male-to-female sex reversal and ectopic expression of SOX9 in XX gonads results in testis development [101,102,103,104], highlighting the essential and central role of SOX9 as a ‘hub’ gene in male sex determination.
The Sox9 signaling pathway, as observed in mammalian testis development, also induces ovary-testis transition in zebrafish [105]. Similar to mammals, the relevant paralog of sox9 for this transition, sox9a, is expressed in the undifferentiated testis before the onset of Amh expression [92]. In addition, suppression of the sox9b (sox9a2) paralog by knockdown of the male switch Dmy in the Japanese rice fish Oryzias latipes (Medaka) promotes the female cascade, resulting in fertile male-to-female sex reversal [106].
Avian sex determination mechanisms have yet to be fully explored, but DMRT1 has been shown to hold a key role in male sex determination [8]. In the developing undifferentiated chicken testis, SOX9 is expressed after DMRT1 from E5.5, and overexpression of DMRT1 in E7.5 female chicken gonads induces localised activation of SOX9, resulting in the development of cord-like structures in masculinized gonads [107].
In contrast to the vertebrate species above, analyses of SOX9 expression in alligator and turtles indicate that SOX9 has an important role in testis differentiation, rather than sex determination, in reptiles with temperature-dependent sex determination (TSD) and reptiles with chromosomal sex determination (GSD) mechanisms, In Alligator mississippiensis, SOX9 is expressed in the embryonic testis during structural organisation at the end of the temperature-sensitive period [5]. The sea turtle Lepidochelys olivacea expresses SOX9 in both male and female gonads until the critical sex determining thermosensitive stage 24, from which SOX9 is maintained in differentiating male gonads at male-promoting temperatures, and downregulated in female gonads within two days as a result of female-promoting temperatures [108,109]. Both Dmrt1 and Sox9 mRNA expression increases in the snapping turtle Chelydra serpentina at male-promoting temperatures, indicating both are part of a core testis-determining mechanisms [110]. Chromosomal sex-determined reptiles such as the Chinese soft-shelled turtle Pelodiscus sinesis (ZZ/ZW) also have a complex network for sex determination. Sox9 expression in this species is first detected during early gonad differentiation. In contrast, Dmrt1 expression precedes male gonad differentiation, and ectopic expression of Dmrt1 can up-regulate Sox9 to induce masculinisation in ZW females [111]. Interestingly, Amh expression also precedes male gonadal differentiation in P. sinesis, and overexpression of Amh leads to ectopic activation and expression of Sox9, and female-to-male sex reversal [112].
Studies in amphibians have shown that the spatiotemporal expression of SOX9 in the gonads can differ quite significantly between closely related species. Like zebrafish and Medaka, the frog Xenopus laevis has two Sox9 paralogs (Sox9a and Sox9b), and both are significantly upregulated in the testes [113]. Both paralogs are expressed early in the undifferentiated developing testes, suggesting a role in male sex determination. This is surprisingly different to the expression pattern of SOX9 observed in Xenopus tropicalis; in both sexes, SOX9 is upregulated only after the gonads have differentiated. In the testis, SOX9 is restricted to the nucleus of Sertoli-like cells similar to that in other vertebrates [114]. However, in the ovary, SOX9 is first localized to the cytoplasm of previtellogenic oocytes then localized to the nucleus of vitellogenic oocytes [114]. These data indicate the crucial role that SOX9 orthologs play in a variety of stages in a tissue-specific manner during embryogenesis, with functions extending beyond that of just testicular development.
Sox9 expression is up-regulated in a male-specific manner for testes development to occur in many vertebrate species, such as chicken, mouse and alligator [115]. The influence of SOX9 on Sertoli cell differentiation appears highly conserved across species, yet the role SOX9 plays within the male sex determination gene cascade varies, implying varied regulatory pathways. SOX9 is observed in a male-specific, testis-specific manner for alligator testis differentiation, with an unknown regulator controlled via environmental temperature [5,116]. In chickens, expression of AMH is detected earlier than that of SOX9, despite SOX9 initiating Amh/AMH expression in mouse and human [117]. This suggests that the genes and processes regulated by SOX9 in gonadal development are not strictly conserved across species. RNA-seq and ChIP-Seq of mouse and chicken developing chondrocytes and Sertoli cells indicates that of the SOX9 target genes examined, there was high similarity in chondrocytes but not in Sertoli cells [118]. This indicates that chicken and mice, two vertebrates that are not phylogenetically close, harbour cell-specific binding preferences of SOX9, and that the regulatory targets of SOX9 in testis development differ between the species. Recently, the evolutionary plasticity of the gonad was further highlighted that essential supporting cell lineages in chickens are not derived from the coelomic epithelium such as in mice, but from mesenchymal origin [119]. This shows that there are fundamental differences between chicken and mouse gonad development. Surprisingly, other XY/XX chromosomal species such as horses, cattle, dogs, and cats have been reported to experience XY Sex Reversal, the mechanism by which this occurs is much less understood [120].
Given that SOX9 has a critical role in the sex determination cascade of many vertebrates and is the ‘hub’ gene in mammalian gonadal development, it might seem surprising that SOX9 did not evolve as the key switch in any of the vertebrate species examined to date. This could be due to the fact that SOX9 has vital roles across in many other developmental processes. The translocation of SOX9 to a sex chromosome may be problematic for complete organogenesis or chondrogenesis if gene dosage were altered; for example, 50%-reduced gene dosage in mice leads to bone and endocrine-specific defects similar to those observed in human haploinsufficiency syndrome Campomelic Dysplasia patients [121,122,123,124]. Thus, gonadogenesis truly highlights great cellular complexity: not only can the genetic (or environmental) trigger differ among vertebrate species, the resulting cellular assembly can too, yet resulting in such similar reproductive structures.
7. Disorders Arising from SOX9 Mutations in Humans
Abnormalities in sex differentiation and gonadogenesis can result in Disorders of Sex Development (DSD). This term encompasses a spectrum of disorders in which chromosomal, gonadal or anatomical sex is atypical [125]. The rate of congenital DSDs is estimated at 1:200 [126]. DSDs are categorised into three types on the basis of sex chromosome content: 46,XY DSD, 46,XX DSD, and sex chromosome DSDs; the most common being Klinefelter syndrome (XXY-XXXY aneuploidy) and Turner syndrome (XO aneuploidy) [127]. Mutations to SRY account for approximately 15% of 46,XY DSD [128]; while mutations in other genes such as NR5A1 or SOX9 are known to cause 46,XY DSD as part of a syndrome. To date, a genetic diagnosis is not achieved in around 50% of XY DSD cases [128,129]. Genetic diagnosis of ambiguous genitalia is particularly challenging and a clinical algorithm can facilitate this [130].
Mutations can occur within the SOX9 coding region itself or in the non-coding regulatory region. Mutations within the gene may affect protein function, resulting in loss-of-function (complete or partial); gain-of-function; or dominant-negative mutations. If a mutation occurs in the non-coding regulatory sequences of genes, this does not impact the protein sequence but may instead affect expression of the gene in a specific tissue or a specific enhancer or repressor.
Heterozygous loss-of-function mutations occur within either the coding or regulatory region for SOX9 in patients with Campomelic Dysplasia (CMPD; OMIM 114290). CMPD is a severe and fatal skeletal malformation syndrome in which 70% of 46,XY patients have either ambiguous genitalia or develop as females due to 46,XY gonadal dysgenesis [131]. Typical skeletal features of CMPD patients with mutations in the SOX9 coding region include bowed lower limbs, hypoplastic scapulae, narrow iliac wings, and non-mineralised thoracic pedicles [132]. An atypical form of CMPD, known as acampomelic campomelic dysplasia occurs as a result of alterations between 50–375 kb upstream of SOX9 resulting in a phenotype similar to that of CMPD but with the absence of bowed limbs [133]. In addition to these phenotypes in the developing bones and gonads, patients with CMPD may also show defects within other tissues in which SOX9 is expressed, such as brain (e.g., the absence of olfactory bulbs), heart, kidney and lung abnormalities [132]. Loss-of-function mutations within the coding sequence of SOX9 occur, for example, in the DNA-binding domain HMG-box, the nuclear localisation signals (NLS), or in the transactivating domain (Figure 6) [82,101,128,131,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156]. In some CMPD patients with associated sex reversal, the SOX9 coding sequence is not affected, but translocation breakpoints have been identified in the SOX9 regulatory region up to several hundred kb upstream of SOX9.
As well as SOX9 being responsible for the syndrome Campomelic Dysplasia with associated 46,XY DSD, SOX9 mutations can also cause isolated DSDs including 46,XX testicular DSD (OMIM: 278850, 300833, and 400045), which involves the development of histologically normal testis in 46,XX individuals; and 46,XX ovotesticular DSD (OMIM: 400045) which involves the development of ovotestis in which both ovarian and testicular tissue is present. 46,XY partial testicular dysgenesis (OMIM: 154230, 300018, 612965, 613762, 615542, 616067, and 616425) can result in ambiguous genitalia varying along a spectrum from almost female phenotype, to an almost male phenotype [157,158]. 46,XY DSD can be caused by heterozygous deletions at the SOX9 locus, removing parts of the upstream SOX9 regulatory region [48]. 46,XX testicular/ovotesticular DSDs can occur in an SRY-independent manner as indicated in a study in which only six of 17 patients with 46,XX ovotesticular/testicular DSD were SRY-positive [159]. Genomic duplications involving SOX9 can be the causative mutation, likely resulting in activation of SOX9 expression in the XX gonad. Three patients with total gene duplication of SOX9 [103,160,161], 17 patients with duplication of the upstream regulatory region [48,63,64,162,163,164,165,166,167], and one patient with a triplication of the regulatory region [65] have been previously identified. Sox9 knock-out mice show sex reversal [102] and overexpression of Sox9 in XX mice induced male development [168]; thus the same ‘disorders’ or developmental differences can be induced in mice as seen in humans. Furthermore, heterozygous deletion of Sox9 in mouse mimics the sex reversed phenotype as seen in CMPD in humans [169].
Recently, the first gain-of-function missense SOX9 variant (p.Glu50Lys) was identified in a patient with 46,XX ovotesticular DSD [170]; suggesting that mutations in the SOX9 gene can result in both loss- or gain-of-function. In vitro experiments showed that the SOX9 variant increased transactivation of an mTESCO-luc reporter when compared to wildtype SOX9, whereas female mice carrying this SOX9 variant did not show abnormalities of external or internal genitalia. However, it is not unusual to experience discordance in DSD-associated gene expressivity between humans and mice [170]. SOX9 mutations identified in DSDs are often involved in human infertility caused by testis gonadal dysgenesis, or XY and XX sex reversal. In a family with two 46, XX infertile males, both have a 96 kb triplication 500 kb upstream of SOX9 and present with hypotrophic testes containing no sperm [165].
SOX9 might also be involved in human hair follicle development, similar to its role in mice [171]. Congenital hypertrichosis is a rare condition characterized by excessive hair growth in humans. In a family with hypertrichosis, a large 2.4 Mb duplication 975 kb upstream of SOX9 was identified and dramatically reduced the expression of SOX9 in hair follicles [172].
Mutations in SOX9 enhancers upstream of SOX9 can also be associated with isolated craniofacial anomalies of the CMPD syndrome called Pierre Robin sequence (PRS; OMIM: 261800) [54]. This congenital syndrome is characterized by underdevelopment of the lower jaw (micrognathia), which can lead to secondary phenotypes including obstruction of the airway and retraction of the tongue. While mutations in this region do not contribute to DSDs, it is important to note the obvious presence of tissue-specific enhancers of SOX9 presenting an excellent opportunity to further investigate long-range regulation of genes crucial for development. Misdiagnosis and mismanagement of disorders due to a lack of genetic information associated with development of the testis can induce psychological and physiological risks, including gonadoblastomas, subfertility, gender dysphoria, anxiety, depression and reduced psychosexual wellbeing [173,174,175], making it ever-more-important that understanding of DSDs pathogenesis continues.
8. Gonad Plasticity: The Role of SOX9 in Transdifferentiation
The question of whether “terminally” differentiated cells, such as Sertoli cells and granulosa cells, can switch from one fate to the other was first posited in 1988, when it was suggested that an ovary-determining signal produced by an XX component may pre-empt the testis-determining action of the Y chromosome [176]. Two decades later, Uhlenhaut et al. [14] indeed demonstrated transdifferentiation of adult mouse ovaries to testes through an inducible deletion of Foxl2, a pro-ovarian gene; and in the reverse, Matson et al. [15] showed transdifferentiation of adult testes to ovaries via loss of Dmrt1.
From observations of the polled intersex syndrome (PIS) in XX female-to-male -sex reversed goats which contain a 11.7 kb deletion of the FOXL2-containing region on chromosome 1, Uhlenhaut et al. provided evidence supporting the theory that maintenance of the male fate of the gonad is a lifelong, active process, counter to the previous idea of terminal differentiation and permanent cell fate. Through inducible deletion of Foxl2 in adult mouse ovaries, upregulation of male-specific markers such as SOX9 was detected, with granulosa cells undergoing transdifferentiation to appear as testicular Sertoli cells (including the tripartite nuclei and cytoplasmic extensions). Histological analysis showed that three weeks after Foxl2 deletion, the ovarian follicles took on the appearance of testicular seminiferous tubules, with granulosa cells and theca cells transdifferentiating into Sertoli-like and Leydig-like cell lineages, respectively. Molecular studies unveiled that in sex-reversed gonads, the deletion of Foxl2 led to not only notable expression of Sox9, but also Dax1, Dhh and Dmrt1. The rapid upregulation of Sox9 indicates a direct transcriptional repression of Sox9 by FOXL2 in the ovary, possibly achieved via the testes-specific enhancer TESCO [58]. Indeed, ChIP assays confirmed that FOXL2 directly bound to Tesco in vivo, and that specific deletion of Foxl2 led to strong TESCO-ECFP activation within the transdifferentiating follicles. Furthermore, in vitro FOXL2 can attenuate TESCO activation by SF1, SRY/SF1, or SOX9/SF1. Additionally, in vitro results showed that FOXL2 synergised with ESR-1 (Estrogen Receptor 1) to repress the TESCO element [14]. Further elucidation by Georges et al. indicate that repression of SOX9 via FOXL2 occurs via multiple pathways, more so through ESR2/E2 and independent of estrogen, not via binding of FOXL2 or ESR1 through the TESCO enhancer [177]. The theory that maintenance of sexual fate requires the repression of genes of the opposite sex was further supported by Matson et al., where the converse male-to-female transdifferentiation highlights the role in which Dmrt1 is essential for maintenance of mammalian testis differentiation [15] and the two-way plasticity of gonadal development. Sertoli-cell specific loss of the DMRT1 protein in mice induced Foxl2 expression, with the antagonistic relationship between pro-ovarian Foxl2 and pro-testes Dmrt1 proving the labile fate of gonadal cells. One month after deletion of Dmrt1, adult XY males had morphologically appearing Sertoli cells (tripartite nuclei) expressing both SOX9 and DMRT1, right next to morphologically appearing granulosa cells expressing only FOXL2, with the entire testicular tissue restructuring to appear more similar to typical ovarian morphology. Theca cells developed, and mRNA for oestrogen precursors HSD17β1 and CYP19A1/aromatase were detected in mutant gonads. Oestrogen signalling acts in conjunction with FOXL2 to repress Sox9 transcription. This indicates that the antagonistic relationship persists into adulthood, with the supporting cells also remaining labile after differentiation.
In addition to Foxl2, targeted deletion of Wnt4 or R-Spondin1 in mice individually results in the partial masculinisation of the embryonic XX gonad, i.e., not complete testes development [178,179]. Testis development can be induced in embryonic XX gonads lacking both Foxl2 and Wnt4, due to the subsequent activation of Sox9 which leads to the development of seminiferous tubules and spermatogenesis [180]. These findings may explain the mechanisms behind the sex-reversal seen in goats with PIS. Since Foxl2/Wnt4 double mutant mice demonstrate that granulosa cells acquire Sertoli-like characteristics, like upregulation of Sox9, Dmrt1, and other testis genes, Wnt signaling is important for ovarian development. Both WNT4 and RSPO1 stabilize β-catenin, and ectopic expression of its stable form in XY gonads can result in male-to-female sex reversal [181]. The Wnt/β-catenin pathway blocks testicular differentiation by repressing the expression of SOX9, possibly by activation of β-catenin preventing SF1 binding to the Sox9 enhancer TESCO [181] and likely other enhancers [67].
Sox9 regulation clearly has a crucial influence in both transdifferentiation pathways. Barrionuevo et al. investigated the effect of Sertoli cell specific Sox9 ablation on a Sox−/− adult testis from postnatal day 60 mouse [182]. Sox8 and Sox9 double knockout within Sertoli cells induced testis-to-ovary reprogramming, with Sertoli to granulosa transdifferentiation as a result of Dmrt1 downregulation [182]. Sox8 and Sox9 maintain basal lamina integrity to prevent testis cord disintegration and both SOX8 and SOX9 actively suppress the ovarian program during testis development [183].
The battle between pro-ovarian and pro-testicular influence for sex determination is evident, with gonadal suppression of Sox9 essential for ovarian development in females, and the absence of FOXL2 (gonadal presence of SOX9) crucial for testicular development (Figure 7). These studies highlight the way in which development is not “final”, in the sense that the organs remain plastic and the constant struggle between the two competing pathways can be manipulated in favour of one over the other, regardless of chromosomal sex.
9. Conclusions
While the early gonad is bipotential with the capacity to develop down either sex differentiation lineage, our understanding of molecular mechanisms that drive testes differentiation is expanding through the identification of conserved elements across species, and through genome analysis of patients with DSDs. Furthermore, abnormalities arising from mutations in the SOX9 regulatory or coding regions indicate the importance of functional SOX9 protein; craniofacial disorders, testicular dysgenesis and infertility can all arise from such mutations. SOX9 is multifunctional, with tissue-specific regulation and roles during embryogenesis, but the plasticity of the regulation of such a conserved transcription factor demonstrates the important role SOX9 plays to mediate both male or female cell fate. Despite the various signals acting upon the SOX9 regulatory region—be it through a ZZ/ZW or XY/XX chromosomal system, or perhaps through temperature sex determination—the common effect of the signals seems to result in upregulation of SOX9 to promote testicular development. Contrasting to the diversity of the regulatory region, the encoded protein is highly similar in vertebrates suggesting common downstream target genes required for testis development. SOX9 mediation of Sertoli cell development can control the downstream cellular differentiation of the bipotential germ cells and supporting cells. Furthermore, the reversal of terminal differentiation of ovarian or testicular supporting cells indicates that cell fate is not canalized, raising the possibility of unappreciated postnatal changes in human DSD.
Author Contributions
Writing—original draft preparation, B.V.; writing—review and editing, B.V., Z.M., S.B.-F., V.H.; visualization, B.V., Z.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Health and Medical Research Council Program Grant 2002426 (to V.H) and Fellowship APP1154870 (to V.H.). This work was supported by the Australian Government Research Training Program Scholarship (to B.V.). This work is supported by the Victorian Government’s Operational Infrastructure Support Program.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
Figures were created using Biorender.com. Thanks to Janelle Ryan, Yuan Chen, Tasman Sutherland, and Hope Newman for critical reading and feedback of the manuscript and figures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stevens, N.M. A study of the germ cells of Aphis rosae and Aphis œnotherae. J. Exp. Zool. 1904, 2, 313–337. [Google Scholar] [CrossRef] [Green Version]
- Bridges, C.B. Non-disjunction as proof of the chromosome theory of heredity. Genetics 1916, 1, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Jost, A. The age factor in the castration of male rabbit fetuses. Proc. Soc. Exp. Biol. Med. 1947, 66, 302. [Google Scholar] [CrossRef]
- Hodgkin, J. Sex determination in the nematode C. Elegans: Analysis of tra-3 suppressors and characterization of fem genes. Genetics 1986, 114, 15–52. [Google Scholar] [CrossRef] [PubMed]
- Western, P.S.; Harry, J.L.; Graves, J.A.M.; Sinclair, A.H. Temperature-dependent sex determination: Upregulation of SOX9 expression after commitment to male development. Dev. Dyn. 1999, 214, 171–177. [Google Scholar] [CrossRef]
- Shoemaker, C.; Ramsey, M.; Queen, J.; Crews, D. Expression of Sox9, Mis, and Dmrt1 in the gonad of a species with temperature-dependent sex determination. Dev. Dyn. 2007, 236, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, C.M.; Queen, J.; Crews, D. Response of candidate sex-determining genes to changes in temperature reveals their involvement in the molecular network underlying temperature-dependent sex determination. Mol. Endocrinol. 2007, 21, 2750–2763. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Roeszler, K.N.; Ohnesorg, T.; Cummins, D.M.; Farlie, P.G.; Doran, T.J.; Sinclair, A.H. The avian Z-linked gene DMRT1 is required for male sex determination in the chicken. Nature 2009, 461. [Google Scholar] [CrossRef] [PubMed]
- da Silva, S.M.; Hacker, A.; Harley, V.; Goodfellow, P.; Swain, A.; Lovell-Badge, R. Sox9 expression during gonadal development implies a conserved role for the gene in testis differentiation in mammals and birds. Nat. Genet. 1996, 14, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Grützner, F.; Rens, W.; Tsend-Ayush, E.; El-Mogharbel, N.; O’Brien, P.C.M.; Jones, R.C.; Ferguson-Smith, M.A.; Marshall Graves, J.A. In the platypus a meiotic chain of ten sex chromosomes shares genes with the bird Z and mammal X chromosomes. Nature 2004, 432, 913–917. [Google Scholar] [CrossRef]
- Rens, W.; O’Brien, P.C.M.; Grützner, F.; Clarke, O.; Graphodatskaya, D.; Tsend-Ayush, E.; Trifonov, V.A.; Skelton, H.; Wallis, M.C.; Johnston, S.; et al. The multiple sex chromosomes of platypus and echidna are not completely identical and several share homology with the avian Z. Genome Biol. 2007, 8, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuyama, H.; Yamada, M.; Kamei, Y.; Fujiwara-Ishikawa, T.; Todo, T.; Nagahama, Y.; Matsuda, M. Dmrt1 mutation causes a male-to-female sex reversal after the sex determination by Dmy in the medaka. Chromosome Res. 2012, 20, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; Nagahama, Y.; Kobayashi, T.; Matsuda, C.; Hamaguchi, S.; Sakaizumi, M. The sex determining gene of medaka: A Y-specific DM domain gene (DMY) is required for male development. Fish Physiol. Biochem. 2003, 28, 135–139. [Google Scholar] [CrossRef]
- Uhlenhaut, N.H.; Jakob, S.; Anlag, K.; Eisenberger, T.; Sekido, R.; Kress, J.; Treier, A.C.; Klugmann, C.; Klasen, C.; Holter, N.I.; et al. Somatic Sex Reprogramming of Adult Ovaries to Testes by FOXL2 Ablation. Cell 2009, 139, 1130–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matson, C.K.; Murphy, M.W.; Sarver, A.L.; Griswold, M.D.; Bardwell, V.J.; Zarkower, D. DMRT1 prevents female reprogramming in the postnatal mammalian testis. Nature 2011, 476, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Furman, B.L.S.; Metzger, D.C.H.; Darolti, I.; Wright, A.E.; Sandkam, B.A.; Almeida, P.; Shu, J.J.; Mank, J.E. Sex Chromosome Evolution: So Many Exceptions to the Rules. Genome Biol. Evol. 2020, 12, 750–763. [Google Scholar] [CrossRef] [PubMed]
- Conover, D.O.; Kynard, B.E. Environmental Sex Determination: Interaction of Temperature and Genotype in a Fish. Science 1981, 213, 577–579. [Google Scholar] [CrossRef] [PubMed]
- Manolakou, P.; Lavranos, G.; Angelopoulou, R. Molecular patterns of sex determination in the animal kingdom: A comparative study of the biology of reproduction. Reprod. Biol. Endocrinol. 2006, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Koopman, P.; Münsterberg, A.; Capel, B.; Vivian, N.; Lovell-Badge, R. Expression of a candidate sex-determining gene during mouse testis differentiation. Nature 1990, 348, 450–452. [Google Scholar] [CrossRef]
- Sinclair, A.H.; Berta, P.; Palmer, M.; Hawkins, J.R.; Griffiths, B.L.; Smith, M.J.; Foster, J.W.; Frischauf, A.; Lovell-Badge, R.; Goodfellow, P.N. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990, 346, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Waters, P.D.; Wallis, M.C.; Marshall Graves, J.A. Mammalian sex - Origin and evolution of the Y chromosome and SRY. Semin. Cell Dev. Biol. 2007, 18, 389–400. [Google Scholar] [CrossRef]
- Sarre, S.D.; Ezaz, T.; Georges, A. Transitions between sex-determining systems in reptiles and amphibians. Annu. Rev. Genomics Hum. Genet. 2011, 12, 391–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brykov, V.A. Mechanisms of sex determination in fish: Evolutionary and practical aspects. Russ. J. Mar. Biol. 2014, 40, 407–417. [Google Scholar] [CrossRef]
- Wallis, M.C.; Delbridge, M.L.; Pask, A.J.; Alsop, A.E.; Grützner, F.; O’Brien, P.C.M.; Rens, W.; Ferguson-Smith, M.A.; Graves, J.A.M. Mapping platypus SOX genes; autosomal location of SOX9 excludes it from sex determining role. Cytogenet. Genome Res. 2007, 116, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Berta, P.; Hawkins, J.R.; Sinclair, A.H.; Taylor, A.; Griffiths, B.L.; Goodfellow, P.N.; Fellous, M. Genetic evidence equating SRY and the testis-determining factor. Nature 1990, 348, 448–450. [Google Scholar] [CrossRef]
- Ferguson-Smith, M. The evolution of sex chromosomes and sex determination in vertebrates and the key role of DMRT1. Sex. Dev. 2006, 1, 2–11. [Google Scholar] [CrossRef]
- Hu, Y.C.; Okumura, L.M.; Page, D.C. Gata4 Is Required for Formation of the Genital Ridge in Mice. PLoS Genet. 2013, 9, e1003629. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, E.; Aksoy, S.; Onel, T.; Yaba, A. Gonadal development and sex determination in mouse. Reprod. Biol. 2020, 20, 115–126. [Google Scholar] [CrossRef]
- Nef, S.; Stévant, I.; Greenfield, A. Characterizing the bipotential mammalian gonad. In Current Topics in Developmental Biology; Capel, B., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 134, pp. 167–194. ISBN 9780128115442. [Google Scholar]
- Molyneaux, K.; Wylie, C. Primordial Germ Cell Migration. Int. J. Dev. Biol. 2004, 48, 537–544. [Google Scholar] [CrossRef]
- Richardson, B.E.; Lehmann, R. Mechanisms guiding primordial germ cell migration: Strategies from different organisms. Nat. Rev. Mol. Cell Biol. 2010, 11, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Ungewitter, E.K.; Yao, H.H.-C. How to make a gonad: Cellular mechanisms governing formation of the testes and ovaries. Sex. Dev. 2013, 7, 7–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginsburg, M.; Snow, M.H.; McLaren, A. Primordial germ cells in the mouse embryo during gastrulation. Development 1990, 110, 521–528. [Google Scholar] [PubMed]
- Schmahl, J.; Capel, B. Cell proliferation is necessary for the determination of male fate in the gonad. Dev. Biol. 2003, 258, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, P.N.; Darling, S.M. Genetics of sex determination in man and mouse. Development 1988, 102, 251–258. [Google Scholar]
- She, Z.Y.; Yang, W.X. Sry and SoxE genes: How they participate in mammalian sex determination and gonadal development? Semin. Cell Dev. Biol. 2017, 63, 13–22. [Google Scholar] [CrossRef]
- Wilhelm, D.; Palmer, S.; Koopman, P. Sex determination and gonadal development in mammals. Physiol. Rev. 2007, 87, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Kanai, Y.; Hiramatsu, R.; Matoba, S.; Kidokoro, T. From SRY to SOX9: Mammalian testis differentiation. J. Biochem. 2005, 138, 13–19. [Google Scholar] [CrossRef]
- Clarkson, M.J.; Harley, V.R. Sex with two SOX on: SRY and SOX9 in testis development. Trends Endocrinol. Metab. 2002, 13, 106–111. [Google Scholar] [CrossRef]
- Sekido, R.; Lovell-Badge, R. Sex determination and SRY: Down to a wink and a nudge? Trends Genet. 2009, 25, 19–29. [Google Scholar] [CrossRef]
- Albrecht, K.H.; Eicher, E.M. Evidence that Sry is expressed in pre-Sertoli cells and Sertoli and granulosa cells have a common precursor. Dev. Biol. 2001, 240, 92–107. [Google Scholar] [CrossRef] [Green Version]
- Mamsen, L.S.; Ernst, E.H.; Borup, R.; Larsen, A.; Olesen, R.H.; Ernst, E.; Anderson, R.A.; Kristensen, S.G.; Andersen, C.Y. Temporal expression pattern of genes during the period of sex differentiation in human embryonic gonads. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacker, A.; Capel, B.; Goodfellow, P.; Lovell-Badge, R. Expression of Sry, the mouse sex determining gene. Development 1995, 121, 1603–1614. [Google Scholar]
- Wilhelm, D.; Martinson, F.; Bradford, S.; Wilson, M.J.; Combes, A.N.; Beverdam, A.; Bowles, J.; Mizusaki, H.; Koopman, P. Sertoli cell differentiation is induced both cell-autonomously and through prostaglandin signaling during mammalian sex determination. Dev. Biol. 2005, 287, 111–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zheng, M.; Lau, Y.F.C. The sex-determining factors SRY and SOX9 regulate similar target genes and promote testis cord formation during testicular differentiation. Cell Rep. 2014, 8, 723–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji-Hosokawa, A.; Kashimada, K.; Kato, T.; Ogawa, Y.; Nomura, R.; Takasawa, K.; Lavery, R.; Coschiera, A.; Schlessinger, D.; Harley, V.R.; et al. Peptidyl arginine deiminase 2 (Padi2) is expressed in Sertoli cells in a specific manner and regulated by SOX9 during testicular development. Sci. Rep. 2018, 8, 13263. [Google Scholar] [CrossRef]
- Gonen, N.; Quinn, A.; O’Neill, H.C.; Koopman, P.; Lovell-Badge, R. Normal Levels of Sox9 Expression in the Developing Mouse Testis Depend on the TES/TESCO Enhancer, but This Does Not Act Alone. PLoS Genet. 2017, 13, e1006520. [Google Scholar] [CrossRef] [Green Version]
- Croft, B.; Ohnesorg, T.; Hewitt, J.; Bowles, J.; Quinn, A.; Tan, J.; Corbin, V.; Pelosi, E.; van den Bergen, J.; Sreenivasan, R.; et al. Human sex reversal is caused by duplication or deletion of core enhancers upstream of SOX9. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larney, C.; Bailey, T.L.; Koopman, P. Switching on sex: Transcriptional regulation of the testis-determining gene Sry. Development 2014, 141, 2195–2205. [Google Scholar] [CrossRef] [Green Version]
- Lei, N.; Hornbaker, K.I.; Rice, D.A.; Karpova, T.; Agbor, V.A.; Heckert, L.L. Sex-specific differences in mouse DMRT1 expression are both cell type- and stage-dependent during gonad development. Biol. Reprod. 2007, 77, 466–475. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Matsuda, M.; Kajiura-Kobayashi, H.; Suzuki, A.; Saito, N.; Nakamoto, M.; Shibata, N.; Nagahama, Y. Two DM domain genes, DMY and DMRT1, involved in testicular differentiation and development in the Medaka, Oryzias latipes. Dev. Dyn. 2004, 231, 518–526. [Google Scholar] [CrossRef]
- Zhang, J. Evolution of DMY, a Newly Emergent Male Sex-Determination Gene of Medaka Fish. Genetics 2004, 166, 1887–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matson, C.; Zarkower, D. Sex and the singular DM domain: Insights into sexual regulation, evolution and plasticity. Nat. Rev. Genet. 2013, 13, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Long, H.K.; Osterwalder, M.; Welsh, I.C.; Hansen, K.; Davies, J.O.J.; Liu, Y.E.; Koska, M.; Adams, A.T.; Aho, R.; Arora, N.; et al. Loss of Extreme Long-Range Enhancers in Human Neural Crest Drives a Craniofacial Disorder. Cell Stem Cell 2020, 27, 765–783.e14. [Google Scholar] [CrossRef] [PubMed]
- Smyk, M.; Akdemir, K.C.; Stankiewicz, P. SOX9 chromatin folding domains correlate with its real and putative distant cis-regulatory elements. Nucleus 2017, 8, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Bagheri-Fam, S.; Barrionuevo, F.; Dohrmann, U.; Günther, T.; Schüle, R.; Kemler, R.; Mallo, M.; Kanzler, B.; Scherer, G. Long-range upstream and downstream enhancers control distinct subsets of the complex spatiotemporal Sox9 expression pattern. Dev. Biol. 2006, 291, 382–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagheri-Fam, S.; Sreenivasan, R.; Bernard, P.; Knower, K.C.; Sekido, R.; Lovell-Badge, R.; Just, W.; Harley, V.R. Sox9 gene regulation and the loss of the XY/XX sex-determining mechanism in the mole vole Ellobius lutescens. Chromosom. Res. 2012, 20, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Sekido, R.; Lovell-Badge, R. Sex determination involves synergistic action of SRY and SF1 on a specific Sox9 enhancer. Nature 2008, 453, 930–934. [Google Scholar] [CrossRef]
- Bagheri-Fam, S.; Sim, H.; Bernard, P.; Jayakody, I.; Taketo, M.M.; Scherer, G.; Harley, V.R. Loss of Fgfr2 leads to partial XY sex reversal. Dev. Biol. 2008, 314, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Symon, A.; Harley, V. SOX9: A genomic view of tissue specific expression and action. Int. J. Biochem. Cell Biol. 2017, 87, 18–22. [Google Scholar] [CrossRef]
- Georg, I.; Bagheri-Fam, S.; Knower, K.C.; Wieacker, P.; Scherer, G.; Harley, V.R. Mutations of the SRY-responsive enhancer of SOX9 are uncommon in XY gonadal dysgenesis. Sex. Dev. 2010, 4, 321–325. [Google Scholar] [CrossRef]
- Bagheri-Fam, S.; Sinclair, A.H.; Koopman, P.; Harley, V.R. Conserved regulatory modules in the Sox9 testis-specific enhancer predict roles for SOX, TCF/LEF, Forkhead, DMRT, and GATA proteins in vertebrate sex determination. Int. J. Biochem. Cell Biol. 2010, 42, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Ohnesorg, T.; van den Bergen, J.A.; Belluoccio, D.; Shankara-Narayana, N.; Kean, A.M.; Vasilaras, A.; Ewans, L.; Ayers, K.L.; Sinclair, A.H. A duplication in a patient with 46,XX ovo-testicular disorder of sex development refines the SOX9 testis-specific regulatory region to 24 kb. Clin. Genet. 2017, 92, 347–349. [Google Scholar] [CrossRef]
- Benko, S.; Gordon, C.T.; Mallet, D.; Sreenivasan, R.; Thauvin-Robinet, C.; Brendehaug, A.; Thomas, S.; Bruland, O.; David, M.; Nicolino, M.; et al. Disruption of a long distance regulatory region upstream of SOX9 in isolated disorders of sex development. J. Med. Genet. 2011, 48, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.J.; Sock, E.; Buchberger, A.; Just, W.; Denzer, F.; Hoepffner, W.; German, J.; Cole, T.; Mann, J.; Seguin, J.H.; et al. Copy number variation of two separate regulatory regions upstream of SOX9 causes isolated 46,XY or 46,XX disorder of sex development. J. Med. Genet. 2015, 52, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Ohnesorg, T.; Croft, B.; Tan, J.; Sinclair, A.H. Using ROADMAP Data to Identify Enhancers Associated with Disorders of Sex Development. Sex. Dev. 2016, 10, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Gonen, N.; Futtner, C.R.; Wood, S.; Garcia-Moreno, A.; Salamone, I.M.; Samson, S.C.; Sekido, R.; Poulat, F.; Maatouk, D.M.; Lovell-Badge, R. Sex reversal following deletion of a single far upstream enhancer of Sox9. Science 2018, 360, 1469–1473. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, H.; Stadler, H.S.; Martin, J.F.; Ishii, T.M.; Beachy, P.A.; Nakamura, T.; De Crombrugghe, B. Misexpression of Sox9 in mouse limb bud mesenchyme induces polydactyly and rescues hypodactyly mice. Matrix Biol. 2007, 26, 224–233. [Google Scholar] [CrossRef]
- Montero, J.A.; Lorda-Diez, C.I.; Francisco-Morcillo, J.; Chimal-Monroy, J.; Garcia-Porrero, J.A.; Hurle, J.M. Sox9 expression in amniotes: Species-specific differences in the formation of digits. Front. Cell Dev. Biol. 2017, 5, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; De Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [CrossRef]
- Song, H.; Park, K.H. Regulation and function of SOX9 during cartilage development and regeneration. Semin. Cancer Biol. 2020, 67, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.K.; Francis, J.C.; Swain, A. The role of Sox9 in prostate development. Differentiation 2008, 76, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Bulanenkova, S.S.; Snezhkov, E.V.; Akopov, S.B. SOX9 as One of the Central Units of Regulation Axis of Pancreas Embryogenesis and Cancer Progression. Mol. Genet. Microbiol. Virol. 2019, 34, 159–169. [Google Scholar] [CrossRef]
- Harley, V.R.; Lovell-Badge, R.; Goodfellow, P.N. Definition of a consensus DNA binding site for SRY. Nucleic Acids Res. 1994, 22, 1500–1501. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Harley, V.R.; Pontiggia, A.; Goodfellow, P.N.; Lovell-Badge, R.; Bianchi, M.E. SRY, like HMG1, recognizes sharp angles in DNA. EMBO J. 1992, 11, 4497–4506. [Google Scholar] [CrossRef]
- Hou, L.; Srivastava, Y.; Jauch, R. Molecular basis for the genome engagement by Sox proteins. Semin. Cell Dev. Biol. 2017, 63, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Mertin, S.; McDowall, S.G.; Harley, V.R. The DNA-binding specificity of SOX9 and other SOX proteins. Nucleic Acids Res. 1999, 27, 1359–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harley, V.R.; Lovell-Badge, R.; Goodfellow, P.N.; Hextall, P.J. The HMG box of SRY is a calmodulin binding domain. FEBS Lett. 1996, 391, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Harley, V.R.; Layfield, S.; Mitchell, C.L.; Forwood, J.K.; John, A.P.; Briggs, L.J.; McDowall, S.G.; Jans, D.A. Defective importin β recognition and nuclear import of the sex-determining factor SRY are associated with XY sex-reversing mutations. Proc. Natl. Acad. Sci. USA 2003, 100, 7045–7050. [Google Scholar] [CrossRef] [Green Version]
- Sim, H.; Rimmer, K.; Kelly, S.; Ludbrook, L.M.; Clayton, A.H.A.; Harley, V.R. Defective calmodulin-mediated nuclear transport of the sex-determining region of the Y chromosome (SRY) in XY sex reversal. Mol. Endocrinol. 2005, 19, 1884–1892. [Google Scholar] [CrossRef] [Green Version]
- Sim, H.; Argentaro, A.; Czech, D.P.; Bagheri-Fam, S.; Sinclair, A.H.; Koopman, P.; Boizet-Bonhoure, B.; Poulat, F.; Harley, V.R. Inhibition of SRY-calmodulin complex formation induces ectopic expression of ovarian cell markers in developing XY gonads. Endocrinology 2011, 152, 2883–2893. [Google Scholar] [CrossRef] [Green Version]
- McDowall, S.; Argentaro, A.; Ranganathan, S.; Weller, P.; Mertin, S.; Mansour, S.; Tolmie, J.; Harley, V. Functional and structural studies of wild SOX9 and mutations causing campomelic dysplasia. J. Biol. Chem. 1999, 274, 24023–24030. [Google Scholar] [CrossRef] [Green Version]
- Haseeb, A.; Lefebvre, V. The SOXE transcription factors-SOX8, SOX9 and SOX10-share a bi-partite transactivation mechanism. Nucleic Acids Res. 2019, 47, 6917–6931. [Google Scholar] [CrossRef] [PubMed]
- Südbeck, P.; Schmitz, M.L.; Baeuerle, P.A.; Scherer, G. Sex reversal by loss of the C-terminal transactivation domain of human SOX9. Nat. Genet. 1996, 13, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Jankowski, A.; Cheah, K.S.E.; Prabhakar, S.; Jauch, R. SOXE transcription factors form selective dimers on non-compact DNA motifs through multifaceted interactions between dimerization and high-mobility group domains. Sci. Rep. 2015, 5, 10398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.F.; Lefebvre, V. The transcription factors SOX9 and SOX5/SOX6 cooperate genome-wide through super-enhancers to drive chondrogenesis. Nucleic Acids Res. 2015, 43, 8183–8203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, R.C.; Yang, H.; Rockowitz, S.; Larsen, S.B.; Nikolova, M.; Oristian, D.S.; Polak, L.; Kadaja, M.; Asare, A.; Zheng, D.; et al. Pioneer factors govern super-enhancer dynamics in stem cell plasticity and lineage choice. Nature 2015, 521, 366–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, H.; Argentaro, A.; Harley, V.R. Boys, girls and shuttling of SRY and SOX9. Trends Endocrinol. Metab. 2008, 19, 213–222. [Google Scholar] [CrossRef]
- Williams, C.A.C.; Soufi, A.; Pollard, S.M. Post-translational modification of SOX family proteins: Key biochemical targets in cancer? Semin. Cancer Biol. 2020, 67, 30–38. [Google Scholar] [CrossRef]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef] [Green Version]
- Voldoire, E.; Brunet, F.; Naville, M.; Volff, J.N.; Galiana, D. Expansion by whole genome duplication and evolution of the sox gene family in teleost fish. PLoS ONE 2017, 12, e180936. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Marí, A.; Yan, Y.L.; BreMiller, R.A.; Wilson, C.; Cañestro, C.; Postlethwait, J.H. Characterization and expression pattern of zebrafish anti-Müllerian hormone (amh) relative to sox9a, sox9b, and cyp19a1a, during gonad development. Gene Expr. Patterns 2005, 5, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, M.; Suzuki, A.; Matsuda, M.; Nagahama, Y.; Shibata, N. Testicular type Sox9 is not involved in sex determination but might be in the development of testicular structures in the medaka, Oryzias latipes. Biochem. Biophys. Res. Commun. 2005, 333, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Aoki, Y.; Saito, D.; Kuroki, Y.; Fujiyama, A.; Naruse, K.; Tanaka, M. Sox9b/sox9a2-EGFP transgenic medaka reveals the morphological reorganization of the gonads and a common precursor of both the female and male supporting cells. Mol. Reprod. Dev. 2008, 75, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Wallis, M.C.; Waters, P.D.; Delbridge, M.L.; Kirby, P.J.; Pask, A.J.; Grützner, F.; Rens, W.; Ferguson-Smith, M.A.; Graves, J.A.M. Sex determination in platypus and echidna: Autosomal location of SOX3 confirms the absence of SRY from monotremes. Chromosom. Res. 2007, 15, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Capel, B. Vertebrate sex determination: Evolutionary plasticity of a fundamental switch. Nat. Rev. Genet. 2017, 18, 675–689. [Google Scholar] [CrossRef]
- Kent, J.; Wheatley, S.C.; Andrews, J.E.; Sinclair, A.H.; Koopman, P. A male-specific role for SOX9 in vertebrate sex determination. Development 1996, 122, 2813–2822. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Zuckerkandl, E.; Pauling, L. Evolutionary Divergence and Convergence in Proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Foster, J.W.; Dominguez-Steglich, M.A.; Guioli, S.; Kwok, C.; Weller, P.A.; Stevanović, M.; Weissenbach, J.; Mansour, S.; Young, I.D.; Goodfellow, P.N.; et al. Campomelic dysplasia and autosomal sex reversal caused by mutations in an SRY-related gene. Nature 1994, 372, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Barrionuevo, F.; Bagheri-Fam, S.; Klattig, J.; Kist, R.; Taketo, M.M.; Englert, C.; Scherer, G. Homozygous inactivation of Sox9 causes complete XY sex reversal in mice. Biol. Reprod. 2006, 74, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Wang, S.; Ning, Y.; Lamb, A.N.; Bartley, J. Autosomal XX sex reversal caused by duplication of SOX9. Am. J. Med. Genet. 1999, 87, 349–353. [Google Scholar] [CrossRef]
- Bishop, C.E.; Whitworth, D.J.; Qin, Y.; Agoulnik, A.I.; Agoulnik, I.U.; Harrison, W.R.; Behringer, R.R.; Overbeek, P.A. A transgenic insertion upstream of Sox9 is associated with dominant XX sex reversal in the mouse. Nat. Genet. 2000, 26, 490–494. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, Y.; Wang, C.; Hua, X.; Zhang, X.A.; Yan, J. Sox9-related signaling controls zebrafish juvenile ovary-testis transformation. Cell Death Dis. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, T.; Zhou, L.Y.; Chaudhari, A.; Iguchi, T.; Nagahama, Y. Dmy initiates masculinity by altering Gsdf/Sox9a2/Rspo1 expression in medaka (Oryzias latipes). Sci. Rep. 2016, 6, 19480. [Google Scholar] [CrossRef] [PubMed]
- Lambeth, L.; Raymond, C.S.; Roeszler, K.N.; Kuroiwa, A.; Nakata, T.; Zarkower, D.; Smith, C.A. Over-expression of DMRT1 induces the male pathway in embryonic chicken gonads. Dev. Biol. 2014, 389, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Mendoza, N.; Harley, V.R.; Merchant-Larios, H. Differential expression of SOX9 in gonads of the sea turtle Lepidochelys olivacea at male- or female-promoting temperatures. J. Exp. Zool. 1999, 284, 705–710. [Google Scholar] [CrossRef]
- Torres-Maldonado, L.; Moreno-Mendoza, N.; Landa, A.; Merchant-Larios, H. Timing of SOX9 downregulation and female sex determination in gonads of the sea turtle Lepidochelys olivacea. J. Exp. Zool. 2001, 290, 498–503. [Google Scholar] [CrossRef]
- Rhen, T.; Metzger, K.; Schroeder, A.; Woodward, R. Expression of putative sex-determining genes during the thermosensitive period of gonad development in the snapping turtle, Chelydra serpentina. Sex. Dev. 2007, 1, 255–270. [Google Scholar] [CrossRef]
- Sun, W.; Cai, H.; Zhang, G.; Zhang, H.; Bao, H.; Wang, L.; Ye, J.; Qian, G.; Ge, C. Dmrt1 is required for primary male sexual differentiation in Chinese soft-shelled turtle Pelodiscus sinensis. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Sun, W.; Cai, H.; Bao, H.; Zhang, Y.; Qian, G.; Ge, C. The role of anti-Müllerian hormone in testis differentiation reveals the significance of the TGF-b pathway in reptilian sex determination. Genetics 2019, 213, 1317–1327. [Google Scholar] [CrossRef]
- Piprek, R.P.; Damulewicz, M.; Kloc, M.; Kubiak, J.Z. Transcriptome analysis identifies genes involved in sex determination and development of Xenopus laevis gonads. Differentiation 2018, 100, 46–56. [Google Scholar] [CrossRef] [PubMed]
- El Jamil, A.; Kanhoush, R.; Magre, S.; Boizet-Bonhoure, B.; Penrad-Mobayed, M. Sex-specific expression of SOX9 during gonadogenesis in the amphibian Xenopus tropicalis. Dev. Dyn. 2008, 237, 2996–3005. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, A.; Smith, C.; Western, P.; McClive, P. A comparative analysis of vertebrate sex determination. Novartis Found. Symp. 2002, 244, 102–114. [Google Scholar] [CrossRef]
- Moreno-Mendoza, N.; Harley, V.R.; Merchant-Larios, H. Temperature regulates SOX9 expression in cultured gonads of Lepidochelys olivacea, a species with temperature sex determination. Dev. Biol. 2001, 229, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Oreal, E.; Pieau, C.; Mattei, M.G.; Josso, N.; Picard, J.Y.; Carré-Eusèbe, D.; Magre, S. Early expression of AMH in chicken embryonic gonads precedes testicular SOX9 expression. Dev. Dyn. 1998, 212, 522–532. [Google Scholar] [CrossRef]
- Yamashita, S.; Kataoka, K.; Yamamoto, H.; Kato, T.; Hara, S.; Yamaguchi, K.; Renard-Guillet, C.; Katou, Y.; Shirahige, K.; Ochi, H.; et al. Comparative analysis demonstrates cell type-specific conservation of SOX9 targets between mouse and chicken. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Estermann, M.A.; Williams, S.; Hirst, C.E.; Roly, Z.Y.; Serralbo, O.; Adhikari, D.; Powell, D.; Major, A.T.; Smith, C.A. Insights into Gonadal Sex Differentiation Provided by Single-Cell Transcriptomics in the Chicken Embryo. Cell Rep. 2020, 31, 107491. [Google Scholar] [CrossRef]
- Parma, P.; Veyrunes, F.; Pailhoux, E. Sex Reversal in Non-Human Placental Mammals. Sex. Dev. 2016, 10, 326–344. [Google Scholar] [CrossRef] [PubMed]
- Kist, R.; Schrewe, H.; Balling, R.; Scherer, G. Conditional inactivation of Sox9: A mouse model for campomelic dysplasia. Genesis 2002, 32, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Seymour, P.A.; Freude, K.K.; Dubois, C.L.; Shih, H.-P.; Patel, N.A.; Sander, M. A Dosage-Dependent Requirement for SOX9 in Pancreatic Endocrine Cell Formation. Dev. Biol. 2008, 323, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Unger, S.; Scherer, G.; Superti-Furga, A. Campomelic Dysplasia. In GeneReviews® [Internet]; Adam, M., Ardinger, H., Pagon, R., Al, E., Eds.; University of Washington: Seattle, WA, USA, 2008. [Google Scholar]
- Piper, K.; Ball, S.G.; Keeling, J.W.; Mansoor, S.; Wilson, D.I.; Hanley, N.A. Novel SOX9 expression during human pancreas development correlates to abnormalities in Campomelic dysplasia. Mech. Dev. 2002, 116, 223–226. [Google Scholar] [CrossRef]
- Hughes, I.A.; Houk, C.; Ahmed, S.F.; Lee, P.A.; LWPES Consensus Group; ESPE Consensus Group. Consensus statement on management of intersex disorders. Arch. Dis. Child 2006, 91, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Nordenvall, A.S.; Frisén, L.; Nordenström, A.; Lichtenstein, P.; Nordenskjöld, A. Population based nationwide study of hypospadias in Sweden, 1973 to 2009: Incidence and risk factors. J. Urol. 2014, 191, 783–789. [Google Scholar] [CrossRef]
- Matzuk, M.M.; Lamb, D.J. The biology of infertility: Research advances and clinical challenges. Nat. Med. 2008, 14, 1197–1213. [Google Scholar] [CrossRef]
- Eggers, S.; Sadedin, S.; van den Bergen, J.A.; Robevska, G.; Ohnesorg, T.; Hewitt, J.; Lambeth, L.; Bouty, A.; Knarston, I.M.; Tan, T.Y.; et al. Disorders of sex development: Insights from targeted gene sequencing of a large international patient cohort. Genome Biol. 2016, 17, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, S.; Guo, S.; Liu, L.; Yang, X.; Liang, H. Functional study of a novel c.630delG (p.Y211Tfs*85) mutation in NR5A1 gene in a Chinese boy with 46,XY disorders of sex development. J. Assist. Reprod. Genet. 2020, 37, 477–486. [Google Scholar] [CrossRef] [PubMed]
- León, N.Y.; Reyes, A.P.; Harley, V.R. A clinical algorithm to diagnose differences of sex development. Lancet Diabetes Endocrinol. 2019, 7, 560–574. [Google Scholar] [CrossRef]
- Wagner, T.; Wirth, J.; Meyer, J.; Zabel, B.; Held, M.; Zimmer, J.; Pasantes, J.; Bricarelli, F.D.; Keutel, J.; Hustert, E.; et al. Autosomal sex reversal and campomelic dysplasia are caused by mutations in and around the SRY-related gene SOX9. Cell 1994, 79, 1111–1120. [Google Scholar] [CrossRef]
- Mansour, S.; Hall, C.M.; Pembrey, M.E.; Young, I.D. A clinical and genetic study of campomelic dysplasia. J. Med. Genet. 1995, 32, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, V.; Dvir-Ginzberg, M. SOX9 and the many facets of its regulation in the chondrocyte lineage. Connect. Tissue Res. 2017, 58, 2–14. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, H.; Liu, Z.; Liu, C.; Luan, J.; Zhou, X.; Han, J. A systematic review of genetic skeletal disorders reported in Chinese biomedical journals between 1978 and 2012. Orphanet J. Rare Dis. 2012, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel-Calemard, L.; Lesca, G.; Morel, Y.; Boggio, D.; Plauchu, H.; Attia-Sobo, J. Campomelic acampomelic dysplasia presenting with increased nuchal translucency in the first trimester. Prenat. Diagn. 2004, 24, 519–523. [Google Scholar] [CrossRef]
- Thong, M.K.; Scherer, G.; Kozlowski, K.; Haan, E.; Morris, L. Brief clinical report: Acampomelic campomelic dysplasia with SOX9 mutation. Am. J. Med. Genet. 2000, 93, 421–425. [Google Scholar] [CrossRef]
- Wada, Y.; Nishimura, G.; Nagai, T.; Sawai, H.; Yoshikata, M.; Miyagawa, S.; Hanita, T.; Sato, S.; Hasegawa, T.; Ishikawa, S.; et al. Mutation analysis of SOX9 and single copy number variant analysis of the upstream region in eight patients with campomelic dysplasia and acampomelic campomelic dysplasia. Am. J. Med. Genet. Part A 2009, 149A, 2882–2885. [Google Scholar] [CrossRef] [PubMed]
- Massardier, J.; Roth, P.; Michel-Calemard, L.; Rudigoz, R.C.; Bouvier, R.; Dijoud, F.; Arnould, P.; Combourieu, D.; Gaucherand, P. Campomelic dysplasia: Echographic suspicion in the first trimester of pregnancy and final diagnosis of two cases. Fetal Diagn. Ther. 2009, 24, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Kitoh, H.; Kaneko, H.; Mishima, K.; Kadono, I.; Ishiguro, N.; Nishimura, G. A novel SOX9 H169Q mutation in a family with overlapping phenotype of mild campomelic dysplasia and small patella syndrome. Am. J. Med. Genet. A 2013, 161A, 2528–2534. [Google Scholar] [CrossRef]
- Tonni, G.; Ventura, A.; Pattacini, P.; Bonasoni, M.P.; Baffico, A.M. p.His165Pro: A novel SOX9 missense mutation of campomelic dysplasia. J. Obstet. Gynaecol. Res. 2013, 39, 1085–1091. [Google Scholar] [CrossRef]
- Staffler, A.; Hammel, M.; Wahlbuhl, M.; Bidlingmaier, C.; Flemmer, A.W.; Pagel, P.; Nicolai, T.; Wegner, M.; Holzinger, A. Heterozygous SOX9 mutations allowing for residual DNA-binding and transcriptional activation lead to the acampomelic variant of campomelic dysplasia. Hum. Mutat. 2010, 31, 1436–1444. [Google Scholar] [CrossRef]
- Karaer, K.; Yüksel, Z.; Yalinbaş, E.; Scherer, G. A case of campomelic dysplasia in whom a new mutation was found in the SOX9 gene. Turk. Pediatr. Ars. 2014, 49, 154–156. [Google Scholar] [CrossRef]
- Preiss, S.; Argentaro, A.; Clayton, A.; John, A.; Jans, D.A.; Ogata, T.; Nagai, T.; Barroso, I.; Schafer, A.J.; Harley, V.R. Compound Effects of Point Mutations Causing Campomelic Dysplasia/Autosomal Sex Reversal upon SOX9 Structure, Nuclear Transport, DNA Binding, and Transcriptional Activation. J. Biol. Chem. 2001, 276, 27864–27872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shotelersuk, V.; Jaruratanasirikul, S.; Sinthuwiwat, T.; Janjindamai, W. A novel nonsense mutation, E150X, in the SOX9 gene underlying campomelic dysplasia. Genet. Mol. Biol. 2006, 29, 617–620. [Google Scholar] [CrossRef] [Green Version]
- Barone, C.; Bartoloni, G.; Baffico, A.M.; Pappalardo, E.; Mura, I.; Ettore, G.; Bianca, S. Novel c.358C>T mutation of SOX9 gene in prenatal diagnosis of campomelic dysplasia. Congenit. Anom. (Kyoto) 2014, 54, 193–194. [Google Scholar] [CrossRef] [Green Version]
- Kwok, C.; Weller, P.A.; Guioli, S.; Foster, J.W.; Mansour, S.; Zuffardi, O.; Punnett, H.H.; Dominguez-Steglich, M.A.; Brook, J.D.; Young, I.D.; et al. Mutations in SOX9, the gene responsible for campomelic dysplasia and autosomal sex reversal. Am. J. Hum. Genet. 1995, 57, 1028–1036. [Google Scholar] [PubMed]
- Gentilin, B.; Forzano, F.; Bedeschi, M.F.; Rizzuti, T.; Faravelli, F.; Izzi, C.; Lituania, M.; Rodriguez-Perez, C.; Bondioni, M.P.; Savoldi, G.; et al. Phenotype of five cases of prenatally diagnosed campomelic dysplasia harboring novel mutations of the SOX9 gene. Ultrasound Obstet. Gynecol. 2010, 36, 315–323. [Google Scholar] [CrossRef]
- Sock, E.; Pagon, R.A.; Keymolen, K.; Lissens, W.; Wegner, M.; Scherer, G. Loss of DNA-dependent dimerization of the transcription factor SOX9 as a cause for campomelic dysplasia. Hum. Mol. Genet. 2003, 12, 1439–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenouchi, T.; Matsuzaki, Y.; Yamamoto, K.; Kosaki, K.; Torii, C.; Takahashi, T.; Kosaki, K. SOX9 dimerization domain mutation mimicking type 2 collagen disorder phenotype. Eur. J. Med. Genet. 2014, 57, 298–301. [Google Scholar] [CrossRef]
- Gogi, K.; Nishijima, E.; Tsugawa, C.; Nishio, H.; Pokharel, R.K.; Matsuo, M. Novel Missense Mutation in the HMG Box of SOX9 Gene in a Japanese XY Male Resulted in Campomelic Dysplasia and Severe Defect in Masculinization. Hum. Mutat. 1998, 116, 114–116. [Google Scholar]
- Posey, J.E.; Harel, T.; Liu, P.; Rosenfeld, J.A.; James, R.A.; Coban Akdemir, Z.H.; Walkiewicz, M.; Bi, W.; Xiao, R.; Ding, Y.; et al. Resolution of Disease Phenotypes Resulting from Multilocus Genomic Variation. N. Engl. J. Med. 2017, 376, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Pop, R.; Zaragoza, M.V.; Gaudette, M.; Dohrmann, U.; Scherer, G. A homozygous nonsense mutation in SOX9 in the dominant disorder campomelic dysplasia: A case of mitotic gene conversion. Hum. Genet. 2005, 117, 43–53. [Google Scholar] [CrossRef]
- Katoh-Fukui, Y.; Igarashi, M.; Nagasaki, K.; Horikawa, R.; Nagai, T.; Tsuchiya, T.; Suzuki, E.; Miyado, M.; Hata, K.; Nakabayashi, K.; et al. Testicular dysgenesis/regression without campomelic dysplasia in patients carrying missense mutations and upstream deletion of SOX9. Mol. Genet. Genomic Med. 2015, 3, 550–557. [Google Scholar] [CrossRef]
- Stoeva, R.; Grozdanova, L.; Scherer, G.; Krasteva, M.; Bausch, E.; Krastev, T.; Linev, A.; Stefanova, M. A novel SOX9 nonsense mutation, Q401X, in a case of campomelic dysplasia with XY sex reversal. Genet. Couns. 2011, 22, 49–53. [Google Scholar]
- Meyer, J.; Südbeck, P.; Held, M.; Wagner, T.; Schmitz, M.L.; Bricarelli, F.D.; Eggermont, E.; Friedrich, U.; Haas, O.A.; Kobelt, A.; et al. Mutational analysis of the SOX9 gene in campomelic dysplasia and autosomal sex reversal: Lack of genotype/phenotype correlations. Hum. Mol. Genet. 1997, 6, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattos, E.P.; Sanseverino, M.T.; Magalhães, J.A.A.; Leite, J.C.L.; Félix, T.M.; Todeschini, L.A.; Cavalcanti, D.P.; Schüler-Faccini, L. Clinical and molecular characterization of a Brazilian cohort of campomelic dysplasia patients, and identification of seven new SOX9 mutations. Genet. Mol. Biol. 2015, 38, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, I.A. Disorders of sex development: A new definition and classification. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.A.; Houk, C.P.; Ahmed, S.F.; Hughes, I.A.; International Consensus Conference on Intersex organized by the Lawson Wilkins Pediatric Endocrine Society and the European Society for Paediatric Endocrinology. Consensus statement on management of intersex disorders. Pediatrics 2006, 118, e488–e500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkauskas, G.; Jaubert, F.; Lortat-Jacob, S.; Malan, V.; Thibaud, E.; Nihoul-Fékété, C. The long-term followup of 33 cases of true hermaphroditism: A 40-year experience with conservative gonadal surgery. J. Urol. 2007, 177, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.M.; Ko, J.M.; Shin, C.H.; Yang, S.W. A Korean boy with 46,XX testicular disorder of sex development caused by SOX9 duplication. Ann. Pediatr. Endocrinol. Metab. 2014, 19, 108–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Hernández, B.; Méndez, J.P.; Coral-Vázquez, R.M.; Benítez-Granados, J.; Zenteno, J.C.; Villegas-Ruiz, V.; Calzada-León, R.; Soderlund, D.; Canto, P. Duplication of SOX9 associated with 46,XX ovotesticular disorder of sex development. Reprod. Biomed. Online 2018, 37, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.J.; Willatt, L.; Homfray, T.; Woods, C.G. A SOX9 Duplication and Familial 46,XX Developmental Testicular Disorder. N. Engl. J. Med. 2011, 364, 91–93. [Google Scholar] [CrossRef] [Green Version]
- Hyon, C.; Chantot-Bastaraud, S.; Harbuz, R.; Bhouri, R.; Perrot, N.; Peycelon, M.; Sibony, M.; Rojo, S.; Piguel, X.; Bilan, F.; et al. Refining the regulatory region upstream of SOX9 associated with 46,XX testicular Disorders of Sex Development (DSD). Am. J. Med. Genet. Part A 2015, 167A, 1851–1858. [Google Scholar] [CrossRef] [Green Version]
- Vetro, A.; Dehghani, M.R.; Kraoua, L.; Giorda, R.; Beri, S.; Cardarelli, L.; Merico, M.; Manolakos, E.; Parada-Bustamante, A.; Castro, A.; et al. Testis development in the absence of SRY: Chromosomal rearrangements at SOX9 and SOX3. Eur. J. Hum. Genet. 2015, 23, 1025–1032. [Google Scholar] [CrossRef]
- Vetro, A.; Ciccone, R.; Giorda, R.; Patricelli, M.G.; Mina, E.D.; Forlino, A.; Zuffardi, O. XX males SRY negative: A confirmed cause of infertility. J. Med. Genet. 2011, 48, 710–712. [Google Scholar] [CrossRef]
- Lybæk, H.; de Bruijn, D.; den Engelsman-van Dijk, A.H.A.; Vanichkina, D.; Nepal, C.; Brendehaug, A.; Houge, G. RevSex duplication-induced and sex-related differences in the SOX9 regulatory region chromatin landscape in human fibroblasts. Epigenetics 2013, 9, 416–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Refai, O.; Friedman, A.; Terry, L.; Jewett, T.; Pearlman, A.; Perle, M.A.; Ostrer, H. De novo 12;17 translocation upstream of SOX9 resulting in 46,XX testicular disorder of sex development. Am. J. Med. Genet. A 2010, 152A, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Vidal, V.P.I.; Chaboissier, M.C.; de Rooij, D.G.; Schedl, A. Sox9 induces testis development in XX transgenic mice. Nat. Genet. 2001, 28, 216–217. [Google Scholar] [CrossRef]
- Bagheri-Fam, S.; Combes, A.N.; Ling, C.K.; Wilhelm, D. Heterozygous deletion of Sox9 in mouse mimics the gonadal sex reversal phenotype associated with campomelic dysplasia in humans. Hum. Mol. Genet. 2021, 29, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Ushijima, K.; Ogawa, Y.; Asakura, Y.; Muroya, K.; Hayashi, M.; Ishii, T.; Hasegawa, T.; Takada, S.; Narumi, S.; Sekido, R.; et al. Identification of the first promoter-specific gain-of-function SOX9 missense variant (p.E50K) in a patient with 46, XX ovotesticular disorder of sex development. Am. J. Hum. Genet. A 2021, 1–9. [Google Scholar] [CrossRef]
- Vidal, V.P.I.; Chaboissier, M.C.; Lützkendorf, S.; Cotsarelis, G.; Mill, P.; Hui, C.C.; Ortonne, N.; Ortonne, J.P.; Schedl, A. Sox9 is essential for outer root sheath differentiation and the formation of the hair stem cell compartment. Curr. Biol. 2005, 15, 1340–1351. [Google Scholar] [CrossRef] [Green Version]
- Fantauzzo, K.A.; Kurban, M.; Levy, B.; Christiano, A.M. Trps1 and Its Target Gene Sox9 Regulate Epithelial Proliferation in the Developing Hair Follicle and Are Associated with Hypertrichosis. PLoS Genet. 2012, 8, e1003002. [Google Scholar] [CrossRef] [Green Version]
- Bashamboo, A.; McElreavey, K. Human sex-determination and disorders of sex-development (DSD). Semin. Cell Dev. Biol. 2015, 45, 77–83. [Google Scholar] [CrossRef]
- Lee, P.A.; Nordenström, A.; Houk, C.P.; Ahmed, S.F.; Auchus, R.; Baratz, A.; Baratz Dalke, K.; Liao, L.M.; Lin-Su, K.; Looijenga, L.H.J.; et al. Global disorders of sex development update since 2006: Perceptions, approach and care. Horm. Res. Paediatr. 2016, 85, 158–180. [Google Scholar] [CrossRef]
- de Vries, A.L.C.; Roehle, R.; Marshall, L.; Frisén, L.; van de Grift, T.C.; Kreukels, B.P.C.; Bouvattier, C.; Köhler, B.; Thyen, U.; Nordenström, A.; et al. Mental Health of a Large Group of Adults with Disorders of Sex Development in Six European Countries. Psychosom. Med. 2019, 81, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, P.S.; Buehr, M.; McLaren, A. XY follicle cells in ovaries of XX ⇆ XY female mouse chimaeras. Development 1988, 104, 683–688. [Google Scholar] [PubMed]
- Georges, A.; L’Hôte, D.; Todeschini, A.L.; Auguste, A.; Legois, B.; Zider, A.; Veitia, R.A. The transcription factor FOXL2 mobilizes estrogen signaling to maintain the identity of ovarian granulosa cells. Elife 2014, 3, e04207. [Google Scholar] [CrossRef] [Green Version]
- Vainio, S.; Heikkilä, M.; Kispert, A.; Chin, N.; McMahon, A.P. Female development in mammals is regulated by Wnt-4 signalling. Nature 1999, 397, 405–409. [Google Scholar] [CrossRef]
- Parma, P.; Radi, O.; Vidal, V.; Chaboissier, M.C.; Dellambra, E.; Valentini, S.; Guerra, L.; Schedl, A.; Camerino, G. R-spondin1 is essential in sex determination, skin differentiation and malignancy. Nat. Genet. 2006, 38, 1304–1309. [Google Scholar] [CrossRef]
- Ottolenghi, C.; Pelosi, E.; Tran, J.; Colombino, M.; Douglass, E.; Nedorezov, T.; Cao, A.; Forabosco, A.; Schlessinger, D. Loss of Wnt4 and Foxl2 leads to female-to-male sex reversal extending to germ cells. Hum. Mol. Genet. 2007, 16, 2795–2804. [Google Scholar] [CrossRef] [Green Version]
- Bernard, P.; Ryan, J.; Sim, H.; Czech, D.P.; Sinclair, A.H.; Koopman, P.; Harley, V.R. Wnt signaling in ovarian development inhibits Sf1 activation of Sox9 via the Tesco enhancer. Endocrinology 2012, 153, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Barrionuevo, F.J.; Hurtado, A.; Kim, G.J.; Real, F.M.; Bakkali, M.; Kopp, J.L.; Sander, M.; Scherer, G.; Burgos, M.; Jiménez, R. Sox9 and Sox8 protect the adult testis from male-to-female genetic reprogramming and complete degeneration. Elife 2016, 5, e15635. [Google Scholar] [CrossRef] [Green Version]
- Georg, I.; Barrionuevo, F.; Wiech, T.; Scherer, G. Sox9 and Sox8 are required for basal lamina integrity of testis cords and for suppression of FOXL2 during embryonic testis development in mice. Biol. Reprod. 2012, 87, 99. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphological changes of the differentiating mouse testis: Between embryonic day (E)8.75 and E9.5, germ cells migrate dorsally towards the developing genital ridge. The testis-determining factor SRY activates Sox9 expression to drive Sertoli cell (blue) differentiation from E10.5. Proliferating Sertoli cells begin to compartmentalize to form testis cords around E12.5. Following Sertoli cell proliferation, fetal Leydig cells (purple) and peritubular myoid cells (green) differentiate. By E15.5, the majority of testis cell types have differentiated, and a distinct mouse testis has formed with typical testis cord structure and interstitial space. The Sertoli cells and peritubular myoid cells secrete various extracellular matrix (ECM) proteins to form the basal lamina, which surrounds the testis cords and maintains their structural integrity. The testis cords are composed of mitotically arrested germ cells enclosed by Sertoli cells, with an outer layer of peritubular myoid cells and extracellular matrix. The interstitial space comprises steroidogenic fetal Leydig cells, mesenchyme, and a prominent blood vasculature.
Figure 1.
Morphological changes of the differentiating mouse testis: Between embryonic day (E)8.75 and E9.5, germ cells migrate dorsally towards the developing genital ridge. The testis-determining factor SRY activates Sox9 expression to drive Sertoli cell (blue) differentiation from E10.5. Proliferating Sertoli cells begin to compartmentalize to form testis cords around E12.5. Following Sertoli cell proliferation, fetal Leydig cells (purple) and peritubular myoid cells (green) differentiate. By E15.5, the majority of testis cell types have differentiated, and a distinct mouse testis has formed with typical testis cord structure and interstitial space. The Sertoli cells and peritubular myoid cells secrete various extracellular matrix (ECM) proteins to form the basal lamina, which surrounds the testis cords and maintains their structural integrity. The testis cords are composed of mitotically arrested germ cells enclosed by Sertoli cells, with an outer layer of peritubular myoid cells and extracellular matrix. The interstitial space comprises steroidogenic fetal Leydig cells, mesenchyme, and a prominent blood vasculature.

Figure 2.
SOX9 upstream regulatory region towards KCNJ2 gene. Green regions represent the enhancers that control SOX9 expression within the testes. Yellow boxes behind the structure indicate regions commonly associated with PRS (Pierre Robin Syndrome) and craniofacial formation, or CMPD (Campomelic Dysplasia) and chondrogenesis. Enhancers eALDI and hTESCO are calculated from Croft et al., [48]. Sox9up7 is identified from bioinformatic analysis by Ohnesorg et al., 2016 [66].
Figure 2.
SOX9 upstream regulatory region towards KCNJ2 gene. Green regions represent the enhancers that control SOX9 expression within the testes. Yellow boxes behind the structure indicate regions commonly associated with PRS (Pierre Robin Syndrome) and craniofacial formation, or CMPD (Campomelic Dysplasia) and chondrogenesis. Enhancers eALDI and hTESCO are calculated from Croft et al., [48]. Sox9up7 is identified from bioinformatic analysis by Ohnesorg et al., 2016 [66].

Figure 3.
Human protein structure: including the dimerization domain (DIM), DNA-binding HMG-box, transactivating domain in the middle of the protein (TAM), the proline, glutamine, and alanine rich region (PQA) and the proline, glutamine, and serine rich region (PQS), both required for transactivation. Adapted from Symon & Harley, 2017 [60].
Figure 3.
Human protein structure: including the dimerization domain (DIM), DNA-binding HMG-box, transactivating domain in the middle of the protein (TAM), the proline, glutamine, and alanine rich region (PQA) and the proline, glutamine, and serine rich region (PQS), both required for transactivation. Adapted from Symon & Harley, 2017 [60].

Figure 4.
Multiple sequence alignment of SOX9 amino acid sequences across 28 species, compared to human SOX9 reference. Human SOX9 protein along the top of the multiple sequence alignment, indicating the dimerization domain (DIM), DNA-binding HMG-box, transactivating domain in the middle of the protein (TAM), the proline, glutamine, and alanine rich region (PQA) and the proline, glutamine, and serine rich region (PQS). In the multiple sequence alignment, grey indicates identical sequence to human SOX9 at each residue, red indicates different residue, and blue indicates an insertion. Images created through NCBI COBALT (Constraint-based multiple alignment tool) with 28 sequences selected through the NCBI Orthologues feature, NCBI Multiple Alignment Sequence Alignment Viewer, Version 1.19.1 (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene/6662 (accessed on 16 February 2021)).
Figure 4.
Multiple sequence alignment of SOX9 amino acid sequences across 28 species, compared to human SOX9 reference. Human SOX9 protein along the top of the multiple sequence alignment, indicating the dimerization domain (DIM), DNA-binding HMG-box, transactivating domain in the middle of the protein (TAM), the proline, glutamine, and alanine rich region (PQA) and the proline, glutamine, and serine rich region (PQS). In the multiple sequence alignment, grey indicates identical sequence to human SOX9 at each residue, red indicates different residue, and blue indicates an insertion. Images created through NCBI COBALT (Constraint-based multiple alignment tool) with 28 sequences selected through the NCBI Orthologues feature, NCBI Multiple Alignment Sequence Alignment Viewer, Version 1.19.1 (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene/6662 (accessed on 16 February 2021)).

Figure 6.
Human SOX9 protein, indicating the dimerization domain (DIM), DNA-binding HMG-box, transactivating domain in the middle of the protein (TAM), the proline, glutamine, and alanine rich region (PQA) and the proline, glutamine, and serine rich region (PQS). Dashes underneath indicate missense single base-pair mutations causing a triplet change and subsequent amino acid change, causing various phenotypes [82,101,128,131,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156].
Figure 6.
Human SOX9 protein, indicating the dimerization domain (DIM), DNA-binding HMG-box, transactivating domain in the middle of the protein (TAM), the proline, glutamine, and alanine rich region (PQA) and the proline, glutamine, and serine rich region (PQS). Dashes underneath indicate missense single base-pair mutations causing a triplet change and subsequent amino acid change, causing various phenotypes [82,101,128,131,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156].

Figure 7.
Molecular mechanisms of mammalian sex determination leading to male or female supporting cell fate.
Figure 7.
Molecular mechanisms of mammalian sex determination leading to male or female supporting cell fate.


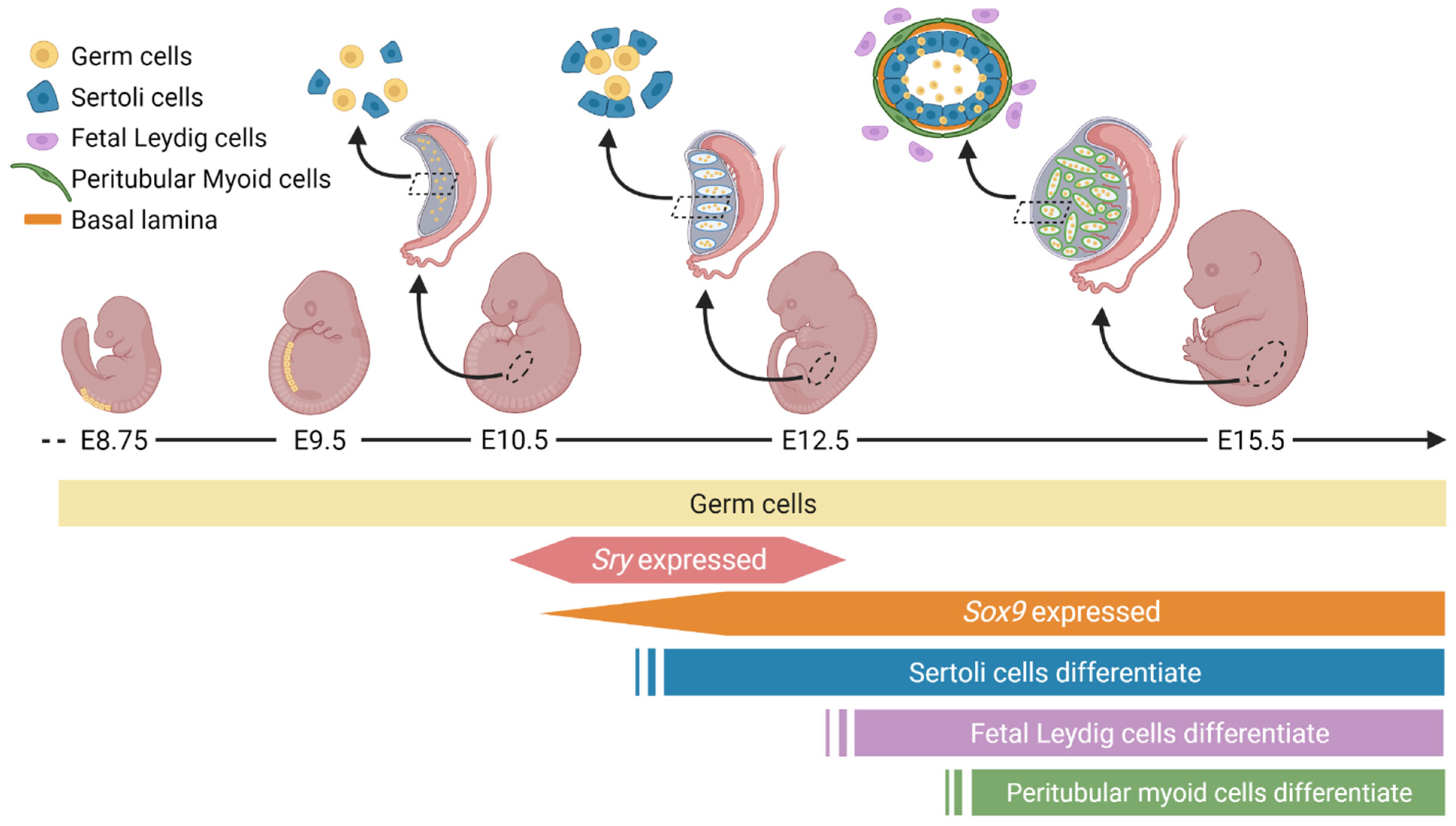{kind=link}
{kind=link}
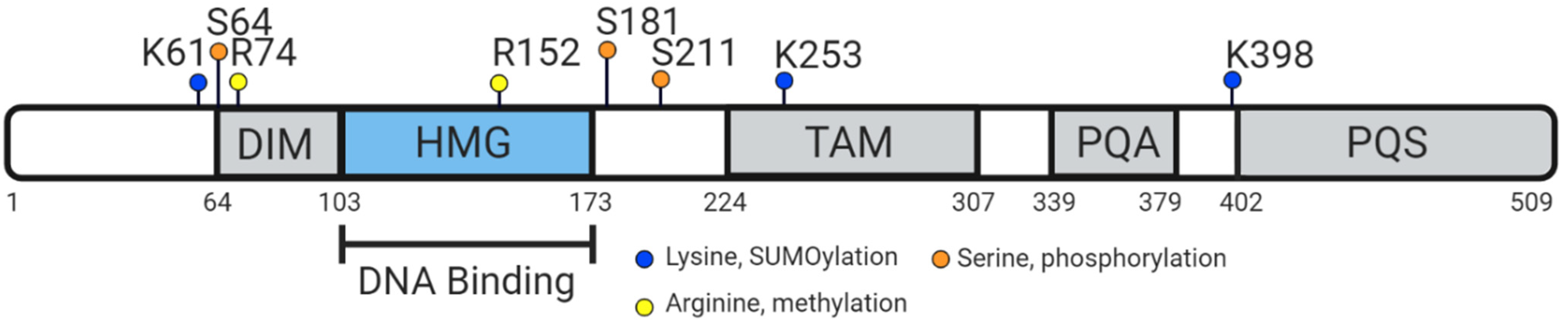{kind=link}
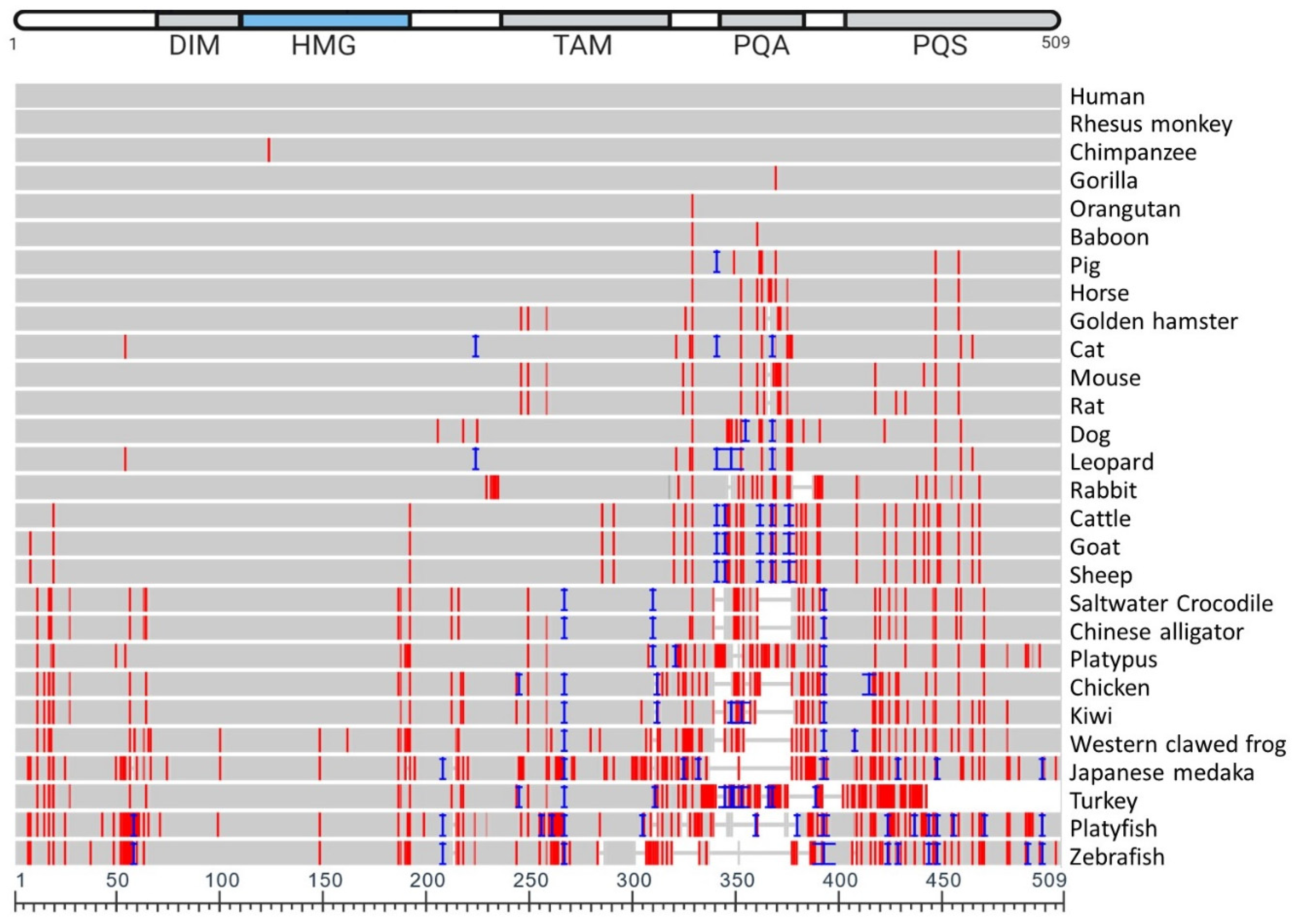{kind=link}
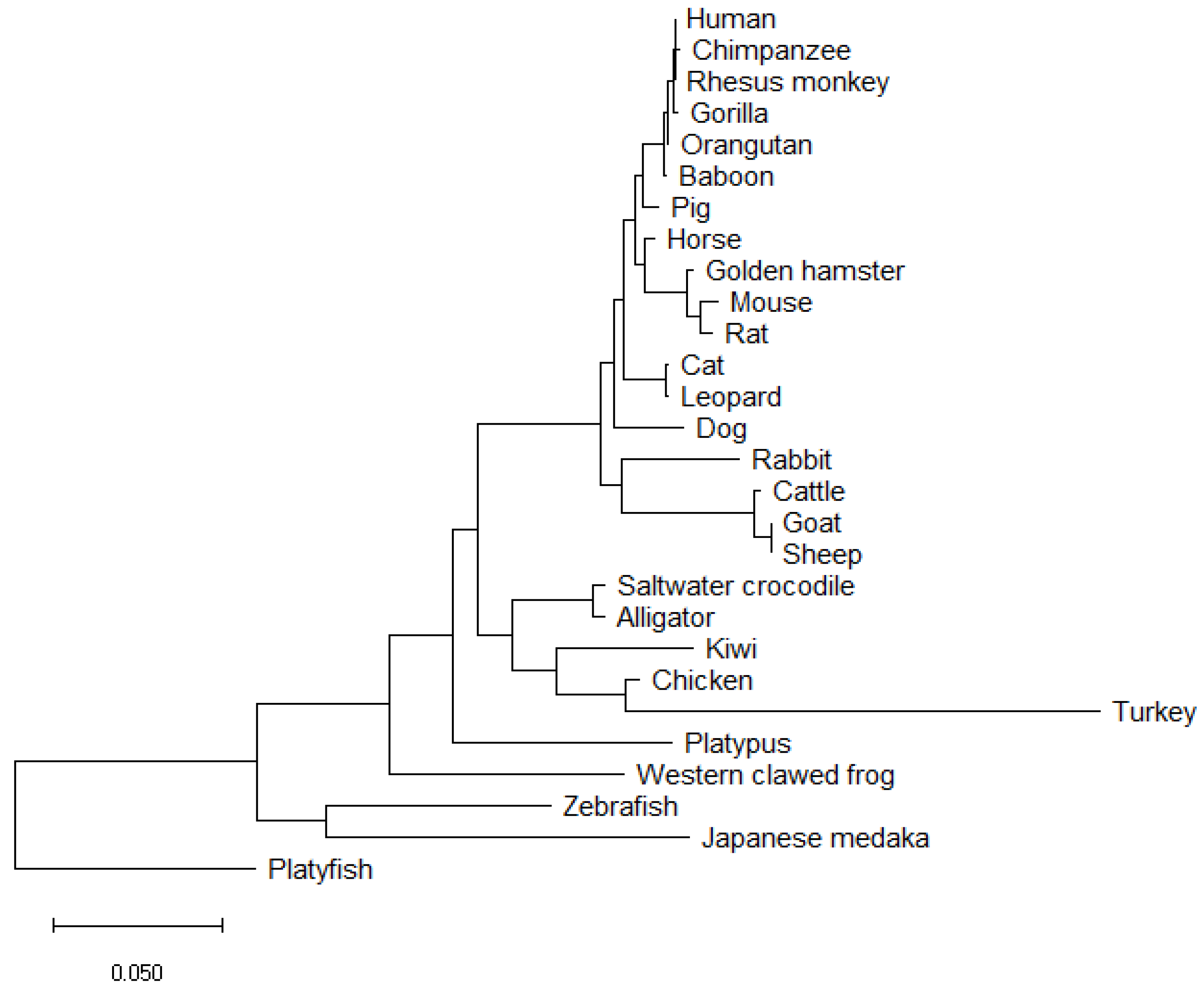{kind=link}
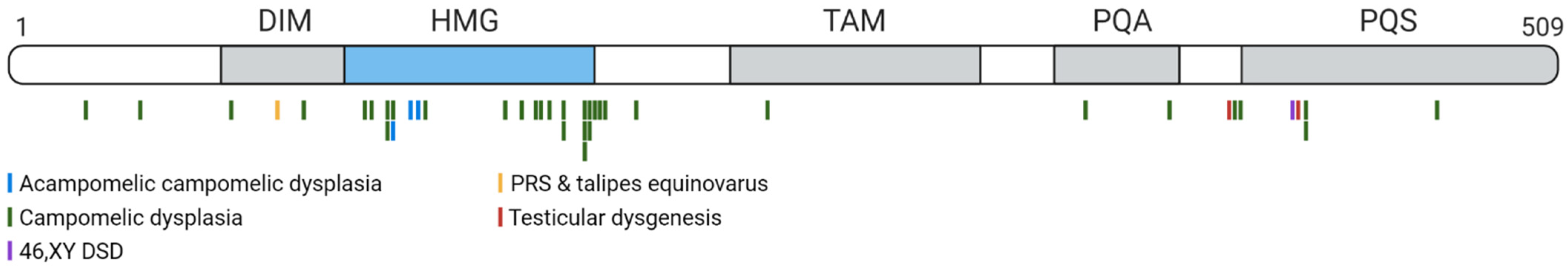{kind=link}
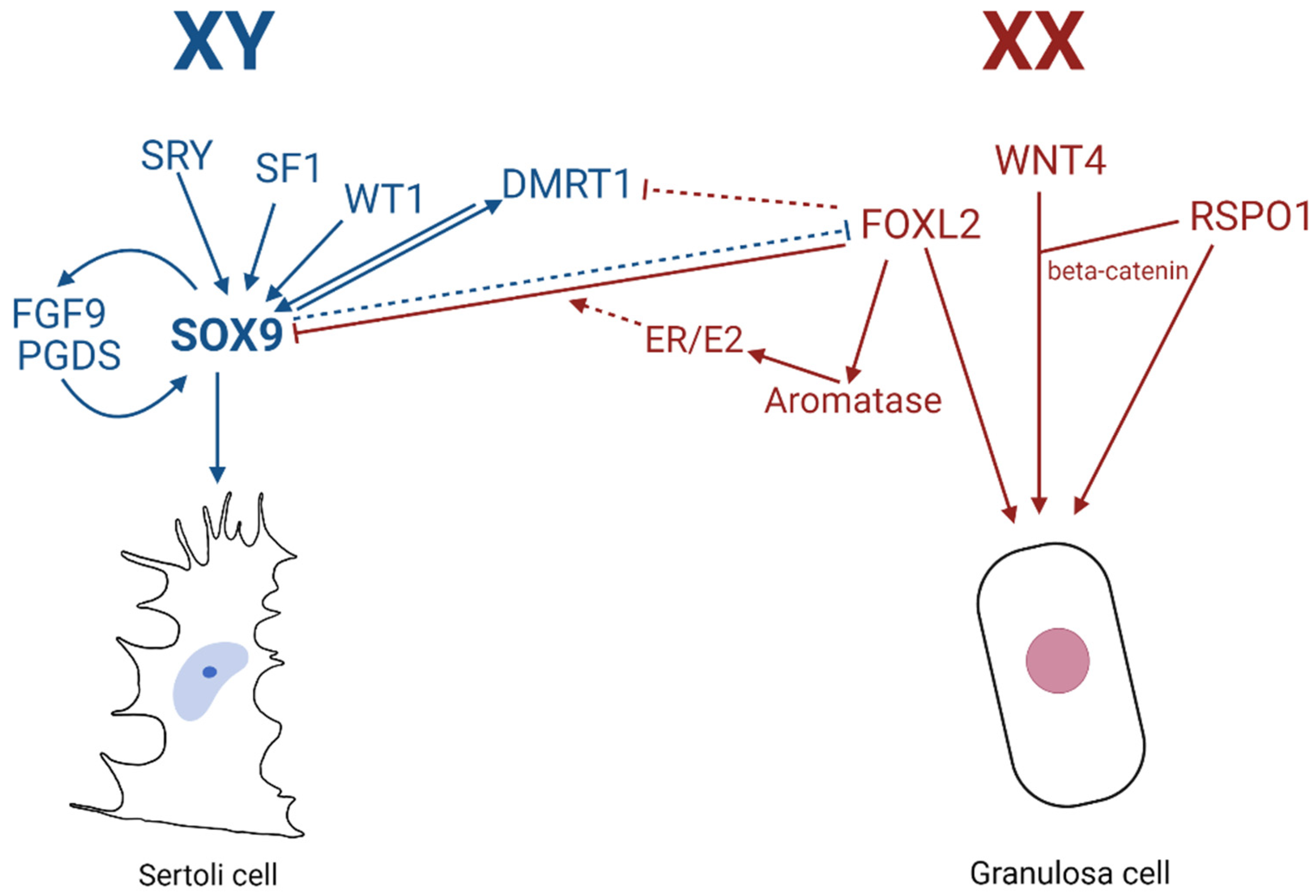{kind=link}
Table 1.
NCBI sequences used to produce N-J phylogenetic tree and COBALT multiple sequence alignment.
Table 1.
NCBI sequences used to produce N-J phylogenetic tree and COBALT multiple sequence alignment.
| NCBI Sequence ID | Amino Acid Length | Species Name | Species Common Name | Identity (%) | Coverage (%) | Mismatches |
|---|---|---|---|---|---|---|
| NP_000337.1 | 509 | Homo sapiens | Human | - | - | |
| NP_001028040.1 | 509 | Macaca mulatta | Rhesus monkey | 100 | 100 | 0 |
| XP_009250264.1 | 509 | Pan troglodytes | Chimpanzee | 99.80 | 100 | 1 |
| XP_018883823.1 | 509 | Pongo abelii | Orangutan | 99.80 | 100 | 1 |
| NP_001009029.1 | 509 | Gorilla gorilla gorilla | Gorilla | 99.80 | 100 | 1 |
| XP_003913405.1 | 509 | Papio Anubis | Baboon | 99.61 | 100 | 2 |
| NP_999008.2 | 511 | Sus scrofa | Pig | 98.04 | 100 | 8 |
| XP_023507898.1 | 509 | Equus caballus | Horse | 98.04 | 100 | 10 |
| XP_005070025.1 | 507 | Mesocricetus auratus | Golden hamster | 96.86 | 99.61 | 14 |
| XP_023099583.1 | 511 | Felis catus | Cat | 96.68 | 99.80 | 13 |
| NP_035578.3 | 507 | Mus musculus | Mouse | 96.46 | 99.61 | 16 |
| XP_032769123.1 | 507 | Rattus rattus | Rat | 96.27 | 99.61 | 17 |
| NP_001002978.1 | 513 | Canis lupus familiaris | Dog | 94.75 | 99.80 | 21 |
| XP_019321937.1 | 529 | Panthera pardus | Leopard | 93.40 | 99.80 | 13 |
| XP_008269985.1 | 497 | Oryctolagus cuniculus | Rabbit | 91.75 | 97.64 | 30 |
| XP_024836864.1 | 524 | Bos taurus | Cattle | 90.84 | 100 | 33 |
| XP_017919394.1 | 525 | Capra hircus | Goat | 90.48 | 100 | 34 |
| XP_027829812.1 | 526 | Ovis aries | Sheep | 90.30 | 100 | 34 |
| XP_020858282.1 | 513 | Phascolarctos cinereus | Koala | 89.60 | 98.82 | 38 |
| XP_006029531.1 | 494 | Alligator sinesis | Chinese alligator | 88.13 | 96.07 | 36 |
| XP_019397875.1 | 494 | Crocodylus porosus | Crocodile | 88.52 | 96.07 | 34 |
| XP_001506094.2 | 508 | Ornithorhynchus anatinus | Platypus | 87.55 | 98.82 | 53 |
| NP_989612.1 | 494 | Gallus gallus | Chicken | 83.52 | 94.50 | 45 |
| NP_001016853.1 | 482 | Xenopus tropicalis | Western clawed frog | 81.64 | 94.11 | 61 |
| NP_001098556.1 | 476 | Oryzias latipes | Japanese medaka | 72.74 | 80.94 | 85 |
| XP_025923282.1 | 500 | Apteryx rowi | Kiwi | 82.73 | 94.70 | 46 |
| XP_010719808.1 | 451 | Meleagris gallopavo | Turkey | 71.64 | 80.94 | 66 |
| XP_005807407.1 | 495 | Xiphosphorus maculatus | Platyfish | 70.58 | 91.75 | 88 |
| NP_571718.1 | 462 | Danio rerio | Zebrafish | 69.96 | 85.46 | 60 |
N.B. Percent identity calculated as the number of mismatches in alignment row relative to human SOX9, where the alignment length is the aligned sequence minus gaps. Percent coverage is calculated as the number of aligned residues in alignment row relative to the length of human SOX9. Mismatches indicate the raw number of residues different per row in comparison to human SOX9. Alignment created via NCBI Orthologues (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gene/6662/ (accessed on 16 February 2021)).and COBALT Multiple Alignment Tool [90].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vining, B.; Ming, Z.; Bagheri-Fam, S.; Harley, V. Diverse Regulation but Conserved Function: SOX9 in Vertebrate Sex Determination. Genes 2021, 12, 486. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040486
AMA Style
Vining B, Ming Z, Bagheri-Fam S, Harley V. Diverse Regulation but Conserved Function: SOX9 in Vertebrate Sex Determination. Genes. 2021; 12(4):486. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040486
Chicago/Turabian StyleVining, Brittany, Zhenhua Ming, Stefan Bagheri-Fam, and Vincent Harley. 2021. "Diverse Regulation but Conserved Function: SOX9 in Vertebrate Sex Determination" Genes 12, no. 4: 486. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12040486
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.