
Analysis of Genetic Diversity and Population Structure of Sesame Accessions from Africa and Asia as Major Centers of Its Cultivation

 , ,
, , 

Abstract
:1. Introduction
2. Experimental Section
2.1. Plant Materials
2.2. DNA Extraction
2.3. PCR and Electrophoresis
2.4. Scoring and Data Analysis
3. Results
3.1. Assessment of the Intra-Accession Variability
3.2. SSR Polymorphism in the Sesame Accessions
3.3. Allele Variation among Geographical Regions
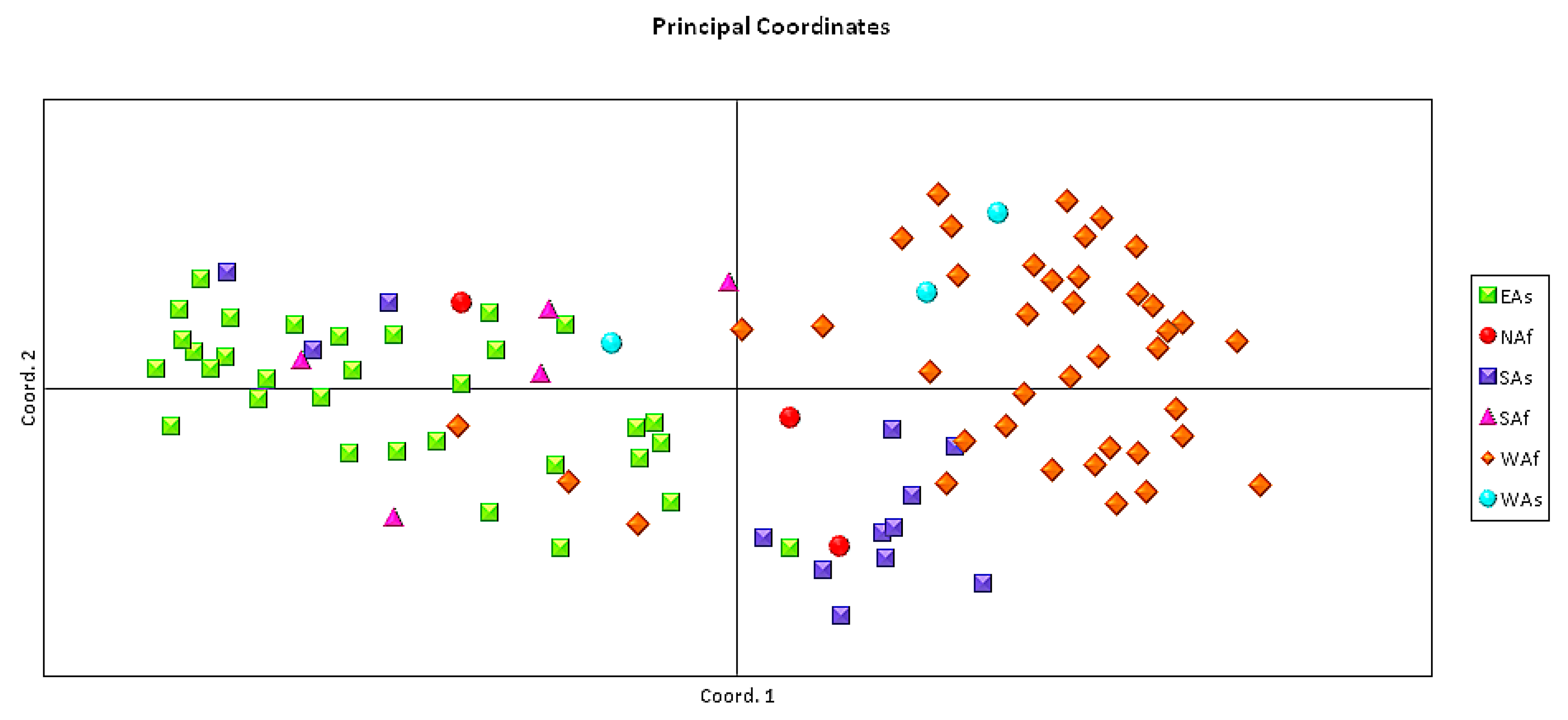
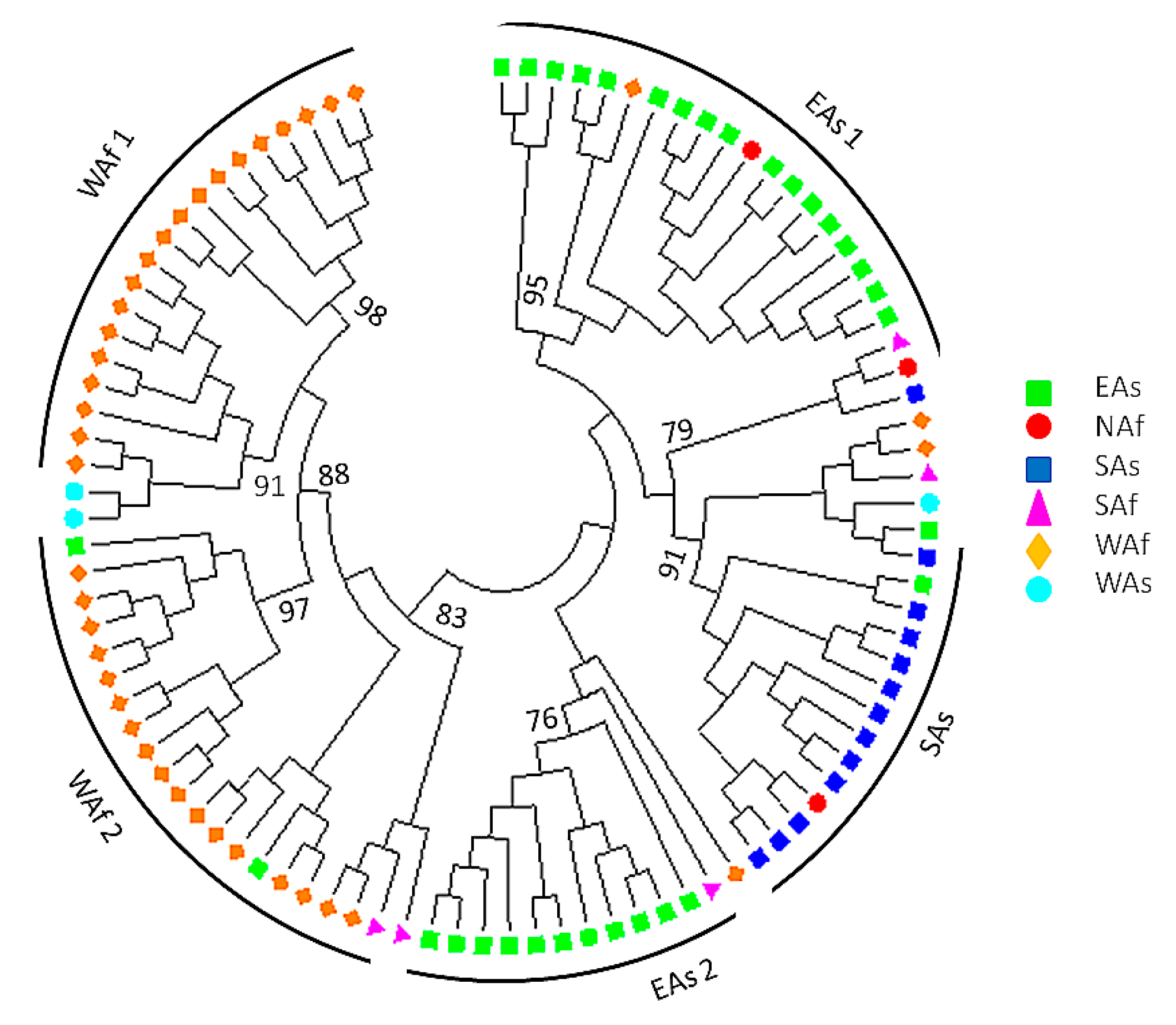
3.4. Pattern of Genetic Diversity and Phylogenetic Relationships
3.5. Analysis of Molecular Variance
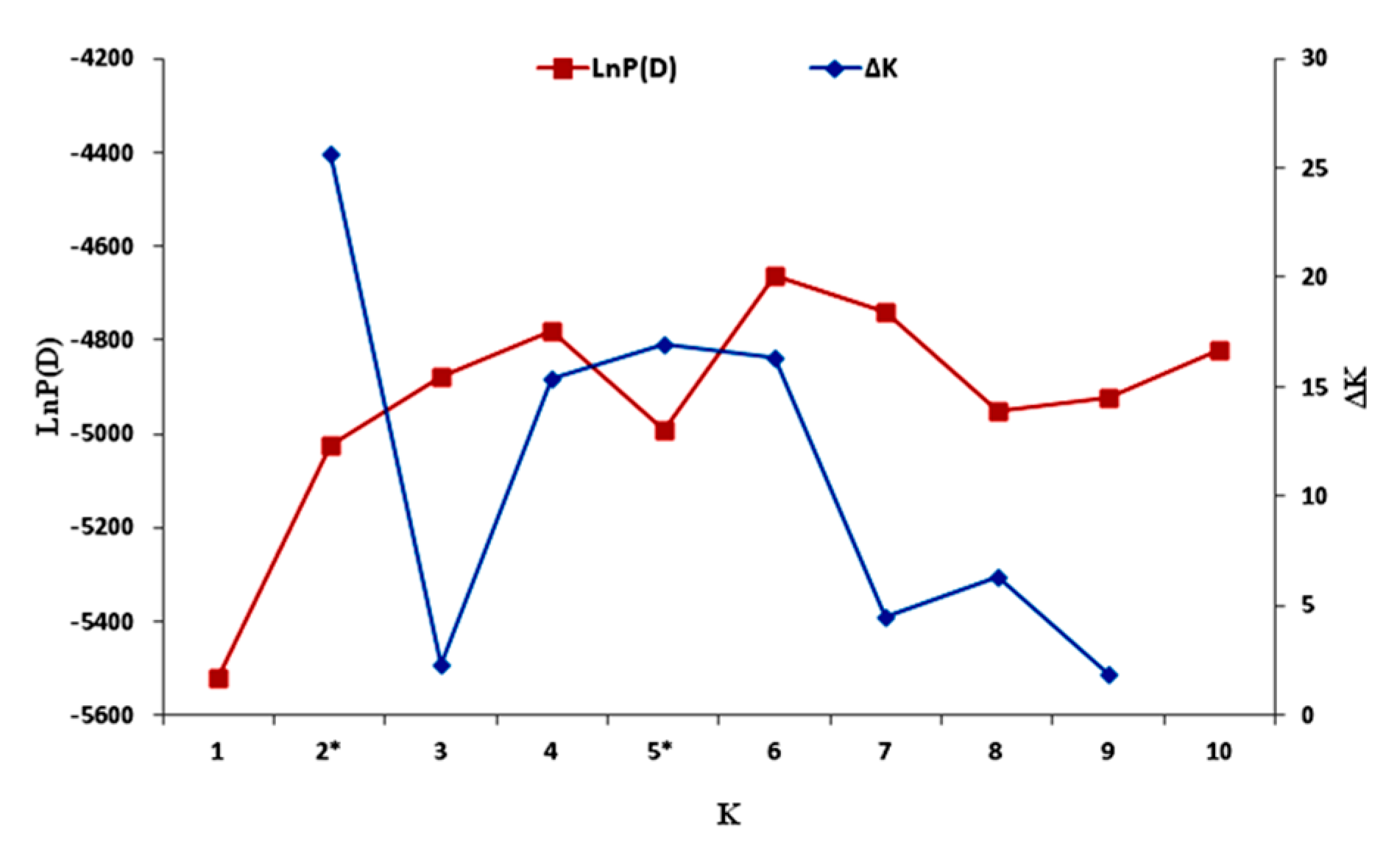
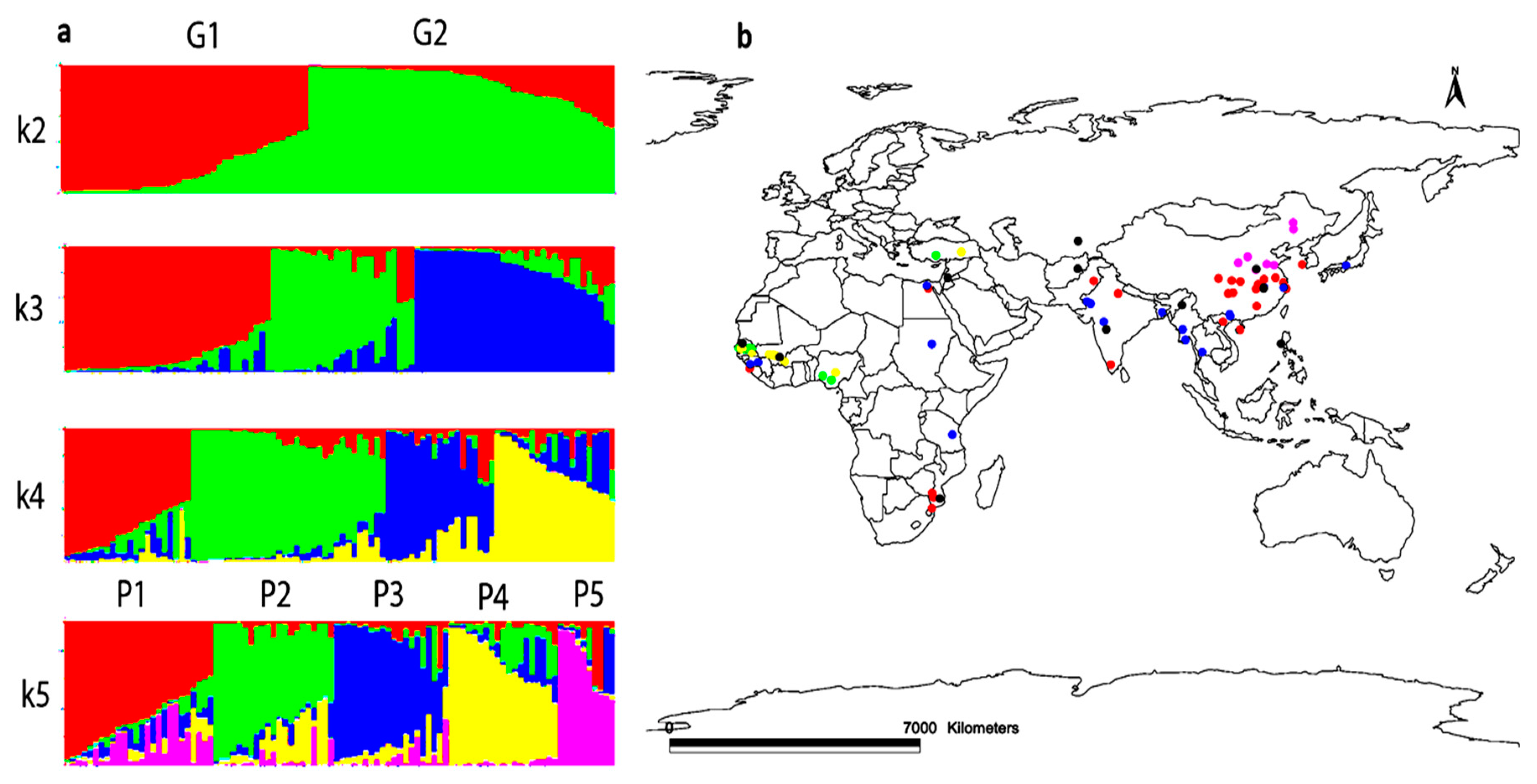
3.6. Population Structure
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Weiss, E.A. Sesame. In Oilseed Crops; Longman: London, UK, 1983. [Google Scholar]
- Mabberley, D.J. The Plant-Book: A Portable Dictionary of the Higher Plants, 2nd ed.; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Nayar, N.M.; Mehra, K.L. Sesame: Its uses, botany, cytogenetics and origin. Econ. Bot. 1970, 24, 20–31. [Google Scholar] [CrossRef]
- Kobayashi, T.; Kinoshita, M.; Hattori, S.; Ogawa, T.; Tsuboi, Y.; Ishida, M.; Ogawa, S.; Saito, H. Development of the sesame metallic fuel performance code. Nucl. Technol. 1990, 89, 183–193. [Google Scholar]
- Ashri, A. Sesame (Sesamum indicum L.). Oilseed Crops. In Genetics Resources, Chromosome Engineering, and Crop Improvement; Singh, R.J., Ed.; CRC Press: Boca Raton, FL, USA, 2007; Volume 4, pp. 231–289. [Google Scholar]
- Wei, X.; Liu, K.; Zhang, Y.; Feng, Q.; Wang, L.; Zhao, Y.; Li, D.; Zhao, Q.; Zhu, X.; Zhu, X.; et al. Genetic discovery for oil production and quality in sesame. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Bedigian, D. Characterization of Sesame (Sesamum indicum L.) Germplasm: A Critique. Genet. Resour. Crop Evol. 2010, 57, 641–647. [Google Scholar] [CrossRef]
- Bedigian, D. Cultivated sesame and wild relatives in the genus Sesamum L. In Sesame: The Genus Sesamum. Medicinal and Aromatic Plants-Industrial Profiles; Bedigian, D., Ed.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2011; pp. 33–77. [Google Scholar]
- Laurentin, H.E.; Karlovsky, P. Genetic relationship and diversity in a sesame (Sesamum indicum L.) germplasm collection using amplified fragment length polymorphism (AFLP). BMC Genet. 2006, 7. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Food and Agriculture Organization statistical databases 2013. Available online: http://faostat.fao.org/ (accessed on 29 August 2015).
- Seegler, C.J.P. Oil Plants in Ethiopia, Their Taxonomy and Agricultural Significance; PUDOC: Wageningen, The Netherlands, 1983. [Google Scholar]
- Burkill, H.M. The Useful Plants of West Tropical Africa, 2nd ed.; RBG Kew: Richmond, UK, 1997; Volume 4, pp. 414–422. [Google Scholar]
- Bedigian, D. Evolution of sesame revisited: Domestication, diversity and prospects. Genet. Resour. Crop Evol. 2003, 50, 779–787. [Google Scholar]
- Pathak, N.; Rai, A.K.; Kumari, R.; Thapa, A.; Bhat, K.V. Sesame Crop: An Underexploited Oilseed Holds Tremendous Potential for Enhanced Food Value. Agric. Sci. 2014, 5, 519–529. [Google Scholar] [CrossRef]
- Hiltebrandt, V.M. Sesame (Sesamum indicum L.). Bull. Appl. Bot. Plant Breed. 1932, 2, 1–114. [Google Scholar]
- De Candolle, A. Origin of Cultivated Plants, 2nd ed.; Noble Offset Printers: New York, NY, USA, 1886. [Google Scholar]
- Vavilov, N.I.; Dorofeev, V.F. Origin and Geography of Cultivated Plants; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Nayar, N.M. Sesame, Sesamum indicum L. (Pedaliaceae). In Evolution of Crop Plants, 2nd ed.; Smartt, J., Simmonds, N.W., Eds.; Wiley: New York, NY, USA, 1995; pp. 404–407. [Google Scholar]
- Ashri, A. Sesame breeding. Plant Breed. Rev. 1998, 16, 179–228. [Google Scholar]
- Weiss, E.A. Oilseed Crops, 2nd ed.; Blackwell Science: Malden, MA, USA, 2000. [Google Scholar]
- Bedigian, D. Sesame in Africa: Origin and dispersals. In Food, Fuel and Fields—Progress in African Archaeobotany, Africa Prae-Historica; Neumann, K., Butler, A., Kahlheber, S., Eds.; Heinrich-Barth-Institute: Cologne, Germany, 2003; pp. 17–36. [Google Scholar]
- Esquinas-Alcazar, J. Protecting crop genetic diversity for food security: Political, ethical and technical challenges. Nat. Rev. Genet. 2005, 6, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing climate change adaptation needs for food security in 2030. Science 2008, 319, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J.; Septiningsih, E.M.; Suwardjo, F.; Santoso, T.J.; Silitonga, T.S.; McCouch, S.R. Genetic diversity analysis of traditional and improved Indonesian rice (Oryza sativa L.) germplasm using microsatellite markers. Theor. Appl. Genet. 2007, 114, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Bhat, K.V.; Babrekar, P.P.; Lakhanpaul, S. Study of genetic diversity in Indian and exotic sesame (Sesamum indicum L.) germplasm using random amplified polymorphic DNA (RAPD) markers. Euphytica 1999, 110, 21–33. [Google Scholar] [CrossRef]
- Ercan, A.G.; Taskin, M.; Turgut, K. Analysis of genetic diversity in Turkish sesame (Sesamum indicum L.) populations using RAPD markers. Genet. Resour. Crop Evol. 2004, 51, 599–607. [Google Scholar] [CrossRef]
- Pham, T.D.; Bui, T.M.; Werlemark, G.; Bui, T.C.; Merker, A.; Carlsson, A.S. A study of genetic diversity of sesame (Sesamum indicum L.) in Vietnam and Cambodia estimated by RAPD markers. Genet. Resour. Crop Evol. 2009, 56, 679–690. [Google Scholar] [CrossRef]
- Kim, D.H.; Zur, G.; Danin-Poleg, Y.; Lee, S.W.; Shim, K.B.; Kang, C.W.; Kashi, Y. Genetic relationships of sesame germplasm collection as revealed by inter-simple sequence repeats. Plant Breed. 2002, 121, 259–262. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Zhang, X.R.; Hua, W.; Wang, L.H.; Che, Z. Analysis of genetic diversity among indigenous landraces from sesame (Sesamum indicum L.) core collection in China as revealed by SRAP and SSR markers. Genes Genom. 2010, 32, 207–215. [Google Scholar] [CrossRef]
- Wei, L.; Zhang, H.; Zheng, Y.; Guo, W.; Zhang, T. Development and utilization of EST-derived microsatellites in sesame (Sesamum indicum L.). Acta Agron. Sin. 2008, 34, 2077–2084. [Google Scholar] [CrossRef]
- Dixit, A.; Jin, M.H.; Chung, J.W.; Yu, J.W.; Chung, H.K.; Ma, K.H.; Park, Y.J.; Cho, E.G. Development of polymorphic microsatellite markers in sesame (Sesamum indicum L.). Mol. Ecol. Notes 2005, 5, 736–738. [Google Scholar] [CrossRef]
- Cho, Y.I.; Park, J.H.; Lee, C.W.; Ra, W.H.; Chung, J.W.; Lee, J.R.; Ma, K.H.; Lee, S.Y.; Lee, K.S.; Lee, M.C.; et al. Evaluation of the genetic diversity and population structure of sesame (Sesamum indicum L.) using microsatellite markers. Genes Genom. 2011, 33, 187–195. [Google Scholar] [CrossRef]
- Wei, X.; Wang, L.; Zhang, Y.; Qi, X.; Wang, X.; Ding, X.; Zhang, J.; Zhang, X. Development of Simple Sequence Repeat (SSR) Markers of Sesame (Sesamum indicum) from a Genome Survey. Molecules 2014, 19, 5150–5162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Zhang, X.R.; Che, Z.; Wang, L.H.; Wei, W.L.; Li, D.H. Genetic diversity assessment of sesame core collection in China by phenotype and molecular markers and extraction of a mini-core collection. BMC Genet. 2012. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Yang, M.; Liu, H.; Tao, Y.; Mei, J.; Zhao, Y. Genetic analysis and molecular characterization of Chinese sesame (Sesamum indicum L.) cultivars using Insertion-Deletion (InDel) and Simple Sequence Repeat (SSR) markers. BMC Genet. 2014. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yu, S.; Tong, C.; Zhao, Y.; Liu, Y.; Song, C.; Zhang, Y.; Zhang, X.; Wang, Y.; Hua, W.; et al. Genome sequencing of the high oil crop sesame provides insight into oil biosynthesis. Genome Biol. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, F.C.; Boyle, T.J.B. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg. J. Bot. 1997, 129, 156–159. [Google Scholar]
- Liu, K.J.; Muse, S.V. PowerMarker, an integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel: Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure, extensions to linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Q.; Börner, A.; Röder, M.S.; Ganal, M.W. Assessing genetic diversity of wheat (Triticum aestivum L.) germplasm using microsatellite markers. Theor. Appl. Genet. 2002, 105, 699–707. [Google Scholar] [PubMed]
- Khlestkina, E.K.; Huang, X.Q.; Quenum, F.J.-B.; Chebotar, S.; Roder, M.S.; Borner, A. Genetic diversity in cultivated plants-loss or stability? Theor. Appl. Genet. 2004, 108, 1466–1472. [Google Scholar] [PubMed]
- Paterson, A. Genetics and Genomics of Cotton. In Plant Genetics and Genomics: Crops and Models; Springer-Verlag: New York, NY, USA, 2009; Volume 3, p. 509. [Google Scholar]
- Aguilar-Meléndez, A.; Morrell, P.L.; Roose, M.L.; Kim, S.C. Genetic diversity and structure in semi wild and domesticated chiles (Capsicum annuum; Solanaceae) from Mexico. Am. J. Bot. 2009, 96, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Qiao, W.H.; Chen, Y.T.; Wang, R.S.; Cao, L.R.; Zhang, W.X.; Yuan, N.N.; Li, Z.C.; Zeng, H.L.; Yang, Q.W. Domestication and geographic origin of Oryza sativa in China: Insights from multilocus analysis of nucleotide variation of O. sativa and O. rufipogon. Mol. Ecol. 2012, 21, 5073–5087. [Google Scholar] [CrossRef] [PubMed]
- Nyongesa, B.; Were, B.; Gudu, S.; Dangasuk, O.; Onkware, A. Genetic diversity in cultivated sesame (Sesamum indicum L.) and related wild species in East Africa. J. Crop. Sci. Biot. 2013, 16, 9–15. [Google Scholar] [CrossRef]
- Hika, G.; Geleta, N.; Jaleta, Z. Genetic Variability, Heritability and Genetic Advance for the Phenotypic Traits in Sesame (Sesamum indicum L.) Populations from Ethiopia. Sci. Technol. Arts Res. J. 2015, 4, 20–26. [Google Scholar] [CrossRef]
- Ali, G.M.; Yasumoto, S.; Seki-Katsuka, M. Assessment of genetic diversity in sesame (Sesamum indicum L.) detected by amplified fragment length polymorphism markers. Electron. J. Biotechnol. 2007, 10, 12–23. [Google Scholar] [CrossRef]
- Yan, W.; Agrama, H.; Jia, M.; Fjellstrom, R.; McClung, A. Geographic Description of Genetic Diversity and Relationships in the USDA Rice World Collection. Crop Sci. 2010, 50, 2406–2417. [Google Scholar] [CrossRef]
- Billot, C.; Ramu, P.; Bouchet, S.; Chantereau, J.; Deu, M.; Gardes, L.; Noyer, J.-L.; Rami, J.-F.; Rivallan, R.; Li, Y.; et al. Massive Sorghum Collection Genotyped with SSR Markers to Enhance Use of Global Genetic Resources. PLoS ONE 2013, 8, e59714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in SOUTHWEST Asia, Europe, and the Mediterranean Basin; Oxford University Press on Demand: Oxford, UK, 2012. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Continents | Geographical Regions | Number of Countries | Number of Accessions |
|---|---|---|---|
| Asia | West Asia (WAs) | 2 | 3 |
| East Asia (EAs) | 5 | 32 | |
| South Asia (SAs) | 7 | 13 | |
| Total | 14 | 48 | |
| Africa | West Africa (WAf) | 5 | 40 |
| North Africa (NAf) | 2 | 3 | |
| Southeast Africa (SAf) | 2 | 5 | |
| Total | 9 | 48 |
| Groups | N | MAF | Na | Ne | He | Ho | I | PIC | Np |
|---|---|---|---|---|---|---|---|---|---|
| Regions | |||||||||
| East Asia | 32 | 0.558 ± 0.138 | 4.030 ± 2.038 | 1.439 ± 0.355 | 0.565 ± 0.130 | 0 | 0.396 ± 0.250 | 0.501 ± 0.146 | 8 |
| West Asia | 3 | 0.585 ± 0.186 | 2.242 ± 0.560 | 1.265 ± 0.382 | 0.484 ± 0.161 | 0 | 0.209 ± 0.301 | 0.399 ± 0.153 | 0 |
| South Asia | 13 | 0.505 ± 0.176 | 3.848 ± 1.543 | 1.458 ± 0.349 | 0.598 ± 0.182 | 0 | 0.414 ± 0.240 | 0.542 ± 0.185 | 5 |
| North Africa | 3 | 0.707 ± 0.246 | 1.878 ± 0.739 | 1.385 ± 0.408 | 0.343 ± 0.261 | 0.006 | 0.303 ± 0.320 | 0.282 ± 0.224 | 5 |
| Southeastern Africa | 5 | 0.581 ± 0.214 | 2.787 ± 1.139 | 1.357 ± 0.377 | 0.509 ± 0.210 | 0 | 0.305 ± 0.297 | 0.445 ± 0.209 | 4 |
| West Africa | 40 | 0.578 ± 0.178 | 4.909 ± 2.350 | 1.379 ± 0.362 | 0.550 ± 0.179 | 0.002 | 0.346 ± 0.263 | 0.502 ± 0.178 | 17 |
| Continents | |||||||||
| Africa | 48 | 0.565 ± 0.184 | 5.242 ± 2.739 | 1.407 ± 0.348 | 0.559 ± 0.181 | 0.003 | 0.382 ± 0.235 | 0.510 ± 0.181 | 30 |
| Asia | 48 | 0.513 ± 0.145 | 4.848 ± 2.575 | 1.4712 ± 0.348 | 0.604 ± 0.134 | 0 | 0.430 ± 0.221 | 0.543 ± 0.151 | 21 |
| Source | df | SS | MS | Est. Var. | %Tv | p |
|---|---|---|---|---|---|---|
| Among geographical regions | 2 | 1103.034 | 551.517 | 20.135 | 44.66 | 0.001 |
| Within geographical regions | 82 | 2246.107 | 27.391 | 24.95 | 55.34 | |
| Total | 84 | 3349.141 | 45.085 | 100 |
| Source | df | SS | MS | Est. Var. | %Tv | p |
|---|---|---|---|---|---|---|
| Among continents | 1 | 615.726 | 615.726 | 18.189 | 34.952 | 0.001 |
| Within continents | 94 | 3182.003 | 33.851 | 33.851 | 65.048 | |
| Total | 95 | 3797.729 | 41.999 | 100 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dossa, K.; Wei, X.; Zhang, Y.; Fonceka, D.; Yang, W.; Diouf, D.; Liao, B.; Cissé, N.; Zhang, X. Analysis of Genetic Diversity and Population Structure of Sesame Accessions from Africa and Asia as Major Centers of Its Cultivation. Genes 2016, 7, 14. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7040014
Dossa K, Wei X, Zhang Y, Fonceka D, Yang W, Diouf D, Liao B, Cissé N, Zhang X. Analysis of Genetic Diversity and Population Structure of Sesame Accessions from Africa and Asia as Major Centers of Its Cultivation. Genes. 2016; 7(4):14. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7040014
Chicago/Turabian StyleDossa, Komivi, Xin Wei, Yanxin Zhang, Daniel Fonceka, Wenjuan Yang, Diaga Diouf, Boshou Liao, Ndiaga Cissé, and Xiurong Zhang. 2016. "Analysis of Genetic Diversity and Population Structure of Sesame Accessions from Africa and Asia as Major Centers of Its Cultivation" Genes 7, no. 4: 14. https://0-doi-org.brum.beds.ac.uk/10.3390/genes7040014