
Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies


Abstract
:1. Introduction
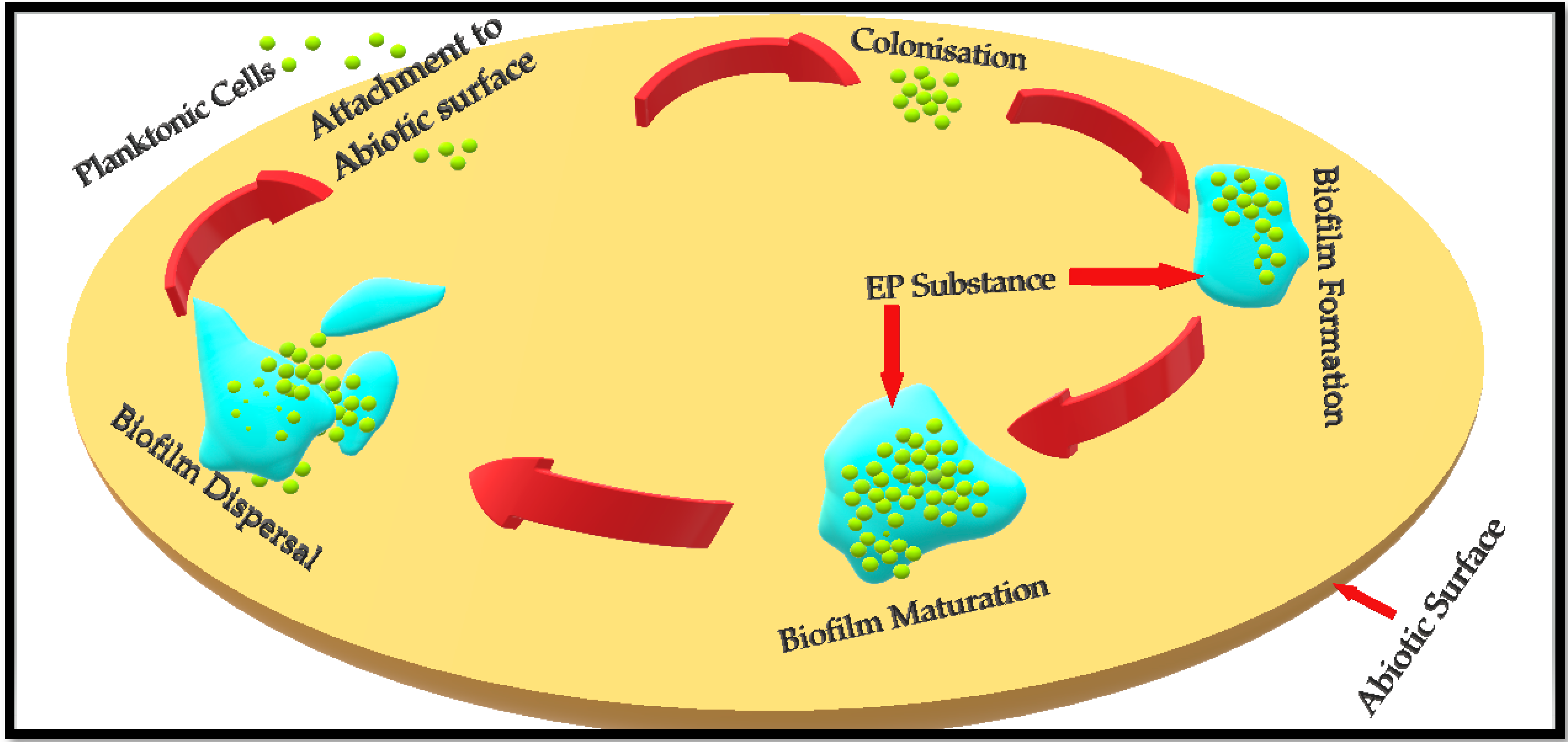
2. Biofilm Formation
- Attachment of planktonic cells to the surface (either a biotic host or any abiotic surface);
- Colonisation and biofilm formation;
- Biofilm maturation;
- Biofilm dispersal.
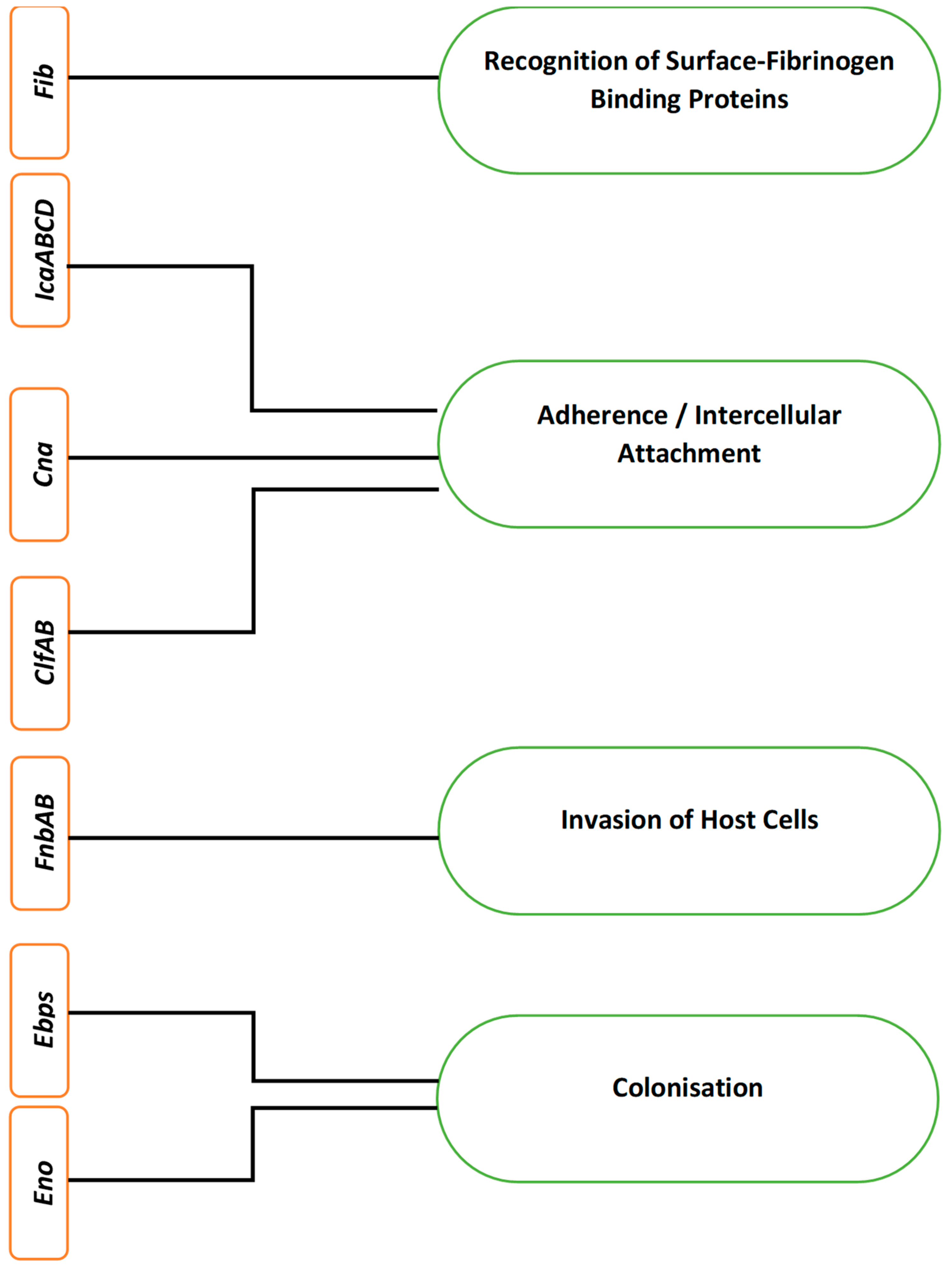
3. Gene Expression during Staph. aureus Biofilm Formation and Dispersal
3.1. Gene Expression and Quorum Sensing in Staphylococcus aureus
3.2. Gene Expression and Staphylococcus aureus Pathogenesis
4. Staphylococcus aureus Biofilm and Antimicrobial Resistance (AMR)
5. Clinical Implications of Staphylococcus aureus Biofilms
6. Techniques and Strategies Used in Studying Staphylococcus aureus Biofilm
6.1. Direct Observation Techniques
6.1.1. Light Microscope and Transmission Electron Microscope
6.1.2. Profilometry and Scanning Electron Microscope
6.1.3. Scanning Transmission X-ray Microscopy (STXM)
6.1.4. Fluorescent Tagging of Biofilm
- (a)
- Confocal laser scanning microscopy (CLSM)
- (b)
- Fluorescent in situ hybridization (FISH)
6.2. Indirect Observational Techniques
6.2.1. Tube Method
6.2.2. Congo Red Agar Method
6.2.3. Detection of Biofilm Production by Microtiter Plate Assay
6.2.4. Biofilm-Associated Genes Detection by PCR
6.2.5. Mass Spectrometry
6.2.6. Atomic Force Microscopy (AFM)
7. Strategies Used to Inhibit and Disrupt Staphylococcus aureus Biofilm
7.1. Antibiofilm Drugs and Functional Excipients
7.2. Other Antibiofilm Molecules
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Adams, M. Staphylococcus aureus and other pathogenic Gram-positive cocci. In Foodborne Pathogens; de Blackburn, C.W., McClure, P.J., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Elsevier: Cambridge, UK, 2009; pp. 802–819. ISBN 9781845693626. [Google Scholar]
- Schaumburg, F.; Pauly, M.; Anoh, E.; Mossoun, A.; Wiersma, L.; Schubert, G.; Flammen, A.; Alabi, A.S.; Muyembe-Tamfum, J.-J.; Grobusch, M.P.; et al. Staphylococcus aureus complex from animals and humans in three remote African regions. Clin. Microbiol. Infect. 2015, 21, 345.e1–345.e8. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.D.; Malachowa, N.; Deleo, F.R. Pathogenesis of Staphylococcus aureus abscesses. Am. J. Pathol. 2015, 185, 1518–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, J.B.; Mlynek, K.D.; Hettiarachchi, H.; Alamneh, Y.A.; Biggemann, L.; Zurawski, D.V.; Black, C.C.; Bane, C.E.; Kim, R.K.; Granick, M.S. Extracellular polymeric substance (EPS)-degrading enzymes reduce staphylococcal surface attachment and biocide resistance on pig skin in vivo. PLoS ONE 2018, 13, e0205526. [Google Scholar] [CrossRef] [PubMed]
- Nazir, R.; Zaffar, M.R.; Amin, I. Bacterial biofilms: The remarkable heterogeneous biological communities and nitrogen fixing microorganisms in lakes. In Freshwater Microbiology: Perspectives of Bacterial Dynamics in Lake Ecosystems; Elsevier: Amsterdam, The Netherlands, 2019; pp. 307–340. ISBN 9780128174951. [Google Scholar]
- Donlan, R.M. Biofilms: Microbial Life on Surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Idrees, M.; Mohammad, A.R.; Karodia, N.; Rahman, A. Multimodal Role of Amino Acids in Microbial Control and Drug Development. Antibiotics 2020, 9, 330. [Google Scholar] [CrossRef]
- Reffuveille, F.; Josse, J.; Vallé, Q.; Gangloff, C.M.; Gangloff, S.C. Staphylococcus aureus Biofilms and their Impact on the Medical Field. In The Rise of Virulence and Antibiotic Resistance in Staphylococcus aureus; InTech: Rijeka, Croatia, 2017; Chapter 11; p. 187. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.T.T.; Nguyen, T.H.; Otto, M. The staphylococcal exopolysaccharide PIA–Biosynthesis and role in biofilm formation, colonization, and infection. Comput. Struct. Biotechnol. J. 2020, 18, 3324–3334. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, R.M.; Rigby, D.; Handley, P.; Foster, T.J. The role of Staphylococcus aureus surface protein SasG in adherence and biofilm formation. Microbiology 2007, 153, 2435–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, A.; Bhattacharyya, S.; Kundu, A.; Dutta, D.; Das, A.K. Macroscopic amyloid fiber formation by staphylococcal biofilm associated SuhB protein. Biophys. Chem. 2016, 217, 32–41. [Google Scholar] [CrossRef]
- Lacey, K.A.; Leech, J.M.; Lalor, S.J.; McCormack, N.; Geoghegan, J.A.; McLoughlin, R.M. The Staphylococcus aureus cell wall-anchored protein clumping factor A is an important T cell antigen. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Taglialegna, A.; Lasa, I.; Valle, J. Amyloid structures as biofilm matrix scaffolds. J. Bacteriol. 2016, 198, 2579–2588. [Google Scholar] [CrossRef] [Green Version]
- Miao, J.; Lin, S.; Soteyome, T.; Peters, B.M.; Li, Y.; Chen, H.; Su, J.; Li, L.; Li, B.; Xu, Z.; et al. Biofilm Formation of Staphylococcus aureus under Food Heat Processing Conditions: First Report on CML Production within Biofilm. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Neu, T.R.; Lawrence, J.R. Extracellular Polymeric Substances in Microbial Biofilms. In Microbial Glycobiology; Elsevier Inc.: Amsterdam, The Netherlands, 2010; pp. 733–758. ISBN 9780123745460. [Google Scholar]
- Algburi, A.; Comito, N.; Kashtanov, D.; Dicks, L.M.T.; Chikindas, M.L. Control of biofilm formation: Antibiotics and beyond. Appl. Environ. Microbiol. 2017, 83, e02508-16. [Google Scholar] [CrossRef] [Green Version]
- Guangyin, Z.; Youcai, Z. Harvest of Bioenergy From Sewage Sludge by Anaerobic Digestion. In Pollution Control and Resource Recovery; Elsevier: Amsterdam, The Netherlands, 2017; pp. 181–273. [Google Scholar]
- Landini, P.; Antoniani, D.; Burgess, J.G.; Nijland, R. Molecular mechanisms of compounds affecting bacterial biofilm formation and dispersal. Appl. Microbiol. Biotechnol. 2010, 86, 813–823. [Google Scholar] [CrossRef]
- Petrova, O.E.; Sauer, K. Sticky situations: Key components that control bacterial surface attachment. J. Bacteriol. 2012, 194, 2413–2425. [Google Scholar] [CrossRef] [Green Version]
- Hamadi, F.; Latrache, H.; Mabrrouki, M.; Elghmari, A.; Outzourhit, A.; Ellouali, M.; Chtaini, A. Effect of pH on distribution and adhesion of Staphylococcus aureus to glass. J. Adhes. Sci. Technol. 2005, 19, 73–85. [Google Scholar] [CrossRef]
- Krasowska, A.; Sigler, K. How microorganisms use hydrophobicity and what does this mean for human needs? Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maikranz, E.; Spengler, C.; Thewes, N.; Thewes, A.; Nolle, F.; Jung, P.; Bischoff, M.; Santen, L.; Jacobs, K. Different binding mechanisms of: Staphylococcus aureus to hydrophobic and hydrophilic surfaces. Nanoscale 2020, 12, 19267–19275. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal Biofilms. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef] [Green Version]
- Paharik, A.E.; Horswill, A.R. The Staphylococcal Biofilm: Adhesins, Regulation, and Host Response. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Archer, N.K.; Mazaitis, M.J.; William Costerton, J.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, regulation and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef] [Green Version]
- Moormeier, D.E.; Bayles, K.W. Staphylococcus aureus biofilm: A complex developmental organism. Mol. Microbiol. 2017, 104, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, K. The genomics and proteomics of biofilm formation. Genome Biol. 2003, 4, 219. [Google Scholar] [CrossRef] [Green Version]
- Nourbakhsh, F.; Namvar, A.E. Detection of genes involved in biofilm formation in Staphylococcus aureus isolates. GMS Hyg. Infect. Control 2016, 11, Doc07. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xie, S.; Lou, X.; Cheng, S.; Liu, X.; Zheng, W.; Zheng, Z.; Wang, H. Biofilm formation and prevalence of adhesion genes among Staphylococcus aureus isolates from different food sources. Microbiologyopen 2020, 9, e00946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azmi, K.; Qrei, W.; Abdeen, Z. Screening of genes encoding adhesion factors and biofilm production in methicillin resistant strains of Staphylococcus aureus isolated from Palestinian patients. BMC Genom. 2019, 20, 571–578. [Google Scholar] [CrossRef]
- O’Neill, E.; Pozzi, C.; Houston, P.; Humphreys, H.; Robinson, D.A.; Loughman, A.; Foster, T.J.; O’Gara, J.P. A Novel Staphylococcus aureus Biofilm Phenotype Mediated by the Fibronectin-Binding Proteins, FnBPA and FnBPB. J. Bacteriol. 2008, 190, 3835. [Google Scholar] [CrossRef] [Green Version]
- Sinha, B.; Francois, P.; Que, Y.-A.; Hussain, M.; Heilmann, C.; Moreillon, P.; Lew, D.; Krause, K.-H.; Peters, G.; Herrmann, M. Heterologously Expressed Staphylococcus aureusFibronectin-Binding Proteins Are Sufficient for Invasion of Host Cells. Infect. Immun. 2000, 68, 6871. [Google Scholar] [CrossRef] [Green Version]
- Wolz, C.; Goerke, C.; Landmann, R.; Zimmerli, W.; Fluckiger, U. Transcription of Clumping Factor A in Attached and Unattached Staphylococcus aureus In Vitro and during Device-Related Infection. Infect. Immun. 2002, 70, 2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman-Bausier, P.; Labate, C.; Towell, A.M.; Derclaye, S.; Geoghegan, J.A.; Dufrêne, Y.F. Staphylococcus aureus clumping factor A is a force-sensitive molecular switch that activates bacterial adhesion. Proc. Natl. Acad. Sci. USA 2018, 115, 5564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabat, A.; Melles, D.C.; Martirosian, G.; Grundmann, H.; Van Belkum, A.; Hryniewicz, W. Distribution of the serine-aspartate repeat protein-encoding sdr genes among nasal-carriage and invasive Staphylococcus aureus strains. J. Clin. Microbiol. 2006, 44, 1135–1138. [Google Scholar] [CrossRef] [Green Version]
- Kot, B.; Sytykiewicz, H.; Sprawka, I. Expression of the Biofilm-Associated Genes in Methicillin-Resistant Staphylococcus aureus in Biofilm and Planktonic Conditions. Int. J. Mol. Sci. 2018, 19, 3487. [Google Scholar] [CrossRef] [Green Version]
- Le, K.Y.; Otto, M. Quorum-sensing regulation in staphylococci-an overview. Front. Microbiol. 2015, 6, 1174. [Google Scholar] [CrossRef] [Green Version]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Saggu, S.K.; Jha, G.; Mishra, P.C. Enzymatic Degradation of Biofilm by Metalloprotease From Microbacterium sp. SKS10. Front. Bioeng. Biotechnol. 2019, 7, 192. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Zhou, R.; Wang, H.; Zeng, Z. Accessory gene regulator in Staphylococcus biofilm formation and infection. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2008, 33, 1066–1070. [Google Scholar]
- Grundstad, M.L.; Parlet, C.P.; Kwiecinski, J.M.; Kavanaugh, J.S.; Crosby, H.A.; Cho, Y.-S.; Heilmann, K.; Diekema, D.J.; Horswill, A.R. Quorum Sensing, Virulence, and Antibiotic Resistance of USA100 Methicillin-Resistant Staphylococcus aureus Isolates. mSphere 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Lepp, D.; Gohari, I.M.; Wu, T.; Zhou, H.; Yin, X.; Yu, H.; Prescott, J.F.; Nie, S.P.; Xie, M.Y.; et al. The Agr-like quorum sensing system is required for pathogenesis of necrotic enteritis caused by Clostridium perfringens in poultry. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Rajasree, K.; Fasim, A.; Gopal, B. Conformational features of the Staphylococcus aureus AgrA-promoter interactions rationalize quorum-sensing triggered gene expression. Biochem. Biophys. Rep. 2016, 6, 124–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oogai, Y.; Matsuo, M.; Hashimoto, M.; Kato, F.; Sugai, M.; Komatsuzawa, H. Expression of virulence factors by Staphylococcus aureus grown in serum. Appl. Environ. Microbiol. 2011, 77, 8097–8105. [Google Scholar] [CrossRef] [Green Version]
- Gordon, R.J.; Lowy, F.D. Pathogenesis of methicillin-resistant Staphylococcus aureus infection. Clin. Infect. Dis. 2008, 46, S350–S359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, M. Staphylococcus aureus toxins. Curr. Opin. Microbiol. 2014, 17, 32–37. [Google Scholar] [CrossRef] [Green Version]
- Vandenesch, F.; Lina, G.; Henry, T. Staphylococcus aureus hemolysins, bi-component leukocidins, and cytolytic peptides: A redundant arsenal of membrane-damaging virulence factors? Front. Cell. Infect. Microbiol. 2012, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Kolar, S.L.; Antonio Ibarra, J.; Rivera, F.E.; Mootz, J.M.; Davenport, J.E.; Stevens, S.M.; Horswill, A.R.; Shaw, L.N. Extracellular proteases are key mediators of Staphylococcus aureus virulence via the global modulation of virulence-determinant stability. Microbiologyopen 2013, 2, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Thammavongsa, V.; Schneewind, O.; Missiakas, D. Recurrent infections and immune evasion strategies of Staphylococcus aureus. Curr. Opin. Microbiol. 2012, 15, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; Van Kessel, K.P.M.; Van Strijp, J.A.G. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on β-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- WHO. New Report Calls for Urgent Action to Avert Antimicrobial Resistance Crisis; WHO: Geneva, Switzerland, 2019; Volume 29, pp. 2019–2021. [Google Scholar]
- Shrestha, P.; Cooper, B.S.; Coast, J.; Oppong, R.; Thuy, N.D.T.; Phodha, T.; Celhay, O.; Guerin, P.J.; Wertheim, H.; Lubell, Y. Enumerating the economic cost of antimicrobial resistance per antibiotic consumed to inform the evaluation of interventions affecting their use. Antimicrob. Resist. Infect. Control 2018, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M. Role of biofilms in antimicrobial resistance. ASAIO J. 2000, 46, S47–S52. [Google Scholar] [CrossRef]
- Rizzato, C.; Torres, J.; Kasamatsu, E.; Camorlinga-Ponce, M.; Bravo, M.M.; Canzian, F.; Kato, I. Potential role of biofilm formation in the development of digestive tract cancer with special reference to helicobacter pylori infection. Front. Microbiol. 2019, 10, 846. [Google Scholar] [CrossRef] [PubMed]
- Fadeeva, E.; Schlie-Wolter, S.; Chichkov, B.N.; Paasche, G.; Lenarz, T. Structuring of biomaterial surfaces with ultrashort pulsed laser radiation. In Laser Surface Modification of Biomaterials: Techniques and Applications; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 145–172. ISBN 9780081009420. [Google Scholar]
- Ou, C.; Shang, D.; Yang, J.; Chen, B.; Chang, J.; Jin, F.; Shi, C. Prevalence of multidrug-resistant Staphylococcus aureus isolates with strong biofilm formation ability among animal-based food in Shanghai. Food Control 2020, 112, 107106. [Google Scholar] [CrossRef]
- Staph Can Lurk Deep within Nose, Study Finds|News Center|Stanford Medicine. Available online: https://med.stanford.edu/news/all-news/2013/12/staph-can-lurk-deep-within-nose-study-finds.html (accessed on 8 July 2021).
- Sakr, A.; Brégeon, F.; Mège, J.L.; Rolain, J.M.; Blin, O. Staphylococcus aureus nasal colonization: An update on mechanisms, epidemiology, risk factors, and subsequent infections. Front. Microbiol. 2018, 9, 2419. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, R.V.; Fowler, V.G., Jr.; Skov, R.; Bruun, N.E. Future challenges and treatment of Staphylococcus aureus bacteremia with emphasis on MRSA. Future Microbiol. 2011, 6, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Amyes, S.G. Treatment of staphylococcal infection. BMJ 2005, 330, 976–977. [Google Scholar] [CrossRef]
- Min, S.H.; Kang, M.H.; Sur, J.H.; Park, H.M. Staphylococcus pseudintermedius infection associated with nodular skin lesions and systemic inflammatory response syndrome in a dog. Can. Vet. J. 2014, 55, 480–483. [Google Scholar] [PubMed]
- Sonesson, A.; Przybyszewska, K.; Eriksson, S.; Mörgelin, M.; Kjellström, S.; Davies, J.; Potempa, J.; Schmidtchen, A. Identification of bacterial biofilm and the Staphylococcus aureus derived protease, staphopain, on the skin surface of patients with atopic dermatitis. Sci. Rep. 2017, 7, 8682–8689. [Google Scholar] [CrossRef]
- Verma, J.; Bag, S.; Saha, B.; Kumar, P.; Ghosh, T.S.; Dayal, M.; Senapati, T.; Mehra, S.; Dey, P.; Desigamani, A.; et al. Genomic plasticity associated with antimicrobial resistance in Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2019, 116, 6226–6231. [Google Scholar] [CrossRef] [Green Version]
- Manzur, A.; Gavalda, L.; de Gopegui, E.R.; Mariscal, D.; Dominguez, M.A.; Perez, J.L.; Segura, F.; Pujol, M. Spanish Network for Research in, I.D. Prevalence of methicillin-resistant Staphylococcus aureus and factors associated with colonization among residents in community long-term-care facilities in Spain. Clin. Microbiol. Infect. 2008, 14, 867–872. [Google Scholar] [CrossRef] [Green Version]
- Marques, S.C.; Rezende, J.D.G.O.S.; Alves, L.A.D.F.; Silva, B.C.; Alves, E.; de Abreu, L.R.; Piccoli, R.H. Formation of biofilms by Staphylococcus aureus on stainless steel and glass surfaces and its resistance to some selected chemical sanitizers. Braz. J. Microbiol. 2007, 38, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Chmielewski, R.A.N.; Frank, J.F. Biofilm formation and control in food processing facilities. Compr. Rev. Food Sci. Food Saf. 2003, 2, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- Characklis, W.G.; Trulear, M.G.; Bryers, J.D.; Zelver, N. Dynamics of biofilm processes: Methods. Water Res. 1982, 16, 1207–1216. [Google Scholar] [CrossRef]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef]
- Kırmusaoğlu, S. The Methods for Detection of Biofilm and Screening Antibiofilm Activity of Agents. In Antimicrobials, Antibiotic Resistance, Antibiofilm Strategies and Activity Methods; IntechOpen: London, UK, 2019. [Google Scholar]
- Lawrence, J.R.; Swerhone, G.D.W.; Leppard, G.G.; Araki, T.; Zhang, X.; West, M.M.; Hitchcock, A.P. Scanning transmission X-ray, laser scanning, and transmission electron microscopy mapping of the exopolymeric matrix of microbial biofilms. Appl. Environ. Microbiol. 2003, 69, 5543–5554. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Biswas, N.; Datta, A.; Dey, R.; Maiti, P. Atomic force microscopy in biofilm study. Microscopy 2014, 63, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Biswas, N.; Datta, A.; Maiti, P.K. Periodicities in the roughness and biofilm growth on glass substrate with etching time: Hydrofluoric acid etchant. PLoS ONE 2019, 14, e0214192. [Google Scholar] [CrossRef]
- Jhass, A.K.; Johnston, D.A.; Gulati, A.; Anand, R.; Stoodley, P.; Sharma, S. A scanning electron microscope characterisation of biofilm on failed craniofacial osteosynthesis miniplates. J. Cranio-Maxillofac. Surg. 2014, 42, e372–e378. [Google Scholar] [CrossRef] [PubMed]
- Warwick, T.; Franck, K.; Kortright, J.B.; Meigs, G.; Moronne, M.; Myneni, S.; Rotenberg, E.; Seal, S.; Steele, W.F.; Ade, H.; et al. A scanning transmission x-ray microscope for materials science spectromicroscopy at the advanced light source. Rev. Sci. Instrum. 1998. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Balhorn, R.; Mazrimas, J.; Kirz, J. Mapping and measuring DNA to protein ratios in mammalian sperm head by XANES imaging. J. Struct. Biol. 1996, 116, 335–344. [Google Scholar] [CrossRef]
- Loo, B.W.; Sauerwald, I.M.; Hitchcock, A.P.; Rothman, S.S. A new sample preparation method for biological soft X-ray microscopy: Nitrogen-based contrast and radiation tolerance properties of glycol methacrylate-embedded and sectioned tissue. J. Microsc. 2001, 204, 69–86. [Google Scholar] [CrossRef] [Green Version]
- Neu, T.R.; Swerhone, G.D.W.; Lawrence, J.R. Assessment of lectin-binding analysis for in situ detection of glycoconjugates in biofilm systems. Microbiology 2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neu, T.R.; Lawrence, J.R. In situ characterization of extracellular polymeric substances (EPS) in biofilm systems. In Microbial Extracellular Polymeric Substances; Springer: Heidelberg/Berlin, Germany, 1999; pp. 21–47. [Google Scholar]
- Van Gijlswijk, R.P.M.; Zijlmans, H.J.M.A.A.; Wiegant, J.; Bobrow, M.N.; Erickson, T.J.; Adler, K.E.; Tanke, H.J.; Raap, A.K. Fluorochrome-labeled tyramides: Use in immunocytochemistry and fluorescence in situ hybridization. J. Histochem. Cytochem. 1997, 45, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, A.; Bishop, P.L.; Okabe, S.; Lee, S.G.; Kim, I.S. Effect of dissolved oxygen concentration on the biofilm and in situ analysis by fluorescence in situ hybridization (FISH) and microelectrodes. Water Sci. Technol. J. Int. Assoc. Water Pollut. Res. 2003, 47, 49–57. [Google Scholar] [CrossRef]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Mathur, T.; Singhal, S.; Khan, S.; Upadhyay, D.J.; Fatma, T.; Rattan, A. Detection of biofilm formation among the clinical isolates of staphylococci: An evaluation of three different screening methods. Indian J. Med. Microbiol. 2006, 24, 25–29. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, 2437. [Google Scholar] [CrossRef]
- Sanchez, C.J.; Mende, K.; Beckius, M.L.; Akers, K.S.; Romano, D.R.; Wenke, J.C.; Murray, C.K. Biofilm formation by clinical isolates and the implications in chronic infections. BMC Infect. Dis. 2013, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoramrooz, S.S.; Mansouri, F.; Marashifard, M.; Malek Hosseini, S.A.A.; Akbarian Chenarestane-Olia, F.; Ganavehei, B.; Gharibpour, F.; Shahbazi, A.; Mirzaii, M.; Darban-Sarokhalil, D. Detection of biofilm related genes, classical enterotoxin genes and agr typing among Staphylococcus aureus isolated from bovine with subclinical mastitis in southwest of Iran. Microb. Pathog. 2016. [Google Scholar] [CrossRef] [PubMed]
- Madigan, M.; Martinko, J.; Bender, K.; Buckley, D.; Stahl, D. Brock Biology of Microorganisms; Pearson Education Ltd.: Edinburgh Gate, UK, 2015. [Google Scholar]
- Park, A.J.; Murphy, K.; Krieger, J.R.; Brewer, D.; Taylor, P.; Habash, M.; Khursigara, C.M. A temporal examination of the planktonic and biofilm proteome of whole cell pseudomonas aeruginosa pao1 using quantitative mass spectrometry. Mol. Cell. Proteom. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shunmugaperumal, T. Biofilm Eradication and Prevention: A Pharmaceutical Approach to Medical Device Infections; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010; ISBN 9780470479964. [Google Scholar]
- Dufrêne, Y.F. Atomic force microscopy in microbiology: New structural and functional insights into the microbial cell surface. MBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Dufrêne, Y.F. Refining our perception of bacterial surfaces with the atomic force microscope. J. Bacteriol. 2004, 186, 3283–3285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, G.; Müller, T.; Meyer, R.L. Single-cell force spectroscopy of bacteria enabled by naturally derived proteins. Langmuir 2014, 30, 4019–4025. [Google Scholar] [CrossRef] [PubMed]
- Relucenti, M.; Familiari, G.; Donfrancesco, O.; Taurino, M.; Li, X.; Chen, R.; Artini, M.; Papa, R.; Selan, L. Microscopy methods for biofilm imaging: Focus on sem and VP-SEM pros and cons. Biology 2021, 10, 51. [Google Scholar] [CrossRef]
- Rutering, J.; Ilmer, M.; Recio, A.; Coleman, M.; Vykoukal, J.; Alt, E.; Orleans, N. Quantitative and Qualitative Assessment Methods for Biofilm Growth: A Mini-review Christina. Nat. Rev Drug Discov. 2016, 5, 1–8. [Google Scholar]
- Wille, G.; Hellal, J.; Ollivier, P.; Richard, A.; Burel, A.; Jolly, L.; Crampon, M.; Michel, C. Cryo-Scanning Electron Microscopy (SEM) and Scanning Transmission Electron Microscopy (STEM)-in-SEM for Bio- and Organo-Mineral Interface Characterization in the Environment. Microsc. Microanal. 2017, 23, 1159–1172. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, O.P.; ka, P. Scanning Electron Microscope: Advantages and Disadvantages in Imaging Components. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1877–1882. [Google Scholar] [CrossRef]
- Magana, M.; Sereti, C.; Ioannidis, A.; Mitchell, C.A.; Ball, A.R.; Magiorkinis, E.; Chatzipanagiotou, S.; Hamblin, M.R.; Hadjifrangiskou, M.; Tegos, G.P. Options and limitations in clinical investigation of bacterial biofilms. Clin. Microbiol. Rev. 2018, 31, e00084-16. [Google Scholar] [CrossRef] [Green Version]
- Gomes, L.C.; Moreira, J.M.R.; Simões, M.; Melo, L.F.; Mergulhão, F.J. Biofilm Localization in the Vertical Wall of Shaking 96-Well Plates. Scientifica 2014, 2014, 1–6. [Google Scholar] [CrossRef]
- Hassan, A.; Usman, J.; Kaleem, F.; Omair, M.; Khalid, A.; Iqbal, M. 05/03/2011 Evaluation of Different Detection Methods of Biofilm Formation in the Clinical Isolates; Elsevier BV: Amsterdam, The Netherlands, 2011; Volume 15, pp. 305–311. [Google Scholar]
- Kaiser, T.D.L.; Pereira, E.M.; dos Santos, K.R.N.; Maciel, E.L.N.; Schuenck, R.P.; Nunes, A.P.F. Modification of the Congo red agar method to detect biofilm production by Staphylococcus epidermidis. Diagn. Microbiol. Infect. Dis. 2013, 75, 235–239. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter plate assay for assessment of Listeria monocytogenes biofilm formation. Appl. Environ. Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latka, A.; Drulis-Kawa, Z. Advantages and limitations of microtiter biofilm assays in the model of antibiofilm activity of Klebsiella phage KP34 and its depolymerase. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Zegaer, B.H.; Ioannidis, A.; Babis, G.C.; Ioannidou, V.; Kossyvakis, A.; Bersimis, S.; Papaparaskevas, J.; Petinaki, E.; Pliatsika, P.; Chatzipanagiotou, S. Detection of Bacteria Bearing Resistant Biofilm Forms, by Using the Universal and Specific PCR is Still Unhelpful in the Diagnosis of Periprosthetic Joint Infections. Front. Med. 2014, 1, 30. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Chen, Y.-P.P.; Qiu, J.-H.H.; Dai, Y.-Z.Z.; Feng, B. Imaging the Microprocesses in Biofilm Matrices; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 37, pp. 214–226. [Google Scholar]
- Agostinho, A.; James, G.; Wazni, O.; Citron, M.; Wilkoff, B.D. Inhibition of Staphylococcus aureus Biofilms by a Novel Antibacterial Envelope for Use with Implantable Cardiac Devices. Clin. Transl. Sci. 2009, 2, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, M.; Wozniak, D.J.; Stoodley, P.; Hall-Stoodley, L. Prevention and treatment of Staphylococcus aureus biofilms. Expert Rev. Anti. Infect. Ther. 2015, 13, 1499–1516. [Google Scholar] [CrossRef] [Green Version]
- Holstege, C.P. Rifampin. In Encyclopedia of Toxicology: Third Edition; Elsevier: Amsterdam, The Netherlands, 2014; pp. 134–136. ISBN 9780123864543. [Google Scholar]
- Rees, C.E.D.; Green, L.H.; Goldman, E.; Loessner, M.J. Practical Handbook of Microbiology; Goldman, E., Green, L.H., Eds.; CRC Press: Boca Raton, FL, USA, 2015; ISBN 9780429168932. [Google Scholar]
- Wells, C.M.; Beenken, K.E.; Smeltzer, M.S.; Courtney, H.S.; Jennings, J.A.; Haggard, W.O. Ciprofloxacin and Rifampin Dual Antibiotic-Loaded Biopolymer Chitosan Sponge for Bacterial Inhibition. Mil. Med. 2018, 183, 433–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vancomycin—StatPearls—NCBI Bookshelf. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK459263/ (accessed on 23 May 2021).
- Salem, A.H.; Elkhatib, W.F.; Noreddin, A.M. Pharmacodynamic assessment of vancomycin-rifampicin combination against methicillin resistant Staphylococcus aureus biofilm: A parametric response surface analysis. J. Pharm. Pharmacol. 2011, 63, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Ramezanpour, M.; Hayes, A.J.; Liu, S.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. Sub-inhibitory clindamycin and azithromycin reduce S. aureus exoprotein induced toxicity, inflammation, barrier disruption and invasion. J. Clin. Med. 2019, 8, 1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.J.; Akers, K.S.; Romano, D.R.; Woodbury, R.L.; Hardy, S.K.; Murray, C.K.; Wenke, J.C. D-amino acids enhance the activity of antimicrobials against biofilms of clinical wound isolates of Staphylococcus aureus and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2014, 58, 4353–4361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warraich, A.A.; Mohammed, A.R.; Perrie, Y.; Hussain, M.; Gibson, H.; Rahman, A. Evaluation of anti-biofilm activity of acidic amino acids and synergy with ciprofloxacin on Staphylococcus aureus biofilms. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Hochbaum, A.I.; Kolodkin-Gal, I.; Foulston, L.; Kolter, R.; Aizenberg, J.; Losick, R. Inhibitory Effects of d-Amino Acids on Staphylococcus aureus Biofilm Development. J. Bacteriol. 2011, 193, 5616. [Google Scholar] [CrossRef] [Green Version]
- Nair, S.; Desai, S.; Poonacha, N.; Vipra, A.; Sharma, U. Antibiofilm activity and synergistic inhibition of Staphylococcus aureus biofilms by bactericidal protein P128 in combination with antibiotics. Antimicrob. Agents Chemother. 2016, 60, 7280–7289. [Google Scholar] [CrossRef] [Green Version]
- Hickey, E.E.; Wong, H.S.; Khazandi, M.; Ogunniyi, A.D.; Petrovski, K.R.; Garg, S.; Page, S.W.; O’Handley, R.; Trott, D.J. Repurposing Ionophores as novel antimicrobial agents for the treatment of bovine mastitis caused by Gram-positive pathogens. J. Vet. Pharmacol. Ther. 2018, 41, 746–754. [Google Scholar] [CrossRef]
- Bazargani, M.M.; Rohloff, J. Antibiofilm activity of essential oils and plant extracts against Staphylococcus aureus and Escherichia coli biofilms. Food Control 2016, 61, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Monteiro-Neto, V.; de Souza, C.D.; Gonzaga, L.F.; da Silveira, B.C.; Sousa, N.C.F.; Pontes, J.P.; Santos, D.M.; Martins, W.C.; Pessoa, J.F.V.; Carvalho Júnior, A.R.; et al. Cuminaldehyde potentiates the antimicrobial actions of ciprofloxacin against Staphylococcus aureus and Escherichia coli. PLoS ONE 2020, 15. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; d’Acierno, A.; Coppola, R.; Jesus Ayala-Zavala, F.; Gomez da Cruz, A.; De Feo, V. Essential Oils and Microbial Communication. In Essential Oils–Oils of Nature; IntechOpen: Lobdon, UK, 2020. [Google Scholar]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Agents that inhibit bacterial biofilm formation. Future Med. Chem. 2015, 7, 647–671. [Google Scholar] [CrossRef]
- Parrino, B.; Schillaci, D.; Carnevale, I.; Giovannetti, E.; Diana, P.; Cirrincione, G.; Cascioferro, S. Synthetic small molecules as anti-biofilm agents in the struggle against antibiotic resistance. Eur. J. Med. Chem. 2019, 161, 154–178. [Google Scholar] [CrossRef] [PubMed]
- Fontecha-Umaña, F.; Ríos-Castillo, A.G.; Ripolles-Avila, C.; Rodríguez-Jerez, J.J. Antimicrobial activity and prevention of bacterial biofilm formation of silver and zinc oxide nanoparticle-containing polyester surfaces at various concentrations for use. Foods 2020, 9, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamida, R.S.; Ali, M.A.; Goda, D.A.; Khalil, M.I.; Al-Zaban, M.I. Novel Biogenic Silver Nanoparticle-Induced Reactive Oxygen Species Inhibit the Biofilm Formation and Virulence Activities of Methicillin-Resistant Staphylococcus aureus (MRSA) Strain. Front. Bioeng. Biotechnol. 2020, 8, 433. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Virulence Factors | Genes | Clinical Implications | |
|---|---|---|---|
| Toxins | Hemolysin | Hla | Food poisoning, toxic shock syndrome, scalded skin syndrome, bullous impetigo and sepsis syndrome |
| Leukotoxin | lukD and E | ||
| Exfoliative toxin | eta and etb | ||
| Toxic shock syndrome toxin 1 | tstH | ||
| Exoenzymes | Lipases | Geh | Tissue destruction and metastatic infections |
| Proteases | aur, sspA, B and C | ||
| Nucleases | nuc1 and nuc2 | ||
| Coagulases | coa and vWbp | ||
| Hyaluronate lyase | hysA | ||
| Immunomodulators | Leucocidin | lukS-PV and lukF-PV | Invasive skin infections, pneumonia and abscesses |
| Extra cellular adherence protein | Eap | ||
| Capsular polysaccharides | cap5 and cap8 | ||
| Phenol-soluble modulins | psm-α | ||
| Other | Attachment | clfA and B, fnbA and B, cna and ica | Endocarditis, septic arthritis, prosthetic devices and catheter infections, cystic fibrosis and relapsing infections |
| Persistence | Ica locus and hemB | ||
| Sr. no | Techniques | Summary | Advantages | Disadvantages | References |
|---|---|---|---|---|---|
| a. | Direct observation techniques | ||||
| 1. | Light microscope | Visualization of biofilm structure. | Cheap, convenient and easy to perform. Simple sample preparation. | Restricted resolution and magnification. Sample staining required. Lacks discriminatory features. | [97,100] |
| 2 | Transmission electron microscope | Images of cell components on the biofilm surface and within the matrix are directly visualized with negative staining. | Biofilm labelling. Several observational modes such as nanometric scale and elemental evaluation. | Expensive method. Long fixative procedure. Possible detachment of biofilm during fixative procedures. | [101] |
| 3. | Scanning electron microscope | Two-dimensional and topographical imaging of biofilm structure. | Allows to examine the biofilm matrix on the growth substratum. Presents observational modes such as nanometric scale imaging. High magnification and resolution. | Expensive and high maintenance. Risk of artifacts. Samples must be solid and non-conductive. Conductive material coating and dehydration causes biofilm shrinkage. Time-consuming technique. | [97,99,102] |
| 4. | Scanning transmission X-ray microscope | Quantitative and qualitative explorations of biofilm structure and analysing the array of microbial communities. | High resolution. Quantitative mapping of biofilm components such as lipids, proteins, nucleic acids and saccharides. Provides spectral outline for each component. | Limited accuracy. Risk of instrumental systematic error. Utilized on thin samples. | [76,103] |
| b | Fluorescent tagging of biofilm | ||||
| 1. | Confocal laser scanning microscope | Three-dimensional morphology and quantitative imaging of biofilm physiology. | Non-invasive technique. Living, hydrated samples. | Interference of biofilm components with the fluorescence probes. Restricted number of reporter molecules. Limited choice of magnification. | [76,97,104] |
| 2. | Fluorescent in situ hybridization (FISH) | Semi-quantitative technique to identify specific organism in a multispecies biofilm population with fluorescent probes. | Applicable to heterogenous biofilm community. Detection of live microorganisms. | Low sensitivity due to non-specific hybridization of complementary probes. Tedious procedure and expensive requirements. | [103,104] |
| C | Indirect observational technique | ||||
| 1. | Tube method | Qualitative detection by presence of visible biofilm lining around the wall and the bottom of the tube. | Identifies strong biofilm producers. | Fails to differentiate between weak, moderate or non-biofilm producers due to the variability in the findings identified by different viewers. | [105] |
| 2. | Congo Red agar method (CRA) | Qualitative method by examining the colony colour change on Congo red agar (CRA) medium. | Cheap and easy to perform. | Substantial low specificity, sensitivity and positive predictive value. | [105,106] |
| 3. | Microtiter plate assays | Quantitative evaluation of biofilm formation in the wells detected by microplate reader. | Quick and simple screening assay to quantify the biofilm formation. Antimicrobial susceptibility assay | Low reproducibility. Non-specificity with crystal violet dyes. Variation in biofilm biomass, depending on the washing step. Limited substratum alternatives. | [107,108] |
| 4. | Biofilm-associated genes detection by PCR | Detection of biofilm-associated genes in microorganisms | Presents sharper specificity, sensitivity and time efficient. Reliable and reproducible | Possibility of sample contamination, false positive results or misinterpretation. High-priced PCR requirements. | [103,109] |
| 5. | Mass spectrometry | Detection of proteins and enzymes expressed with EPS matrix. | Provides identification of proteins, chemical components and mass-based variation of analogous molecules. Detection of chemical heterogeneity and secondary metabolites, even in multispecies biofilms. Cell-level and macroscopic chemical alterations. | Imaging artifacts. Sample surface requires chemical modification. | [103,110] |
| 6. | Atomic force microscopy | Utilized to map distributions of EPS, biomass, chemical and molecular compounds with a physical probe tip scanning the sample surface. | Minimal pre-treatment procedures and artifacts. Three dimensional images. Qualitative imaging of EPS. High resolution. Elucidation of cellular and molecular interactions. Imaging samples at the nanometre to micrometre scale. | Small scanning area (max 150 × 150 µm). Risk of surface damage due to sample-probe tip interactions. | [77,95,96,97] |
| Antimicrobial Agents | Examples | Mechanism of Action Against S. aureus Biofilm |
|---|---|---|
| AMPs | NA-CATH: ATRA1-ATRA1 | Prevention of gene expression that encodes the formation of biofilm-related proteins. Attachment to eDNA |
| Functional Excipients | D-amino acids | Disruption of eDNA |
| Plant Extracts and Essential Oils | Garlic and ginseng extracts | Disruption of bacterial quorum sensing |
| Melaleuca bracteate leaves oil | Inhibition of biofilm-related proteins formation and disruption of quorum sensing | |
| Enzymes | DNase I, Dispersin B | Degradation of eDNA |
| a-amylase | Degradation of exopolysaccharides | |
| Nanoparticles | Silver, zinc oxide | Unknown |
| Antibiotics | Azithromycin | Inhibition of EPS related proteins formation and disruption of quorum sensing |
| Ciprofloxacin, rifampin, amoxicillin, clindamycin, vancomycin, etc. | Except azithromycin, mechanism of action for antibiofilm activity of most of these antibiotics is not fully understood | |
| Ionophores | Unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies. Int. J. Environ. Res. Public Health 2021, 18, 7602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18147602
Idrees M, Sawant S, Karodia N, Rahman A. Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies. International Journal of Environmental Research and Public Health. 2021; 18(14):7602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18147602
Chicago/Turabian StyleIdrees, Muhammad, Sheeba Sawant, Nazira Karodia, and Ayesha Rahman. 2021. "Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies" International Journal of Environmental Research and Public Health 18, no. 14: 7602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18147602