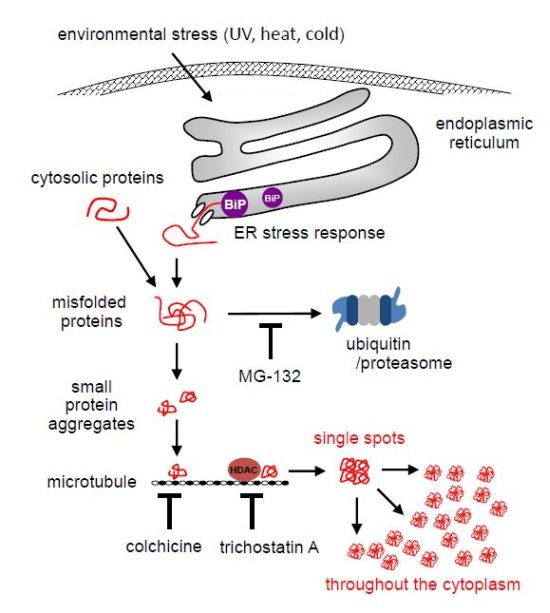
Environmental Stresses Induce Misfolded Protein Aggregation in Plant Cells in a Microtubule-Dependent Manner



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
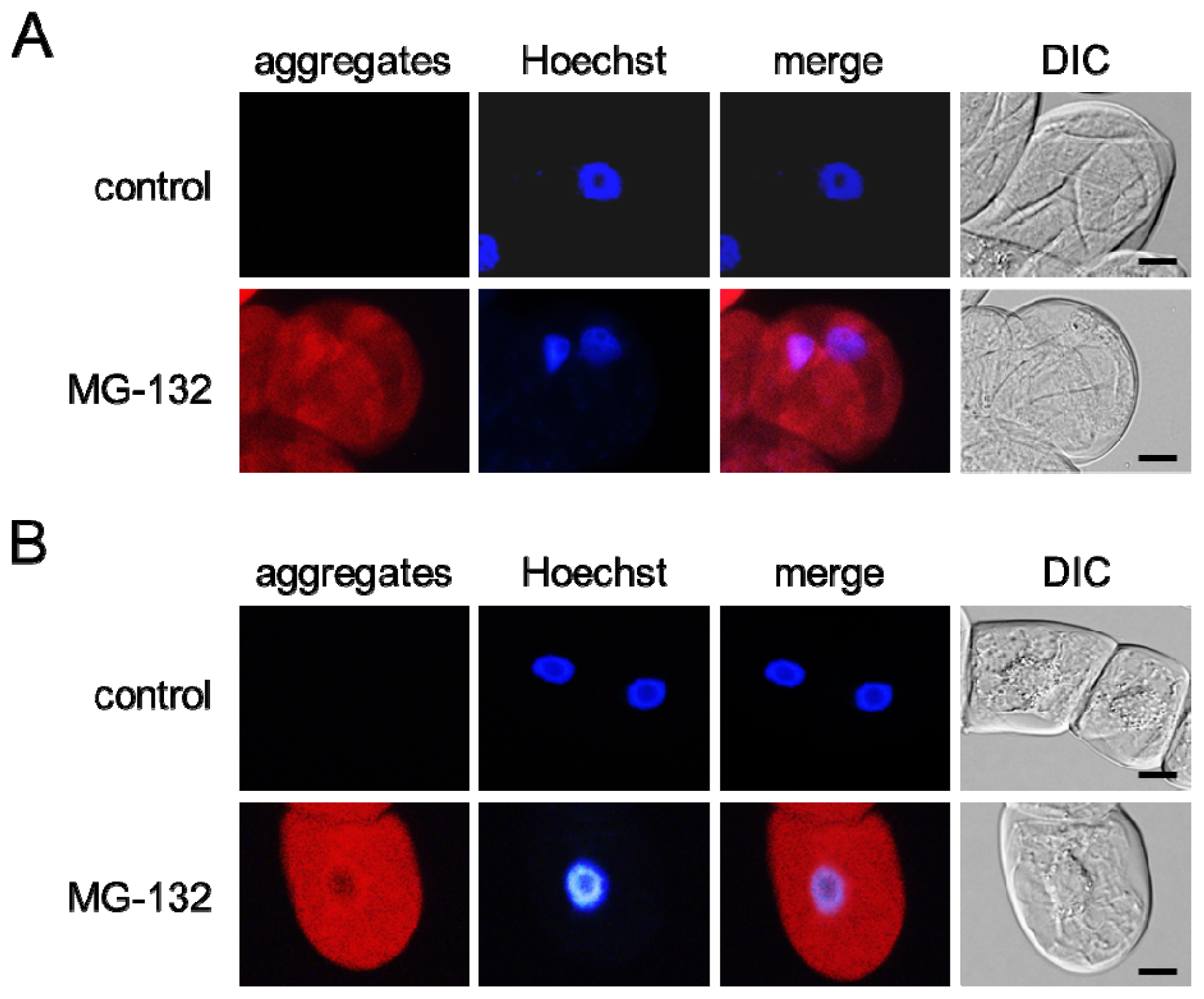
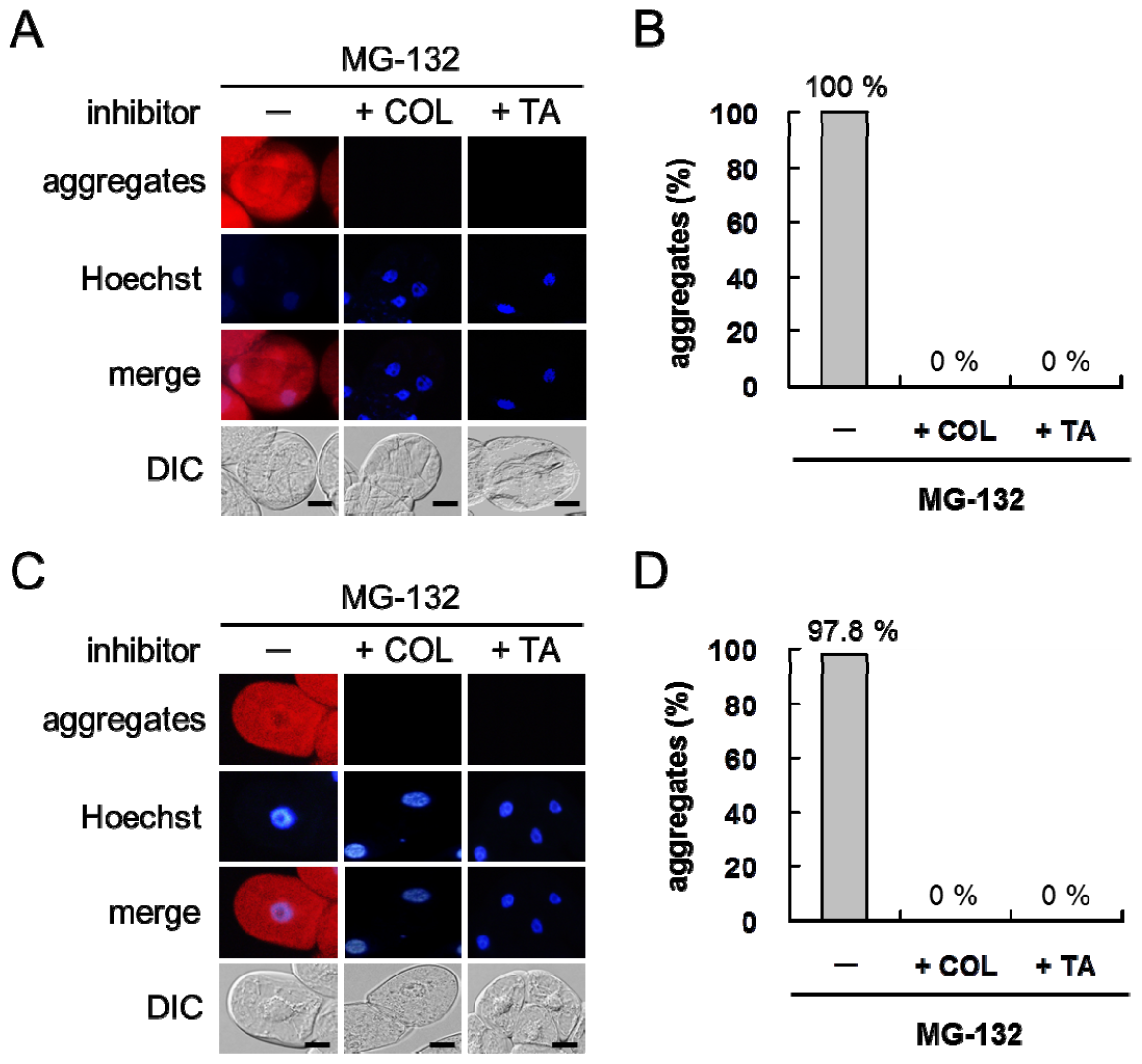
2.1. MG-132 Induces Misfolded Protein Aggregation in Plant Cells in a Microtubule-Dependent Manner
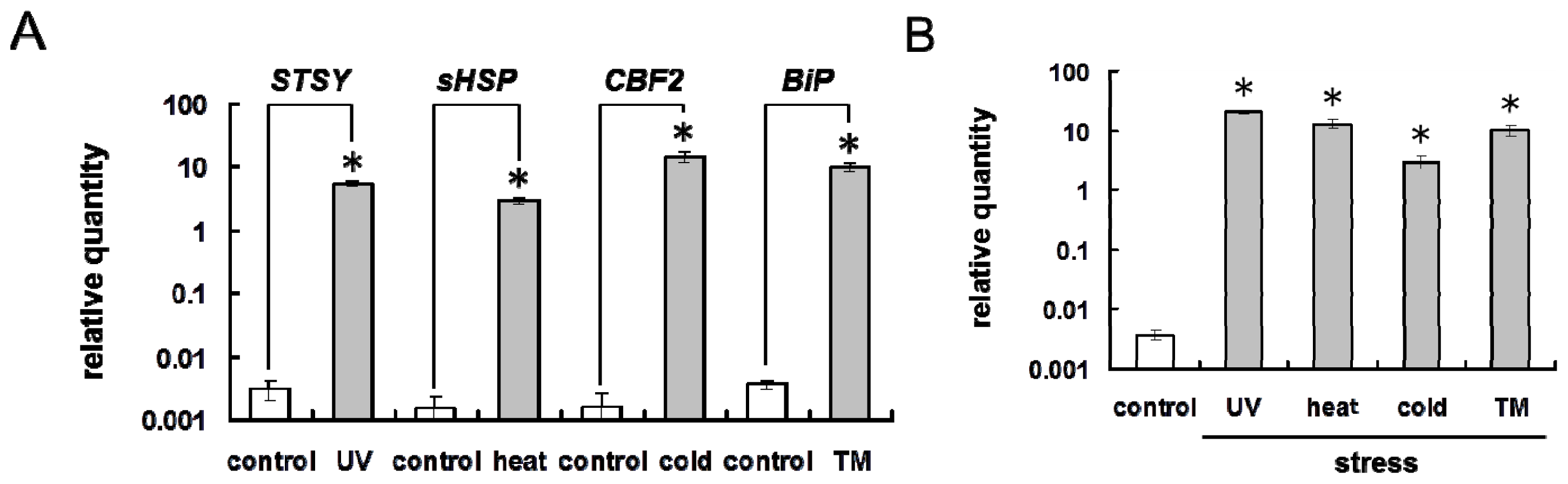
2.2. Environmental Stresses Induce ER Stress Response in Plant Cells
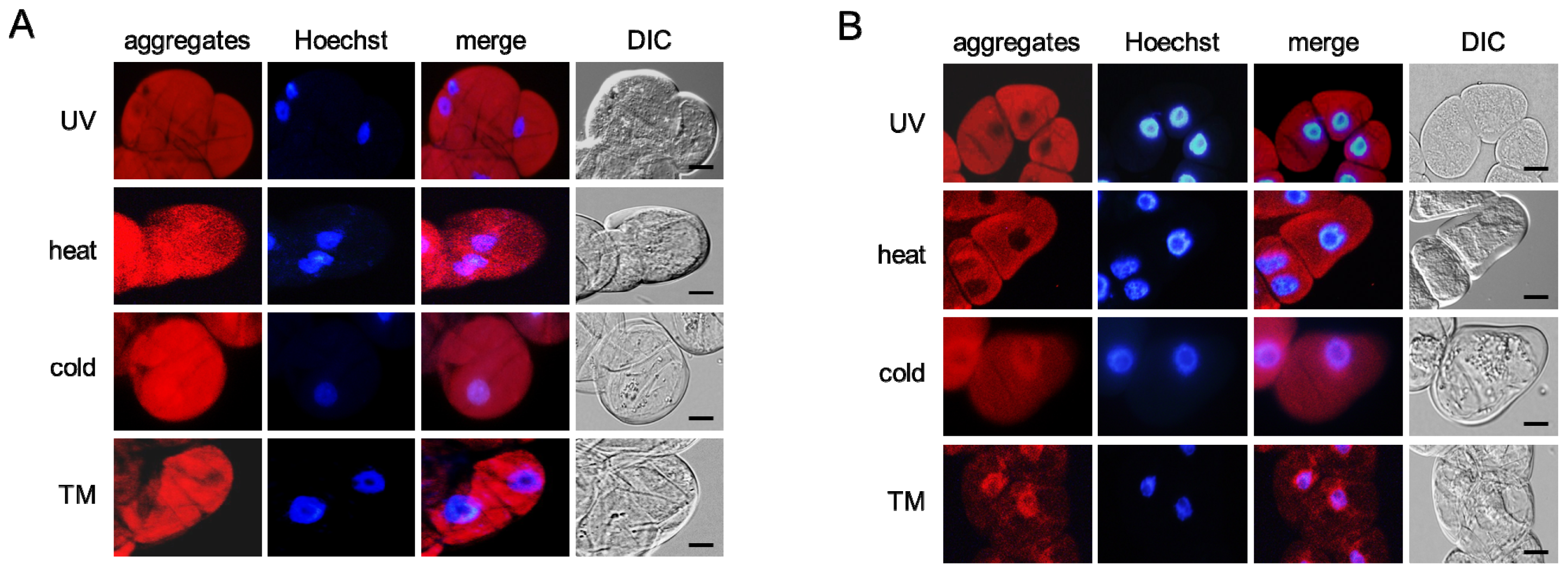
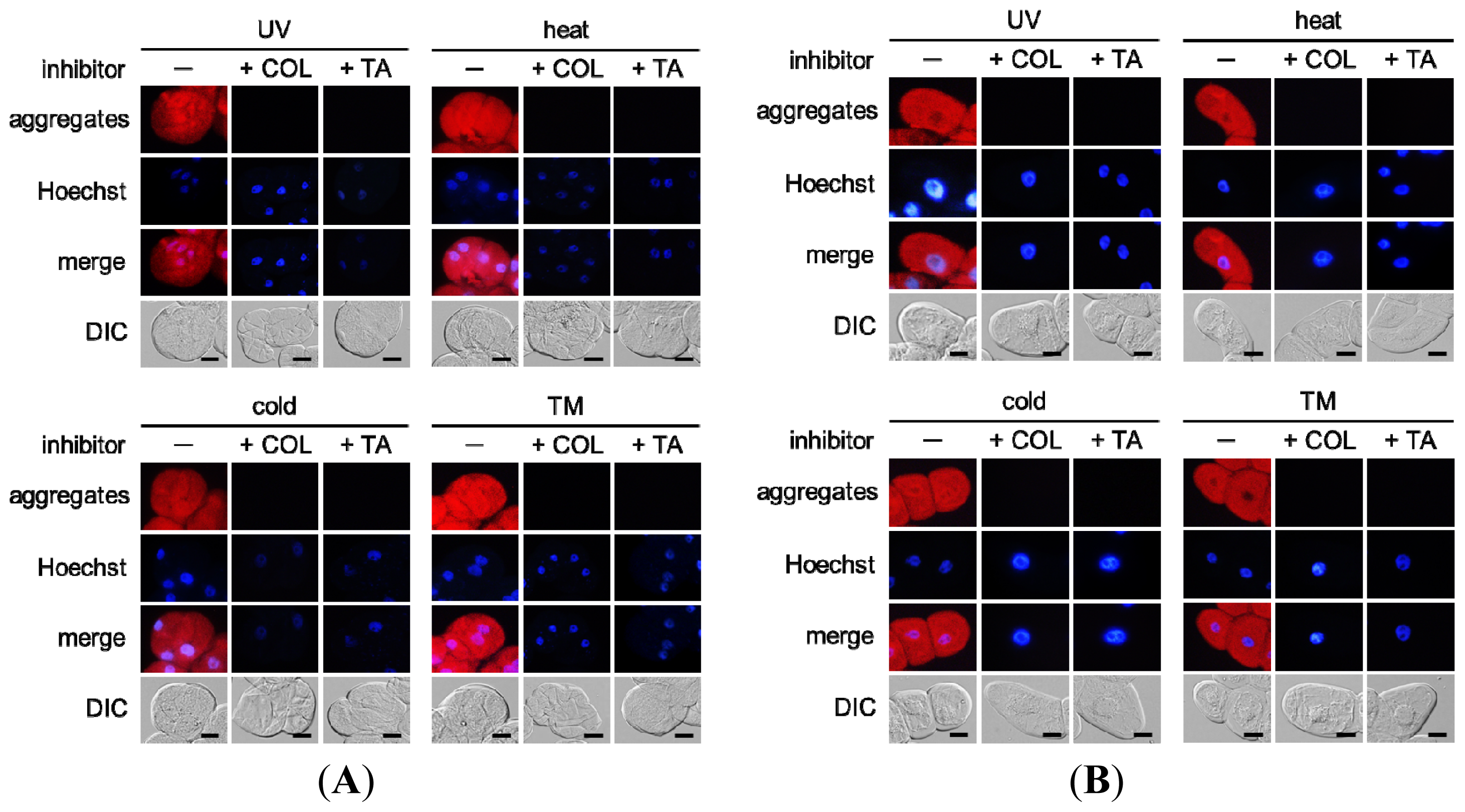
2.3. Environmental Stresses Induce Misfolded Protein Aggregation in Plant Cells in a Microtubule-Dependent Manner
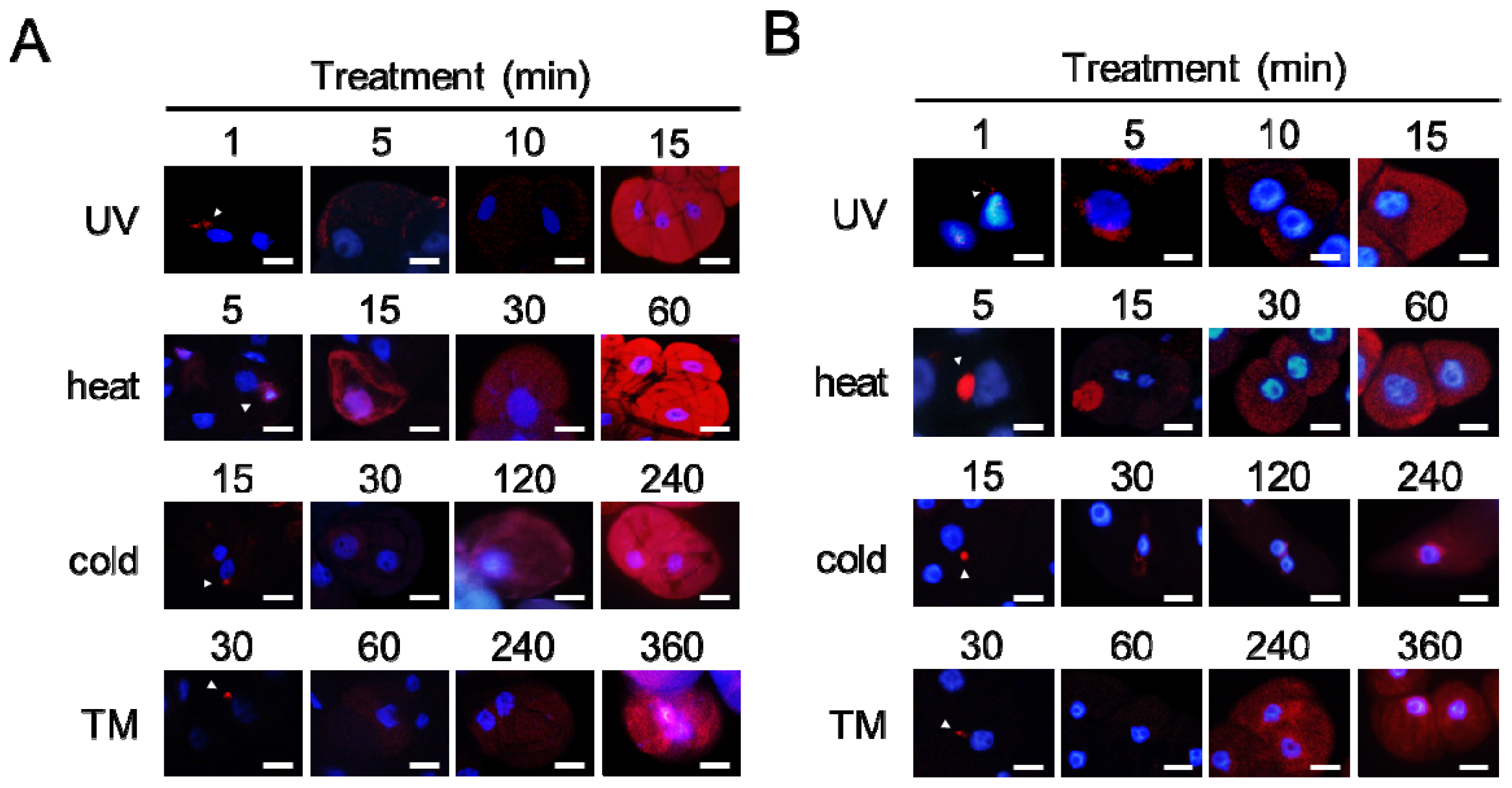
2.4. Aggregates Are Rapidly Accumulated
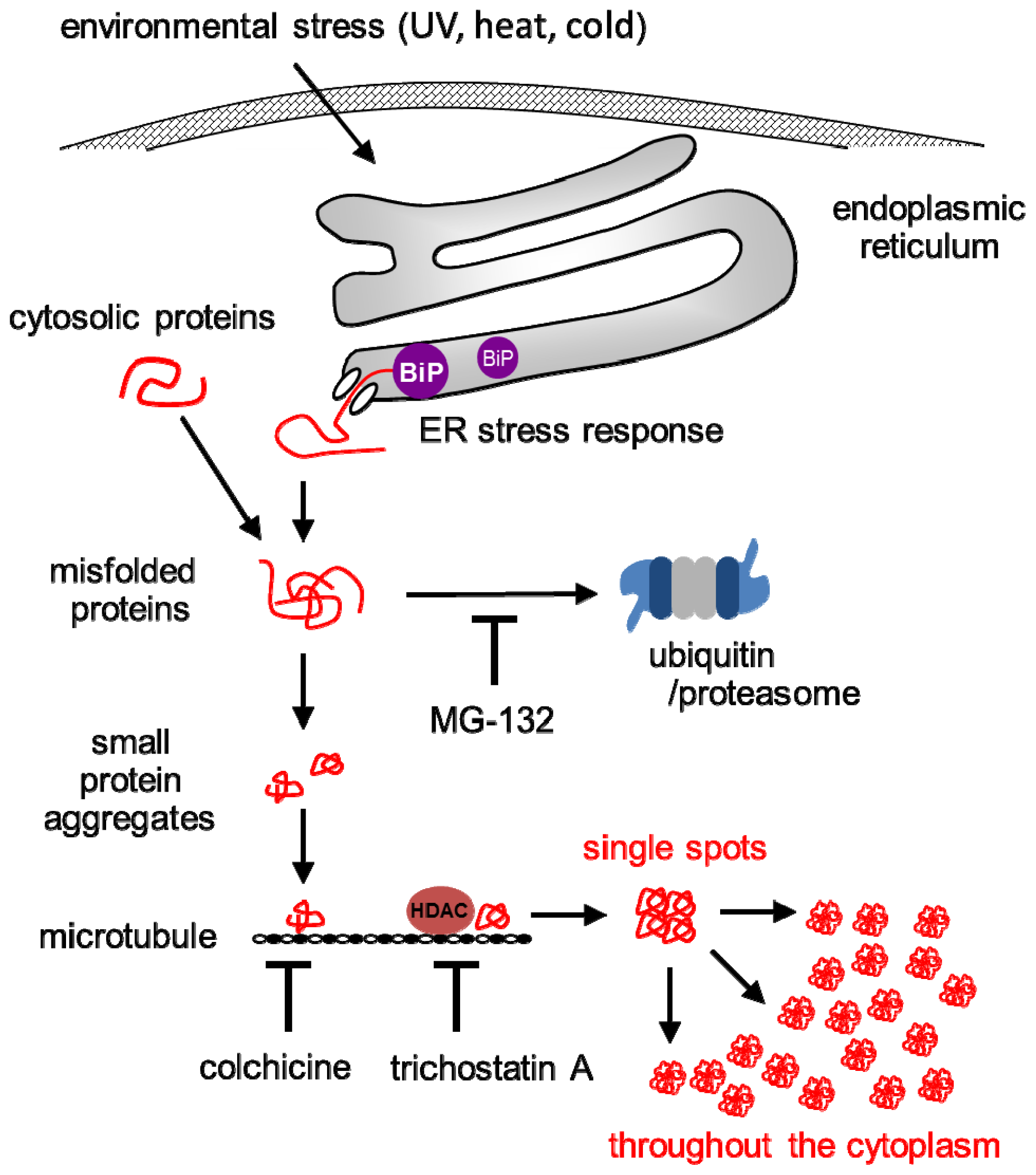
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Materials
4.3. Chemical Treatment
4.4. Stress Treatment
4.5. Staining of Misfolded Proteins
4.6. RNA Isolation
4.7. Quantitative RT-PCR Analysis
4.8. Statistical Analysis
5. Conclusion
Acknowledgements
References
- Vembar, S.S.; Brodsky, J.L. One step at a time: Endoplasmic reticulum-associated degradation. Nature Rev. Mol. Cell Biol 2008, 9, 944–957. [Google Scholar]
- Kamauchi, S.; Nakatani, H.; Nakano, C.; Urade, R. Gene expression in response to endoplasmic reticulum stress in Arabidopsis thaliana. FEBS J 2005, 272, 3461–3476. [Google Scholar]
- Vidair, A.; Huang, R.N.; Doxsey, S.J. Heat shock causes protein aggregation and reduced protein solubility at the centrosome and other cytoplasmic locations. Int. J. Hyperth 1996, 12, 12681–12695. [Google Scholar]
- Buchner, J. Supervising the fold: Functional principles of molecular chaperones. FASEB J 1996, 10, 10–19. [Google Scholar]
- Levitskaya, J.; Sharipo, A.; Leonchiks, A.; Ciechanover, A.; Masucci, M.G. Inhibition of ubiquitin/proteasome-dependent protein degradation by the Gly-Ala repeat domain of the Epstein-Barr virus nuclear antigen 1. Proc. Natl. Acad. Sci. USA 1997, 94, 12616–12621. [Google Scholar]
- Liu, J.X.; Howell, S.H. Endoplasmic reticulum protein quality control and its relationship to environmental stress responses in plants. Plant Cell 2010, 22, 2930–2942. [Google Scholar]
- Katoh, H.; Fujita, K.; Takuhara, Y.; Ogawa, A.; Suzuki, S. ER stress-induced protein, VIGG, disturbs plant cation homeostasis, which is correlated with growth retardation and robustness to ER stress. Biochem. Biophys. Res. Commun 2011, 405, 514–520. [Google Scholar]
- Vogel, F.; Hofius, D.; Sonnewald, U. Intracellular trafficking of Potato leafroll virus movement protein in transgenic Arabidopsis. Traffic 2007, 8, 1205–1214. [Google Scholar]
- Martinière, A.; Gargani, D.; Uzest, M.; Lautredou, N.; Blanc, S.; Drucker, M. A role for plant microtubules in the formation of transmission-specific inclusion bodies of Cauliflower mosaic virus. Plant J 2009, 58, 135–146. [Google Scholar]
- Niehl, A.; Pena, E.J.; Amari, K.; Heinlein, M. Microtubules in viral replication and transport. Plant J 2013. [Google Scholar] [CrossRef]
- Niehl, A.; Amari, K.; Gereige, D.; Brandner, K.; Mély, Y.; Heinlein, M. Control of Tobacco mosaic virus movement protein fate by CELL-DIVISION-CYCLE Protein48. Plant Physiol 2012, 160, 2093–2108. [Google Scholar]
- Saliba, R.S.; Munro, P.M.G.; Luthert, P.J.; Cheetham, M.E. The cellular fate of mutant rhodopsin: quality control, degradation and aggresome formation. J. Cell Sci 2002, 115, 2907–2918. [Google Scholar]
- Kothawala, A.; Kilpatrick, K.; Novoa, J.A.; Segatori, L. Quantitative analysis of α-synuclein solubility in living cells using split GFP complementation. PLoS One 2012, 7, e43505. [Google Scholar]
- Ohnishi, K.; Nakahata, E.; Irie, K.; Murakami, A. Zerumbone, an electrophilic sesquiterpene, induces cellular proteo-stress leading to activation of ubiquitin-proteasome system and autophagy. Biochem. Biophys. Res. Commun 2013, 430, 616–622. [Google Scholar]
- Kobayashi, M.; Katoh, H.; Takayanagi, T.; Suzuki, S. Characterization of thermotolerance-related genes in grapevine (Vitis vinifera). J. Plant Physiol 2010, 167, 812–819. [Google Scholar]
- Takuhara, Y.; Kobayashi, M.; Suzuki, S. Low-temperature-induced transcription factors in grapevine enhance cold tolerance in transgenic Arabidopsis plants. J. Plant Physiol 2011, 168, 967–975. [Google Scholar]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic interaction of BiP and ER stress transducers in the unfolded protein response. Nat. Cell Biol 2000, 2, 326–332. [Google Scholar]
- Johnston, J.A.; Ward, C.L.; Kopito, R.R. Aggresomes: A cellular response to misfolded proteins. J. Cell Biol 1998, 143, 1883–1898. [Google Scholar]
- Garcia-Mata, R.; Gao, Y.S.; Sztul, E. Hassles with taking out the garbage: Aggravating aggresomes. Traffic 2002, 3, 388–396. [Google Scholar]
- Murata, T.; Hasebe, M. Microtubule-dependent microtubule nucleation in plant cells. J. Plant Res 2007, 120, 73–78. [Google Scholar]
- Kawaguchi, Y.; Kovacs, J.J.; McLaurin, A.; Vance, J.M.; Ito, A.; Yao, T.P. The deacetylase HDAC6 regulates aggresome formation and cell viability in response to misfolded protein stress. Cell 2003, 115, 727–738. [Google Scholar]
- Nagata, T.; Nemoto, Y.; Hasezawa, S. Tobacco BY-2 cell line as the “HeLa” cell in the cell biology of higher plants. Int. Rev. Cytol 1992, 132, 1–30. [Google Scholar]
- Katoh, H.; Suzuki, S.; Saitoh, T.; Takayanagi, T. Cloning and characterization of VIGG, a novel virus-induced grapevine protein, correlated with fruit quality. Plant Physiol. Biochem 2009, 47, 291–299. [Google Scholar]







© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nakajima, Y.; Suzuki, S. Environmental Stresses Induce Misfolded Protein Aggregation in Plant Cells in a Microtubule-Dependent Manner. Int. J. Mol. Sci. 2013, 14, 7771-7783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14047771
Nakajima Y, Suzuki S. Environmental Stresses Induce Misfolded Protein Aggregation in Plant Cells in a Microtubule-Dependent Manner. International Journal of Molecular Sciences. 2013; 14(4):7771-7783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14047771
Chicago/Turabian StyleNakajima, Yuko, and Shunji Suzuki. 2013. "Environmental Stresses Induce Misfolded Protein Aggregation in Plant Cells in a Microtubule-Dependent Manner" International Journal of Molecular Sciences 14, no. 4: 7771-7783. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms14047771