The Wheat E Subunit of V-Type H+-ATPase Is Involved in the Plant Response to Osmotic Stress

Abstract
:1. Introduction
2. Results
2.1. cDNA Isolation and Homology Analysis
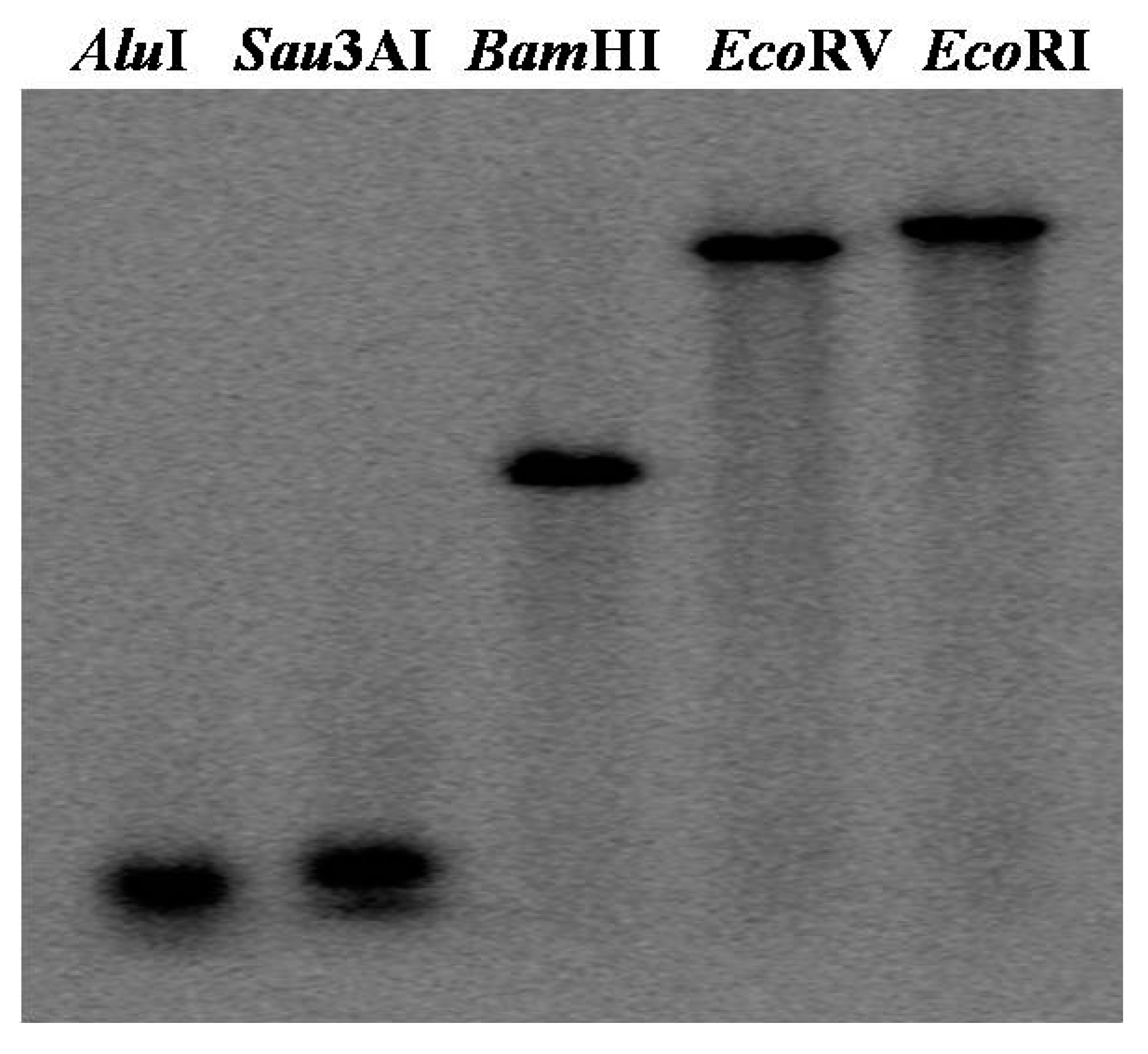
2.2. W36 Copy Number Analysis


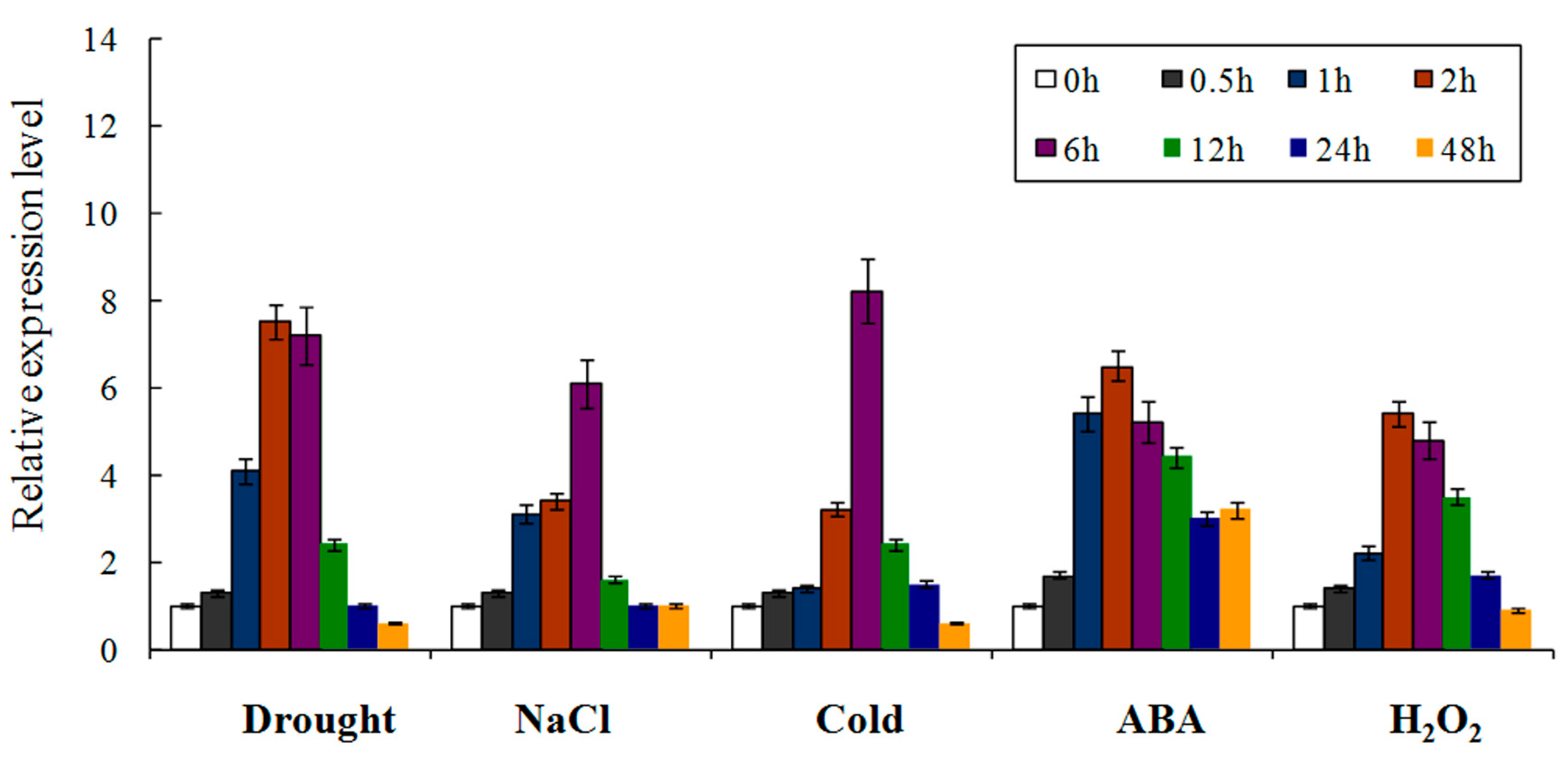
2.3. W36 Expression Characteristics under Various Stress Conditions
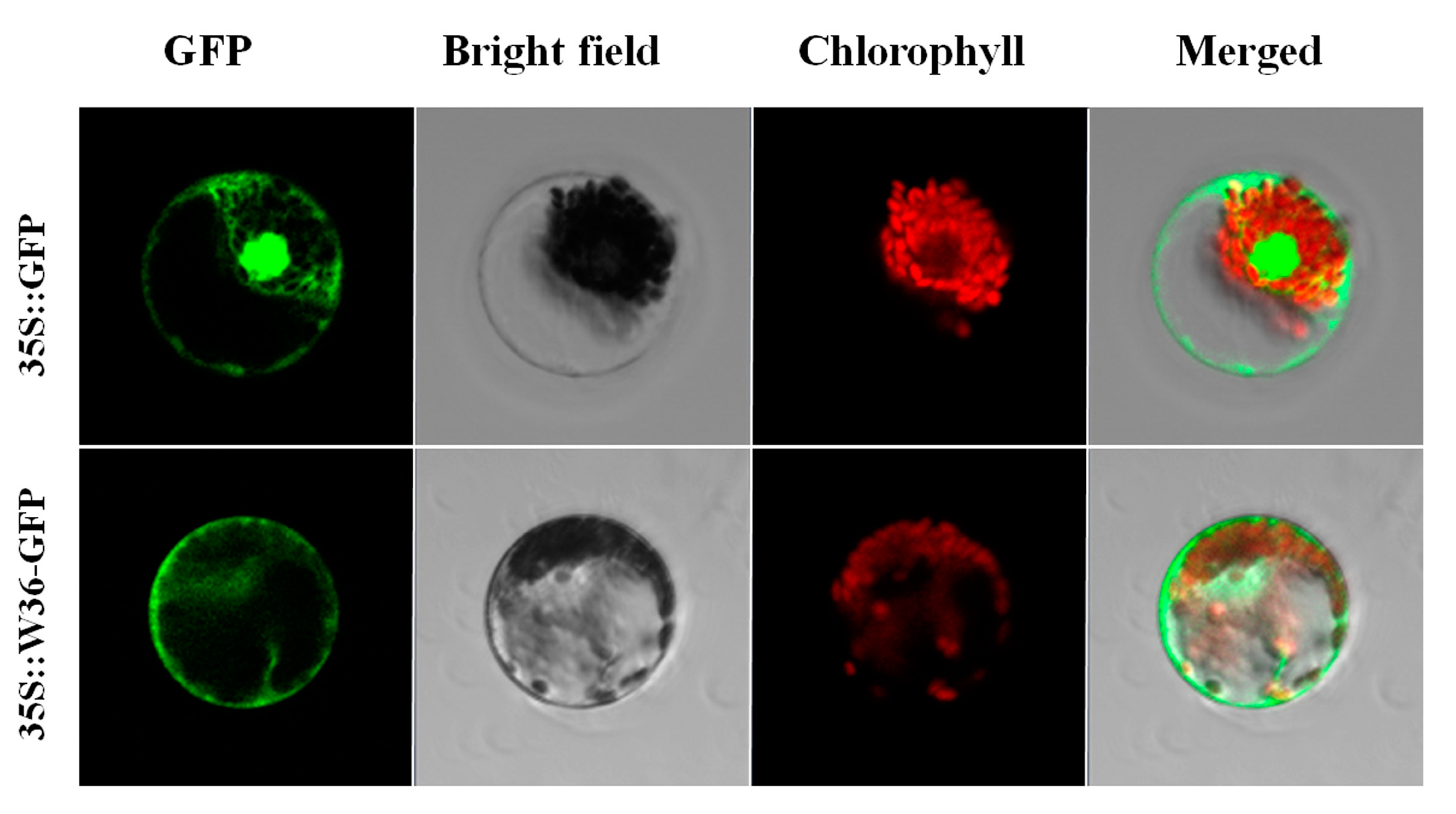
2.4. W36 Subcellular Localization


2.5. Characterization of the 5'-Flanking Region of W36

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elements | Core Sequence | Functions |
|---|---|---|
| TATA-box | TAATA | Core promoter |
| ABRE | ACGTG | ABA and drought responsive elements |
| ARE | TGGTTT | Anaerobic responsive element |
| CGTCA-motif | CGTCA | MeJA responsive elements |
| ERE | ATTTCAAA | Ethylene responsive elements |
| TGA-element | AACGAC | Auxin responsive elements |
| GARE-motif | AAACAGA | Gibberellin responsive elements |
| GC-motif | CCCCCG | Anoxic responsive elements |
| HSE | AAAAAATTTC | Heat shock responsive element |
| TC-rich repeats | GTTTTCTTAC | Element involved in defense and stress responsiveness |
| TCA-element | CCATCTTTTT | Salicylic acid responsive elements |
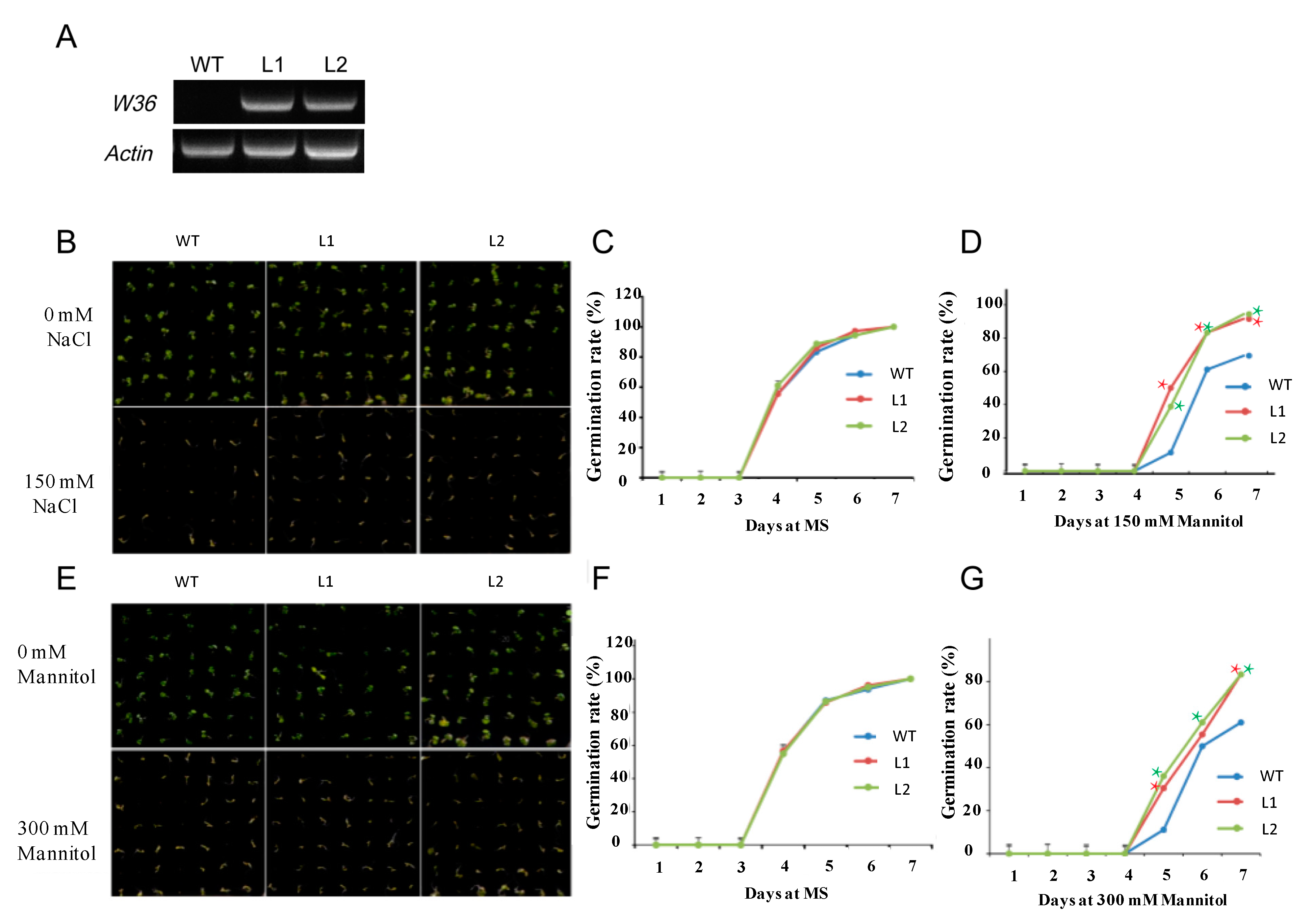
2.6. Overexpression of W36 Improves Tolerance to Osmotic Stress in Transgenic Plants

3. Experimental Section
3.1. Plant Materials and Stress Treatments
3.2. De Novo Transcriptome Sequencing of Wheat
3.3. Homology Analysis
3.4. DNA Extraction and Southern Blotting
3.5. qRT-PCR Analysis
3.6. W36 Subcellular Localization
3.7. Amplification of the 5'-Flanking Region of W36
3.8. Arabidopsis Transformation and Stress Treatment
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Alvarez, I.; Tomaro, M.L.; Benavides, M.P. Changes in polyamines, proline and ethylene in sunflower calluses treated with NaCl. Plant Cell Tissue Organ Cult. 2003, 74, 51–59. [Google Scholar] [CrossRef]
- Niu, X.M.; Narasimhan, M.L.; Salzman, R.A.; Bressan, R.A.; Hasegawa, P.M. NaCl regulation of plasma membrane H+-ATPase gene expression in a glycophyte and halophyte. Plant Physiol. 1993, 103, 713–718. [Google Scholar]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Li, P.H.; Chen, M.; Wang, B.S. Effect of K+ nutrition on growth and activity of leaf tonoplast V-H+-ATPase and V-PPase of Suaed salsa under NaCl stress. Acta Bot. Sin. 2002, 44, 433–440. [Google Scholar]
- Hunte, C.; Screpanti, E.; Venturi, M.; Rimon, A.; Padan, E.; Michel, H. Structure of a Na+/H+ anti-porter and insights into mechanism of action and regulation by pH. Nature 2005, 435, 1197–1202. [Google Scholar] [CrossRef]
- Otoch, M.D.L.O.; Sobreira, A.C.M.; Aragao, M.E.F.D.; Orellano, E.G.; Lima, M.D.G.S.; Melo, D.F.D. Salt modulation of vacuolar H+-ATPase and H+-Pyrophosphatase activities in Vigna unguiculata. Plant Physiol. 2001, 158, 545–551. [Google Scholar] [CrossRef]
- Qiu, N.W.; Chen, M.; Guo, J.R.; Bao, H.Y.; Ma, X.L.; Wang, B.S. Coordinate up-regulation of V-H+-ATPase and vacuolar Na+/H+ antiporter as a response to NaCl treatment in a C3 halophyte Suaeda salsa. Plant Sci. 2007, 172, 1218–1225. [Google Scholar] [CrossRef]
- Yang, M.F.; Song, J.; Wang, B.S. Organ-specific responses of vacuolar H-ATPase in the shoots and roots of C halophyte Suaeda salsa to NaCl. J. Integr. Plant Biol. 2010, 52, 308–314. [Google Scholar] [CrossRef]
- Gaxiola, R.A.; Palmgren, M.G.; Schumacher, K. Plant proton pumps. FEBS Lett. 2007, 581, 2204–2214. [Google Scholar] [CrossRef]
- Rober-Kleber, N.; Albrechtova, J.T.; Fleig, S.; Huck, N.; Michalke, W.; Wagner, E.; Speth, V.; Neuhaus, G.; Fischer-Iglesias, C. Plasma membrane H+-ATPase is involved in auxin-mediated cell elongation during wheat embryo development. Plant Physiol. 2003, 131, 1302–1312. [Google Scholar] [CrossRef]
- Fuglsang, A.T.; Guo, Y.; Cuin, T.A.; Qiu, Q.; Song, C.; Kristiansen, K.A.; Bych, K.; Schulz, A.; Shabala, S.; Schumaker, K.S.; et al. Arabidopsis protein kinase PKS5 inhibits the plasma membrane H+-ATPase by preventing interaction with 14-3-3 protein. Plant Cell 2007, 19, 1617–1634. [Google Scholar] [CrossRef]
- Janicka-Russak, M.; Kabała, K.; Wdowikowska, A.; Kłobus, G. Response of plasma membrane H+-ATPase to low temperature in cucumber roots. J. Plant Res. 2012, 125, 291–300. [Google Scholar] [CrossRef]
- Palmgren, M.G. Plant plasma membrane H+-ATPases: Power-houses for nutrient uptake. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 817–845. [Google Scholar] [CrossRef]
- Seidel, T.; Kluge, C.; Hanitzsch, M.; Ross, J.; Sauer, M.; Dietz, K.J.; Golldack, D. Colocalization and FRET analysis of subunits c and a of the vacuolar H+-ATPase in living plant cells. J. Biotechnol. 2004, 112, 165–175. [Google Scholar] [CrossRef]
- Merlot, S.; Leonhardt, N.; Fenzi, F.; Valon, C.; Costa, M.; Piette, L.; Vavasseur, A.; Genty, B.; Boivin, K.; Muller, A.; et al. Constitutive activation of a plasma membrane H+-ATPase prevents abscisic acid-mediated stomatal closure. EMBO J. 2007, 26, 3216–3226. [Google Scholar] [CrossRef]
- Gevaudant, F.; Duby, G.; von Stedingk, E.; Zhao, R.; Morsomme, P.; Boutry, M. Expression of a constitutively activated plasma membrane H+-ATPase alters plant development and increases salt tolerance. Plant Physiol. 2007, 144, 1763–1776. [Google Scholar] [CrossRef]
- Nishi, T.; Forgac, M. The vacuolar (H+)-ATPases—Nature’s most versatile proton pumps. Nat. Rev. Mol. Cell Biol. 2002, 3, 94–103. [Google Scholar] [CrossRef]
- Seidel, T. Structure and regulation of plant vacuolar H+-ATPase. In Progress in Botany; Lüttge, U., Ed.; Springer Berlin Heidelberg: Berlin, Germany, 2009; Volume 70, pp. 93–126. [Google Scholar]
- Sze, H.; Schumacher, K.; Müller, M.L.; Padmanaban, S.; Taiz, L. A simple nomenclature for a complex proton pump: VHA genes encode the vacuolar H+-ATPase. Trends Plant Sci. 2002, 7, 157–161. [Google Scholar] [CrossRef]
- Padmanaban, S.; Lin, X.; Perera, I.; Kawamura, Y.; Sze, H. Differential expression of vacuolar H+-ATPase subunit c genes in tissues active in membrane trafficking and their roles in plant growth as revealed by RNAi. Plant Physiol. 2004, 134, 1514–1526. [Google Scholar] [CrossRef]
- Gogarten, J.P.; Kibak, H.; Dittrich, P.; Taiz, L.; Bowman, E.J.; Bowman, B.J.; Manolson, M.F.; Poole, R.J.; Date, T.; Oshima, T.; et al. Evolution of the vacuolar H+-ATPase: Implications for the origin of eukaryotes. Proc. Natl. Acad. Sci. USA 1989, 86, 6661–6665. [Google Scholar] [CrossRef]
- PlantCARE. Available online: http://bioinformatics.psb.ugent.be/webtools/ plantcare/html/ (accessed on 12 September 2014).
- Liu, P.; Xu, Z.S.; Lu, P.P.; Hu, D.; Chen, M.; Li, L.C.; Ma, Y.Z. A wheat PI4K gene whose product possesses threonine autophophorylation activity confers tolerance to drought and salt in Arabidopsis. J. Exp. Bot. 2013, 64, 2915–2927. [Google Scholar] [CrossRef]
- Jiang, A.L.; Xu, Z.S.; Zhao, G.Y.; Cui, X.Y.; Chen, M.; Li, L.C.; Ma, Y.Z. Genome-wide analysis of the C3H zinc finger transcription factor family and drought responses of members in Aegilops tauschii. Plant Mol. Biol. Rep. 2014. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Li, Z.Y.; Xu, Z.S.; He, G.Y.; Yang, G.X.; Chen, M.; Li, L.C.; Ma, Y.Z. Overexpression of soybean GmCBL1 enhances abiotic stress tolerance and promotes hypocotyl elongation in Arabidopsis. Biochem. Biophys. Res. Commun. 2012, 427, 731–736. [Google Scholar] [CrossRef]
- PLACE. Available online: http://www.dna.affrc.go.jp/PLACE/ (accessed on 12 September 2014).
- Xu, Z.S.; Ni, Z.Y.; Liu, L.; Nie, L.N.; Li, L.C.; Chen, M.; Ma, Y.Z. Characterization of the TaAIDFa gene encoding a CRT/DRE-binding factor responsive to drought, high-salt, and cold stress in wheat. Mol. Genet. Genomics 2008, 280, 497–508. [Google Scholar] [CrossRef]
- Xu, Z.S.; Xia, L.Q.; Chen, M.; Cheng, X.G.; Zhang, R.Y.; Li, L.C.; Zhao, Y.X.; Lu, Y.; Ni, Z.Y.; Liu, L.; et al. Isolation and molecular characterization of the Triticum aestivum L. ethylene-responsive factor 1 (TaERF1) that increases multiple stress tolerance. Plant Mol. Biol. 2007, 65, 719–732. [Google Scholar] [CrossRef]
- Strompen, G.; Dettmer, J.; Stierhof, Y.D.; Schumacher, K.; Jürgens, G.; Mayer, U. Arabidopsis vacuolar H+-ATPase subunit E isoform 1 is required for Golgi organization and vacuole function in embryogenesis. Plant J. 2005, 41, 125–132. [Google Scholar]
- Ratajczak, R. Structure, function and regulation of the plant vacuolar H+-translocating ATPase. Biochim. Biophys. Acta 2000, 1465, 17–36. [Google Scholar]
- Reuveni, M.; Bennet, A.B.; Bressan, R.A.; Hasegawa, P.M. Enhanced H+ transport capacity and ATP hydrolysis activity of the tonoplast H+-ATPase after NaCl adaptation. Plant Physiol. 1990, 94, 524–530. [Google Scholar] [CrossRef]
- Barkla, B.J.; Zingarelli, L.; Blumwald, E.; Smith, J.A.C. Tonoplast Na+/H+ antiport activity and its energization by the vacuolar H+-ATPase in the hallophytic plant Mesembryanthemum crystallinum L. Plant Physiol. 1995, 109, 549–556. [Google Scholar]
- Wang, B.S.; Rafael, R.; Zhang, J.H. Activity, amount, and subunit composition of vacuolar-type H+-ATPase and H+-PPase in wheat roots under severe NaCl stress. J. Plant Physiol. 2000, 157, 109–166. [Google Scholar] [CrossRef]
- Dietz, K.J.; Tavakoli, N.; Kluge, C.; Mimura, T.; Sharma, S.S.; Harris, G.C.; Chardonnens, A.N.; Golldack, D. Significance of the V-type ATPase for the adaptation to stressful growth conditions and its regulation on the molecular and biochemical level. J. Exp. Bot. 2001, 52, 1969–1980. [Google Scholar] [CrossRef]
- Binzel, M.L. NaCl-induced accumulation of tonoplast and plasma membrane H+-ATPase message in tomato. Physiol. Plant 1995, 94, 722–728. [Google Scholar] [CrossRef]
- Kasai, M.; Yamamoto, Y.; Maeshima, M.; Matsumoto, H. Effects of in vivo treatment with abscisic acid and/or cytokinin on activities of vacuolar H+ pumps of tonoplast-enriched membrane vesicles prepared from barley roots. Plant Cell Physiol. 1993, 34, 1107–1115. [Google Scholar]
- Batelli, G.; Verslues, P.E.; Agius, F.; Qiu, Q.; Fujii, H.; Pan, S.; Schumaker, K.S.; Grillo, S.; Zhu, J.K. SOS2 promotes salt tolerance in part by interacting with the vacuolar H+-ATPase and upregulating its transport activity. Mol. Cell. Biol. 2007, 27, 7781–7790. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.K. Salt stress signaling and mechanisms of plant salt tolerance. Genet. Eng. 2006, 27, 141–177. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Kłobus, G.; Janicka-Russak, M. Modulation by cytosolic components of proton pump activities in plasma membrane and tonoplast from Cucumis sativus roots during salt stress. Physiol. Plant 2004, 121, 84–92. [Google Scholar] [CrossRef]
- Yang, Y.Q.; Qin, Y.X.; Xie, C.G.; Zhao, F.Y.; Zhao, J.F.; Liu, D.F.; Chen, S.Y.; Fuglsang, A.T.; Palmgren, M.G.; Schumaker, K.S.; et al. The Arabidopsis chaperone J3 regulates the plasma membrane H+-ATPase through interaction with the PKS5 kinase. Plant Cell 2010, 22, 1313–1332. [Google Scholar] [CrossRef]
- Janicka-Russak, M.; Kłobus, G. Modification of plasma membrane and vacuolar H+-ATPases in response to NaCL and ABA. J. Plant Physiol. 2007, 164, 295–302. [Google Scholar] [CrossRef]
- ZhI, R.; TO, P. Function of transport H+-ATPases in plant cell plasma and vacuolar membranes of maize under salt stress conditions and effect of adaptogenic preparations. Ukrainskiĭ Biokhimicheskiĭ Zhurnal 2011, 83, 63–68. [Google Scholar]
- Ogawa, M.; Hanada, A.; Yamauchi, Y.; Kuwahara, A.; Kamiya, Y.; Yamaguchi, S. Gibberellin biosynthesis and response during Arabidopsis seed germination. Plant Cell 2003, 15, 1591–1604. [Google Scholar] [CrossRef]
- Razem, F.A.; Baron, K.; Hill, R.D. Turning on gibberellin and abscisic acid signaling. Curr. Opin. Plant Biol. 2006, 9, 454–459. [Google Scholar] [CrossRef]
- Toh, S.; Imamura, A.; Watanabe, A.; Nakabayashi, K.; Okamoto, M.; Jikumaru, Y.; Hanada, A.; Aso, Y.; Ishiyama, K.; Tamura, N.; et al. High temperatureinduced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds. Plant Physiol. 2008, 146, 1368–1385. [Google Scholar] [CrossRef]
- Pitzschke, A.; Hirt, H. Mitogen-activated protein kinases and reactive oxygen species signaling in plants. Plant Physiol. 2006, 141, 351–356. [Google Scholar] [CrossRef]
- Liu, Y.G.; Ye, N.H.; Liu, R.; Chen, M.X.; Zhang, J.H. H2O2 mediates the regulation of ABA catabolism and GA biosynthesis in Arabidopsis seed dormancy and germination. J. Exp. Bot. 2010, 61, 2979–2990. [Google Scholar] [CrossRef]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA induced NO generation and stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, X.-H.; Li, B.; Hu, Y.-G.; Chen, L.; Min, D.-H. The Wheat E Subunit of V-Type H+-ATPase Is Involved in the Plant Response to Osmotic Stress. Int. J. Mol. Sci. 2014, 15, 16196-16210. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150916196
Zhang X-H, Li B, Hu Y-G, Chen L, Min D-H. The Wheat E Subunit of V-Type H+-ATPase Is Involved in the Plant Response to Osmotic Stress. International Journal of Molecular Sciences. 2014; 15(9):16196-16210. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150916196
Chicago/Turabian StyleZhang, Xiao-Hong, Bo Li, Yin-Gang Hu, Liang Chen, and Dong-Hong Min. 2014. "The Wheat E Subunit of V-Type H+-ATPase Is Involved in the Plant Response to Osmotic Stress" International Journal of Molecular Sciences 15, no. 9: 16196-16210. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms150916196