Insights into Vibrio parahaemolyticus CHN25 Response to Artificial Gastric Fluid Stress by Transcriptomic Analysis

Abstract
:1. Introduction
2. Results and Discussion
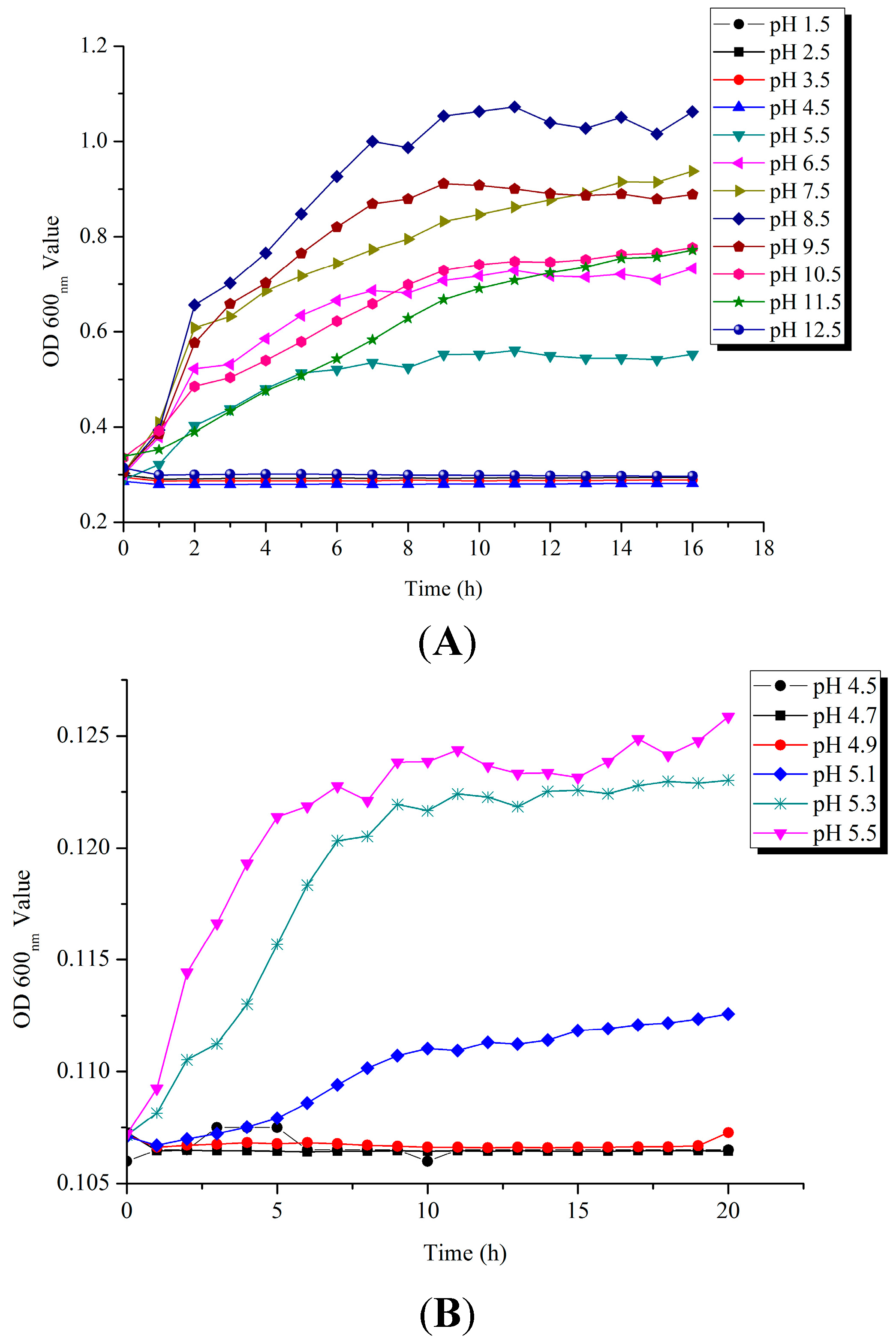
2.1. Survival of V. parahaemolyticus CHN25 under Acid pH Conditions

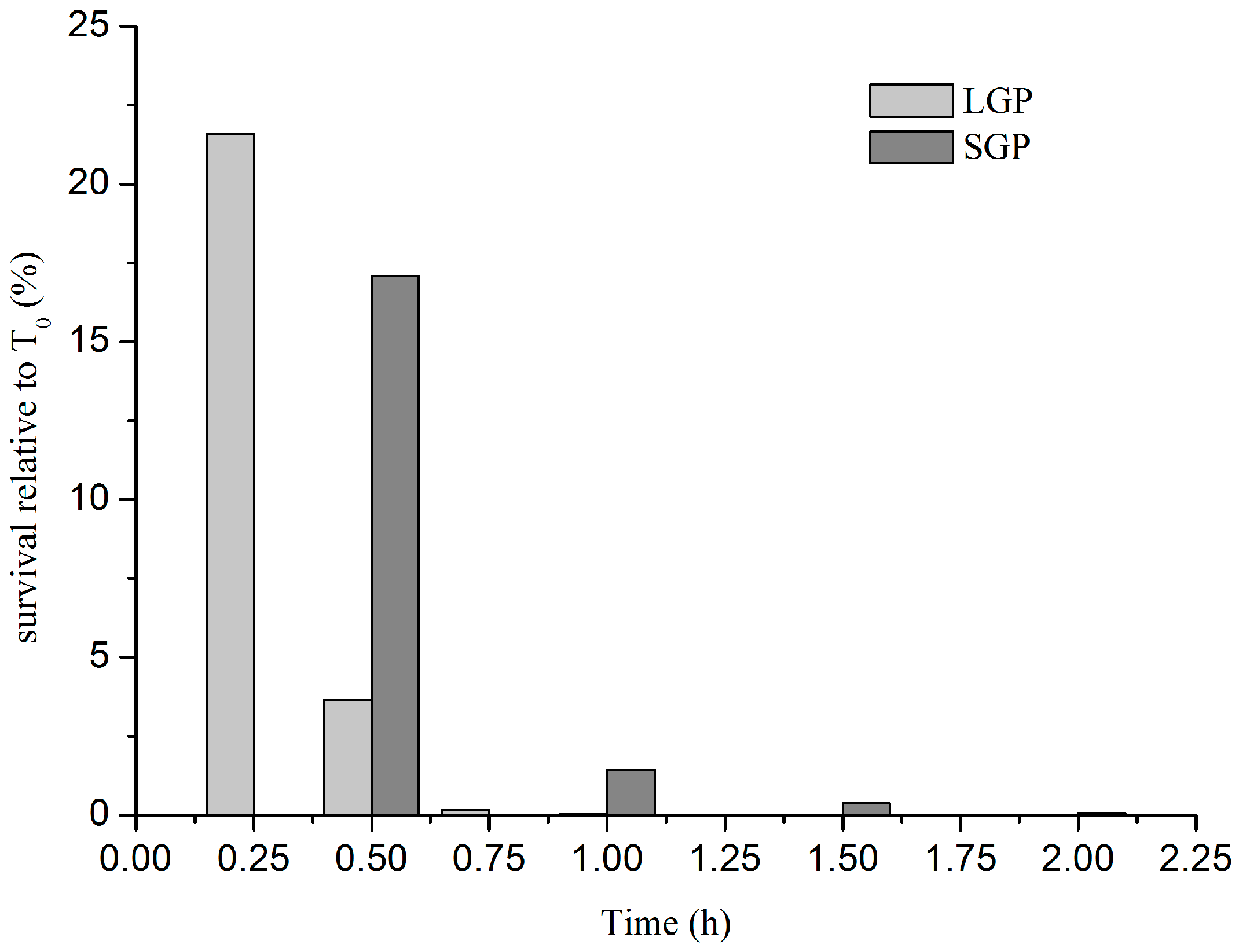
2.2. Tolerance of V. parahaemolyticus CHN25 at Logarithmic Growth Phase (LGP) and Stationary Growth Phase (SGP) to the AGF (Artificial Gastric Fluid) Stress

2.3. Transcriptome Profiles of V. parahaemolyticus CHN25 in the Response to the AGF Stress
2.4. Major Metabolic Pathways Involved in the Response of V. parahaemolyticus CHN25 to the AGF Stress
2.4.1. Major Metabolic Pathways Involved in V. parahaemolyticus CHN25 Cells at LGP in the AGF Stress

{kind=link}
{kind=link}
| Metabolic Pathway/Cellular Function | Locus/Gene in V. parahaemolyticus | Fold Change | Description of Encoded Protein | |
|---|---|---|---|---|
| RIMD2210633 | CHN25 | |||
| LGP cells | ||||
| Phosphotransferase system (PTS) | VPA1667 | Chn25A_1555 | 0.0384 | Glucose-specific IIBC component |
| VPA1424 | Chn25A_1312 | 27.9696 | Fructose-specific IIABC component | |
| VPA1422 | Chn25A_1310 | 2.2688 | Nitrogen regulatory IIA component | |
| VPA1421 | Chn25A_1309 | 2.3425 | Fructose-specific IIB component | |
| VPA1420 | Chn25A_1308 | 2.8724 | Fructose-specific IIABC component | |
| VPA0501 | Chn25A_1196 | 2.6299 | Mannitol-specific enzyme II component | |
| VPA0500 | Chn25A_1197 | 2.6362 | Mannitol-specific enzyme II component | |
| VPA0297 | Chn25A_0302 | 8.029 | fructose-specific IIBC component | |
| VPA0298 | Chn25A_0303 | 2.8846 | Fructose-specific IIA component | |
| VPA0231 | Chn25A_0232 | 2.0079 | Phosphotransferase enzyme II, A component | |
| VPA0230 | Chn25A_0231 | 5.501 | Putative sugar phosphotransferase component II B | |
| VP2637 | Chn25_2566 | 2.2518 | Cellobiose-specific IIB component | |
| VP2636 | Chn25_2565 | 2.6756 | Cellobiose-specific IIC component | |
| VP2046 | Chn25_1932 | 2.6196 | Glucose-specific IIBC components | |
| VP0831 | Chn25_0826 | 4.8477 | N-acetylglucosamine-specific IIABC component | |
| VP0370 | Chn25_0356 | 2.7113 | Mannitol-specific IIABC component | |
| VPA0229 (ulaA) | Chn25A_0230 | 6.4614 | Ascorbate-specific enzyme IIC | |
| VP0366 | Chn25_0352 | 3.2996 | Putative PTS, enzyme I | |
| Galactose metabolism | VPA0879 | Chn25A_0828 | 0.3848 | UDP-glucose 4-epimerase |
| VP2400 | Chn25_2264 | 4.517 | UDP-glucose 4-epimerase | |
| VP2400 | Chn25A_1071 | 3.5838 | UDP-glucose 4-epimerase | |
| VP2399 | Chn25A_1070 | 4.4699 | Galactose-1-phosphate uridylyltransferase | |
| VP2398 | Chn25_2262 | 3.8991 | Galactokinase | |
| VP2398 | Chn25A_1069 | 2.497 | Galactokinase | |
| VP2077 | Chn25_1963 | 2.0087 | Maltodextrin glucosidase | |
| Galactose metabolism | VP0839 | Chn25_0834 | 2.4836 | Phosphoglucomutase |
| VP2403 (ebgA) | Chn25_2266 | 2.4115 | Cryptic beta-d-galactosidase subunit alpha | |
| VP2404 (ebgC) | Chn25_2267 | 2.9965 | Cryptic beta-d-galactosidase subunit beta | |
| VP2397 (galM) | Chn25_2261 | 2.1978 | Aldose 1-epimerase | |
| VP2855 (pfkA) | Chn25_2776 | 0.3195 | 6-phosphofructokinase | |
| Ribosome biosynthesis | VP2772 | Chn25_2698 | 3.0673 | 30S ribosomal protein S7 |
| VP1210 | Chn25_1217 | 2.6329 | 50S ribosomal protein L25 | |
| VP2925 (rplA) | Chn25_2832 | 2.3329 | 50S ribosomal protein L1 | |
| VP2926 (rplK) | Chn25_2833 | 2.1866 | 50S ribosomal protein L11 | |
| VP2923 (rplL) | Chn25_2830 | 2.7378 | 50S ribosomal protein L7/L12 | |
| VP0264 (rplP) | Chn25_0255 | 2.2242 | 50S ribosomal protein L16 | |
| VP1282 (rplT) | Chn25_1289 | 6.5113 | 50S ribosomal protein L20 | |
| VP0262 (rplV) | Chn25_0253 | 2.5181 | 50S ribosomal protein L22 | |
| VP0259 (rplW) | Chn25_0250 | 2.2719 | 50S ribosomal protein L23 | |
| VP0329 (rpmA) | Chn25_0317 | 5.5738 | 50S ribosomal protein L27 | |
| VP0185 (rpmB) | Chn25_0181 | 2.7392 | 50S ribosomal protein L28 | |
| VP0265 (rpmC) | Chn25_0256 | 2.5323 | 50S ribosomal protein L29 | |
| VP0255 (rpmE) | Chn25_0246 | 5.0193 | 50S ribosomal protein L31 | |
| VP0186 (rpmG) | Chn25_0182 | 7.2192 | 50S ribosomal protein L33 | |
| VP2030 (rpsA) | Chn25_1918 | 3.9866 | 30S ribosomal protein S1 | |
| VP0263 (rpsC) | Chn25_0254 | 2.6074 | 30S ribosomal protein S3 | |
| VP2740 (rpsF) | Chn25_2668 | 2.818 | 30S ribosomal protein S6 | |
| VP0439 (rpsI) | Chn25_0398 | 2.1455 | 30S ribosomal protein S9 | |
| VP2453 (rpsO) | Chn25_2391 | 3.7221 | 30S ribosomal protein S15 | |
| VP0266 | Chn25_0257 | 3.3225 | Ribosomal protein S17 | |
| VP0531 (rpsT) | Chn25_0481 | 6.127 | 30S ribosomal protein S20 | |
| Fructose and mannose metabolism | VPA1425 | Chn25A_1313 | 23.9303 | Mannose-6-phosphate isomerase |
| VP2599 | Chn25_2528 | 0.2922 | Fructose-bisphosphate aldolase | |
| VP2488 | Chn25_2426 | 4.6724 | Putative phosphoglucomutase/phosphomannomutase | |
| Amino sugar and nucleotide sugar metabolism | VP0543 | Chn25_0493 | 2.2962 | N-acetylmuramic acid-6-phosphate etherase |
| VPA0833 (glgC) | Chn25A_0780 | 0.4431 | Glucose-1-phosphate adenylyltransferase | |
| VP1023 (glgC) | Chn25_1047 | 2.336 | Glucose-1-phosphate adenylyltransferase | |
| VP0829 (nagA) | Chn25_0825 | 2.027 | N-acetylglucosamine-6-phosphate deacetylase | |
| VPA0038 (nagB) | Chn25A_0033 | 2.5647 | Glucosamine-6-phosphate deaminase | |
| Glycolysis/gluconeogenesis | VPA0566 | Chn25A_1136 | 5.8196 | Alcohol dehydrogenase |
| VPA0180 | Chn25A_0182 | 2.2002 | Phospho-beta-glucosidase B | |
| VP2157 | Chn25_2026 | 4.6439 | Glyceraldehyde-3-phosphate dehydrogenase | |
| Aminoacyl-tRNA biosynthesis | VP2470 | Chn25_2407 | 2.0793 | Tyrosyl-tRNA synthetase |
| VP1280 | Chn25_1286 | 0.4891 | Threonyl-tRNA synthetase | |
| VP2548 (alaS) | Chn25_2482 | 2.6177 | Alanyl-tRNA synthetase | |
| VP0861 (argS) | Chn25_0854 | 2.7778 | Arginyl-tRNA synthetase | |
| VP1150 (cysS) | Chn25_1159 | 2.0297 | Cysteinyl-tRNA synthetase | |
| VP0021 (glyS) | Chn25_0010 | 3.0031 | Glycyl-tRNA synthetase subunit beta | |
| VP0534 (ileS) | Chn25_0484 | 2.3183 | Isoleucyl-tRNA synthetase | |
| VP0727 (leuS) | Chn25_0685 | 2.1816 | Leucyl-tRNA synthetase | |
| VP2069 (metG) | Chn25_1955 | 2.2087 | Methionyl-tRNA synthetase | |
| VP1291 (pheT) | Chn25_1298 | 5.4809 | Phenylalanyl-tRNA synthetase subunit beta | |
| VP2646 (valS) | Chn25_2575 | 2.077 | Valyl-tRNA synthetase | |
| Pyruvate metabolism | VPA1567 | Chn25A_1456 | 2.1699 | Putative pyruvate formate lyase |
| VPA1123 | Chn25A_0560 | 0.4528 | Putative acyl-CoA thiolase | |
| VPA0823 | Chn25A_0771 | 2.0537 | Pyruvate kinase | |
| VPA0646 | Chn25A_1010 | 0.3383 | Putative pyruvate dehydrogenase E1 component, beta subunit | |
| Pyruvate metabolism | VPA0620 | Chn25A_1034 | 0.4695 | Putative acyl-CoA thiolase |
| VPA0611 | Chn25A_1091 | 0.3706 | Acetate kinase | |
| VPA0372 | Chn25A_0367 | 3.9537 | Phosphoenolpyruvate synthase | |
| VPA0144 | Chn25A_0145 | 0.4364 | d-lactate dehydrogenase | |
| VP2881 | Chn25_2801 | 3.0293 | Acetyl-CoA carboxylase, biotin carboxylase subunit | |
| VP2878 | Chn25_2798 | 2.514 | Acetyl-CoA synthetase | |
| VP2545 | Chn25_2479 | 2.1607 | Oxaloacetate decarboxylase subunit gamma | |
| VP2517 | Chn25_2451 | 3.4256 | Dihydrolipoamide dehydrogenase | |
| VP2039 | Chn25_1927 | 2.6156 | Pyruvate kinase II | |
| VP1627 | Chn25_1620 | 3.6175 | Acylphosphatase | |
| VP1258 | Chn25_1264 | 2.6731 | Malate dehydrogenase | |
| VP0325 | Chn25_0313 | 3.5504 | Malate dehydrogenase | |
| VP2519 (aceE) | Chn25_2453 | 2.132 | Pyruvate dehydrogenase subunit E1 | |
| VPA1499 (lldD) | Chn25A_1389 | 8.5267 | l-lactate dehydrogenase | |
| Oxidative phosphorylation | VPA0631 | Chn25A_1023 | 0.464 | Putative protoheme IX farnesyltransferase |
| VPA0544 | Chn25A_1156 | 0.4221 | Protoheme IX farnesyltransferase | |
| VP2841 | Chn25_2763 | 2.102 | Fumarate reductase iron-sulfur subunit | |
| VPA0539 | Chn25A_1161 | 0.2437 | Cytochrome c oxidase, subunit III | |
| VP1543 | Chn25_1521 | 2.2303 | Cytochrome c oxidase, subunit CcoO | |
| VP1541 | Chn25_1519 | 2.2683 | Cytochrome c oxidase, subunit CcoP | |
| VP1054 | Chn25_1074 | 2.0957 | Cytochrome d ubiquinol oxidase, subunit II | |
| VP1053 | Chn25_1073 | 2.3358 | Cytochrome d ubiquinol oxidase, subunit I | |
| VPA0628 | Chn25A_1026 | 2.0087 | Cytochrome o ubiquinol oxidase, subunit I | |
| VP1165 | Chn25_1174 | 2.4205 | Putative manganese-dependent inorganic pyrophosphatase | |
| VP0443 | Chn25_0401 | 2.4995 | Ubiquinol-cytochrome c reductase, cytochrome c1 | |
| VP0442 | Chn25_0400 | 2.0404 | Ubiquinol-cytochrome c reductase, cytochrome b | |
| Oxidative phosphorylation | VP3076 | Chn25_2976 | 0.4698 | F0F1 ATP synthase subunit I |
| VP3068 (atpC) | Chn25_2968 | 3.0803 | F0F1 ATP synthase subunit epsilon | |
| VP0844 | Chn25_0839 | 2.0288 | Succinate dehydrogenase, hydrophobic membrane anchor protein | |
| VP0843 (sdhC) | Chn25_0838 | 2.1718 | Succinate dehydrogenase cytochrome b556 large membrane subunit | |
| Ciliary or bacterial-type flagellar motility | VP0772 (flgA) | Chn25_0767 | 0.2303 | Flagellar basal body P-ring biosynthesis protein FlgA |
| VP2235 (flhA) | Chn25_2102 | 0.4555 | Flagellar biosynthesis protein FlhA | |
| VP2236 (flhB) | Chn25_2103 | 0.463 | Flagellar biosynthesis protein FlhB | |
| VP2255 | Chn25_2122 | 0.436 | Polar flagellar rod protein FlaI | |
| VP2256 (fliD) | Chn25_2123 | 0.4776 | Flagellar capping protein | |
| VP2257 | Chn25_2124 | 2.0357 | Flagellar protein FlaG | |
| VP2261 | Chn25_2127 | 0.2235 | Flagellin | |
| VPA0263 | Chn25A_0264 | 0.3335 | Flagellar basal body P-ring biosynthesis protein | |
| Polyamine transport | VP1332 | Chn25_1334 | 0.4055 | Binding protein component of ABC transporter |
| VP1336 | Chn25_1337 | 0.3839 | ABC transporter ATP-binding protein | |
| VP1337 | Chn25_1338 | 0.419 | Putative permease of ABC transporter | |
| VP1338 | Chn25_1339 | 0.371 | ABC transporter permease | |
| d-ribose transport | VPA1087 | Chn25A_0593 | 12.5376 | d-ribose pyranase |
| VPA1086 | Chn25A_0594 | 14.963 | d-ribose transporter ATP binding protein | |
| VPA1086 (rbsC) | Chn25A_0595 | 8.678 | Ribose ABC transporter permease protein | |
| VPA1084 | Chn25A_0596 | 7.7049 | d-ribose transporter subunit RbsB | |
| Maltose transport | VPA1399 (malG) | Chn25A_1076 | 2.0352 | Maltose transporter permease |
| VPA1400 (malF) | Chn25A_1077 | 2.8009 | Maltose transporter membrane protein | |
| VPA1401(malE) | Chn25A_1078 | 3.0226 | Maltose ABC transporter periplasmic protein | |
| VPA1402 | Chn25A_1079 | 2.561 | Maltose/maltodextrin transporter ATP-binding protein | |
| VPA1644 (lamB) | Chn25A_1532 | 3.35 | Maltoporin | |
| SGP cells | ||||
| Pyrimidine and purine metabolism | VPA1243 | Chn25A_0442 | 0.491 | Cytosine deaminase |
| VP0524 (thyA) | Chn25_0476 | 2.1909 | Thymidylate synthase | |
| VP1760 | Chn25_1387 | 2.1851 | Putative adenylate cyclase | |
| VPA1159 | Chn25A_0527 | 0.4775 | Guanosine 5'-monophosphate oxidoreductase | |
| VPA0855 | Chn25A_0801 | 0.3912 | Putative 5'-nucleotidase | |
| VPA0074 | Chn25A_0069 | 2.3652 | Putative DNA polymerase III, epsilon subunit | |
| VP2303 (dnaE) | Chn25_2169 | 0.4586 | DNA polymerase III subunit alpha | |
| Iron ion transport | VP2491 | Chn25_2429 | 0.4159 | Iron (III) ABC transporter, periplasmic Iron-compound-binding protein |
| VPA0310 | Chn25A_0316 | 2.1424 | Hypothetical protein | |
| PTS | VP2674 | Chn25_2602 | 2.2237 | Phosphocarrier protein NPr |
| VPA0297 | Chn25A_0302 | 0.4786 | PTS system, fructose-specific IIBC component | |
| VPA0298 | Chn25A_0303 | 0.4004 | PTS system, fructose-specific IIA component | |
| VPA1424 | Chn25A_1312 | 0.3946 | PTS system, fructose-specific IIABC component | |
| Quaternary ammonium group transport | VPA1111 | Chn25A_0571 | 0.453 | Putative glycine betaine-binding ABC transporter |
| Aromatic compound catabolic process | VP0240 | Chn25_0231 | 2.2949 | Putative 5-carboxymethyl-2-hydroxymuconate delta isomerase |
| Glycine betaine biosynthetic process from choline | VPA1114 | Chn25A_0568 | 0.4435 | Transcriptional regulator BetI |
| VPA1112 | Chn25A_0570 | 0.3776 | Choline dehydrogenase | |
| Pilus | VPA0725 | Chn25A_0670 | 3.0408 | Putative TadB |
| ATP binding | VPA0380 | Chn25A_0375 | 0.481 | Hypothetical protein |
| VPA1302 | Chn25A_0400 | 0.2424 | Hypothetical protein | |
| Outer membrane-bounded periplasmic space and nitrite reductase activity | VP1928 | Chn25_1816 | 2.0205 | Cytochrome c nitrite reductase pentaheme subunit |
| Cytolysis | VP3048 | Chn25_2947 | 2.0825 | Putative hemolysin III |
| Betaine-aldehyde dehydrogenase activity | VPA1113 | Chn25A_0569 | 0.3189 | Betaine aldehyde dehydrogenase |
| Phosphoglycerate transport | VPA0825 | Chn25A_0773 | 0.3614 | Putative phosphoglycerate transport regulatory protein PgtC |
| NADPH dehydrogenase activity | VPA0465 | Chn25A_1233 | 2.0487 | Putative NAD(P)H oxidoreductase |
| Regulation of DNA repair | VP2945 | Chn25_2852 | 2.205 | LexA repressor |
| 3-isopropylmalate dehydratase complex and 3-isopropylmalate dehydratase activity | VP0343 | Chn25_0331 | 2.0142 | Isopropylmalate isomerase large subunit |
| Triglyceride lipase activity | VP1181 | Chn25_1190 | 2.2305 | lactonizing lipase |
| Lactoylglutathione lyase activity | VP2166 | Chn25_2034 | 0.3883 | Putative lactoylglutathione lyase |
| Anaerobic electron transport chain and nitrogen compound metabolic process | VP1928 | Chn25_1816 | 2.0205 | Cytochrome c nitrite reductase pentaheme subunit |
| Alkaline phosphatase activity | VP2163 | Chn25_2032 | 0.4035 | Alkaline phosphatase |
| Zinc ion transmembrane transporter activity | VPA1287 | Chn25A_0416 | 0.2585 | Putative transporter |
| Transmembrane transport | VP1359 | Chn25_1745 | 2.0727 | Hypothetical protein |
2.4.2. Major Metabolic Pathways Involved in V. parahaemolyticus CHN25 Cells at SGP in the AGF Stress
2.5. Other Altered Biological Functions in V. parahaemolyticus CHN25 in the Response to the AGF Stress
2.6. Regulators Involved in the Response of V. parahaemolyticus CHN25 to the AGF Stress
2.7. Possible Acid Stress Mechanisms in V. parahaemolyticus CHN25
3. Experimental Section
3.1. Bacterial Growth Conditions
3.2. AGF Survival Assay
3.3. RNA Extraction and Microarray Analysis
3.4. qRT-PCR Analysis
3.5. Microarray Data Accession Number
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ceccarelli, D.; Hasan, N.A.; Huq, A.; Colwell, R.R. Distribution and dynamics of epidemic and pandemic Vibrio parahaemolyticus virulence factors. Front. Cell. Infect. Microbiol. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Fujino, T.; Okuno, Y.; Nakada, D.; Aoyama, A.; Fukai, K.; Mukai, T.; Ueho, T. On the bacteriological examination of Shirasu food poisoning. Med. J. Osaka Univ. 1953, 4, 299–304. [Google Scholar]
- Okuda, J.; Ishibashi, M.; Hayakawa, E.; Nishino, T.; Takeda, Y.; Mukhopadhyay, A.K.; Garg, S.; Bhattacharya, S.; Nair, G.B.; Nishibuchi, M. Emergence of a unique O3:K6 clone of Vibrio parahaemolyticus in Calcutta, India, and isolation of strains from the same clonal group from Southeast Asian travelers arriving in Japan. J. Clin. Microbiol. 1997, 35, 3150–3155. [Google Scholar] [PubMed]
- Velazquez-Roman, J.; León-Sicairos, N.; de Jesus Hernández-Díaz, L.; Canizalez-Roman, A. Pandemic Vibrio parahaemolyticus O3:K6 on the American continent. Front. Cell. Infect. Microbiol. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Twedt, R.M.; Spaulding, P.L.; Hall, H.E. Morphological, cultural, biochemical, and serological comparison of Japanese strains of Vibrio parahemolyticus with related cultures isolated in the United States. J. Bacteriol. 1969, 98, 511–518. [Google Scholar] [PubMed]
- Joseph, S.W.; Colwell, R.R.; Kaper, J.B. Vibrio parahaemolyticus and related halophilic Vibrios. Crit. Rev. Microbiol. 1982, 10, 77–124. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.L. Gastric exocrine and endocrine secretion. Curr. Opin. Gastroenterol. 2009, 25, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Russell, T.L.; Berardi, R.R.; Barnett, J.L.; Dermentzoglou, L.C.; Jarvenpaa, K.M.; Schmaltz, S.P.; Dressman, J.B. Upper gastrointestinal pH in seventy-nine healthy, elderly, North American men and women. Pharm. Res. 1993, 10, 187–196. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.; Boyd, A. Manipulation of intestinal epithelial cell function by the cell contact-dependent type III secretion systems of Vibrio parahaemolyticus. Front. Cell. Infect. Microbiol. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Lund, P.; Tramonti, A.; Biase, D. Coping with low pH: Molecular strategies in neutralophilic bacteria. FEMS Microbiol. Rev. 2014, 38. [Google Scholar] [CrossRef]
- Merrell, D.S.; Camilli, A. Regulation of Vibrio cholerae genes required for acid tolerance by a member of the “ToxR-like” family of transcriptional regulators. J. Bacteriol. 2000, 182, 5342–5350. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.E.; Jeong, H.G.; Lee, J.H.; Choi, S.H. AphB influences acid tolerance of Vibrio vulnificus by activating expression of the positive regulator CadC. J. Bacteriol. 2006, 188, 6490–6497. [Google Scholar] [CrossRef] [PubMed]
- Kalburge, S.S.; Whitaker, W.B.; Boyd, E.F. High-salt preadaptation of Vibrio parahaemolyticus enhances survival in response to lethal environmental stresses. J. Food Prot. 2014, 77, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, W.B.; Parent, M.A.; Boyd, A.; Richards, G.P.; Boyd, E.F. The Vibrio parahaemolyticus ToxRS regulator is required for stress tolerance and colonization in a novel orogastric streptomycin-induced adult murine model. Infect Immun. 2012, 80, 1834–1845. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Yu, P.; Li, B.; Pan, Y.; Zhang, X.; Cong, J.; Zhao, Y.; Wang, H.; Chen, L. The mosaic accessory gene structures of the SXT/R391-like integrative and conjugative elements derived from Vibrio spp. isolated from aquatic products and environment in the Yangtze River estuary, China. BMC Microbiol. 2013, 13. [Google Scholar] [CrossRef]
- Martín-Galiano, A.J.; Overweg, K.; Ferrándiz, M.J.; Reuter, M.; Wells, J.M.; Adela, G. Transcriptional analysis of the acid tolerance response in Streptococcus pneumoniae. Microbiology 2005, 151, 3935–3946. [Google Scholar] [CrossRef] [PubMed]
- Bore, E.; Langsrud, S.; Langsrud, Ø.; Rode, T.M.; Holck, A. Acid-shock responses in Staphylococcus aureus investigated by global gene expression analysis. Microbiology 2007, 153, 2289–2303. [Google Scholar] [CrossRef]
- Hellweg, C.; Pühler, A.; Weidner, S. The time course of the transcriptome response of Sinorhizobium meliloti 1021 following a shift to acidic pH. BMC Microbiol. 2009, 9. [Google Scholar] [CrossRef]
- Ivy, R.A.; Wiedmann, M.; Boor, K.J. Listeria monocytogenes grown at 7 °C shows reduced acid survival and an altered transcriptional response to acid shock compared to L. monocytogenes grown at 37 °C. Appl. Environ. Microbiol. 2012, 78, 3824–3836. [Google Scholar] [CrossRef] [PubMed]
- Wall, T.; Båth, K.; Britton, R.A.; Jonsson, H.; Versalovic, J.; Roos, S. The early response to acid shock in Lactobacillus reuteri involves the ClpL chaperone and a putative cell wall-altering esterase. Appl. Environ. Microbiol. 2007, 73, 3924–3935. [Google Scholar] [CrossRef] [PubMed]
- Postma, P.; Lengeler, J.; Jacobson, G. Phosphoenolpyruvate: Carbohydrate phosphotransferase systems of bacteria. Microbiol. Rev. 1993, 57, 543–594. [Google Scholar] [PubMed]
- Bickle, T.A.; Howard, G.A.; Traut, R.R. Ribosome heterogeneity the nonuniform distribution of specific ribosomal proteins among different functional classes of ribosomes. J. Biol. Chem. 1973, 248, 4862–4864. [Google Scholar] [PubMed]
- Moll, I.; Engelberg-Kulka, H. Selective translation during stress in Escherichia coli. Trends Biochem. Sci. 2012, 37, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Okuno, D.; Iino, R.; Noji, H. Rotation and structure of FoF1-ATP synthase. J. Biochem. 2011, 149, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, Z.; Zhang, H.; Wang, J.; Liu, Y.; Chen, G. Rapid detection of several foodborne pathogens by F0F1-ATPase molecular motor biosensor. J. Microbiol. Methods 2013, 93, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Oldham, M.L.; Chen, S.; Chen, J. Structural basis for substrate specificity in the Escherichia coli maltose transport system. Proc. Natl. Acad. Sci. USA 2013, 110, 18132–18137. [Google Scholar] [CrossRef] [PubMed]
- Bron, P.A.; Grangette, C.; Mercenier, A.; de Vos, W.M.; Kleerebezem, M. Identification of Lactobacillus plantarum genes that are induced in the gastrointestinal tract of mice. J. Bacteriol. 2004, 186, 5721–5729. [Google Scholar] [CrossRef]
- De Jonge, R.; Ritmeester, W.S.; van Leusden, F.M. Adaptive responses of Salmonella enterica serovar Typhimurium DT104 and other S. Typhimurium strains and Escherichia coli O157 to low pH environments. J. Appl. Microbiol. 2003, 94, 625–632. [Google Scholar] [CrossRef]
- Hickey, E.W.; Hirshfield, I.N. Low-pH-induced effects on patterns of protein synthesis and on internal pH in Escherichia coli and Salmonella typhimurium. Appl. Environ. Microbiol. 1990, 56, 1038–1045. [Google Scholar] [PubMed]
- Maurer, L.M.; Yohannes, E.; Bondurant, S.S.; Radmacher, M.; Slonczewski, J.L. pH regulates genes for flagellar motility, catabolism, and oxidative stress in Escherichia coli K-12. J. Bacteriol. 2005, 187, 304–319. [Google Scholar] [CrossRef] [PubMed]
- Tomich, M.; Fine, D.H.; Figurski, D.H. The TadV protein of Actinobacillus actinomycetemcomitans is a novel aspartic acid prepilin peptidase required for maturation of the Flp1 pilin and TadE and TadF pseudopilins. J. Bacteriol. 2006, 188, 6899–6914. [Google Scholar] [CrossRef] [PubMed]
- Swint-Kruse, L.; Matthews, K.S. Allostery in the LacI/GalR family: Variations on a theme. Curr. Opin. Microbiol. 2009, 12, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Rkenes, T.; Lamark, T.; Strøm, A.R. DNA-binding properties of the BetI repressor protein of Escherichia coli: The inducer choline stimulates BetI-DNA complex formation. J. Bacteriol. 1996, 178, 1663–1670. [Google Scholar] [PubMed]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant. Cell Environ. 2002, 25, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Laub, M.T.; Goulian, M. Specificity in two-component signal transduction pathways. Annu. Rev. Genet. 2007, 41, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, H.; Xie, J. Regulatory and pathogenesis roles of Mycobacterium Lrp/AsnC family transcriptional factors. J. Cell. Biochem. 2011, 112, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Maddocks, S.E.; Oyston, P.C. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef] [PubMed]
- Gallegos, M.T.; Schleif, R.; Bairoch, A.; Hofmann, K.; Ramos, J.L. Arac/XylS family of transcriptional regulators. Microbiol. Mol. Biol. Rev. 1997, 61, 393–410. [Google Scholar] [PubMed]
- Ramos, J.L.; Martínez-Bueno, M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [PubMed]
- Bakhlanova, I.; Dudkina, A.; Baĭtin, D. Enzymatic control of homologous recombination in Escherichia coli cells and hyper-recombination. Mol. Biol. 2012, 47, 205–217. [Google Scholar]
- Brinkrolf, K.; Brune, I.; Tauch, A. The transcriptional regulatory network of the amino acid producer Corynebacterium glutamicum. J. Biotechnol. 2007, 129, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Justino, G.C.; Santos, M.R.; Canário, S.; Borges, C.; Florêncio, M.H.; Mira, L. Plasma quercetin metabolites: Structure-antioxidant activity relationships. Arch. Biochem. Biophys. 2004, 432, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Leyn, S.A.; Li, X.; Zheng, Q.; Novichkov, P.S.; Reed, S.; Romine, M.F.; Fredrickson, J.K.; Yang, C.; Osterman, A.L.; Rodionov, D.A. Control of proteobacterial central carbon metabolism by the HexR transcriptional regulator a case study in shewanella oneidensis. J. Biol. Chem. 2011, 286, 35782–35794. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, J. Role and regulation of bacterial LuxR-like regulators. J. Cell. Biochem. 2011, 112, 2694–2702. [Google Scholar] [CrossRef] [PubMed]
- Brzostek, K.; Skorek, K.; Raczkowska, A. OmpR, a central integrator of several cellular responses in Yersinia enterocolitica. In Advances in Yersinia Research; Springer-Verlag: New York, NY, USA, 2012; pp. 325–334. [Google Scholar]
- Padan, E.; Schuldiner, S. Molecular physiology of the Na+/H+ antiporter in Escherichia coli. J. Exp. Biol. 1994, 196, 443–456. [Google Scholar] [PubMed]
- Merrell, D.S.; Camilli, A. Acid tolerance of gastrointestinal pathogens. Curr. Opin. Microbiol. 2002, 5, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Kovacikova, G.; Lin, W.; Skorupski, K. The LysR-type virulence activator AphB regulates the expression of genes in Vibrio cholerae in response to low pH and anaerobiosis. J. Bacteriol. 2010, 192, 4181–4191. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Kimura, B.; Takahashi, H.; Watanabe, T.; Obata, H.; Kai, A.; Morozumi, S.; Fujii, T. Lysine decarboxylase of Vibrio parahaemolyticus: Kinetics of transcription and role in acid resistance. J. Appl. Microbiol. 2008, 104, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, W.B.; Parent, M.A.; Naughton, L.M.; Richards, G.P.; Blumerman, S.L.; Boyd, E.F. Modulation of responses of Vibrio parahaemolyticus O3:K6 to pH and temperature stresses by growth at different salt concentrations. Appl. Environ. Microbiol. 2010, 76, 4720–4729. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Press: New York, NY, USA, 2001. [Google Scholar]
- Tessema, G.T.; Moretro, T.; Snipen, L.; Heir, E.; Holck, A.; Naterstad, K.; Axelsson, L. Microarray-based transcriptome of Listeria monocytogenes adapted to sublethal concentrations of acetic acid, lactic acid, and hydrochloric acid. Can. J. Microbiol. 2012, 58, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Coutard, F.; Lozach, S.; Pommepuy, M.; Hervio-Heath, D. Real-time reverse transcription-PCR for transcriptional expression analysis of virulence and housekeeping genes in viable but nonculturable Vibrio parahaemolyticus after recovery of culture ability. Appl. Environ. Microbiol. 2007, 73, 5183–5189. [Google Scholar] [CrossRef] [PubMed]
- Funahashi, T.; Moriya, K.; Uemura, S.; Miyoshi, S.I.; Shinoda, S.; Narimatsu, S.; Yamamoto, S. Identification and characterization of pvuA, a gene encoding the ferric vibrioferrin receptor protein in Vibrio parahaemolyticus. J. Bacteriol. 2002, 184, 936–946. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Liu, T.; Peng, X.; Chen, L. Insights into Vibrio parahaemolyticus CHN25 Response to Artificial Gastric Fluid Stress by Transcriptomic Analysis. Int. J. Mol. Sci. 2014, 15, 22539-22562. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151222539
Sun X, Liu T, Peng X, Chen L. Insights into Vibrio parahaemolyticus CHN25 Response to Artificial Gastric Fluid Stress by Transcriptomic Analysis. International Journal of Molecular Sciences. 2014; 15(12):22539-22562. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151222539
Chicago/Turabian StyleSun, Xuejiao, Taigang Liu, Xu Peng, and Lanming Chen. 2014. "Insights into Vibrio parahaemolyticus CHN25 Response to Artificial Gastric Fluid Stress by Transcriptomic Analysis" International Journal of Molecular Sciences 15, no. 12: 22539-22562. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms151222539