
Transcriptome Analysis in Rat Kidneys: Importance of Genes Involved in Programmed Hypertension



Abstract
:1. Introduction
2. Results
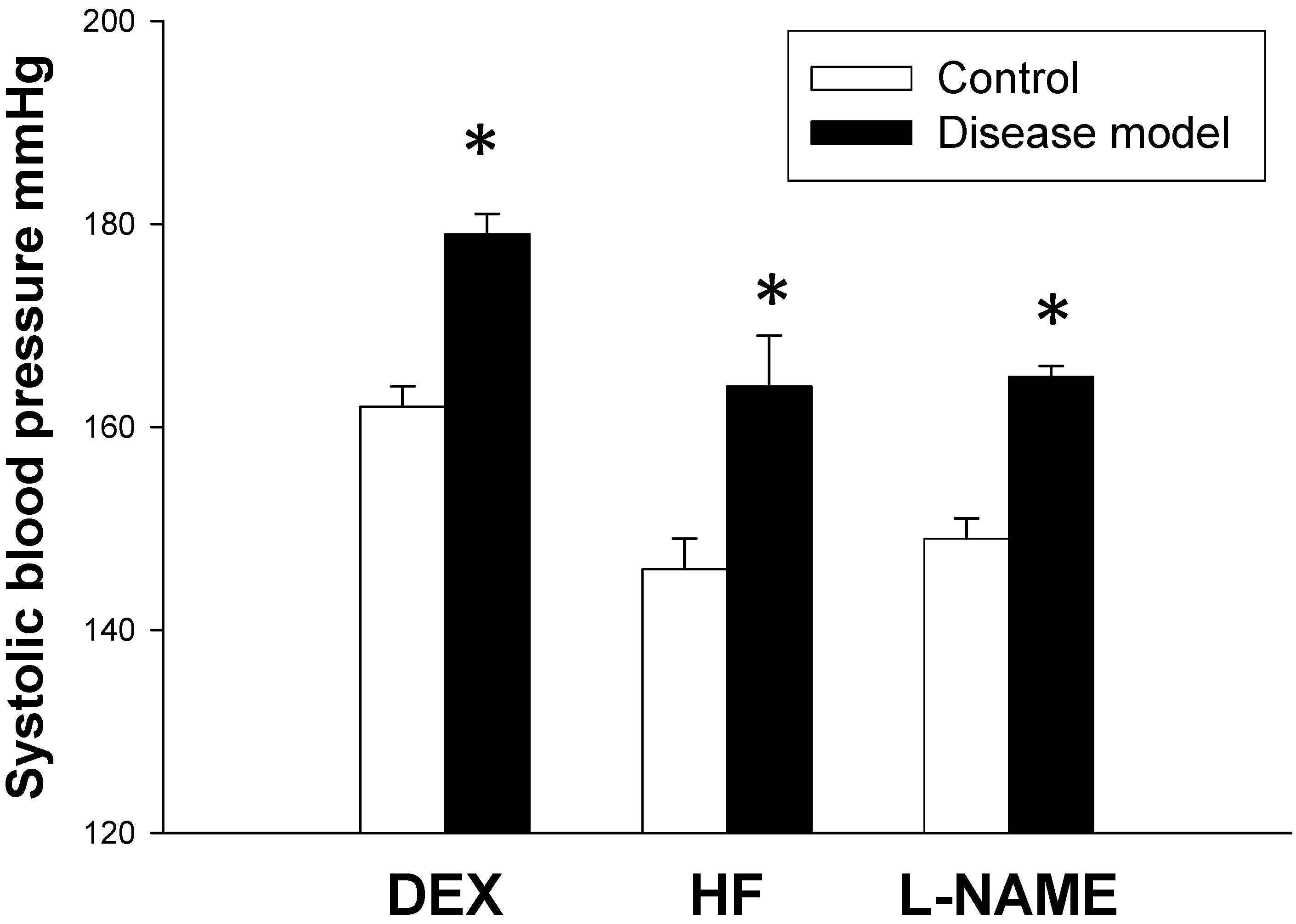
2.1. Hypertension Is a Common Phenotype in Response to DEX, HF, and l-NAME

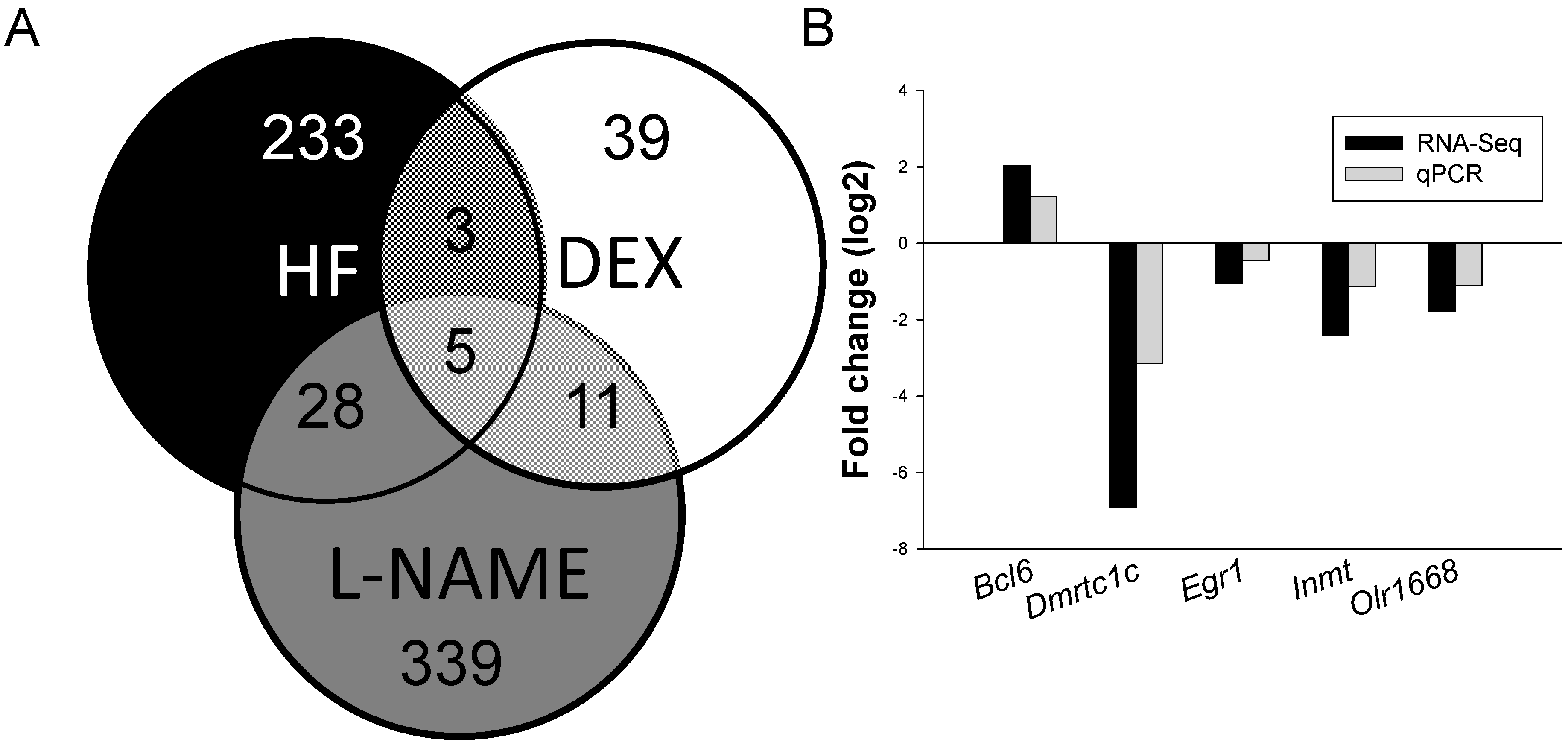
2.2. The Effects of DEX, HF, and l-NAME on Renal Transcriptome


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Symbol | Fold Changes | ||
|---|---|---|---|---|
| 3 groups | HF | DEX | l-NAME | |
| ENSRNOG00000001843 | Bcl6 | 5.401 | 0.217 | 3.277 |
| ENSRNOG00000037923 | Dmrtc1c | 12.62 | 38.46 | 0.018 |
| ENSRNOG00000019422 | Egr1 | 2.885 | 3.947 | 0.480 |
| ENSRNOG00000011250 | Inmt | 0.283 | 0.275 | 0.188 |
| ENSRNOG00000029622 | Olr1668 | 4.562 | 4.106 | 0.060 |
| 2 groups: HF and DEX | HF | DEX | ||
| ENSRNOG00000012531 | Ephb2 | 2.254 | 5.554 | |
| ENSRNOG00000017286 | Ephx2 | 24.34 | 16.76 | |
| ENSRNOG00000029651 | Rdh2 | 0.228 | 0.195 | |
| 2 groups: HF and l-NAME | HF | l-NAME | ||
| ENSRNOG00000005542 | Apob | 0.254 | 2.167 | |
| ENSRNOG00000007830 | Apold1 | 2.658 | 0.369 | |
| ENSRNOG00000003745 | Atf3 | 4.408 | 0.294 | |
| ENSRNOG00000043249 | B0BNK5_RAT | 2.148 | 0.345 | |
| ENSRNOG00000021027 | Dbp | 4.491 | 4.966 | |
| ENSRNOG00000000577 | Ddit4 | 2.319 | 2.003 | |
| ENSRNOG00000016301 | Dmrt2 | 2.361 | 2.061 | |
| ENSRNOG00000003977 | Dusp1 | 2.379 | 0.380 | |
| 2 groups: HF and l-NAME | HF | l-NAME | ||
| ENSRNOG00000029919 | F1M7Y9_RAT | 4.44 × 10−4 | 8.03 × 10−6 | |
| ENSRNOG00000011631 | Fst | 4.452 | 0.403 | |
| ENSRNOG00000013090 | Gadd45g | 2.732 | 0.156 | |
| ENSRNOG00000026053 | GREM1_RAT | 2.798 | 2.093 | |
| ENSRNOG00000003616 | Grem2 | 3.557 | 9.922 | |
| ENSRNOG00000015724 | Gucy2g | 46.59 | 7.268 | |
| ENSRNOG00000029886 | Hba-a2 | 2.009 | 0.131 | |
| ENSRNOG00000033465 | Hbb | 2.608 | 0.049 | |
| ENSRNOG00000019120 | Hmgcs2 | 0.318 | 3.421 | |
| ENSRNOG00000010999 | K1731_RAT | 0.428 | 2.833 | |
| ENSRNOG00000032857 | Klks3 | 0.284 | 0.233 | |
| ENSRNOG00000012871 | LOC100360318 | 26291 | 153519 | |
| ENSRNOG00000030492 | LOC100364577 | 0.199 | 0.135 | |
| ENSRNOG00000013872 | P2ry14 | 3.641 | 2.316 | |
| ENSRNOG00000018187 | Racgap1 | 3.481 | 2.055 | |
| ENSRNOG00000005447 | RGD1311564 | 0.494 | 0.443 | |
| ENSRNOG00000001414 | Serpine1 | 2.878 | 0.421 | |
| ENSRNOG00000019996 | Slc16a1 | 0.430 | 0.458 | |
| ENSRNOG00000006096 | Slc26a7 | 2.423 | 2.269 | |
| ENSRNOG00000015394 | Trpv5 | 0.445 | 2.570 | |
| 2 groups: DEX and l-NAME | DEX | l-NAME | ||
| ENSRNOG00000037206 | Ccdc77 | 4.727 | 0.188 | |
| ENSRNOG00000024899 | Cxcl13 | 0.160 | 0.414 | |
| ENSRNOG00000029128 | Cyp2d5 | 3.162 | 0.284 | |
| ENSRNOG00000021699 | D4A508_RAT | 5.273 | 0.456 | |
| ENSRNOG00000039874 | D4A8E2_RAT | 8.207 | 0.410 | |
| ENSRNOG00000015716 | Gp2 | 6.667 | 2.250 | |
| ENSRNOG00000015518 | Rbp4 | 5.991 | 0.463 | |
| ENSRNOG00000025670 | Shisa3 | 0.101 | 0.441 | |
| ENSRNOG00000030500 | Tcf24 | 0.114 | 1.74 × 10−5 | |
| ENSRNOG00000020057 | Tex101 | 0.061 | 1.71 × 10−5 | |
| ENSRNOG00000000768 | Ubd | 39.33 | 3.45 × 10−6 |
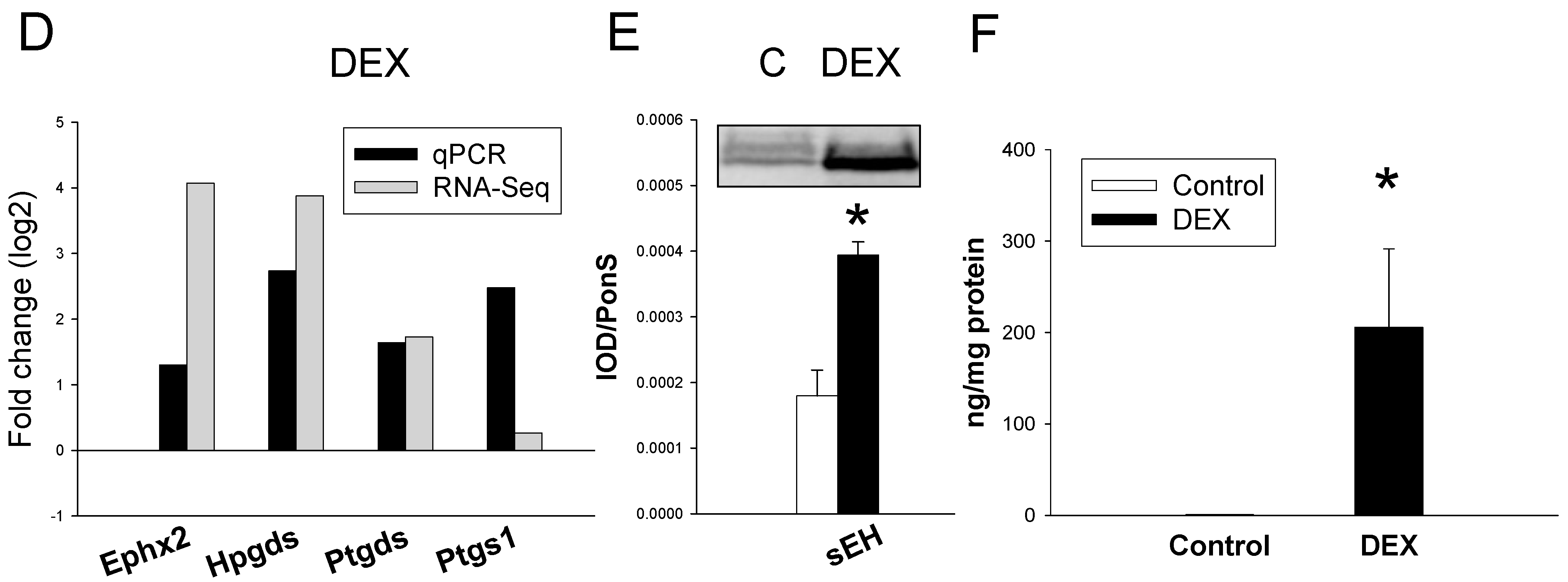
2.3. Gene Expression of Arachidonic Acid Metabolism Pathway


3. Discussion
4. Experimental Section
4.1. Animals
4.2. Next-Generation Sequencing and Analysis
4.3. Quantitative Real-Time PCR Analysis
| Gene | Forward | Reverse |
|---|---|---|
| Bcl6 | 5'-CTGAGGGAAGGCAACATCAT-3' | 5'-CGGCTGTTCAGGAACTCTTC-3' |
| Dmrtc1c | 5'-ACATACAAGTCACGCTGGCA-3' | 5'-TTGGCCTGTTTGAGGGGTTT-3' |
| Egr1 | 5'-CAGGAGTGATGAACGCAAGA-3' | 5'-AGCCCGGAGAGGAGTAAGAG-3' |
| Inmt | 5'-CAGGAGTGATGAACGCAAGA-3' | 5'-AGCCCGGAGAGGAGTAAGAG-3' |
| Olr1668 | 5'-ACGTGGCTATCTGCAGACCT-3' | 5'-CTCCCCACAGGCAGTTTTTA-3' |
| R18s | 5'-GCCGCGGTAATTCCAGCTCCA-3' | 5'-CCCGCCCGCTCCCAAGATC-3' |
4.4. Western Blot
4.5. Statistics
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pereira, M.; Lunet, N.; Azevedo, A.; Barros, H. Differences in prevalence, awareness, treatment and control of hypertension between developing and developed countries. J. Hypertens. 2009, 27, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Feig, D.I.; Nakagawa, T.; Sanchez-Lozada, L.G.; Rodriguez-Iturbe, B. Pathogenesis of essential hypertension: Historical paradigms and modern insights. J. Hypertens. 2008, 26, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, C.S. Oxidative stress and nitric oxide deficiency in the kidney: A critical link to hypertension? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R913–R935. [Google Scholar] [CrossRef] [PubMed]
- Paixão, A.D.; Alexander, B.T. How the kidney is impacted by the perinatal maternal environment to develop hypertension. Biol. Reprod. 2013, 89, 1–10. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsieh, C.S.; Lin, I.C.; Chen, C.C.; Sheen, J.M.; Huang, L.T. Effects of maternal l-citrulline supplementation on renal function and blood pressure in offspring exposed to maternal caloric restriction: The impact of nitric oxide pathway. Nitric Oxide 2010, 23, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N.; Lee, C.T. Melatonin therapy prevents programmed hypertension and nitric oxide deficiency in offspring exposed to maternal caloric restriction. Oxid. Med. Cell Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Lee, W.C.; Hsu, C.N.; Lee, W.C.; Huang, L.T.; Lee, C.T.; Lin, C.Y. Asymmetric dimethylarginine is associated with developmental programming of adult kidney disease and hypertension in offspring of streptozotocin-treated mothers. PLoS One 2013, 8, e55420. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Leu, S.; Wu, K.L.; Lee, W.C.; Chan, J.Y. Melatonin prevents maternal fructose intake-induced programmed hypertension in the offspring: Roles of nitric oxide and arachidonic acid metabolites. J. Pineal Res. 2014, 57, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Huang, L.T.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal citrulline supplementation prevents prenatal NG-nitro-l-arginine-methyl ester (l-NAME)-induced programmed hypertension in rats. Biol. Reprod. 2015, 92, 7. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Chen, C.C.; Sheen, J.M.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Melatonin attenuates prenatal dexamethasone-induced blood pressure increase in a rat model. J. Am. Soc. Hypertens. 2014, 8, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Sheen, J.M.; Chen, C.C.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Maternal citrulline supplementation prevents prenatal dexamethasone-induced programmed hypertension. Free Radic. Res. 2014, 48, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Ritz, E.; Amann, K.; Koleganova, N.; Benz, K. Prenatal programming-effects on blood pressure and renal function. Nat. Rev. Nephrol. 2011, 7, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Lee, C.T.; Huang, L.T. Long-term effects of maternal citrulline supplementation on renal transcriptome prevention of nitric oxide depletion-related programmed hypertension: The impact of gene-nutrient interactions. Int. J. Mol. Sci. 2014, 15, 23255–23268. [Google Scholar] [CrossRef] [PubMed]
- NIH DAVID Bioinformatics Resources 6.7. Available online: http://david.abcc.ncifcrf.gov/ (accessed on 28 December 2014).
- Chawengsub, Y.; Gauthier, K.M.; Campbell, W.B. Role of arachidonic acid lipoxygenase metabolites in the regulation of vascular tone. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H495–H507. [Google Scholar] [CrossRef] [PubMed]
- McMullen, S.; Langley-Evans, S.C.; Gambling, L.; Lang, C.; Swali, A.; McArdle, H.J. A common cause for a common phenotype: The gatekeeper hypothesis in fetal programming. Med. Hypotheses 2012, 78, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Campbell, W.B.; Falck, J.R. Arachidonic acid metabolites as endothelium-derived hyperpolarizing factors. Hypertension 2007, 49, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Sporková, A.; Kopkan, L.; Varcabová, S.; Husková, Z.; Hwang, S.H.; Hammock, B.D.; Imig, J.D.; Kramer, H.J.; Cervenka, L. Role of cytochrome P-450 metabolites in the regulation of renal function and blood pressure in 2-kidney 1-clip hypertensive rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1468–R1475. [Google Scholar] [CrossRef] [PubMed]
- Hatano, R.; Mukouchi, H.; Matsumoto, Y.; Kawaguchi, K.; Kazama, I.; Endo, Y.; Toyama, H.; Ejima, Y.; Kurosawa, S.; Kanai, Y.; et al. Glucocorticoid mediates the transcription of OAT-PG, a kidney-specific prostaglandin transporter. Pflugers. Arch. 2014, 466, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.S.; Park, B.Y.; Saint-Jeannet, J.P. The function of Dmrt genes in vertebrate development: It is not just about sex. Dev. Biol. 2007, 310, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pagel, J.I.; Deindl, E. Disease progression mediated by egr-1 associated signaling in response to oxidative stress. Int. J. Mol. Sci. 2012, 13, 13104–13117. [Google Scholar] [CrossRef] [PubMed]
- Kärkkäinen, J.; Forsström, T.; Tornaeus, J.; Wähälä, K.; Kiuru, P.; Honkanen, A.; Stenman, U.H.; Turpeinen, U.; Hesso, A. Potentially hallucinogenic 5-hydroxytryptamine receptor ligands bufotenine and dimethyltryptamine in blood and tissues. Scand. J. Clin. Lab. Investig. 2005, 65, 189–199. [Google Scholar] [CrossRef]
- Pluznick, J.L.; Zou, D.J.; Zhang, X.; Yan, Q.; Rodriguez-Gil, D.J.; Eisner, C.; Wells, E.; Greer, C.A.; Wang, T.; Firestein, S.; et al. Functional expression of the olfactory signaling system in the kidney. Proc. Natl. Acad. Sci. USA 2009, 106, 2059–2064. [Google Scholar] [CrossRef] [PubMed]
- Koeners, M.P.; Wesseling, S.; Ulu, A.; Sepúlveda, R.L.; Morisseau, C.; Braam, B.; Hammock, B.D.; Joles, J.A. Soluble epoxide hydrolase in the generation and maintenance of high blood pressure in spontaneously hypertensive rats. Am. J. Physiol. Endocrinol. MeTable 2011, 300, E691–E698. [Google Scholar] [CrossRef]
- Procino, G.; Romano, F.; Torielli, L.; Ferrari, P.; Bianchi, G.; Svelto, M.; Valenti, G. Altered expression of renal aquaporins and α-adducin polymorphisms may contribute to the establishment of salt-sensitive hypertension. Am. J. Hypertens. 2011, 24, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.L.; Bowie, L.J.; Jiang, H. Blood pressure changes after intravenous administration of cell-free hemoglobin A and hemoglobin H in the rat. Nitric Oxide 2000, 4, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Kalea, A.Z.; Batlle, D. Apelin and ACE2 in cardiovascular disease. Curr. Opin. Investig. Drugs 2010, 11, 273–282. [Google Scholar] [PubMed]
- Thoonen, R.; Sips, P.Y.; Bloch, K.D.; Buys, E.S. Pathophysiology of hypertension in the absence of nitric oxide/cyclic GMP signaling. Curr. Hypertens. Rep. 2013, 15, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Abraham, N.G.; Kappas, A. Heme oxygenase and the cardiovascular-renal system. Free Radic. Biol. Med. 2005, 39, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Shanks, J.; Herring, N. Peripheral cardiac sympathetic hyperactivity in cardiovascular disease: Role of neuropeptides. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1411–R1420. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Campain, A.E.; Tomaszewski, M.; Zukowska-Szczechowska, E.; Yang, Y.H.; Charchar, F.J.; Morris, B.J. Gene expression profiling reveals renin mRNA overexpression in human hypertensive kidneys and a role for microRNAs. Hypertension 2011, 58, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Witlin, A.G.; Gangula, P.R.; Thompson, M.L.; Yallampalli, C. Growth and fertility rates in the offspring of pregnant rats treated with l-ω nitro-l-arginine methyl ester (l-NAME), a nitric oxide inhibitor. Am. J. Obstet. Gynecol. 2002, 186, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Vaiman, D.; Gascoin-Lachambre, G.; Boubred, F.; Mondon, F.; Feuerstein, J.M.; Ligi, I.; Grandvuillemin, I.; Barbaux, S.; Ghigo, E.; Achard, V.; et al. The intensity of IUGR-induced transcriptome deregulations is inversely correlated with the onset of organ function in a rat model. PLoS One 2011, 6, e21222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almon, R.R.; Lai, W.; DuBois, D.C.; Jusko, W.J. Corticosteroid-regulated genes in rat kidney: Mining time series array data. Am. J. Physiol. Endocrinol. MeTable 2005, 289, E870–E882. [Google Scholar] [CrossRef]
- Smeds, L.; Künstner, A. ConDeTri—A content dependent read trimmer for Illumina data. PLoS One 2011, 6, e26314. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tain, Y.-L.; Huang, L.-T.; Chan, J.Y.H.; Lee, C.-T. Transcriptome Analysis in Rat Kidneys: Importance of Genes Involved in Programmed Hypertension. Int. J. Mol. Sci. 2015, 16, 4744-4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16034744
Tain Y-L, Huang L-T, Chan JYH, Lee C-T. Transcriptome Analysis in Rat Kidneys: Importance of Genes Involved in Programmed Hypertension. International Journal of Molecular Sciences. 2015; 16(3):4744-4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16034744
Chicago/Turabian StyleTain, You-Lin, Li-Tung Huang, Julie Y. H. Chan, and Chien-Te Lee. 2015. "Transcriptome Analysis in Rat Kidneys: Importance of Genes Involved in Programmed Hypertension" International Journal of Molecular Sciences 16, no. 3: 4744-4758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms16034744